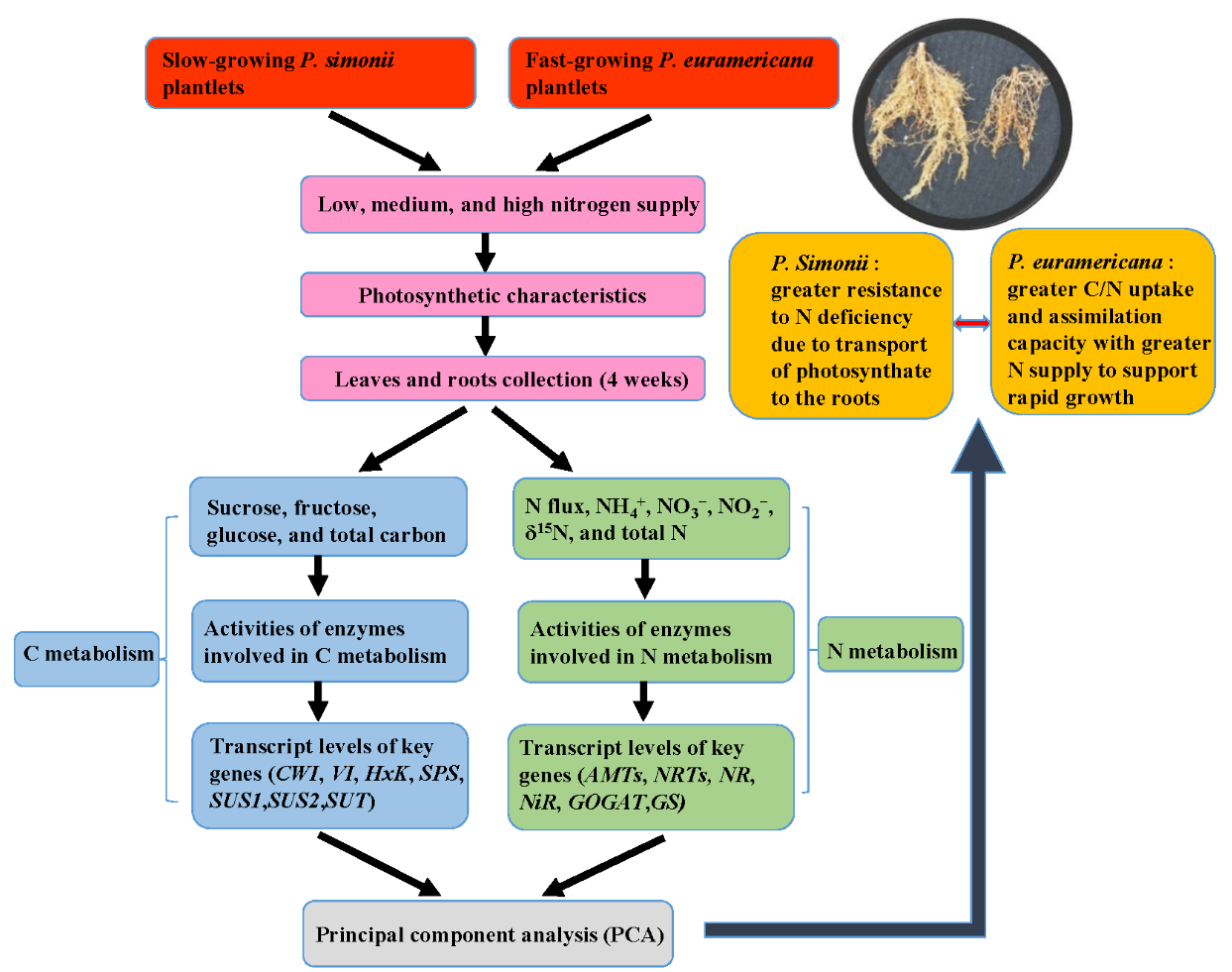
Distinct Carbon and Nitrogen Metabolism of Two Contrasting Poplar Species in Response to Different N Supply Levels

Abstract
:
1. Introduction
2. Results
2.1. Root Growth and Photosynthetic Characteristics
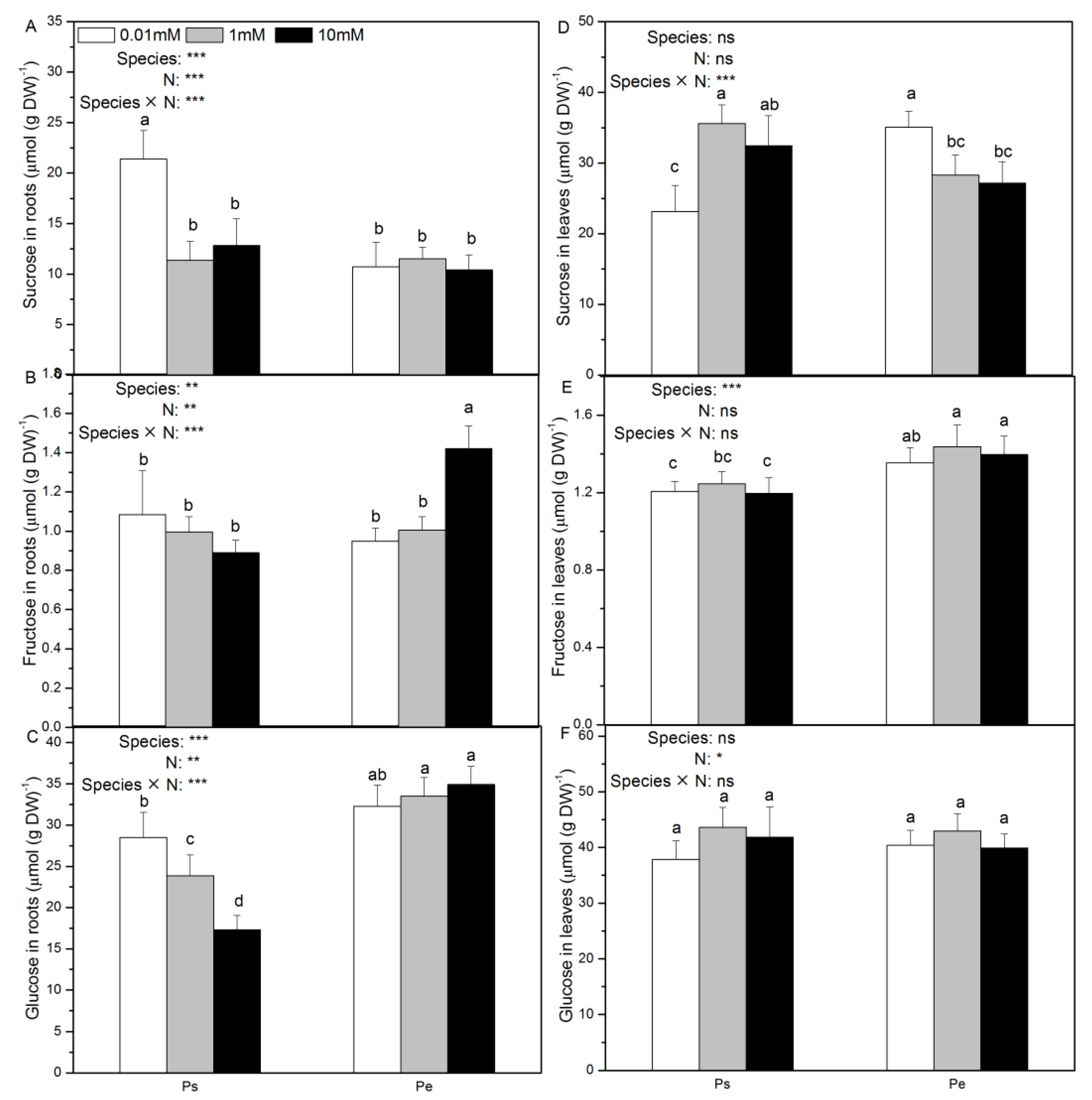
2.2. Contents of Sucrose, Fructose, Glucose, and Total Carbon
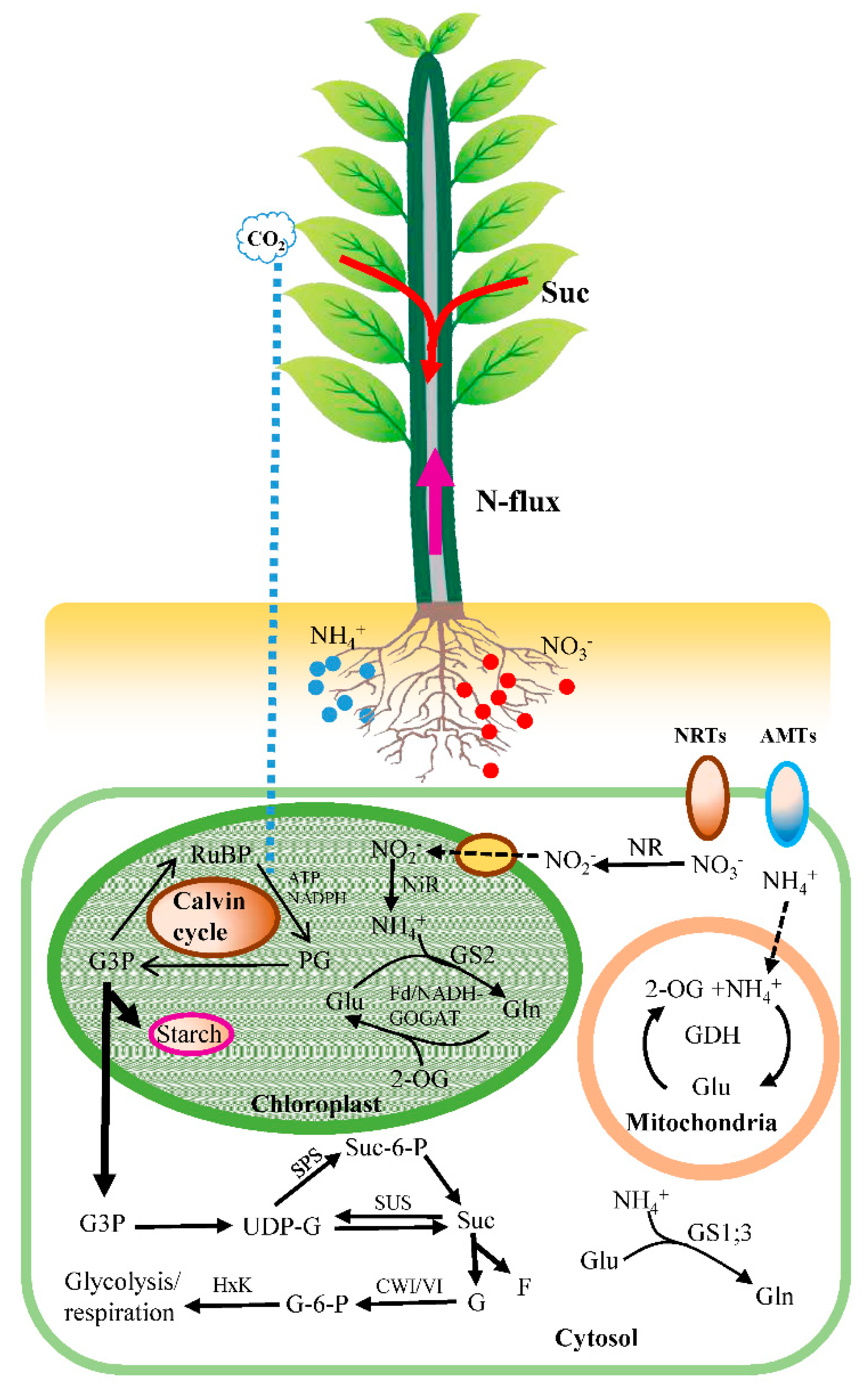
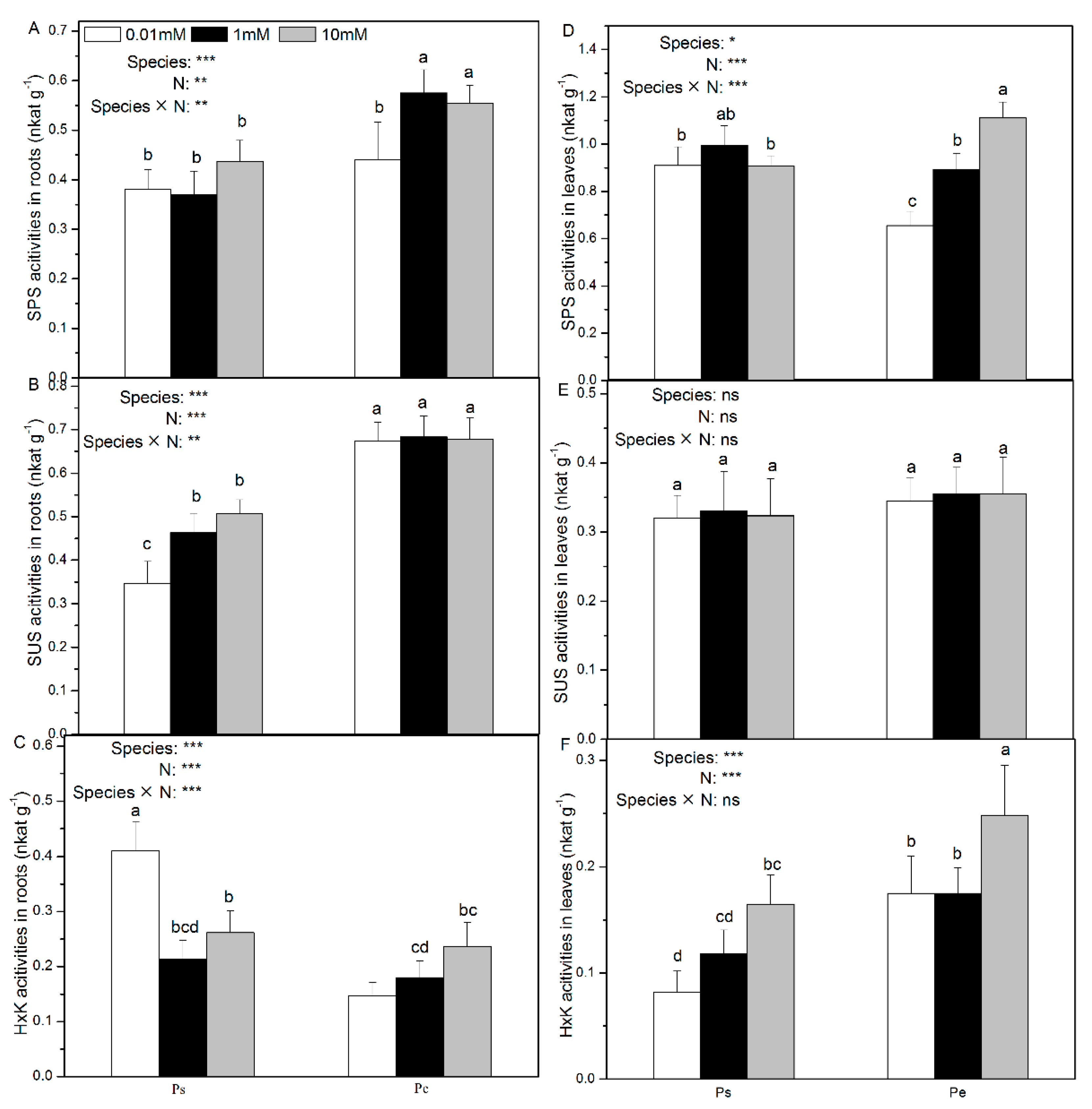
2.3. Root and Leaf Activities of Enzymes Involved in C Assimilation
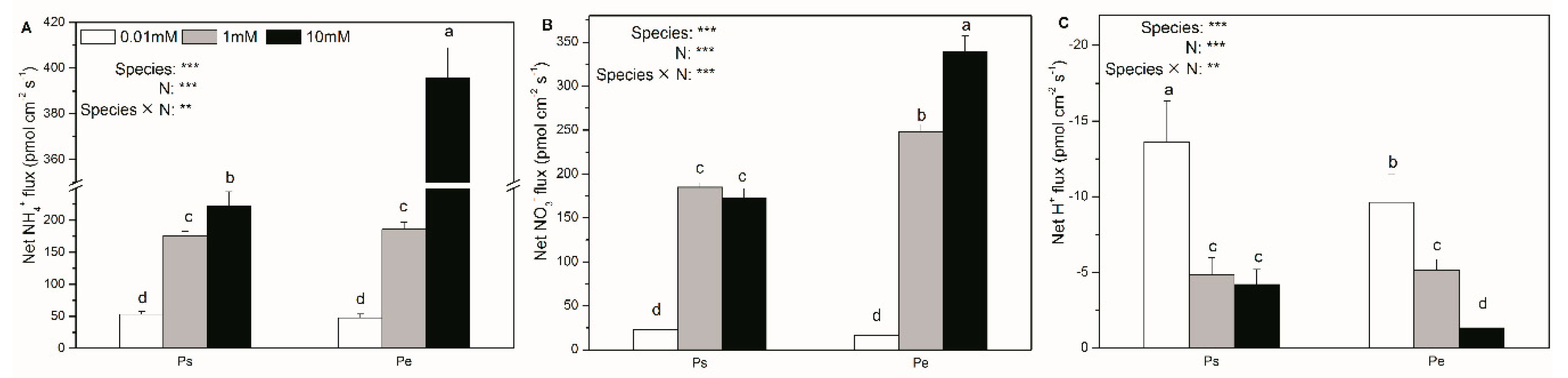
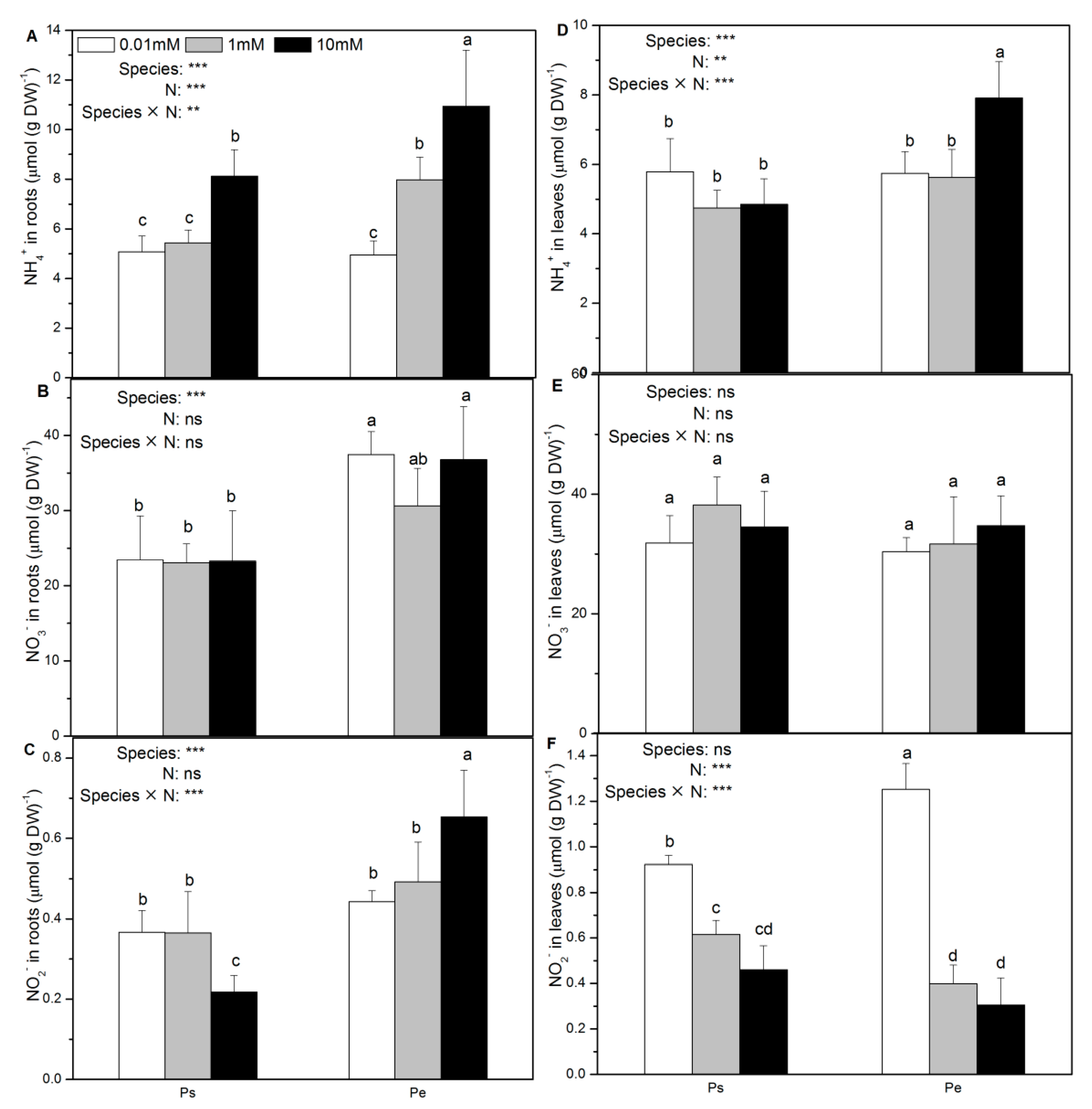
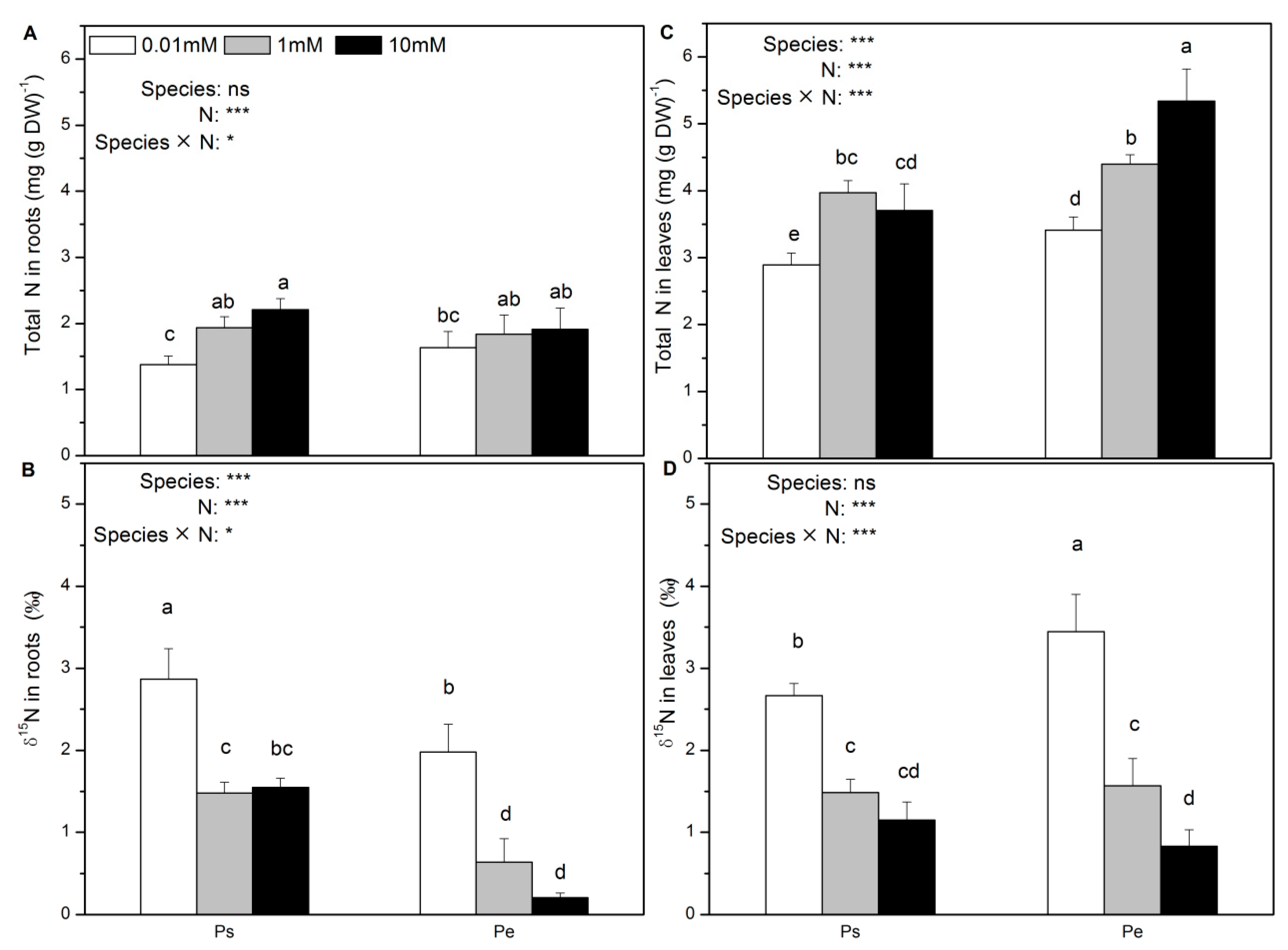
2.4. Net Fluxes of NH4+, NO3–, and H+; Accumulation of NH4+, NO3−, and NO2−; and δ15N Content
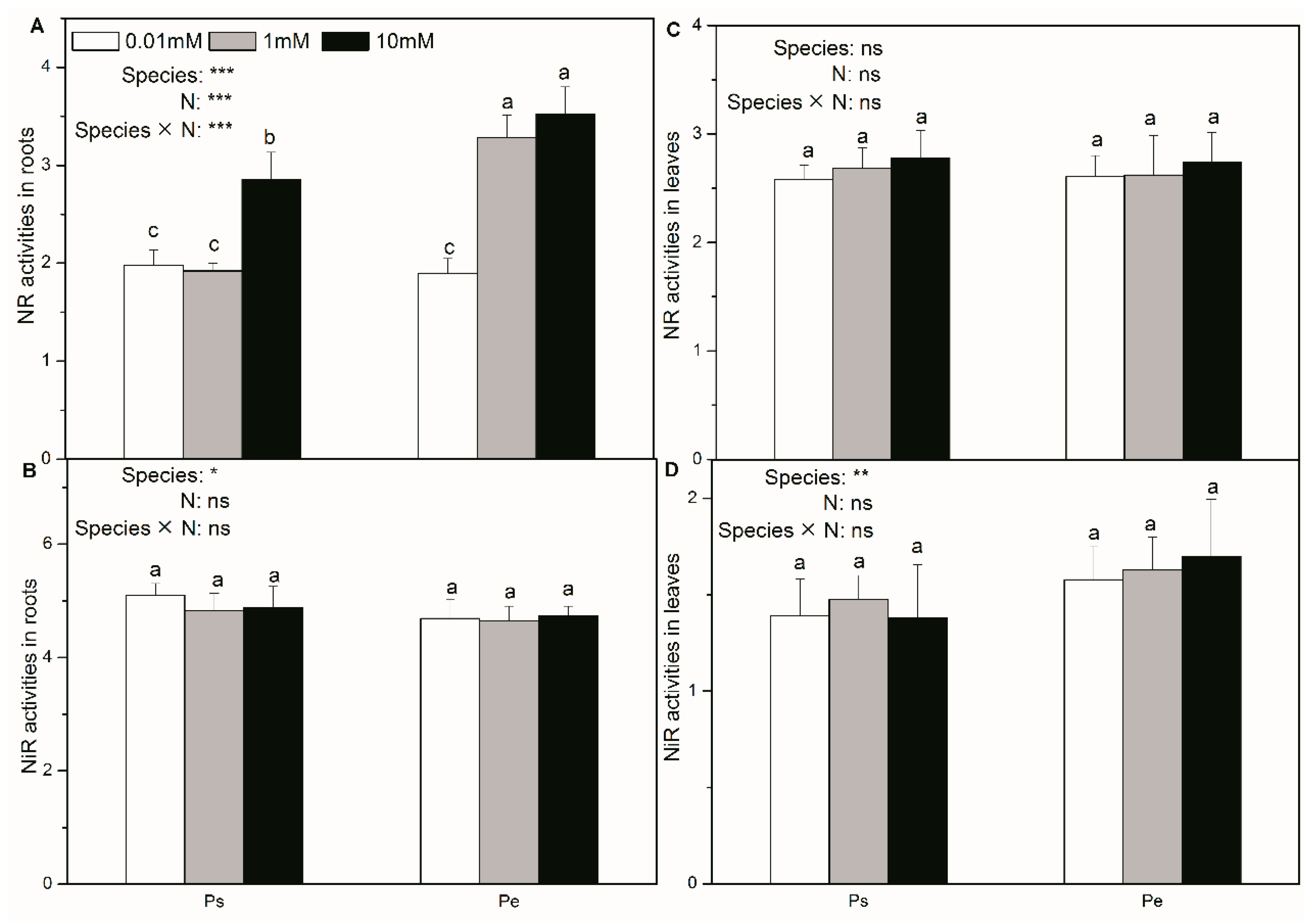
2.5. Activities of Enzymes in Roots and Leaves Involved in N Assimilation
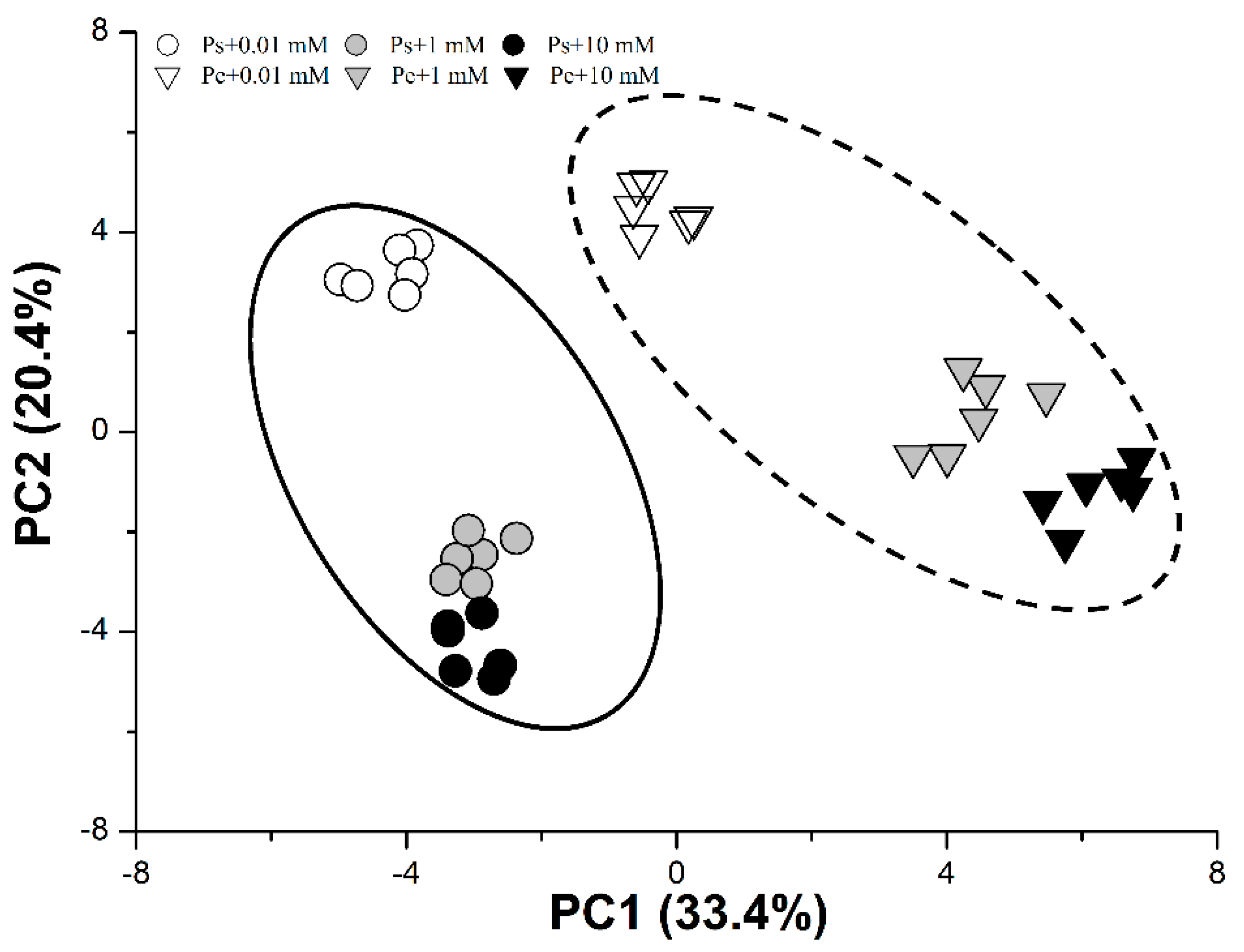
2.6. PCA of Root Characteristics and C/N Physiological Responses
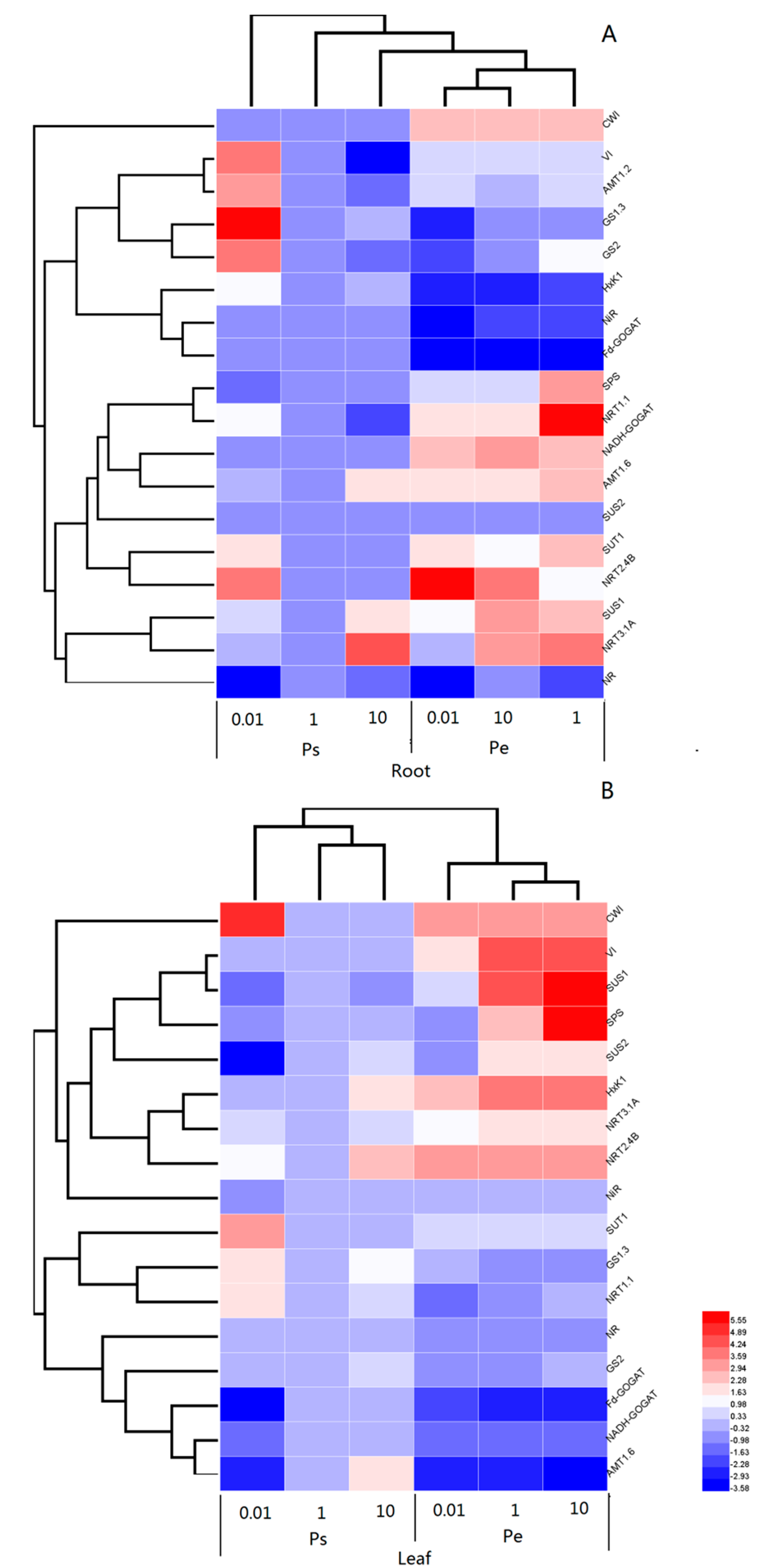
2.7. Transcript Levels of Key Genes Involved in C/N Metabolisms
3. Discussion
3.1. The Slow-Growing Ps Exhibits Enhanced Resistance to N Deficiency
3.2. The Fast Growth of Pe Relies on Sufficient N Supply
4. Materials and Methods
4.1. Plant Growth Conditions and Treatments
4.2. Photosynthetic Characteristics and Root Development
4.3. Determination of Contents of Sucrose, Fructose, Glucose, and Total Carbon
4.4. Determination of Activities of Enzymes Involved in C Metabolism in Roots and Leaves
4.5. Determination of Net Fluxes of NH4+, NO3−, and H+
4.6. Determination of Contents of NH4+, NO3−, NO2−, δ15N, and Total N
4.7. Determination of Activities of Enzymes Involved in N Metabolism in Roots and Leaves
4.8. RNA Extraction and Analysis of the Transcript Levels of Key Genes Involved in C/N Metabolism
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
| AMTs | Ammonium transporters |
| CWI | Cell wall invertase |
| GDH | Glutamate dehydrogenase |
| GOGAT | Glutamate synthase |
| GS | Glutamine synthetase |
| HxKs | Hexokinases |
| NiR | Nitrite reductase |
| NR | Nitrate reductase |
| NRTs | Nitrate transporters |
| SPP | Sucrose-6-phosphate phosphatase |
| SPS | Sucrose phosphate synthase |
| SUS | Sucrose synthase |
| SUT | Sucrose transporter |
| UDP-G | Uridine diphosphate glucose |
| VI | Vacuolar invertase |
References
- Liu, L.; Peng, D.; Wang, Z.; Hu, Y. Improving artificial forest biomass estimates using afforestation age information from time series Landsat stacks. Environ. Monit. Assess. 2014, 186, 7293–7306. [Google Scholar] [CrossRef] [PubMed]
- Polle, A.; Janz, D.; Teichmann, T.; Lipka, V. Poplar genetic engineering: Promoting desirable wood characteristics and pest resistance. Appl. Microbiol. Biotechnol. 2013, 97, 5669–5679. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Li, H.; Liu, T.; Polle, A.; Peng, C.; Luo, Z.-B. Nitrogen metabolism of two contrasting poplar species during acclimation to limiting nitrogen availability. J. Exp. Bot. 2013, 64, 4207–4224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Z.B.; Ke, L.; Jiang, X.; Polle, A. Ectomycorrhizal fungus (Paxillus involutus) and hydrogels affect performance of Populus euphratica exposed to drought stress. Ann. For. Sci. 2009, 66, 106. [Google Scholar] [CrossRef]
- Euring, D.; Löfke, C.; Teichmann, T.; Polle, A. Nitrogen fertilization has differential effects on N allocation and lignin in two Populus species with contrasting ecology. Trees 2012, 26, 1933–1942. [Google Scholar] [CrossRef]
- Novaes, E.; Osorio, L.; Drost, D.R.; Miles, B.L.; Boaventura-Novaes, C.R.D.; Benedict, C.; Dervinis, C.; Yu, Q.; Sykes, R.; Davis, M. Quantitative genetic analysis of biomass and wood chemistry of Populus under different nitrogen levels. New Phytol. 2009, 182, 878–890. [Google Scholar] [CrossRef] [PubMed]
- Cooke, J.E.; Weih, M. Nitrogen storage and seasonal nitrogen cycling in Populus: Bridging molecular physiology and ecophysiology. New Phytol. 2005, 167, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Li, M.; Luo, J.; Cao, X.; Qu, L.; Gai, Y.; Jiang, X.; Liu, T.; Bai, H.; Janz, D. N-fertilization has different effects on the growth, carbon and nitrogen physiology, and wood properties of slow- and fast-growing Populus species. J. Exp. Bot. 2012, 63, 6173–6185. [Google Scholar] [CrossRef] [PubMed]
- Ruan, Y.L. Sucrose metabolism: Gateway to diverse carbon use and sugar signaling. Plant Biol. 2014, 65, 33–67. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Tao, S.; Huo, L.; Wei, H.; Yang, J.; Xu, L.; Ma, F. Unveiling the mechanism of melatonin impacts on maize seedling growth: Sugar metabolism as a case. J. Pineal Res. 2015, 59, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Zhang, J. Research progresses on the key enzymes involved in sucrose metabolism in maize. Carbohydr. Res. 2013, 368, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Kühn, C.; Grof, C.P. Sucrose transporters of higher plants. Curr. Opin. Plant Biol. 2010, 13, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Ganeteg, U. Uptake of organic nitrogen by plants. New Phytol. 2009, 182, 31–48. [Google Scholar] [Green Version]
- Luo, J.; Qin, J.; He, F.; Li, H.; Liu, T.; Polle, A.; Peng, C.; Luo, Z.B. Net fluxes of ammonium and nitrate in association with H+ fluxes in fine roots of Populus popularis. Planta 2013, 237, 919–931. [Google Scholar] [CrossRef] [PubMed]
- Bai, H.; Euring, D.; Volmer, K.; Janz, D.; Polle, A. The nitrate transporter (NRT) gene family in poplar. PLoS ONE 2013, 8, e72126. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, B.; Boukcim, H.; Plassard, C. A comparison of ammonium, nitrate and proton net fluxes along seedling roots of Douglas-fir and lodgepole pine grown and measured with different inorganic nitrogen sources. Plant Cell Environ. 2008, 31, 278–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, S.; Zhang, C.-X.; Su, L.; Li, Y.-M.; Cao, Y.; Zhao, Z. Distinct effect of pH on N uptake and assimilation in two conifer species. Trees 2016, 30, 1607–1618. [Google Scholar] [CrossRef]
- Sorgona, A.; Lupini, A.; Mercati, F.; Di Dio, L.; Sunseri, F.; Abenavoli, M.R. Nitrate uptake along the maize primary root: An integrated physiological and molecular approach. Plant Cell Environ. 2011, 34, 1127–1140. [Google Scholar] [CrossRef] [PubMed]
- Pii, Y.; Alessandrini, M.; Guardini, K.; Zamboni, A.; Varanini, Z. Induction of high-affinity NO3– uptake in grapevine roots is an active process correlated to the expression of specific members of the NRT2 and plasma membrane H+-ATPase gene families. Funct. Plant Biol. 2014, 41, 353–365. [Google Scholar] [CrossRef]
- Kalcsits, L.A.; Buschhaus, H.A.; Guy, R.D. Nitrogen isotope discrimination as an integrated measure of nitrogen fluxes, assimilation and allocation in plants. Physiol. Plant. 2014, 151, 293–304. [Google Scholar] [CrossRef] [PubMed]
- Evans, R.D. Physiological mechanisms influencing plant nitrogen isotope composition. Trends Plant Sci. 2001, 6, 121–126. [Google Scholar] [CrossRef]
- Meng, S.; Su, L.; Li, Y.; Wang, Y.; Zhang, C.; Zhao, Z. Nitrate and ammonium contribute to the distinct nitrogen metabolism of Populus simonii during moderate salt stress. PLoS ONE 2016, 11, e0150354. [Google Scholar] [CrossRef] [PubMed]
- Meng, S.; Zhang, C.; Su, L.; Li, Y.; Zhao, Z. Nitrogen uptake and metabolism of Populus simonii in response to PEG-induced drought stress. Environ. Exp. Bot. 2016, 123, 78–87. [Google Scholar] [CrossRef]
- Liu, Z.P.; Shao, M.A.; Wang, Y.Q. Large-scale spatial interpolation of soil pH across the Loess Plateau, China. Environ. Earth Sci. 2013, 69, 2731–2741. [Google Scholar] [CrossRef]
- Chapman, N.; Miller, A.J.; Lindsey, K.; Whalley, W.R. Roots, water, and nutrient acquisition: Let’s get physical. Trends Plant Sci. 2012, 17, 701–710. [Google Scholar] [CrossRef] [PubMed]
- Kraiser, T.; Gras, D.E.; Gutiérrez, A.G.; González, B.; Gutiérrez, R.A. A holistic view of nitrogen acquisition in plants. J. Exp. Bot. 2011, 62, 1455–1466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.; Yang, Y.; Wang, G.; Yang, L.; Sun, X. Ecophysiological responses of Abies fabri seedlings to drought stress and nitrogen supply. Physiol. Plant. 2010, 139, 335–347. [Google Scholar] [PubMed]
- Zhao, H.; Xu, L.; Tao, S.; Yang, J.; Hu, L.; Ma, F. Melatonin regulates carbohydrate metabolism and defenses against Pseudomonas syringae pv. tomato DC3000 infection in Arabidopsis thaliana. J. Pineal Res. 2015, 59, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Schlüter, U.; Mascher, M.; Colmsee, C.; Scholz, U.; Bräutigam, A.; Fahnenstich, H.; Sonnewald, U. Maize source leaf adaptation to nitrogen deficiency affects not only nitrogen and carbon metabolism but also control of phosphate homeostasis. Plant Physiol. 2012, 160, 1384–1406. [Google Scholar] [CrossRef] [PubMed]
- Karve, R.; Lauria, M.; Virnig, A.; Xia, X.X.; Rauh, B.L.; Moore, B.D. Evolutionary lineages and functional diversification of plant hexokinases. Mol. Plant 2010, 3, 334–346. [Google Scholar] [CrossRef] [PubMed]
- Jia, J.J.; Li, S.L.; Cao, X.; Li, H.; Shi, W.; Polle, A.; Liu, T.X.; Peng, C.; Luo, Z.B. Physiological and transcriptional regulation in poplar roots and leaves during acclimation to high temperature and drought. Physiol. Plant. 2016, 157, 38–53. [Google Scholar] [CrossRef] [PubMed]
- Tcherkez, G. Natural 15N/14N isotope composition in C3 leaves: Are enzymatic isotope effects informative for predicting the 15N-abundance in key metabolites? Funct. Plant Biol. 2010, 38, 1–12. [Google Scholar] [CrossRef]
- Cernusak, L.A.; Winter, K.; Turner, B.L. Plant δ15N correlates with the transpiration efficiency of nitrogen acquisition in tropical trees. Plant Physiol. 2009, 151, 1667–1676. [Google Scholar] [CrossRef] [PubMed]
- Yousfi, S.; Serret, M.D.; Araus, J.L. Comparative response of δ13C, δ18O and δ15N in durum wheat exposed to salinity at the vegetative and reproductive stages. Plant Cell Environ. 2013, 36, 1214–1227. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, P.P.; Lamothe, M.; Mahé, A.; Molero, G.; Nogués, S.; Hodges, M.; Tcherkez, G. Metabolic origin of δ 15N values in nitrogenous compounds from Brassica napus L. leaves. Plant Cell Environ. 2012, 36, 128–137. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Gao, Y.; Ding, L.; Shen, Q.; Guo, S. Ammonium enhances the tolerance of rice seedlings (Oryza sativa L.) to drought condition. Agric. Water Manag. 2009, 96, 1746–1750. [Google Scholar] [CrossRef]
- Bonneau, L.; Huguet, S.; Wipf, D.; Pauly, N.; Truong, H.N. Combined phosphate and nitrogen limitation generates a nutrient stress transcriptome favorable for arbuscular mycorrhizal symbiosis in Medicago truncatula. New Phytol. 2013, 199, 188–202. [Google Scholar] [CrossRef] [PubMed]
- Kiba, T.; Feria-Bourrellier, A.-B.; Lafouge, F.; Lezhneva, L.; Boutet-Mercey, S.; Orsel, M.; Bréhaut, V.; Miller, A.; Daniel-Vedele, F.; Sakakibara, H. The Arabidopsis nitrate transporter NRT2.4 plays a double role in roots and shoots of nitrogen-starved plants. Plant Cell 2012, 24, 245–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Couturier, J.; Montanini, B.; Martin, F.; Brun, A.; Blaudez, D.; Chalot, M. The expanded family of ammonium transporters in the perennial poplar plant. New Phytol. 2007, 174, 137–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pii, Y.; Alessandrini, M.; Dal’Osto, L.; Guardini, K.; Prinsi, B.; Espen, L.; Zamboni, A.; Varanini, Z. Time-resolved investigation of molecular components involved in the induction of high affinity transport system in maize roots. Front. Plant Sci. 2016, 7, 1657. [Google Scholar] [CrossRef] [PubMed]
- Sperandio, M.V.; Santos, L.A.; Bucher, C.A.; Fernandes, M.S.; de Souza, S.R. Isoforms of plasma membrane H+-ATPase in rice root and shoot are differentially induced by starvation and resupply of NO3− or NH4+. Plant Sci. 2011, 180, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Selle, A.; Willmann, M.; Grunze, N.; Geßler, A.; Weiß, M.; Nehls, U. The high-affinity poplar ammonium importer PttAMT1.2 and its role in ectomycorrhizal symbiosis. New Phytol. 2005, 168, 697–706. [Google Scholar] [CrossRef] [PubMed]
- Lawlor, D.W. Carbon and nitrogen assimilation in relation to yield: Mechanisms are the key to understanding production systems. J. Exp. Bot. 2002, 53, 773–787. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Fan, X.; Miller, A.J. Plant nitrogen assimilation and use efficiency. Annu. Rev. Plant Biol. 2012, 63, 153–182. [Google Scholar] [CrossRef] [PubMed]
- Frew, A.J.; Kay, A.B. Gene expression associated with N-induced shifts in resource allocation in poplar. Plant Cell Environ. 2003, 26, 757–770. [Google Scholar] [Green Version]
- Falxa-Raymond, N.; Patterson, A.E.; Schuster, W.S.; Griffin, K.L. Oak loss increases foliar nitrogen, δ15N and growth rates of Betula lenta in a northern temperate deciduous forest. Tree Physiol. 2012, 32, 1092–1101. [Google Scholar] [CrossRef] [PubMed]
- Gan, H.; Jiao, Y.; Jia, J.; Wang, X.; Li, H.; Shi, W.; Peng, C.; Polle, A.; Luo, Z.B. Phosphorus and nitrogen physiology of two contrasting poplar genotypes when exposed to phosphorus and/or nitrogen starvation. Tree Phys. 2016, 36, 22–38. [Google Scholar] [CrossRef] [PubMed]
- Zamboni, A.; Astolfi, S.; Zuchi, S.; Pii, Y.; Guardini, K.; Tononi, P.; Varanini, Z. Nitrate induction triggers different transcriptional changes in a high and a low nitrogen use efficiency maize inbred line. J. Integr. Plant Biol. 2014, 56, 1080–1094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, X.; Jia, J.; Zhang, C.; Li, H.; Liu, T.; Jiang, X.; Polle, A.; Peng, C.; Luo, Z.B. Anatomical, physiological and transcriptional responses of two contrasting poplar genotypes to drought and re-watering. Physiol. Plant. 2014, 151, 480–494. [Google Scholar] [CrossRef] [PubMed]
- Link, M.; Rausch, T.; Greiner, S. In Arabidopsis thaliana, the invertase inhibitors AtC/VIF1 and 2 exhibit distinct target enzyme specificities and expression profiles. FEBS Lett. 2004, 573, 105–109. [Google Scholar] [CrossRef] [PubMed]
- Patterson, K.; Cakmak, T.; Cooper, A.; Lager, I.; Rasmusson, A.G.; Escobar, M.A. Distinct signalling pathways and transcriptome response signatures differentiate ammonium-and nitrate-supplied plants. Plant Cell Environ. 2010, 33, 1486–1501. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, T.; Fukuoka, H.; Yano, H.; Ohkawa, Y. Relationships between nitrite reductase activity and genotype-dependent callus growth in rice cell cultures. Plant Cell Rep. 1999, 18, 576–581. [Google Scholar] [CrossRef]
- Natali, S.M.; Sañudo-Wilhelmy, S.A.; Lerdau, M.T. Effects of elevated carbon dioxide and nitrogen fertilization on nitrate reductase activity in sweetgum and loblolly pine trees in two temperate forests. Plant Soil. 2009, 314, 197–210. [Google Scholar] [CrossRef]
- Seith, B.; Setzer, B.; Flaig, H.; Mohr, H. Appearance of nitrate reductase, nitrite reductase and glutamine synthetase in different organs of the Scots pine (Pinus sylvestris) seedling as affected by light, nitrate and ammonium. Physiol. Plant. 1994, 91, 419–426. [Google Scholar] [CrossRef]
- Wang, L.; Zhou, Q.; Ding, L.; Sun, Y. Effect of cadmium toxicity on nitrogen metabolism in leaves of Solanum nigrum L. as a newly found cadmium hyperaccumulator. J. Hazard. Mater. 2008, 154, 818–825. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.C.; Kao, C.H. Disturbed ammonium assimilation is associated with growth inhibition of roots in rice seedlings caused by NaCl. Plant Growth Regul. 1996, 18, 233–238. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | N Supply (mM) | Root Biomass (mg DW) | Total Fine Root Length (m) | Total Fine Root Surface Area (cm2) | Total Root Volume(cm3) | SPAD Values | A (μmol CO2 m−2 s−1) | gs (mol H2O m−2 s−1) | E (mmol H2O m−2 s−1) |
|---|---|---|---|---|---|---|---|---|---|
| Ps | 0.01 | 5.74 ± 0.79 bc | 13.62 ± 1.60 c | 57.55 ± 5.62 c | 12.72 ± 1.38 bc | 22.90 ± 3.44 b | 11.86 ± 1.03 a | 0.22 ± 0.02 ab | 4.85 ± 0.37 a |
| 1 | 3.93 ± 0.53 d | 11.77 ± 1.58 cd | 40.25 ± 3.53 d | 11.73 ± 1.13 c | 28.82 ± 2.74 a | 9.75 ± 0.61 b | 0.26 ± 0.02 a | 4.97 ± 0.46 a | |
| 10 | 2.73 ± 0.35 e | 9.53 ± 1.18 d | 48.21 ± 3.67 cd | 9.73 ± 0.98 c | 31.23 ± 2.88 a | 10.23 ± 0.82 ab | 0.22 ± 0.05 ab | 4.93 ± 0.41 a | |
| Pe | 0.01 | 4.93 ± 0.58 c | 15.14 ± 1.60 bc | 98.66 ± 11.45 b | 12.72 ± 2.21 bc | 14.58 ± 1.11 c | 6.25 ± 1.03 c | 0.09 ± 0.02 d | 2.67 ± 0.21 c |
| 1 | 7.37 ± 0.54 a | 22.95 ± 1.59 a | 130.24 ± 12.89 a | 17.92 ± 3.05 a | 29.77 ± 1.87 a | 9.45 ± 0.71 b | 0.15 ± 0.02 c | 3.26 ± 0.32 bc | |
| 10 | 6.18 ± 0.49 b | 17.75 ± 4.05 b | 109.05 ± 12.47 b | 15.85 ± 1.30 ab | 31.22 ± 2.75 a | 11.09 ± 1.56 ab | 0.18 ± 0.03 bc | 3.69 ± 0.20 b | |
| p-values | Species | *** | *** | *** | *** | ** | *** | *** | *** |
| N | *** | *** | ns | * | *** | ** | *** | ** | |
| Species × N | *** | *** | *** | *** | *** | *** | ** | ** |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meng, S.; Wang, S.; Quan, J.; Su, W.; Lian, C.; Wang, D.; Xia, X.; Yin, W. Distinct Carbon and Nitrogen Metabolism of Two Contrasting Poplar Species in Response to Different N Supply Levels. Int. J. Mol. Sci. 2018, 19, 2302. https://doi.org/10.3390/ijms19082302
Meng S, Wang S, Quan J, Su W, Lian C, Wang D, Xia X, Yin W. Distinct Carbon and Nitrogen Metabolism of Two Contrasting Poplar Species in Response to Different N Supply Levels. International Journal of Molecular Sciences. 2018; 19(8):2302. https://doi.org/10.3390/ijms19082302
Chicago/Turabian StyleMeng, Sen, Shu Wang, Jine Quan, Wanlong Su, Conglong Lian, Dongli Wang, Xinli Xia, and Weilun Yin. 2018. "Distinct Carbon and Nitrogen Metabolism of Two Contrasting Poplar Species in Response to Different N Supply Levels" International Journal of Molecular Sciences 19, no. 8: 2302. https://doi.org/10.3390/ijms19082302
APA StyleMeng, S., Wang, S., Quan, J., Su, W., Lian, C., Wang, D., Xia, X., & Yin, W. (2018). Distinct Carbon and Nitrogen Metabolism of Two Contrasting Poplar Species in Response to Different N Supply Levels. International Journal of Molecular Sciences, 19(8), 2302. https://doi.org/10.3390/ijms19082302