Ectopic Expression of a Thellungiella salsuginea Aquaporin Gene, TsPIP1;1, Increased the Salt Tolerance of Rice

Abstract
:1. Introduction
2. Results
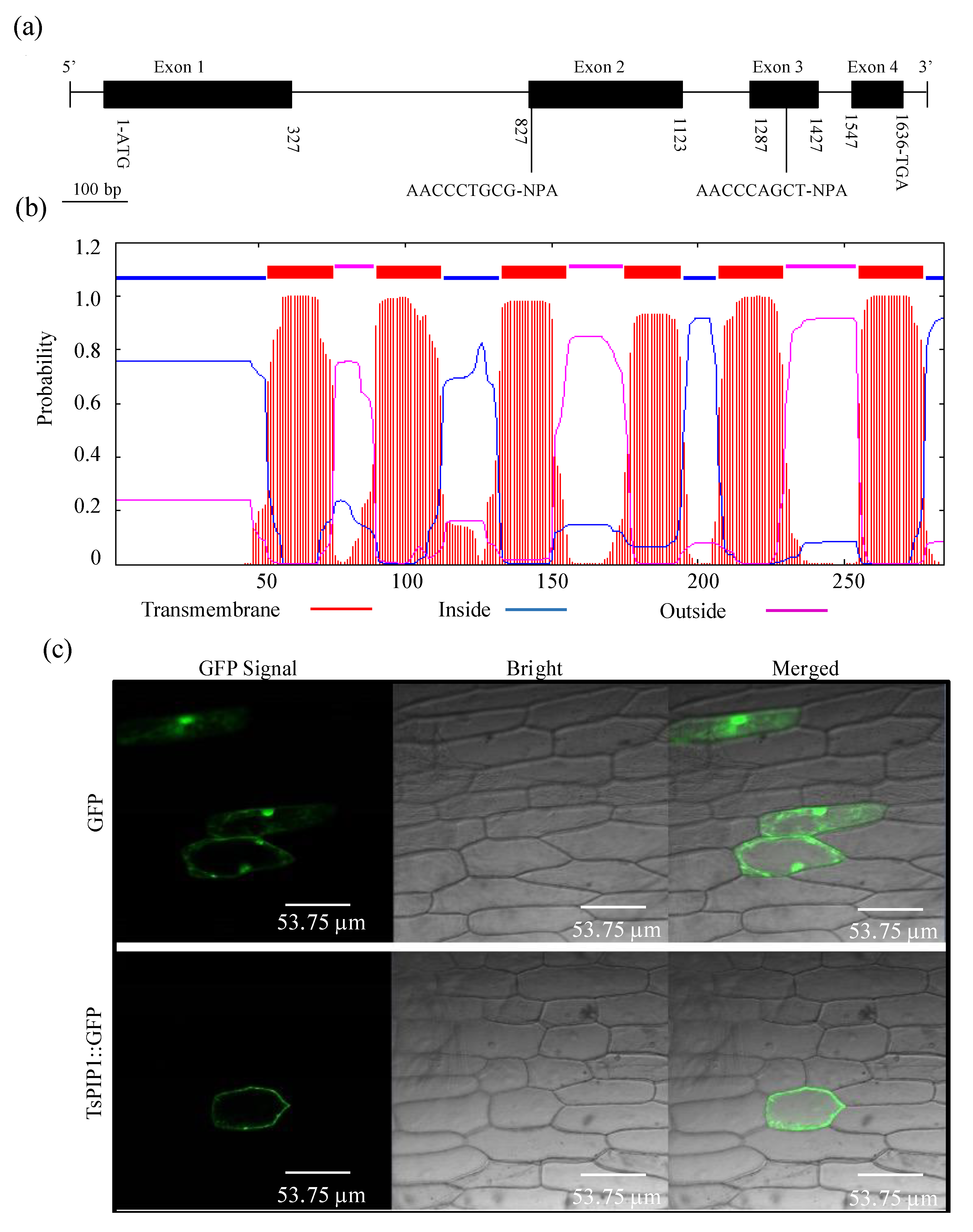
2.1. Analyses of the TsPIP1;1 Gene’s Structural Characteristics
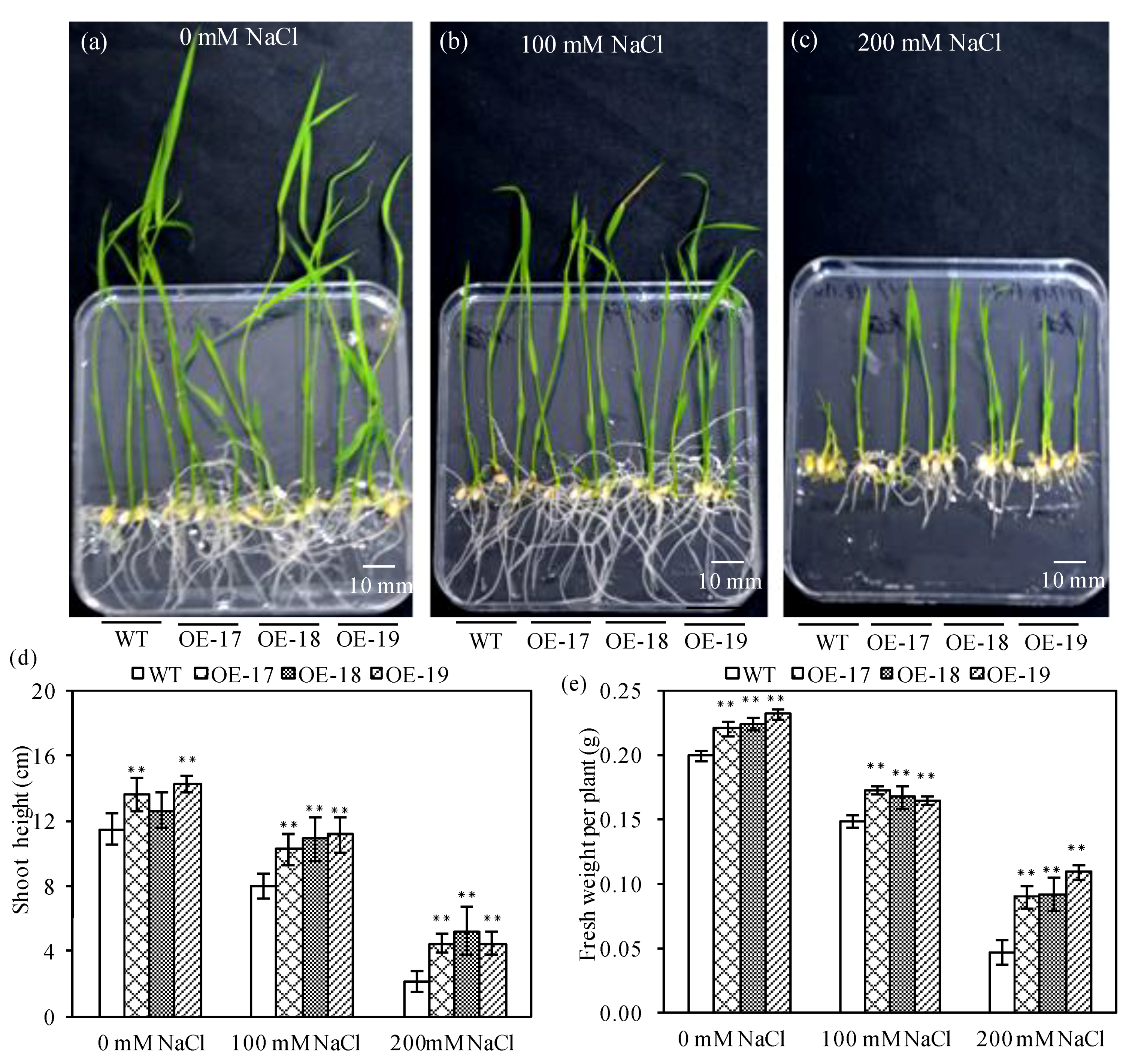
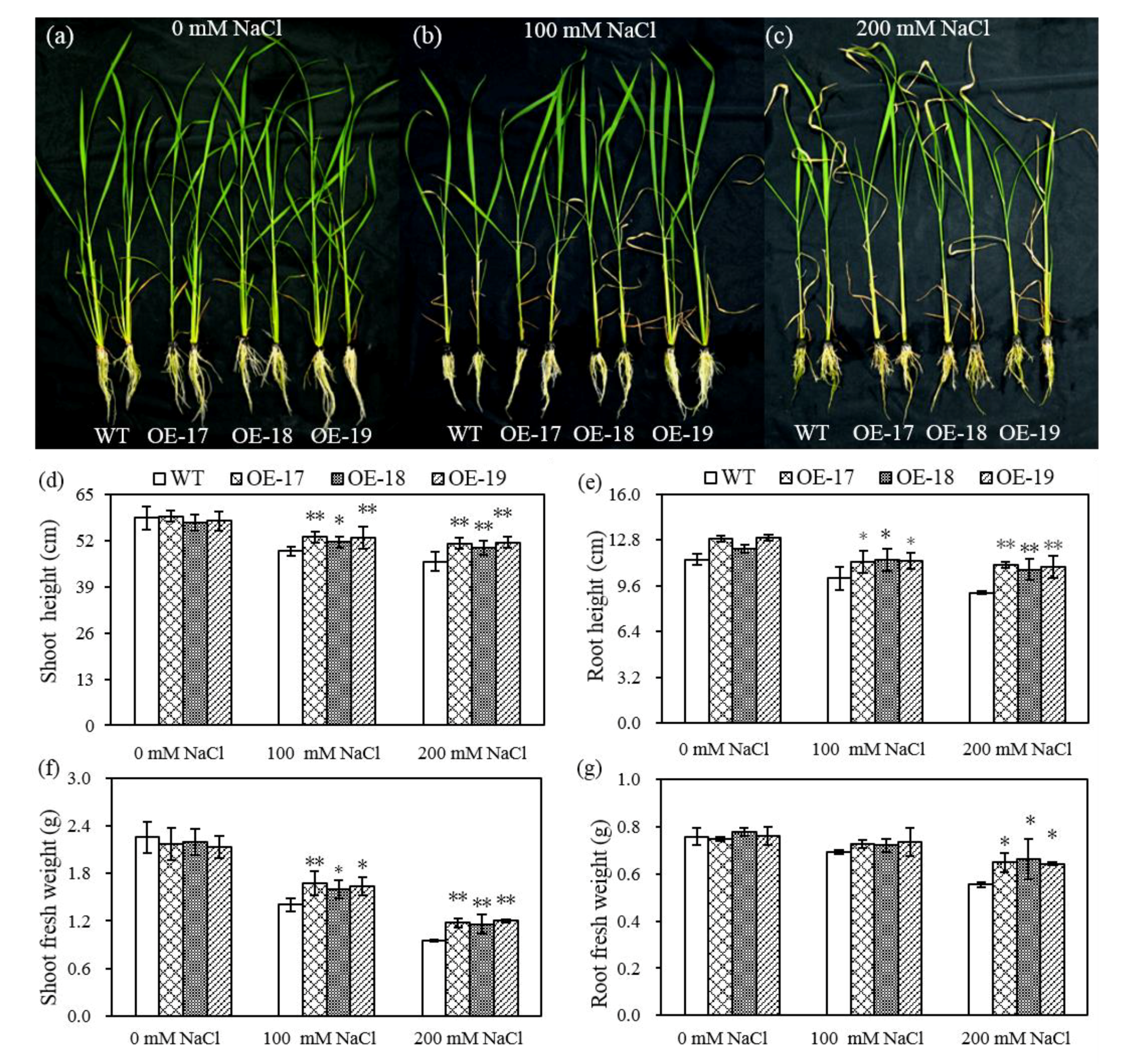
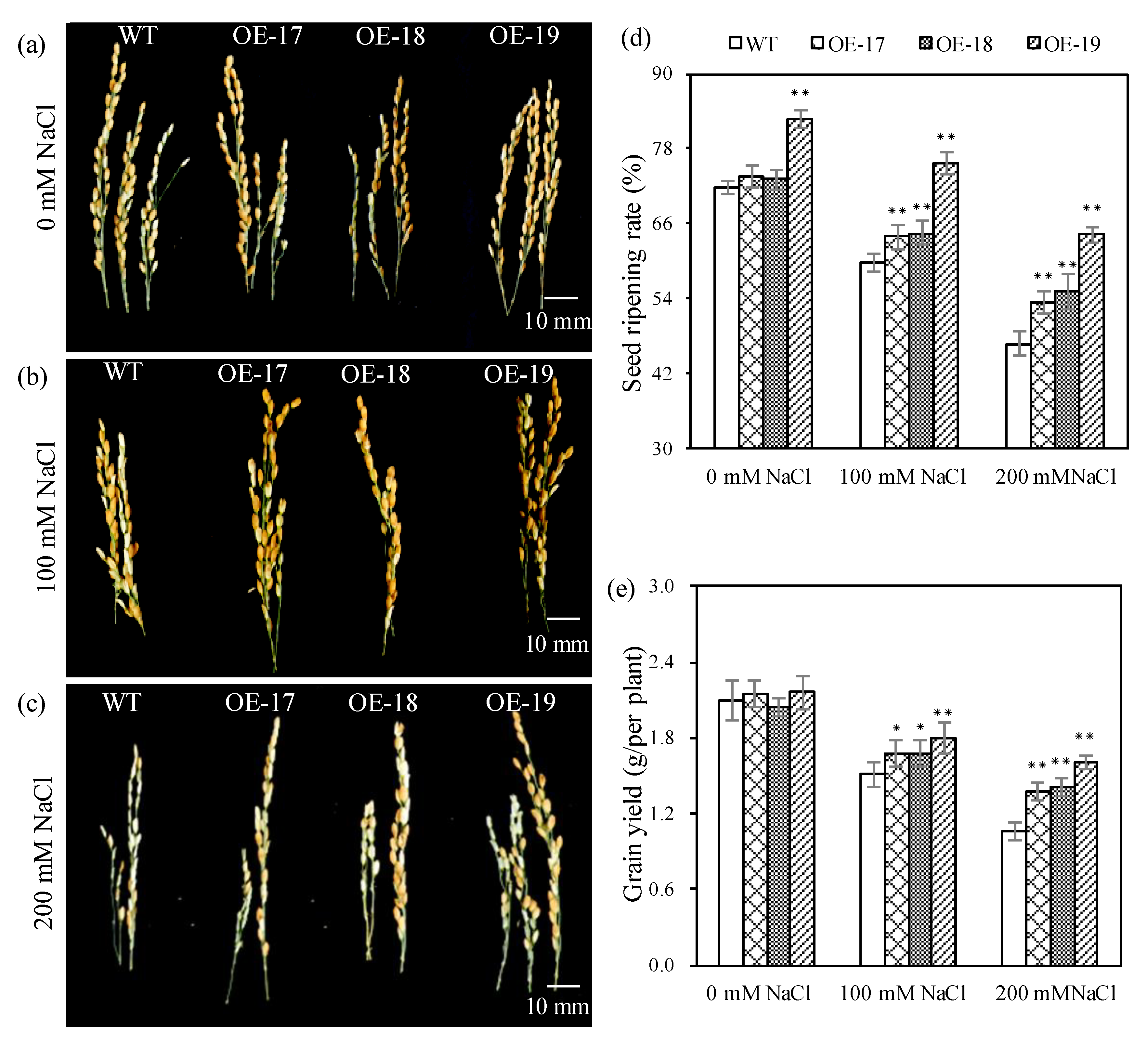
2.2. The Phenotypic Characteristics of Transgenic Rice under Salt Stress
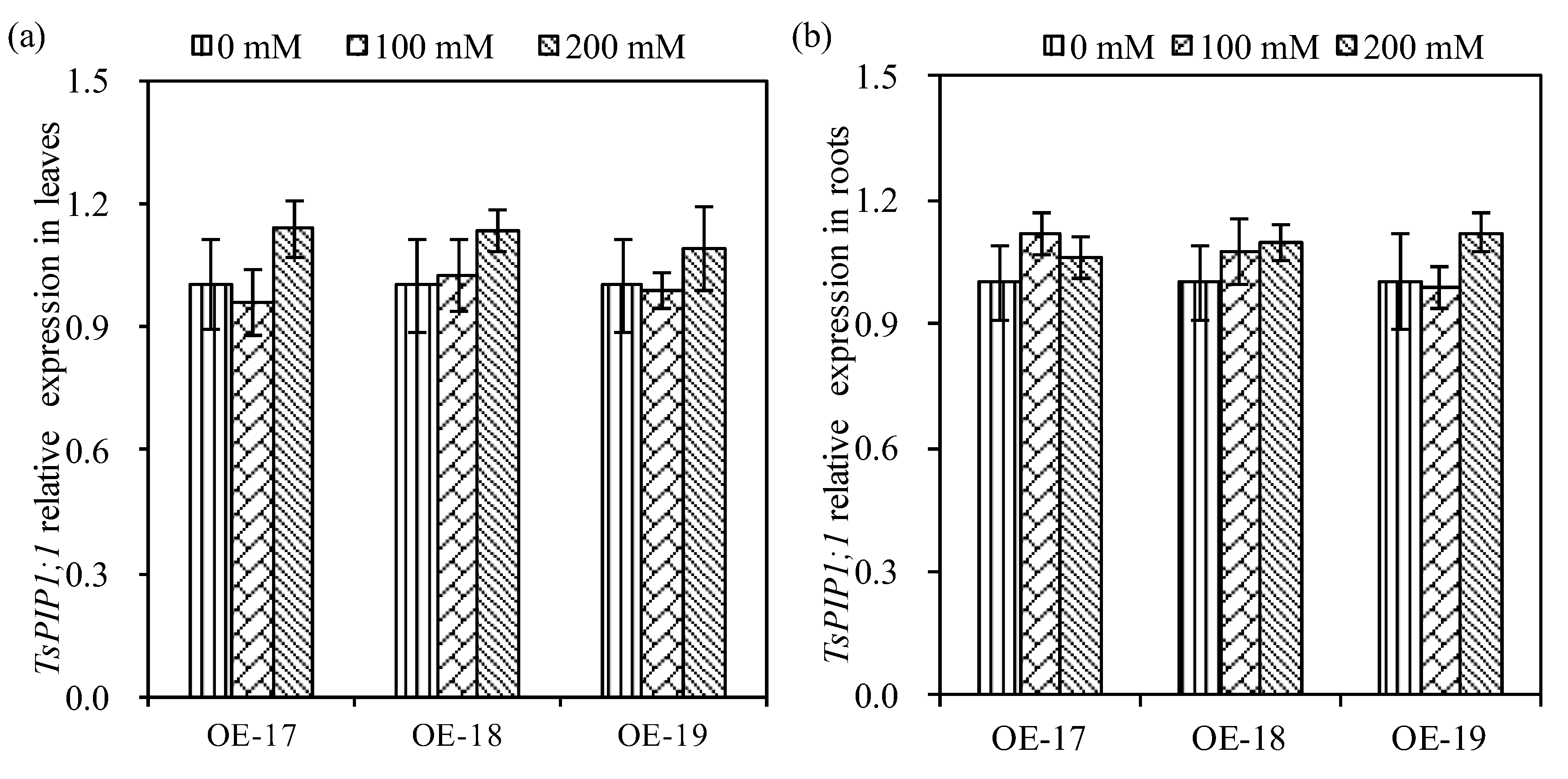
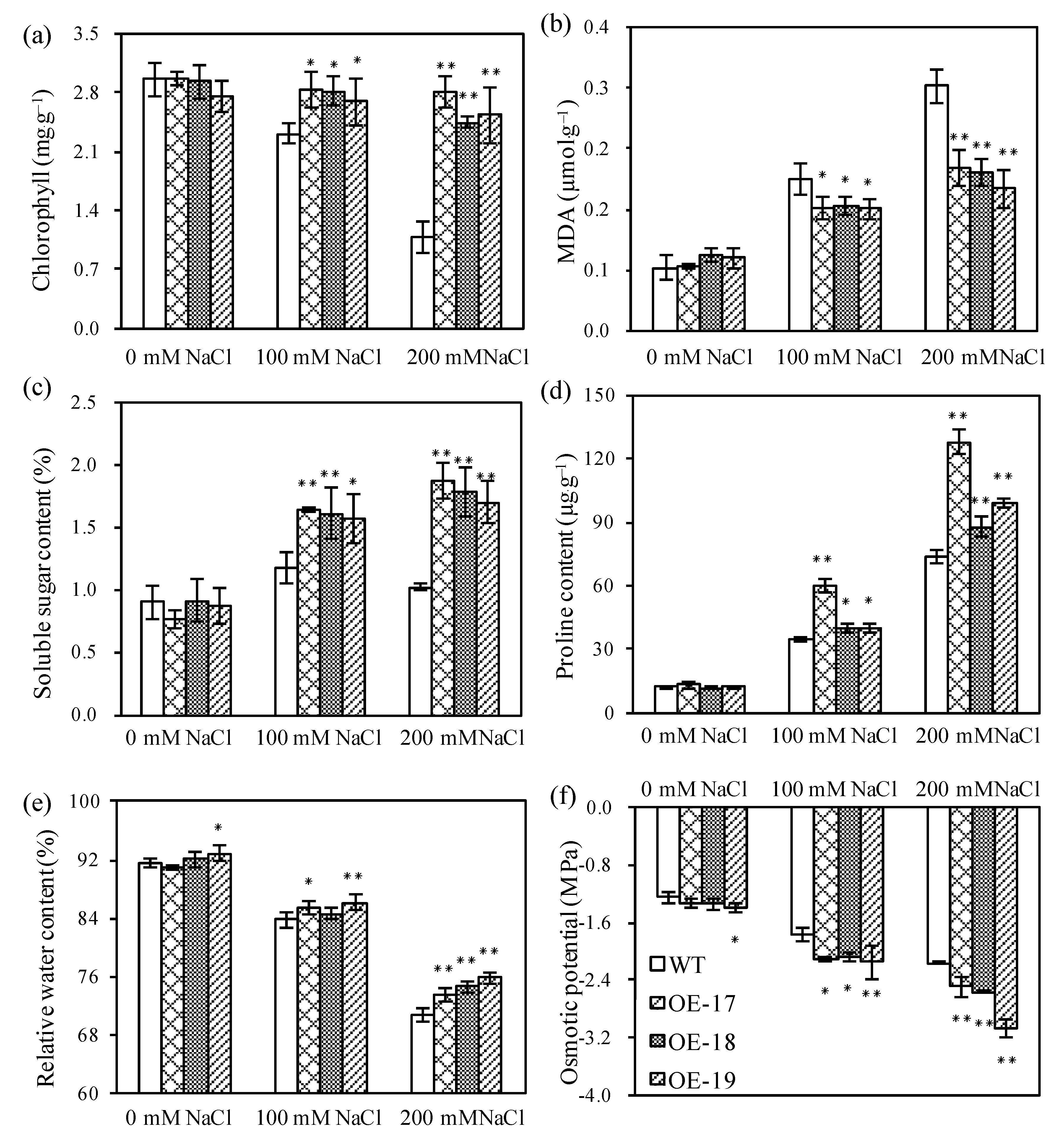
2.3. Relative Expressions of the TsPIP1;1 Gene and Physiological Changes
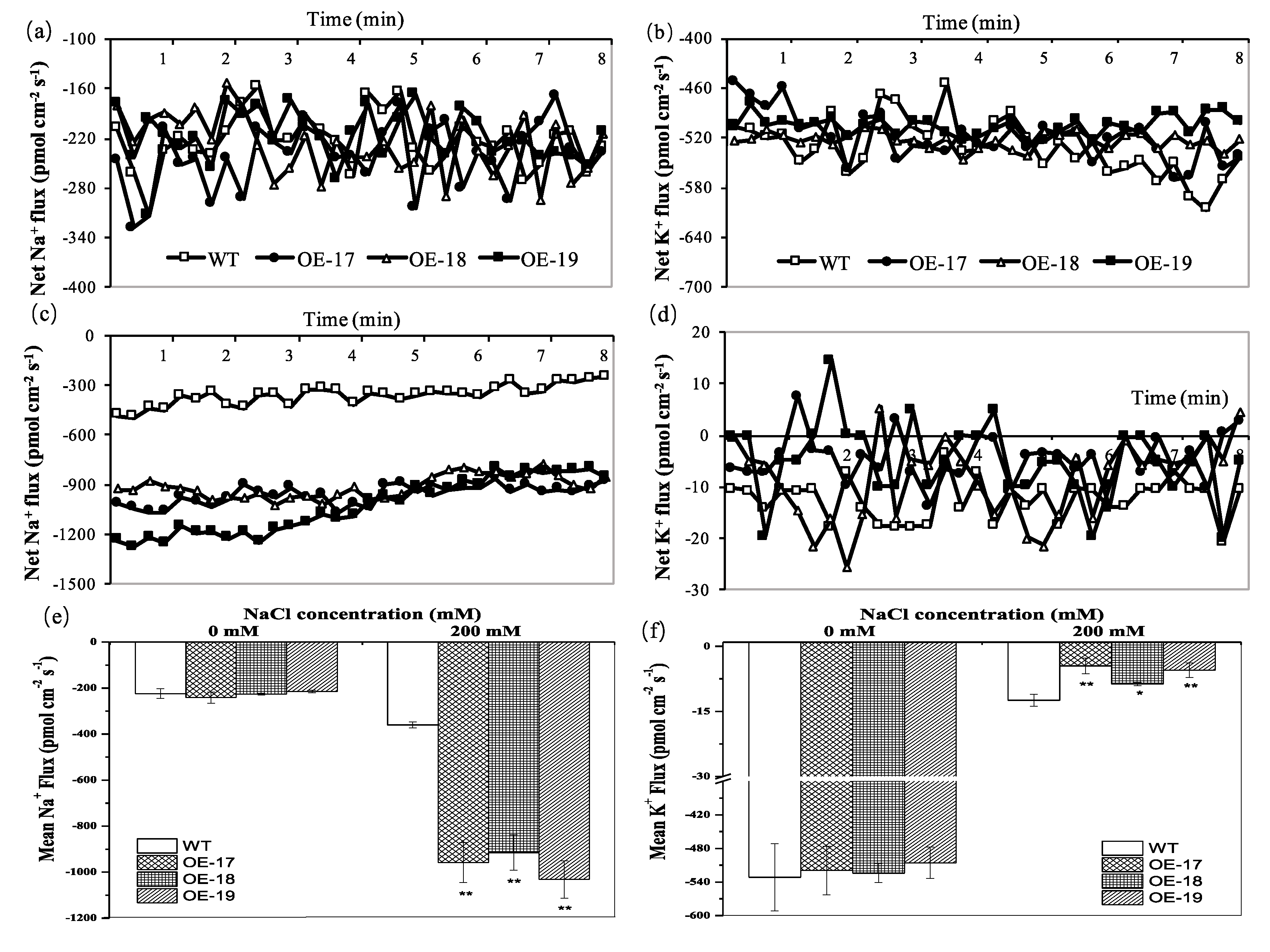
2.4. The Accumulation Profiles of K+ and Na+ in Transgenic Rice
2.5. The Flux Analyses of Na and K in the Roots of Transgenic Rice
2.6. Transgenic Rice Overexpressing the TsPIP1;1 Gene Showed Enhanced Photosynthesis
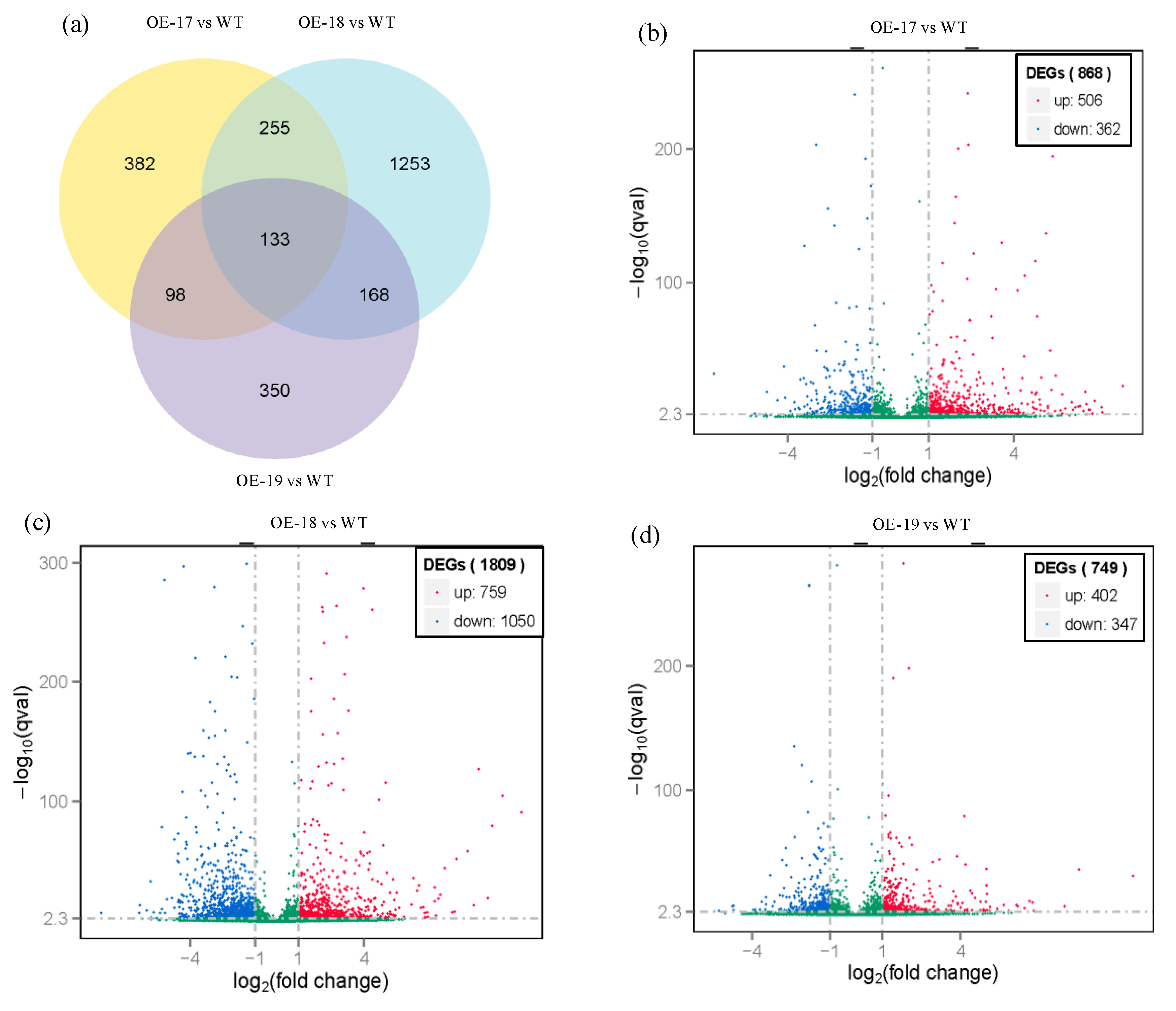
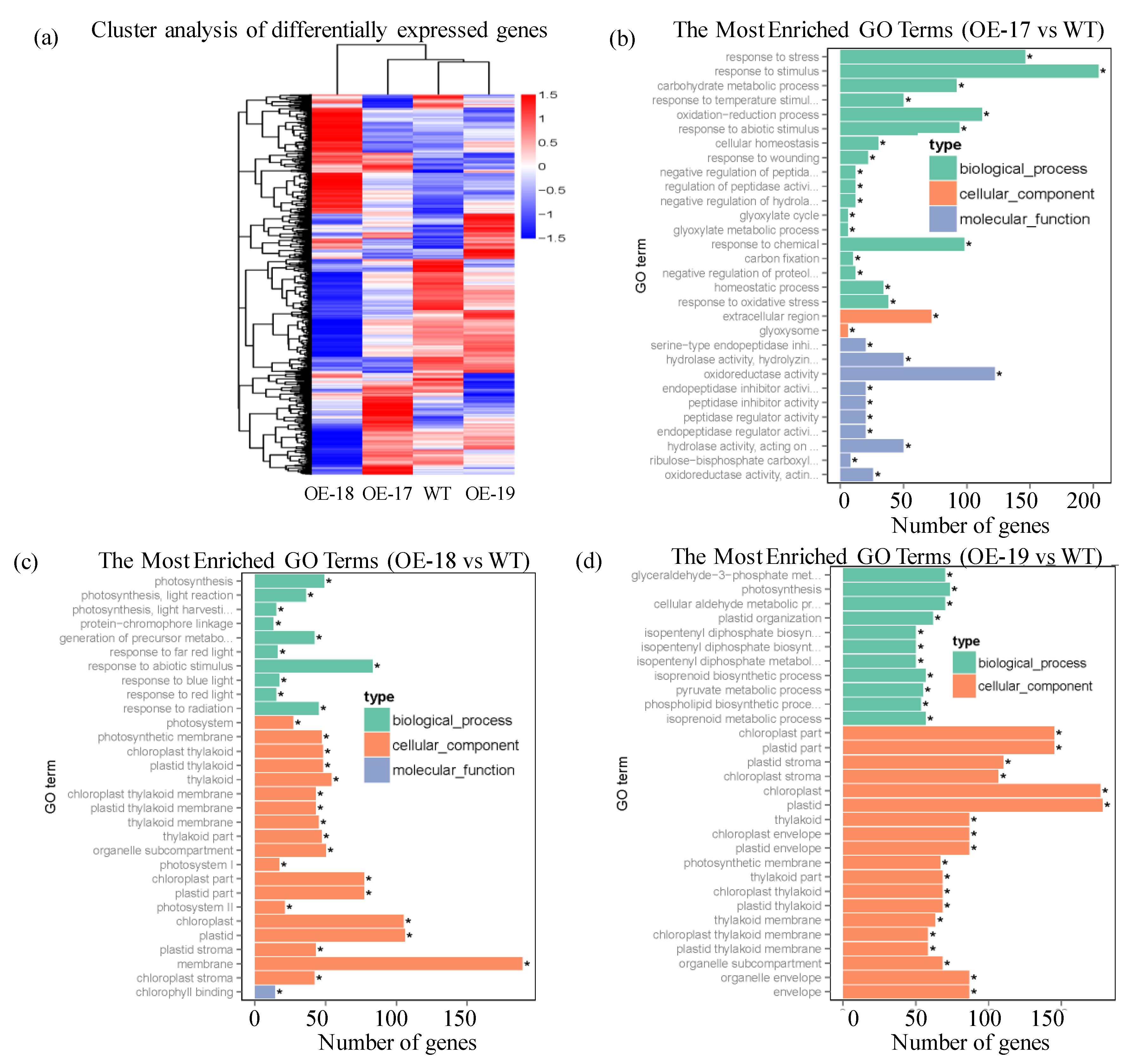
2.7. Responses of Differentially Expressed Genes in the Transgenic Lines to Salt Stress
3. Discussion
4. Materials and Methods
4.1. Plant Culture and Treatments
4.2. Plant Transformation and Subcellular Localization
4.3. RNA Extraction and qPCR Analyses
4.4. Morphological Parameters and Physiological Measurements
4.5. Measurement of Na and K Ion Flux
4.6. Analyses of Transcriptome Sequencing
4.7. Bioinformatics Analysis and Statistical Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| PIP | Plasma membrane intrinsic proteins |
| TIP | Tonoplast intrinsic proteins |
| NIP | Nod26-like intrinsic proteins |
| SIP | Small and basic intrinsic proteins |
| XIP | Uncharacterized intrinsic proteins |
| DEG | Differentially expressed genes |
| MDA | Malondialdehyde |
| NMT | Non-invasive micro-test technique |
| MS | Murashige and Skoog medium |
| ERF109 | Ethylene response factor109 |
| bHLH35 | Basic helix-loop-helix 35 |
| GLK1 | Golden-like 1 transcription factor |
| CaATP2 | Plant-type II Ca2+ATPases |
| RBCS | Ribulose bisphosphate carboxylase small chain |
| LHCA4 | Light-harvesting complex I chlorophyll a/b binding protein 4 |
References
- Parida, A.K.; Das, A.B.; Mittra, B. Effects of salt on growth ion accumulation photosynthesis and leaf anatomy of the mangrove Bruguiera parviflora. Trees 2004, 18, 167–174. [Google Scholar] [CrossRef]
- Niu, X.; Bressan, R.A.; Hasegawa, P.M.; Pardo, J.M. Ion homeostasis in NaCl stress environments. Plant Physiol. 1995, 109, 735–742. [Google Scholar] [CrossRef] [PubMed]
- Maurel, C.; Verdoucq, L.; Luu, D.T.; Santoni, V. Plant aquaporins: Membrane channels with multiple integrated functions. Annu. Rev. Plant Biol. 2008, 59, 595–624. [Google Scholar] [CrossRef] [PubMed]
- Törnroth, H.S.; Wang, Y.; Hedfalk, K.; Johanson, U.; Karlsson, M.; Tajkhorshid, E.; Neutze, R.; Kjellbom, P. Structural mechanism of plant aquaporin gating. Nature 2006, 439, 688–694. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Zhou, M.; Dong, X.; Zou, S.; Xiao, H.; Ma, X. Molecular mechanisms of foliar water uptake in a desert tree. AoB Plants 2015, 7, plv129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumoto, T.; Lian, H.L.; Su, W.A.; Tanaka, D.; Liu, C.; Iwasaki, I.; Kitagawa, Y. Role of the aquaporin PIP1 subfamily in the chilling tolerance of rice. Plant Cell Physiol. 2009, 50, 216–229. [Google Scholar] [CrossRef] [PubMed]
- Uehlein, N.; Lovisolo, C.; Siefritz, F.; Kaldenhoff, R. The tobacco aquaporin NtAQP1 is a membrane CO2 pore with physiological functions. Nature 2003, 425, 734–737. [Google Scholar] [CrossRef] [PubMed]
- Prak, S.; Hem, S.; Boudet, J.; Viennois, G.; Sommerer, N.; Rossignol, M.; Maurel, C.; Santoni, V. Multiple phosphorylations in the C-terminal tail of plant plasma membrane aquaporins: Role in subcellular trafficking of AtPIP2;1 in response to salt stress. Mol. Cell. Proteom. 2008, 7, 1019–1030. [Google Scholar] [CrossRef] [PubMed]
- Dean, R.M.; Rivers, R.L.; Zeidel, M.L.; Roberts, D.M. Purification and functional reconstitution of soybean nodulin 26. An aquaporin with water and glycerol transport properties. Biochemistry 1999, 38, 347–353. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Chen, A.; Zhong, N.; Liu, N.; Wu, X.; Wang, F.; Yang, C.L.; Romero, M.F.; Xia, G.X. The Thellungiella salsuginea tonoplast aquaporin TsTIP1;2 functions in protection against multiple abiotic stresses. Plant Cell Physiol. 2014, 55, 148–161. [Google Scholar] [CrossRef] [PubMed]
- Amtmann, A. Learning from evolution: Thellungiella generates new knowledge on essential and critical components of abiotic stress tolerance in plants. Mol. Plant 2009, 2, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Yue, C.; Cao, H.; Wang, L.; Zhou, Y.; Hao, X.; Zeng, J.; Wang, X.; Yang, Y. Molecular cloning and expression analysis of tea plant aquaporin (AQP) gene family. Plant Physiol. Biochem. 2014, 83, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Maurel, C.; Verdoucq, L.; Rodrigues, O. Aquaporins and plant transpiration. Plant Cell Environ. 2016, 39, 2580–2587. [Google Scholar] [CrossRef] [PubMed]
- Vera, E.R.; Barkla, B.J.; Bohnert, H.J.; Pantoja, O. Novel regulation of aquaporins during osmotic stress. Plant Physiol. 2004, 135, 2318–2329. [Google Scholar] [CrossRef] [PubMed]
- Postaire, O.; Tournaire, R.C.; Grondin, A.; Boursiac, Y.; Morillon, R.; Schäffner, A.R.; Maurel, C. A PIP1 aquaporin contributes to hydrostatic pressure-induced water transport in both the root and rosette of Arabidopsis. Plant Physiol. 2010, 152, 1418–1430. [Google Scholar] [CrossRef] [PubMed]
- Sakurai, J.; Ishikawa, F.; Yamaguchi, T.; Uemura, M.; Maeshima, M. Identification of 33 Rice aquaporin genes and analysis of their expression and function. Plant Cell Physiol. 2005, 46, 1568–1577. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Fukumoto, T.; Matsumoto, T.; Gen, P.; Frascaria, D.; Kaneko, T.; Katsuhara, M.; Zhong, S.; Sun, X.; Zhu, Y.; et al. Aquaporin OsPIP1;1 promotes rice salt resistance and seed germination. Plant Physiol. 2013, 63, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Mahajan, S.; Narendra, T. Cold salinity and drought stresses: An overview. Arch. Biochem. Biophys. 2005, 444, 139–158. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.L.; Yu, G.H.; Han, X.R.; Xin, S.C.; Qiang, X.J.; Jiang, L.L.; Zhang, S.; Cheng, X. TsMIP6 enhances the tolerance of transgenic rice to salt stress and interacts with target proteins. J. Plant Biol. 2015, 58, 285–292. [Google Scholar] [CrossRef]
- Xu, Y.; Hu, W.; Liu, J.; Zhang, J.; Jia, C.; Miao, H.; Xu, B.; Jin, Z. A banana aquaporin gene MaPIP1;1 is involved in tolerance to drought and salt stresses. BMC Plant Biol. 2014, 14, 59–72. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Li, Q.; Lei, Q.; Feng, C.; Gao, Y.; Zheng, X.; Zhao, Y.; Wang, Z.; Kong, J. MzPIP2;1: An aquaporin involved in radial water movement in both water uptake and transportation altered the drought and salt tolerance of transgenic Arabidopsis. PLoS ONE 2015, 10, e0142446. [Google Scholar] [CrossRef] [PubMed]
- Xin, S.C.; Yu, G.H.; Sun, L.L.; Qiang, X.J.; Xu, N.; Cheng, X.G. Expression of tomato SlTIP2;2 enhances the tolerance to salt stress in the transgenic Arabidopsis and interacts with target proteins. J. Plant Res. 2014, 127, 695–708. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yu, H.; Zhang, Y.; Wang, Y.; Li, M.; Zhang, J.; Duan, L.; Zhang, M.; Li, Z. Increased abscisic acid levels in transgenic maize overexpressing AtLOS5 mediated root ion fluxes and leaf water status under salt stress. J. Exp. Bot. 2016, 67, 1339–1355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerganova, M.; Popova, A.V.; Stanoeva, D.; Velitchkova, M. Tomato plants acclimate better to elevated temperature and high light than to treatment with each factor separately. Plant Physiol. Biochem. 2016, 25, 234–241. [Google Scholar] [CrossRef] [PubMed]
- Shareena, S.; Upendra, K.; Singh, S.; Thumballi, R.G. Constitutive and stress-inducible overexpression of a native aquaporin gene (MusaPIP2;6) in transgenic banana plants signals its pivotal role in salt tolerance. Plant Mol. Biol. 2015, 88, 41–52. [Google Scholar]
- Gao, F.; Zhou, Y.J.; Zhu, W.P.; Li, X.F.; Fan, L.M.; Zhang, G.F. Proteomic analysis of cold stress-responsive proteins in Thellungiella rosette leaves. Planta 2009, 230, 1033–1046. [Google Scholar] [CrossRef] [PubMed]
- Sade, N.; Gallé, A.; Flexas, J.; Lerner, S.; Peleg, G.; Yaaran, A.; Moshelion, M. Differential tissue-specific expression of NtAQP1 in Arabidopsis thaliana reveals a role for this protein in stomatal and mesophyll conductance of CO2 under standard and salt-stress conditions. Planta 2014, 239, 357–366. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Wang, C.; Liu, R.; Han, Q.; Vandeleur, R.K.; Du, J.; Tyerman, S.; Shou, H. Constitutive overexpression of soybean plasma membrane intrinsic protein GmPIP1;6 confers salt tolerance. BMC Plant Biol. 2014, 14, 181–194. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.B.; Foley, R.C.; Onate-Sanchez, L. Transcription factors in plant defense and stress responses. Curr. Opin. Plant Biol. 2002, 5, 430–436. [Google Scholar] [CrossRef]
- Xu, S.M.; Wang, X.C.; Chen, J. Zinc finger protein 1 (ThZF1) from salt cress (Thellungiella halophila) is a Cys-2/His-2-type transcription factor involved in drought and salt stress. Plant Cell Rep. 2007, 26, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.Y.; Guan, Y.C.; Wu, Y.R.; Chen, H.L.; Chen, F.; Chu, C.C. Overexpression of a rice OsDREB1F gene increases salt, drought, and low temperature tolerance in both Arabidopsis and rice. Plant Mol. Biol. 2008, 67, 589–602. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Li, F.; Wang, J.L.; Ma, Y.; Chong, K.; Xu, Y.Y. Basic helix-loop-helix transcription factor from wild rice (OrbHLH2) improves tolerance to salt- and osmotic stress in Arabidopsis. J. Plant Physiol. 2009, 166, 1296–1306. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, Y.; Ebisu, Y.; Kinoshita, T.; Doi, M.; Okuma, E.; Murata, Y.; Shimazaki, K. bHLH transcription factors that facilitate K+ uptake during stomatal opening are repressed by abscisic acid through phosphorylation. Sci. Signal. 2013, 6, ra48. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Wang, N.N.; Gong, S.Y.; Lu, R.; Li, Y.; Li, X.B. Overexpression of a cotton (Gossypium hirsutum) WRKY gene, GhWRKY34, in Arabidopsis enhances salt-tolerance of the transgenic plants. Plant Physiol. Biochem. 2015, 96, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Tian, S.; Liu, C.; Dong, H. Identification of a MYB3R gene involved in drought, salt and cold stress in wheat (Triticumaestivum L.). Gene 2011, 485, 46–152. [Google Scholar] [CrossRef] [PubMed]
- Saijo, Y.; Hata, S.; Kyozuka, J.; Shimamoto, K.; Izui, K. Overexpression of a single Ca2+-dependent protein kinase confers both cold and salt/drought tolerance on rice plants. Plant J. 2000, 23, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Murata, Y.; Yoshihashi, M.; Obi, I.; Kakutani, T. Ca2+ Regulation of Outward Rectifying K+ Channel in the Plasma Membrane of Tobacco Cultured Cells in Suspension: A Role of the K+ Channel in Mitigation of Salt-Stress Effects by External Ca2+. Plant Cell Physiol. 1998, 39, 1039–1044. [Google Scholar] [CrossRef]
- Kazi, Md.; Kamrul, H.; Sandep, Y.; Mst, S.A.B.; Dipesh, K.T.; Narendra, T. Genome-wide analysis of plant-type II Ca2+ATPases gene family from rice and Arabidopsis: Potential role in abiotic stresses. Plant Physiol. Biochem. 2013, 65, 32–47. [Google Scholar]
- Muller, P.; Li, X.P.; Niyogi, K.K. Non-Photochemical Quenching, A Response to Excess Light Energy. Plant Physiol. 2001, 125, 1558–1566. [Google Scholar] [CrossRef] [PubMed]
- Flexas, J.; Bota, J.; Galmes, J.; Medrano, H.; Ribas-Carbó, M. Keeping a positive carbon balance under adverse conditions: Responses of photosynthesis and respiration to water stress. Physiol. Plant 2006, 127, 343–352. [Google Scholar] [CrossRef]
- Galmés, J.; Ribas-Carbó, M.; Medrano, H.; Flexas, J. Rubisco activity in Mediterranean species is regulated by the chloroplastic CO2 concentration under water stress. J. Exp. Bot. 2011, 62, 653–665. [Google Scholar] [CrossRef] [PubMed]
- Juan, A.P.; Sebastià, C.B.; Elizabete, C.S.; Jeroni, G. Rubisco and Rubisco Activase Play an Important Role in the Biochemical Limitations of Photosynthesis in Rice, Wheat, and Maize under High Temperature and Water Deficit. Front. Plant Sci. 2017, 8, 490–504. [Google Scholar]
- Sauer, K.; Scheer, H. Energy transfer calculations for two C-phycocyanins based on refined X-ray crystal structure coordinates of chromophores. In Photosynthetic Light-Harvesting Systems; FAO: Rome, Italy, 1988; pp. 508–511. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Sandra, G.; Habib, K. Co-expression of vacuolar Na+/H+ antiporter and H+-pyrophosphatase with an IRES-mediated dicistronic vector improves salinity tolerance and enhances potassium biofortification of tomato. Phytochemistry 2015, 117, 537–546. [Google Scholar]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NaCl Treatment | Lines | Na+ Content (mg/g, DW) | K+ Content (mg/g, DW) | Na+/K+ Ratio |
|---|---|---|---|---|
| 0 mM | WT | 2.44 ± 0.33 | 16.50 ± 1.02 | 0.15 ± 0.02 |
| OE-17 | 2.53 ± 0.42 | 16.73 ± 0.59 | 0.15 ± 0.02 | |
| OE-18 | 2.58 ± 0.17 | 17.51 ± 0.69 | 0.15 ± 0.01 | |
| OE-19 | 2.53 ± 0.26 | 17.77 ± 0.65 | 0.14 ± 0.02 | |
| 100 mM | WT | 14.42 ± 0.42 | 8.61 ± 0.61 | 1.68 ± 0.08 |
| OE-17 | 14.01 ± 0.66 | 10.03 ± 0.72 | 1.40 ± 0.06 * | |
| OE-18 | 14.08 ± 0.80 | 10.28 ± 1.31 * | 1.38 ± 0.18 * | |
| OE-19 | 13.54 ± 0.95 | 9.92 ± 0.44 | 1.37 ± 0.12 ** | |
| 200 mM | WT | 27.90 ± 0.65 | 5.28 ± 0.72 | 5.35 ± 0.58 |
| OE-17 | 25.21 ± 1.27 ** | 6.70 ± 0.95 * | 3.80 ± 0.35 ** | |
| OE-18 | 25.62 ± 1.39 * | 7.80 ± 0.80 ** | 3.32 ± 0.48 ** | |
| OE-19 | 25.60 ± 0.89 * | 7.88 ± 0.54 ** | 3.26 ± 0.33 ** |
| NaCl | Lines | Pn μmol·m−2·s−1 | Gs mol·m−2·s−1 | Ci μmol/mol | Tr mmol m−2·s−1 | Wue μmol/mmol |
|---|---|---|---|---|---|---|
| 0 mM | WT | 12.91 ± 0.09 | 0.44 ± 0.01 | 165.85 ± 0.57 | 4.22 ± 0.13 | 3.06 ± 0.09 |
| OE-17 | 13.11 ± 0.11 * | 0.43 ± 0.01 | 166.59 ± 2.55 | 4.17 ± 0.08 | 3.14 ± 0.03 * | |
| OE-18 | 13.07 ± 0.31 | 0.45 ± 0.01 | 166.23 ± 1.48 | 4.25 ± 0.10 | 3.07 ± 0.08 | |
| OE-19 | 14.04 ± 0.16 ** | 0.49 ± 0.01 ** | 164.37 ± 2.48 | 4.36 ± 0.11 ** | 3.22 ± 0.08 ** | |
| 100 mM | WT | 6.13 ± 0.49 | 0.30 ± 0.01 | 142.91 ± 0.83 | 3.70 ± 0.07 | 1.66 ± 0.13 |
| OE-17 | 6.88 ± 0.55 ** | 0.32 ± 0.01 ** | 144.92 ± 0.65 ** | 3.83 ± 0.25 | 1.80 ± 0.09 ** | |
| OE-18 | 6.89 ± 0.19 ** | 0.31 ± 0.01 * | 143.49 ± 0.25 * | 3.61 ± 0.18 | 1.91 ± 0.08 ** | |
| OE-19 | 7.43 ± 0.37 ** | 0.35 ± 0.01 ** | 144.86 ± 0.47 ** | 3.92 ± 0.09 ** | 1.90 ± 0.12 ** | |
| 200 mM | WT | 3.45 ± 0.29 | 0.21 ± 0.01 | 117.42 ± 1.19 | 2.64 ± 0.11 | 1.31 ± 0.10 |
| OE-17 | 4.30 ± 0.27 ** | 0.27 ± 0.01 ** | 129.84 ± 0.61 ** | 2.96 ± 0.04 ** | 1.45 ± 0.08 ** | |
| OE-18 | 4.21 ± 0.22 ** | 0.24 ± 0.01 ** | 133.43 ± 1.49 ** | 2.95 ± 0.05 ** | 1.43 ± 0.07 ** | |
| OE-19 | 4.94 ± 0.30 ** | 0.29 ± 0.02 ** | 132.99 ± 0.63 ** | 3.13 ± 0.32 ** | 1.59 ± 0.12 ** |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, W.; Qiang, X.-J.; Han, X.-R.; Jiang, L.-L.; Zhang, S.-H.; Han, J.; He, R.; Cheng, X.-G. Ectopic Expression of a Thellungiella salsuginea Aquaporin Gene, TsPIP1;1, Increased the Salt Tolerance of Rice. Int. J. Mol. Sci. 2018, 19, 2229. https://doi.org/10.3390/ijms19082229
Li W, Qiang X-J, Han X-R, Jiang L-L, Zhang S-H, Han J, He R, Cheng X-G. Ectopic Expression of a Thellungiella salsuginea Aquaporin Gene, TsPIP1;1, Increased the Salt Tolerance of Rice. International Journal of Molecular Sciences. 2018; 19(8):2229. https://doi.org/10.3390/ijms19082229
Chicago/Turabian StyleLi, Wei, Xiao-Jing Qiang, Xiao-Ri Han, Lin-Lin Jiang, Shu-Hui Zhang, Jiao Han, Rui He, and Xian-Guo Cheng. 2018. "Ectopic Expression of a Thellungiella salsuginea Aquaporin Gene, TsPIP1;1, Increased the Salt Tolerance of Rice" International Journal of Molecular Sciences 19, no. 8: 2229. https://doi.org/10.3390/ijms19082229
APA StyleLi, W., Qiang, X. -J., Han, X. -R., Jiang, L. -L., Zhang, S. -H., Han, J., He, R., & Cheng, X. -G. (2018). Ectopic Expression of a Thellungiella salsuginea Aquaporin Gene, TsPIP1;1, Increased the Salt Tolerance of Rice. International Journal of Molecular Sciences, 19(8), 2229. https://doi.org/10.3390/ijms19082229