The γ-Aminobutyric Acid (GABA) Alleviates Salt Stress Damage during Seeds Germination of White Clover Associated with Na+/K+ Transportation, Dehydrins Accumulation, and Stress-Related Genes Expression in White Clover



 ,
,
Abstract
:1. Introduction
2. Results
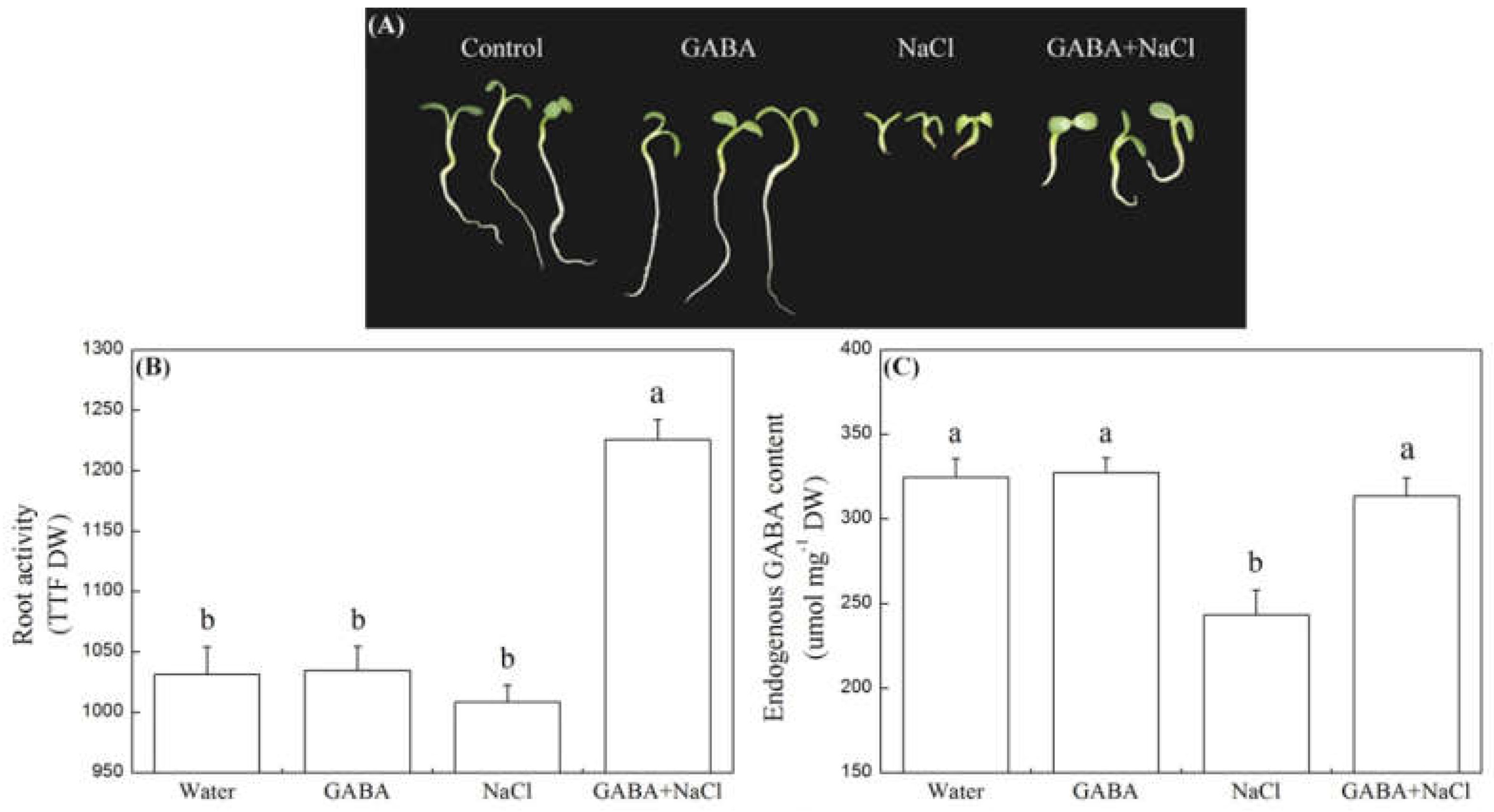
2.1. Effects of the GABA on Seed Germination Characteristics
2.2. Effect of the GABA on Root Activity and Endogenous GABA Content
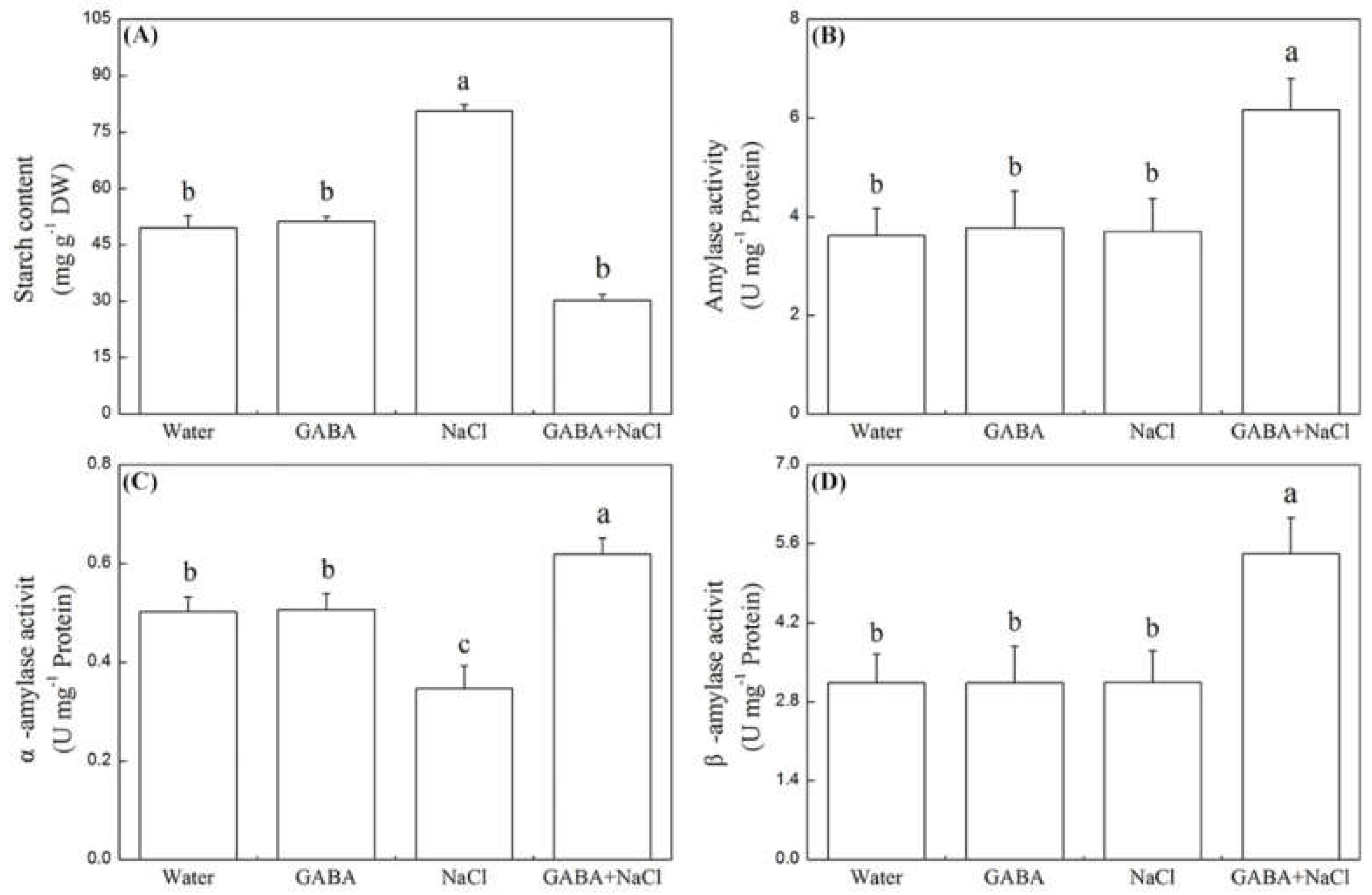
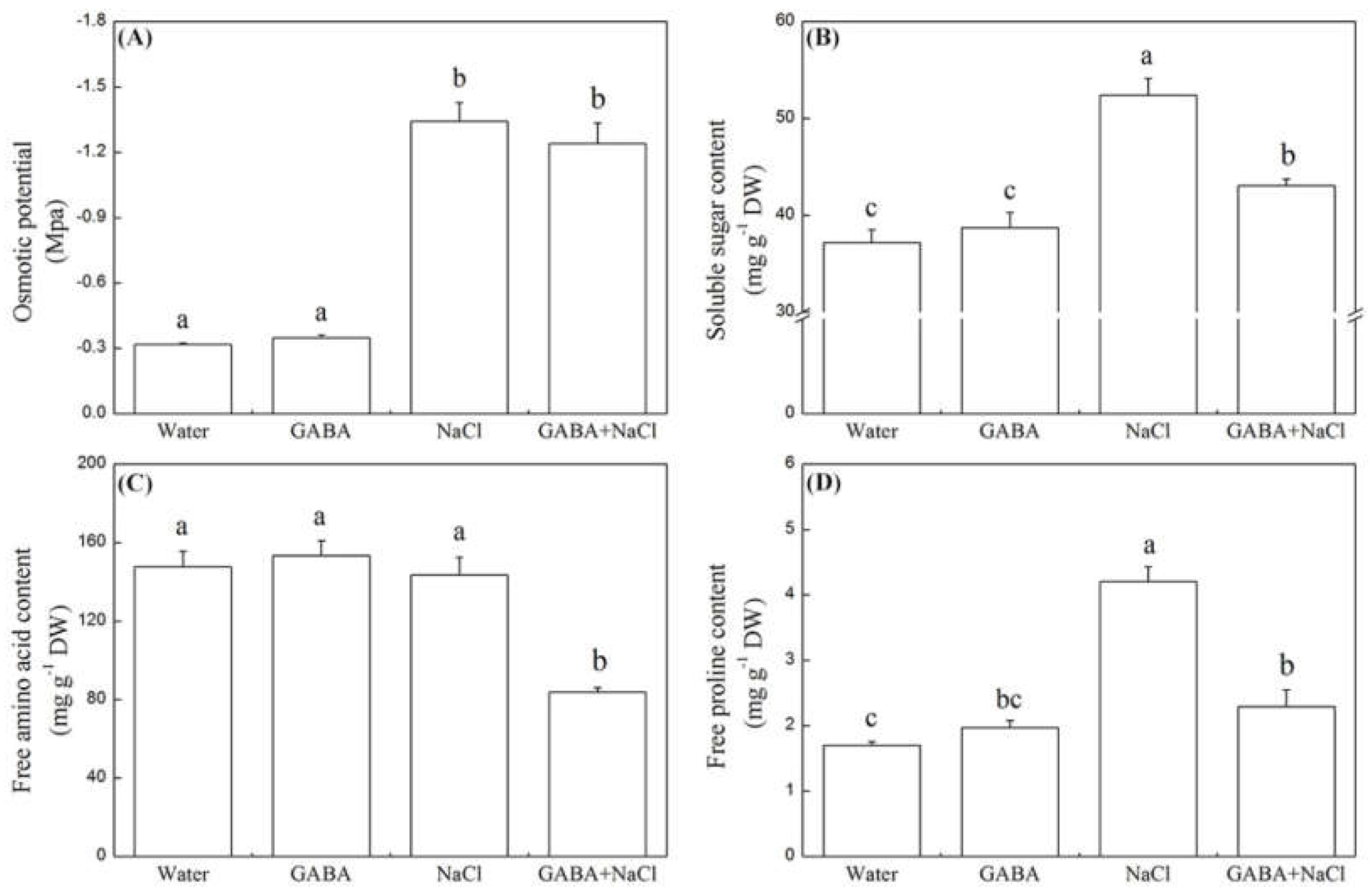
2.3. Effects of the GABA on Starch Metabolism and Osmotic Adjustment
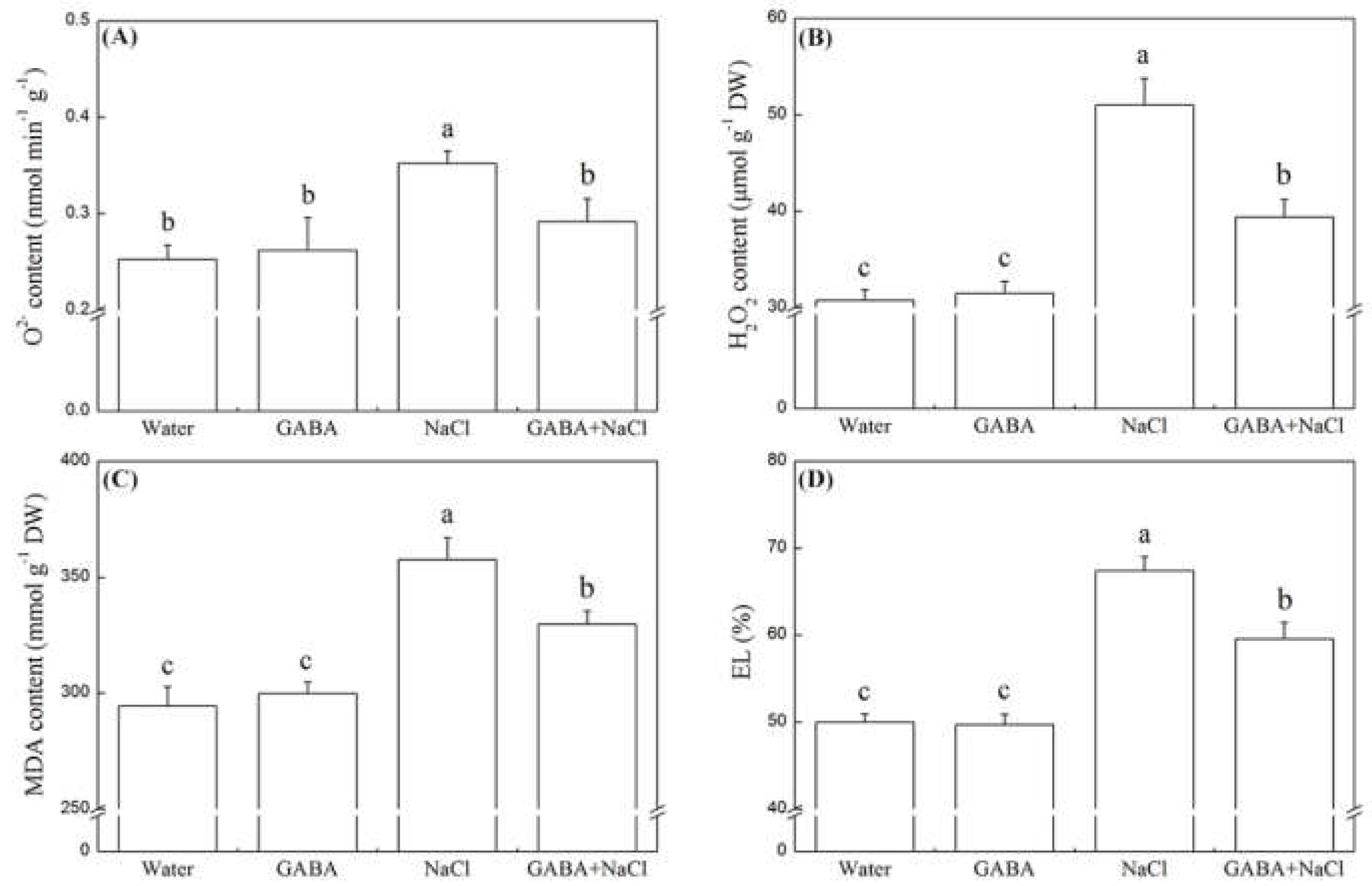
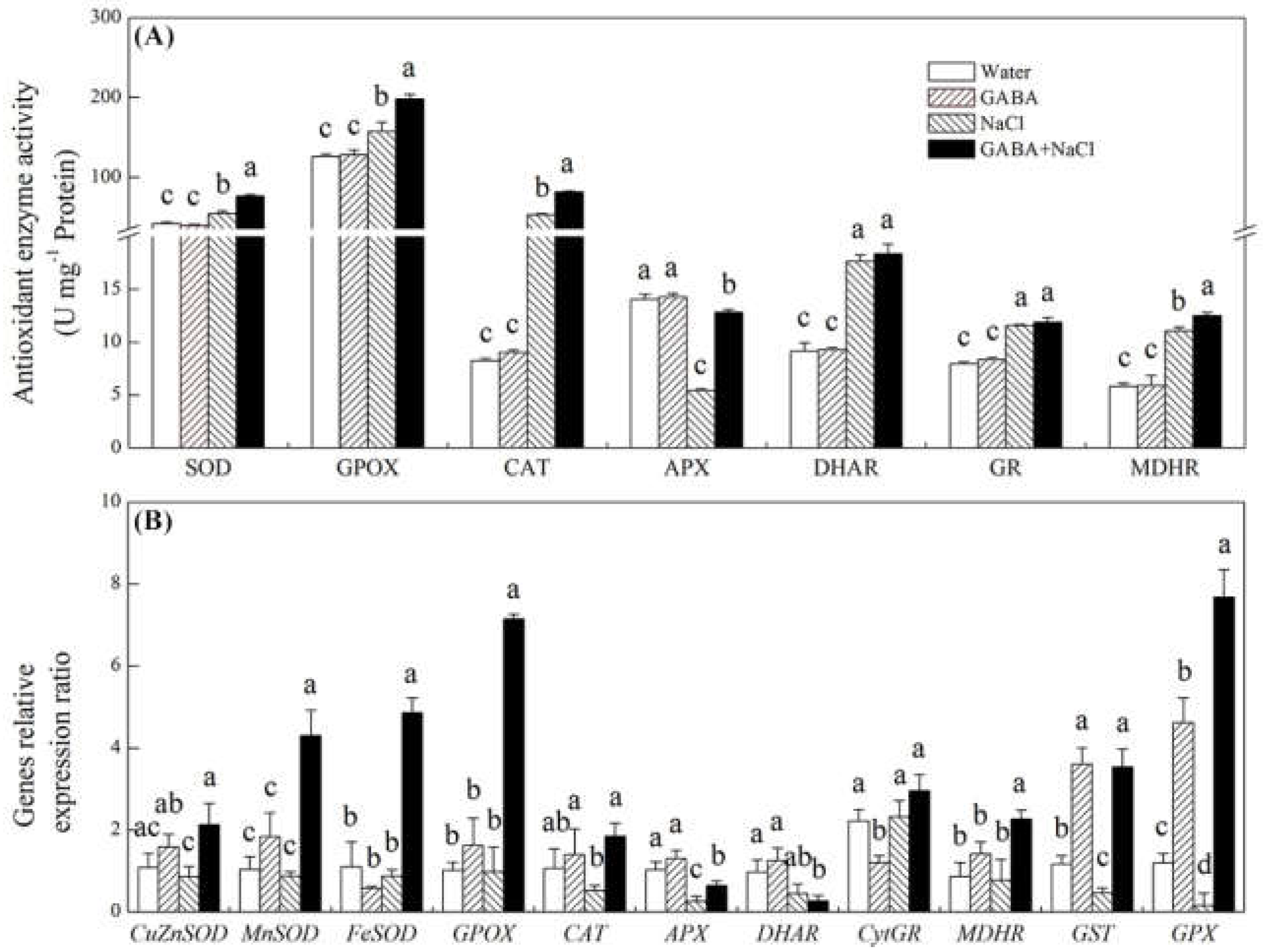
2.4. Effects of the GABA on Antioxidant Defense and Oxidative Damage
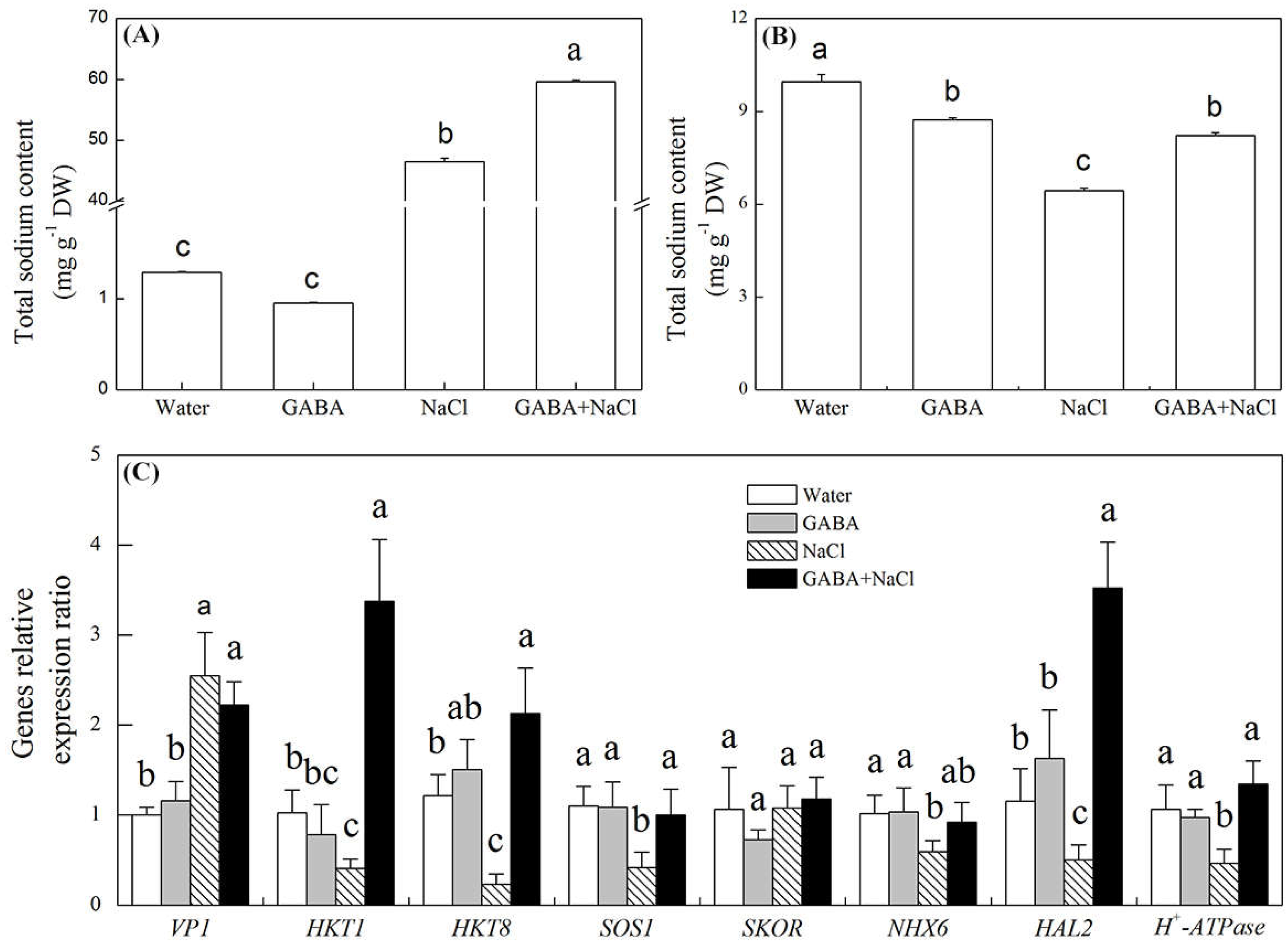
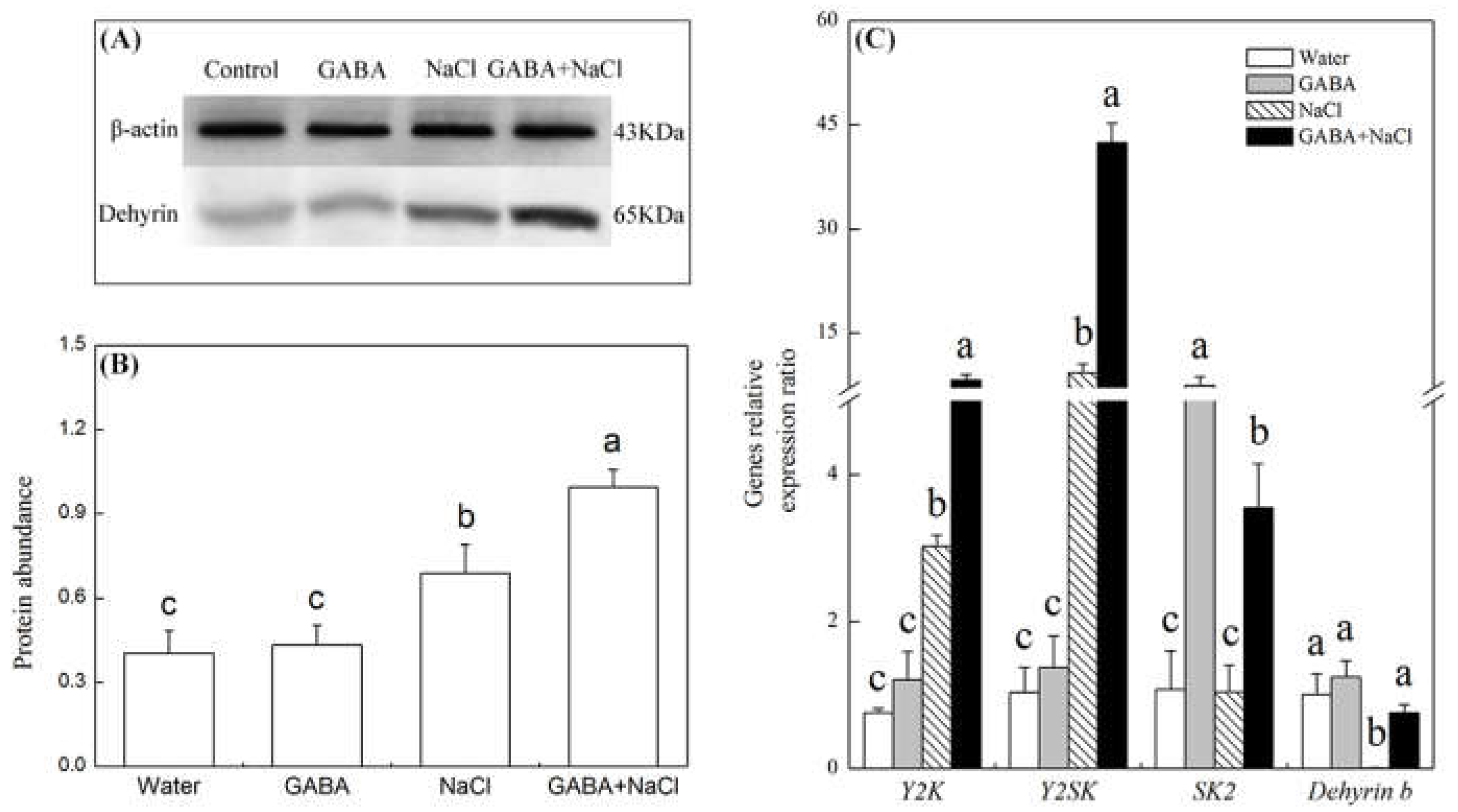
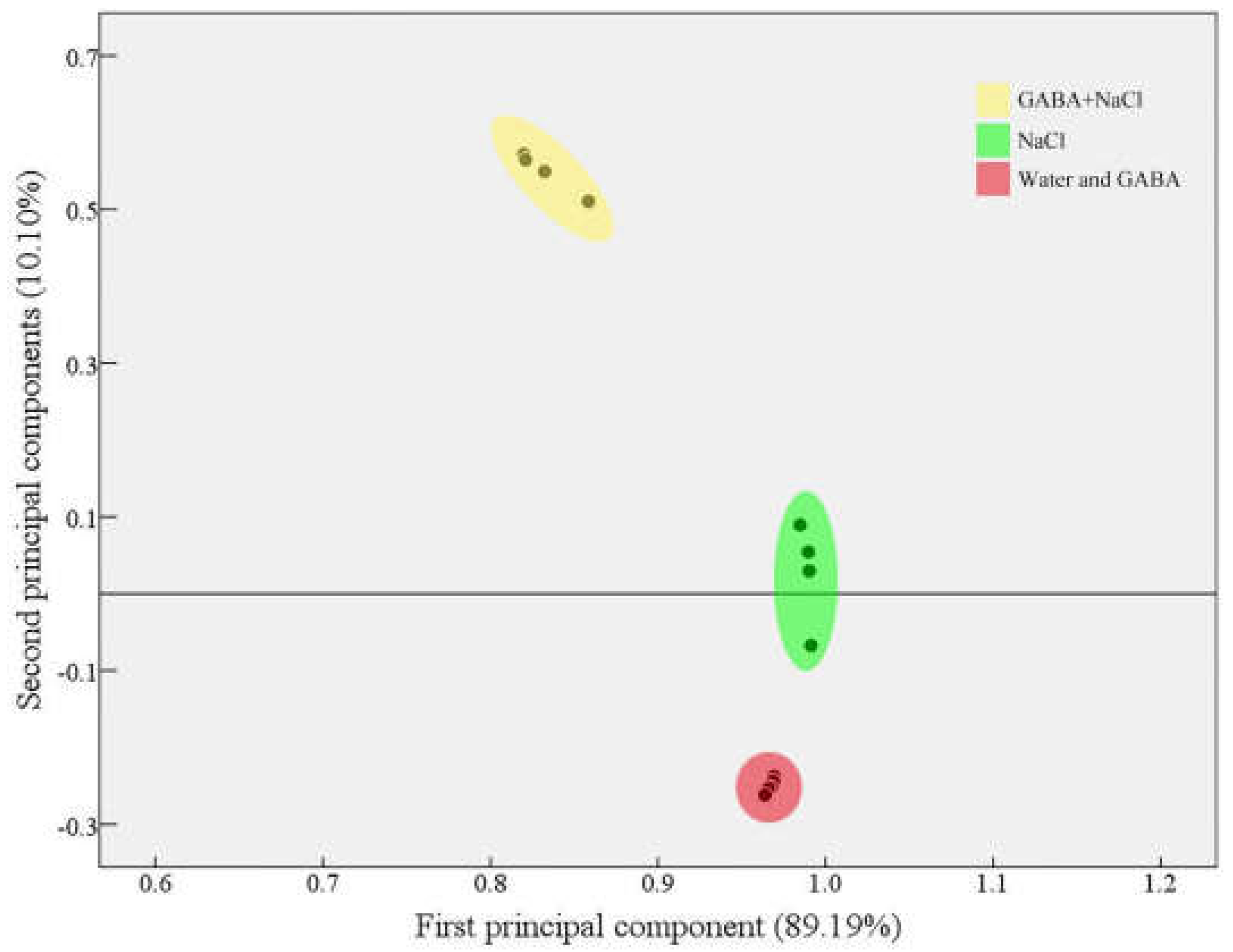
2.5. Effects of the GABA on Accumulation of Dehydrins and Genes Relative Expression
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Treatments
4.2. Determination of Seed Germination Characteristics, Root Viability and Endogenous GABA Content
4.3. Determination of Starch Metabolism, Amino Acids, and Osmotic Potential
4.4. Determination of Antioxidant Enzyme Activities and Oxidative Damage
4.5. Determination of Na+/K+ Content and Western Blot Analysis
4.6. Genes Expression Analysis
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Munns, R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashraf, M. Effect of sodium chloride on water relations and some organic osmotica in arid zone plant species Melilotus indica (L.) All. Tropenlandwirt 1993, 94, 95–102. [Google Scholar]
- Al-Karaki, G. Barley response to salt stress at varied levels of phosphorus. J. Plant Nutr. 1997, 20, 1635–1643. [Google Scholar] [CrossRef]
- Bybordi, A.; Tabatabaei, S.J.; Ahmedov, A. Effect of salinity on the growth and peroxidase and iaa oxidase activities in canola. J. Food Agric. Environ. 2010, 8, 109–112. [Google Scholar]
- Bybordi, A.; Tabatabaei, S.J.; Ahmedov, A. Effects of salinity on fatty acid composition of canola (Brassica napus L.). J. Food Agric. Environ. 2010, 8, 113–115. [Google Scholar]
- Woodrow, P.; Ciarmiello, L.F.; Annunziata, M.G.; Pacifico, S.; Iannuzzi, F.; Mirto, A.; D’Amelia, L.; Dell’Aversana, E.; Piccolella, S.; Fuggi, A.; et al. Durum wheat seedling responses to simultaneous high light and salinity involve a fine reconfiguration of amino acids and carbohydrate metabolism. Physiol. Plant. 2016, 159, 290–312. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Xu, S.; Xuan, W.; Ling, T.; Cao, Z.; Huang, B.; Sun, Y.; Fang, L.; Liu, Z.; Zhao, N. Carbon monoxide counteracts the inhibition of seed germination and alleviates oxidative damage caused by salt stress in oryza sativa. Plant Sci. 2007, 172, 544–555. [Google Scholar] [CrossRef]
- Cuartero, J.; Bolarín, M.C.; Asíns, M.J.; Moreno, V. Increasing salt tolerance in the tomato. J. Exp. Bot. 2006, 57, 1045–1058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbagallo, A.; Nicola, A.D.; Missikoff, M. The influence of salt stress on seed germination, growth and yield of canola cultivars. Notulae Bot. Horti Agrobotanici Cluj-Napoca. 2010, 38, 128–133. [Google Scholar]
- Zhang, S.; Hu, J.; Zhang, Y.; Xie, X.J.; Knapp, A. Seed priming with brassinolide improves lucerne (Medicago sativa L.) seed germination and seedling growth in relation to physiological changes under salinity stress. Aust. J. Agric. Res. 2007, 58, 811–815. [Google Scholar] [CrossRef]
- Duan, D.Y.; Li, W.Q.; Liu, X.J.; Ouyang, H.; An, P.; Duan, D.Y.; Liu, W.Q. Seed germination and seedling growth of suaeda salsa under salt stress. Ann. Bot. Fenn. 2007, 44, 161–169. [Google Scholar]
- Kinnersley, A.M.; Turano, F.J. Gamma aminobutyric acid (GABA) and plant responses to stress. Crit. Rev. Plant Sci. 2000, 19, 479–509. [Google Scholar] [CrossRef]
- Shelp, B.J.; Bown, A.W.; Mclean, M.D. Metabolism and function of γ-aminobutyric acid. Trends Plant Sci. 1999, 4, 446–452. [Google Scholar] [CrossRef]
- Shi, S.Q.; Zheng, S.; Jiang, Z.P.; Qi, L.W.; Sun, X.M.; Li, C.X.; Liu, J.F.; Xiao, W.F.; Zhang, S.G. Effects of exogenous GABA on gene expression of caragana intermedia roots under NaCl stress: Regulatory roles for H2O2 and ethylene production. Plant Cell Environ. 2010, 33, 149–162. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Fan, Y.; Ma, Y.; Zhang, Z.; Yue, H.; Wang, L.; Li, J.; Jiao, Y. Effects of exogenous γ-aminobutyric acid (GABA) on photosynthesis and antioxidant system in pepper (Capsicum annuum L.) seedlings under low light stress. J. Plant Growth Regul. 2017, 36, 1–14. [Google Scholar] [CrossRef]
- Barbosa, J.M.; Singh, N.K.; Cherry, J.H.; Locy, R.D. Nitrate uptake and utilization is modulated by exogenous γ-aminobutyric acid in arabidopsis thaliana seedlings. Plant Physiol. Biochem. 2010, 48, 443–450. [Google Scholar] [CrossRef] [PubMed]
- Carillo, P. GABA shunt in durum wheat. Front. Plant Sci. 2018, 9, 100. [Google Scholar] [CrossRef] [PubMed]
- Vijayakumari, K.; Puthur, J.T. γ-Aminobutyric acid (GABA) priming enhances the osmotic stress tolerance in piper nigrum linn. plants subjected to PEG-induced stress. Plant Growth Regul. 2015, 78, 1–11. [Google Scholar] [CrossRef]
- Yu, C.; Zeng, L.; Sheng, K.; Chen, F.; Zhou, T.; Zheng, X.; Yu, T. γ-Aminobutyric acid induces resistance against penicillium expansum by priming of defence responses in pear fruit. Food Chem. 2014, 159, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Malekzadeh, P.; Khara, J.; Heydari, R. Alleviating effects of exogenous gamma-aminobutiric acid on tomato seedling under chilling stress. Physiol. Mol. Biol. Plants 2014, 20, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.Y.; Yang, L.W.; Gao, H.B.; Xiao-Lei, W.U.; Liu, H.H. Physiological mechanism of GABA soaking to tomato seed germination and seedling development under NaCl stress. Acta Bot. Boreal.-Occident. Sin. 2011, 31, 2235–2242. [Google Scholar]
- Li, Z.; Peng, D.D.; Zhang, X.Q.; Peng, Y.; Chen, M.; Ma, X.; Huang, L.K.; Yan, Y.H. Na+ induces the tolerance to water stress in white clover associated with osmotic adjustment and aquaporins-mediated water transport and balance in root and leaf. Environ. Exp. Bot. 2017, 144, 1271–1279. [Google Scholar] [CrossRef]
- Khalid, M.; Bilal, M.; Hassani, D.; Hmn, I.; Wang, H.; Huang, D. Mitigation of salt stress in white clover (Trifolium repens) by azospirillum brasilense and its inoculation effect. Bot. Stud. 2017, 58, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Zadeh, H.M.; Naeini, M.S.B. Effects of salinity stress on the morphology and yield of two cultivars of canola (Brassica napus L.). J. Agron. 2007, 6, 409–414. [Google Scholar]
- Hakim, M.A.; Juraimi, A.S.; Hanafi, M.M.; Ali, E.; Ismail, M.R.; Selamat, A.; Karim, S.M. Effect of salt stress on morpho-physiology, vegetative growth and yield of rice. J. Environ. Biol. 2014, 35, 317–326. [Google Scholar] [PubMed]
- Yacoubi, R.; Job, C.; Belghazi, M.; Chaibi, W.; Job, D. Proteomic analysis of the enhancement of seed vigour in osmoprimed alfalfa seeds germinated under salinity stress. Seed Sci. Res. 2013, 23, 99–110. [Google Scholar] [CrossRef]
- Wang, Y.; Gu, W.; Yao, M.; Xie, T.; Li, L.; Jing, L.; Shi, W. γ-Aminobutyric acid imparts partial protection from salt stress injury to maize seedlings by improving photosynthesis and upregulating osmoprotectants and antioxidants. Sci. Rep. 2017, 7, 43609. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.K.; Son, T.K.; Park, S.Y.; Lee, I.J.; Lee, B.H.; Kim, H.Y.; Lee, S.C. Influences of gibberellin and auxin on endogenous plant hormone and starch mobilization during rice seed germination under salt stress. J. Environ. Biol. 2006, 27, 181–186. [Google Scholar]
- Li, Z.; Peng, Y.; Zhang, X.Q.; Ma, X.; Hang, L.K.; Yan, Y.H. Exogenous spermidine improves seed germination of white clover under water stress via involvement in starch metabolism, antioxidant defenses and relevant gene expression. Molecules 2014, 19, 18003–18024. [Google Scholar] [CrossRef] [PubMed]
- Ghoulam, C.; Foursy, A.; Fares, K. Effects of salt stress on growth, inorganic ions and proline accumulation in relation to osmotic adjustment in five sugar beet cultivars. Environ. Exp. Bot. 2002, 47, 39–50. [Google Scholar] [CrossRef]
- Saito, T.; Matsukura, C. Effect of Salt Stress on the Growth and Fruit Quality of Tomato Plants. In Abiotic Stress Biology in Horticultural Plants; Kanayama, Y., Kochetov, A., Eds.; Springer: Tokyo, Japan, 2015; pp. 3–16. [Google Scholar]
- Zhu, J.K. Regulation of ion homeostasis under salt stress. Curr. Opin. Plant Biol. 2003, 6, 441–445. [Google Scholar] [CrossRef]
- Shabala, S. Learning from halophytes: Physiological basis and strategies to improve abiotic stress tolerance in crops. Ann. Bot. 2013, 112, 1209–1221. [Google Scholar] [CrossRef] [PubMed]
- Annunziata, M.G.; Ciarmiello, L.F.; Woodrow, P.; Maximova, E.; Fuggi, A.; Carillo, P. Durum wheat roots adapt to salinity remodeling the cellular content of nitrogen metabolites and sucrose. Front. Plant Sci. 2016, 7, 2035. [Google Scholar] [CrossRef] [PubMed]
- Nayyar, H.; Kaur, R.; Kaur, S.; Singh, R. γ-Aminobutyric acid (GABA) imparts partial protection from heat stress injury to rice seedlings by improving leaf turgor and upregulating osmoprotectants and antioxidants. J. Plant Growth Regul. 2014, 33, 408–419. [Google Scholar] [CrossRef]
- Krishnan, S.; Laskowski, K.; Shukla, V.; Merewitz, E.B. Mitigation of drought stress damage by exogenous application of a non-protein amino acid gamma aminobutyric acid on perennial ryegrass. J. Am. Soc. Hort. Sci. 2013, 138, 358–366. [Google Scholar]
- Hanin, M.; Brini, F.; Ebel, C.; Toda, Y.; Takeda, S.; Masmoudi, K. Plant dehydrins and stress tolerance. Plant Signal. Behav. 2011, 6, 1503–1509. [Google Scholar] [CrossRef] [PubMed]
- Marček, T.; Tkalec, M.; Vidaković-Cifrek, Ž.; Ježić, M.; Ćurković-Perica, M. Expression of dehydrins, Hsp70, Cu/Zn Sod, andRubisco in leaves of tobacco (Nicotiana tabacum L.) dihaploids under salt stress. Vitro Cell. Dev. Biol. Plant 2016, 52, 233–240. [Google Scholar] [CrossRef]
- Ruibal, C.; Salamó, I.P.; Carballo, V.; Castro, A.; Bentancor, M.; Borsani, O.; Szabados, L.; Vidal, S. Differential contribution of individual dehydrin genes from physcomitrella patens to salt and osmotic stress tolerance. Plant Sci. 2012, 190, 89–102. [Google Scholar] [CrossRef] [PubMed]
- Hundertmark, M.; Buitink, J.; Leprince, O.; Hincha, D.K. Reduction of seed-specific dehydrins reduces seed longevity in arabidopsis thaliana. Seed Sci. Res. 2011, 21, 165–173. [Google Scholar] [CrossRef]
- Han, B.; Kermode, A.R. Dehydrin-like proteins in castor bean seeds and seedlings are differentially produced in response to ABA and water-deficit-related stresses. J. Exp. Bot. 1996, 47, 933–939. [Google Scholar] [CrossRef] [Green Version]
- Han, B.; Hughes, D.W.; Galau, G.A.; Bewley, J.D.; Kermode, A.R. Changes in late-embryogenesis-abundant (LEA) messenger rnas and dehydrins during maturation and premature drying of Ricinus communis L. seeds. Planta 1997, 201, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.P.; Chen, T.H. Molecular cloning of abscisic acid-responsive mRNAs expressed during the induction of freezing tolerance in bromegrass (Bromus inermis leyss) suspension culture. Plant Physiol. 1993, 101, 1089–1096. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Jing, W.; Peng, Y.; Zhang, X.Q.; Ma, X.; Hang, L.K.; Yan, Y.H. Spermine alleviates drought stress in white clover with different resistance by influencing carbohydrate metabolism and dehydrins synthesis. PLoS ONE 2015, 10, e0120708. [Google Scholar] [CrossRef] [PubMed]
- Knievel, D.P. Procedure for estimating ratio of live to dead root dry matter in root core samples1. Cropence 1973, 13, 124–126. [Google Scholar] [CrossRef]
- Fu, J.M.; Dernoeden, P.H. Carbohydrate level, photosynthesis, and respiration in creeping bentgrass as influenced by spring and summer coring. J. Am. Soc. Hort. Sci. 2009, 134, 41–47. [Google Scholar]
- Smith, D. Removing and analyzing total nonstructural carbohydrates from plant tissue. Jpn. J. Grassl. Sci. 1969, 17, 75–82. [Google Scholar]
- Tárrago, J.F.; Nicolás, G. Starch degradation in the cotyledons of germinating lentils. Plant Physiol. 1976, 58, 618–621. [Google Scholar] [CrossRef] [PubMed]
- Kishorekumar, A.; Jaleel, C.A.; Manivannan, P.; Sankar, B.; Sridharan, R.; Panneerselvam, R. Comparative effects of different triazole compounds on growth, photosynthetic pigments and carbohydrate metabolism of solenostemon rotundifolius. Colloids Surf. B Biointerfaces 2007, 60, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Blum, A. Osmotic adjustment and growth of barley genotypes under drought stress. Crop Sci. 1989, 29, 230–233. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Ries, S.K. Superoxide dismutases: I. Occurrence in higher plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Dhindsa, R.S.; Plumb-Dhindsa, P.; Thorpe, T.A. Leaf senescence: Correlated with increased levels of membrane permeability and lipid peroxidation, and decreased levels of superoxide dismutase and catalase. J. Exp. Bot. 1981, 32, 93–101. [Google Scholar] [CrossRef]
- Elstner, E.F.; Heupel, A. Inhibition of nitrite formation from hydroxylammonium chloride: A simple assay for superoxide dismutase. Anal. Biochem. 1976, 70, 616–620. [Google Scholar] [CrossRef]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Blum, A.; Ebercon, A. Cell membrane stability as a measure of drought and heat tolerance in wheat. Crop Sci. 1981, 21, 43–47. [Google Scholar] [CrossRef]
- Khedr, A.H.; Abbas, M.A.; Wahid, A.A.; Quick, W.P.; Abogadallah, G.M. Proline induces the expression of salt-stress-responsive proteins and may improve the adaptation of pancratium maritimum L. to salt-stress. J. Exp. Bot. 2003, 54, 2553–2562. [Google Scholar] [CrossRef] [PubMed]
- Vaseva, I.; Akiscan, Y.; Demirevska, K.; Anders, I.; Feller, U. Drought stress tolerance of red and white clover–comparative analysis of some chaperonins and dehydrins. Sci. Hortic. 2011, 130, 653–659. [Google Scholar] [CrossRef]
- Close, T.J.; Fenton, R.D.; Moonan, F. A view of plant dehydrins using antibodies specific to the carboxy terminal peptide. Plant Mol. Biol. 1993, 23, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.J.; Wang, Y.J.; Zhou, Y.H.; Yuan, T.; Mao, W.H.; Kai, S.; Asami, T.; Chen, Z.X.; Yu, J.Q. Reactive oxygen species are involved in brassinosteroid-induced stress tolerance in cucumber. Plant Physiol. 2009, 150, 801–814. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GABA (µM) | Germination Percentage (%) | Germination Vigor (%) | Germination Index | Mean Germination Time (d) | Seed Vigour Index | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Water | NaCl | Water | NaCl | Water | NaCl | Water | NaCl | Water | NaCl | |
| 0.00 | 95.33 ± 1.15 a | 51.50 ± 1.00 cd* | 90.80 ± 1.10 a | 33.33 ± 2.31 b* | 35.66 ± 1.28 a | 9.29 ± 0.75 c* | 1.61 ± 0.05 a | 3.28 ± 0.29 a* | 1.58 ± 0.13 a | 0.34 ± 0.02 d* |
| 0.25 | 96.00 ± 1.00 a | 54.67 ± 1.15 bc* | 93.20 ± 1.10 a | 40.67 ± 1.15 a* | 35.66 ± 1.47 a | 10.46 ± 0.16 b* | 1.63 ± 0.02 a | 3.10 ± 0.21 a* | 1.59 ± 0.04 a | 0.38 ± 0.01 c* |
| 0.50 | 95.60 ± 1.67 a | 58.00 ± 5.29 ab* | 93.33 ± 1.15 a | 39.33 ± 3.06 a* | 35.86 ± 1.97 a | 11.05 ± 0.88 ab* | 1.85 ± 0.52 a | 3.07 ± 0.14 a* | 1.59 ± 0.09 a | 0.41 ± 0.03 b* |
| 1.00 | 96.00 ± 1.41 a | 63.33 ± 4.16 a* | 93.50 ± 1.00 a | 42.00 ± 2.00 a* | 34.87 ± 3.00 a | 11.73 ± 0.11 a* | 1.75 ± 0.15 a | 3.06 ± 0.12 a* | 1.53 ± 0.19 a | 0.45 ± 0.01 a* |
| 2.50 | 96.00 ± 2.83 a | 50.50 ± 4.43 d* | 92.50 ± 1.91 a | 32.67 ± 1.15 b* | 33.92 ± 1.57 a | 9.15 ± 0.29 c* | 1.75 ± 0.09 a | 3.07 ± 0.09 a* | 1.48 ± 0.09 a | 0.35 ± 0.01 d* |
| 5.00 | 95.60 ± 0.90 a | 46.00 ± 4.90 d* | 92.50 ± 1.91 a | 32.00 ± 3.46 b* | 34.19 ± 2.15 a | 8.77 ± 0.26 c* | 1.68 ± 0.06 a | 3.12 ± 0.19 a* | 1.51 ± 0.11 a | 0.32 ± 0.01 d* |
| GABA (µM) | Seedling Fresh Weight (mg·10 Seedling−1) | Seedling Dry Weight (mg·10 Seedling−1) | Root Length (cm) | Shoot Length (cm) | Shoot-Root Ratio | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Water | NaCl | Water | NaCl | Water | NaCl | Water | NaCl | Water | NaCl | |
| 0.00 | 44.26 ± 2.44 a | 36.25 ± 0.96 c* | 3.36 ± 0.15 a | 3.08 ± 0.2 bc* | 1.03 ± 0.09 a | 0.21 ± 0.02 c* | 0.35 ± 0.03 a | 0.25 ± 0.01 b* | 2.85 ± 0.23 a | 0.85 ± 0.11 b* |
| 0.25 | 45.38 ± 0.88 a | 36.30 ± 0.70 bc* | 3.33 ± 0.21 a | 3.18 ± 0.27 ab | 1.07 ± 0.10 a | 0.24 ± 0.01 bc* | 0.37 ± 0.02 a | 0.27 ± 0.02 b* | 2.80 ± 0.30 a | 0.87 ± 0.06 b* |
| 0.50 | 45.58 ± 3.24 a | 37.33 ± 0.60 ab* | 3.30 ± 0.20 a | 3.13 ± 0.12 ab | 1.10 ± 0.06 a | 0.23 ± 0.01 bc* | 0.36 ± 0.02 a | 0.27 ± 0.01 b* | 2.98 ± 0.08 a | 0.87 ± 0.09 b* |
| 1.00 | 44.82 ± 1.78 a | 37.97 ± 0.87 a* | 3.35 ± 0.10 a | 3.45 ± 0.13 a | 1.06 ± 0.06 a | 0.29 ± 0.04 a* | 0.37 ± 0.02 a | 0.30 ± 0.02 a* | 2.85 ± 0.17 a | 0.98 ± 0.10 a* |
| 2.50 | 42.68 ± 0.81 a | 37.77 ± 0.59 a* | 3.33 ± 0.12 a | 3.15 ± 0.26 ab | 1.00 ± 0.01 a | 0.26 ± 0.04 ab* | 0.37 ± 0.02 a | 0.26 ± 0.02 b* | 2.77 ± 0.12 a | 0.87 ± 0.11 b* |
| 5.00 | 42.80 ± 1.77 a | 36.07 ± 0.58 c* | 3.26 ± 0.06 a | 2.83 ± 0.21 c* | 1.01 ± 0.03 a | 0.21 ± 0.02 c* | 0.37 ± 0.01 a | 0.26 ± 0.01 b* | 2.88 ± 0.30 a | 0.76 ± 0.05 b* |
| Targetgene | Accession No. | Forward Primer (5′–3′) | Reverse Primer (5′–3′) | Tm (°C) |
|---|---|---|---|---|
| Cu/ZnSOD | JQ321597.1 | AACTGTGTACCACGAGGACTTC | AGACTAACAGGTGCTAACAACG | 58 |
| FeSOD | KP202173 | ACACGATTTCTCAGGGTTACGAC | GCGGCCAAGACTATCAGTTCCAT | 58 |
| MnSOD | JQ321598.1 | TAAGGGAACCTACCCGATAACT | CCAGGACCAAACGTCACCAAAG | 66 |
| CAT | JQ321596.1 | AACAGGACGGGAATAGCACG | ACCAGGTTCAGACACGGAGACA | 58 |
| GPOX | JQ321606.1 | CACTTGGTTTAGTTTTGTCGCC | AACACGGTCTTGTCTGCTACG | 64 |
| APX | JQ321599.1 | TAAAGATAGTCAACCCACCTCAACA | ACCAGTCTTGGGAAACAACGTA | 58 |
| MDHR | KP202172 | CCAACTGCCTAAAGCCACATCT | GAAGAAAGGAAACTAACGGAGCAT | 64 |
| DHAR | KP202171 | TGGTTACCTCCCGACCCTAT | TCTTACCAAGGAACTTTAGTCAGG | 58 |
| GPX | JQ321604.1 | ATGTGCCTTGAGAGCGTGAATATAC | CCTTTAAGACGAAACTTGGACC | 58 |
| CytGR | JQ321602.1 | TAAACTTCCACTCCCTTTCTATCG | CTACAATATGGGTTGAGGACAGGT | 58 |
| GST | JQ321600.1 | TCGAGCTAAGACCCGACTAATAC | GAGGTTGTTAAACTACCGAAGATAC | 58 |
| SK2 | GU443960.1 | TGGAACAGGAGTAACAACAGGTGGA | TGCCAGTTGAGAAAGTTGAGGTTGT | 58 |
| Y2K | JF748410.1 | AGCCACGCAACAAGGTTCTAA | TTGAGGATACGGGATGGGTG | 60 |
| Y2SK | GU443965.1 | GTGCGATGGAGATGCTGTTTG | CCTAATCCAACTTCAGGTTCAGC | 60 |
| dehydrin b | GU443960.1 | TCCAGTCATCCAGCCTGTTG | CCAGCCACAACACTTGTCA | 60 |
| VP1 | MF405364 | GTCCAATCAGTGACAATGCCG | AGAGGGCAAGAGACACAAGAGC | 58 |
| HKT1 | MF405365 | TGCATCACCGAAAGACAAAGC | ATCGACAACCCTACATTCCCATA | 57 |
| HKT8 | MF405366 | TTCAAGACACGCTGGAGAAACTAT | CGATGGCAGGAATGAGGTGT | 57 |
| SKOR | MF405367 | GTTTCATTTGATATGGTTCTCGGTG | GGCCCTTTATTTGTTCACGGA | 58 |
| HAL2 | MF405368 | TTGTGAACCAGTTGAGAAGGCC | TCGGCATCTCCACGACCTATT | 61 |
| H+̶ATPase | MF405369 | CGTATAGTGTTTGGCTTCATGTTCA | AATGGAGATGGCACCACCCTA | 60 |
| SOS1 | MF405370 | TGGTCCATCTGAAAGTGACAATAAC | TCATCAAGCATCTCCCAGTAAGC | 57 |
| NHX6 | MF405371 | CAGTCTGGTTTCAGTCTTGCTCC | ACCAAACATCAGGCACTCAACA | 60 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, B.; Li, Z.; Liang, L.; Cao, Y.; Zeng, W.; Zhang, X.; Ma, X.; Huang, L.; Nie, G.; Liu, W.; et al. The γ-Aminobutyric Acid (GABA) Alleviates Salt Stress Damage during Seeds Germination of White Clover Associated with Na+/K+ Transportation, Dehydrins Accumulation, and Stress-Related Genes Expression in White Clover. Int. J. Mol. Sci. 2018, 19, 2520. https://doi.org/10.3390/ijms19092520
Cheng B, Li Z, Liang L, Cao Y, Zeng W, Zhang X, Ma X, Huang L, Nie G, Liu W, et al. The γ-Aminobutyric Acid (GABA) Alleviates Salt Stress Damage during Seeds Germination of White Clover Associated with Na+/K+ Transportation, Dehydrins Accumulation, and Stress-Related Genes Expression in White Clover. International Journal of Molecular Sciences. 2018; 19(9):2520. https://doi.org/10.3390/ijms19092520
Chicago/Turabian StyleCheng, Bizhen, Zhou Li, Linlin Liang, Yiqin Cao, Weihang Zeng, Xinquan Zhang, Xiao Ma, Linkai Huang, Gang Nie, Wei Liu, and et al. 2018. "The γ-Aminobutyric Acid (GABA) Alleviates Salt Stress Damage during Seeds Germination of White Clover Associated with Na+/K+ Transportation, Dehydrins Accumulation, and Stress-Related Genes Expression in White Clover" International Journal of Molecular Sciences 19, no. 9: 2520. https://doi.org/10.3390/ijms19092520
APA StyleCheng, B., Li, Z., Liang, L., Cao, Y., Zeng, W., Zhang, X., Ma, X., Huang, L., Nie, G., Liu, W., & Peng, Y. (2018). The γ-Aminobutyric Acid (GABA) Alleviates Salt Stress Damage during Seeds Germination of White Clover Associated with Na+/K+ Transportation, Dehydrins Accumulation, and Stress-Related Genes Expression in White Clover. International Journal of Molecular Sciences, 19(9), 2520. https://doi.org/10.3390/ijms19092520