Signalling Alterations in Bones of Pituitary Adenylate Cyclase Activating Polypeptide (PACAP) Gene Deficient Mice




Abstract
:1. Introduction
2. Results
2.1. Bone Morphology Is Only Slightly Modified in PACAP Gene Deficient Mice
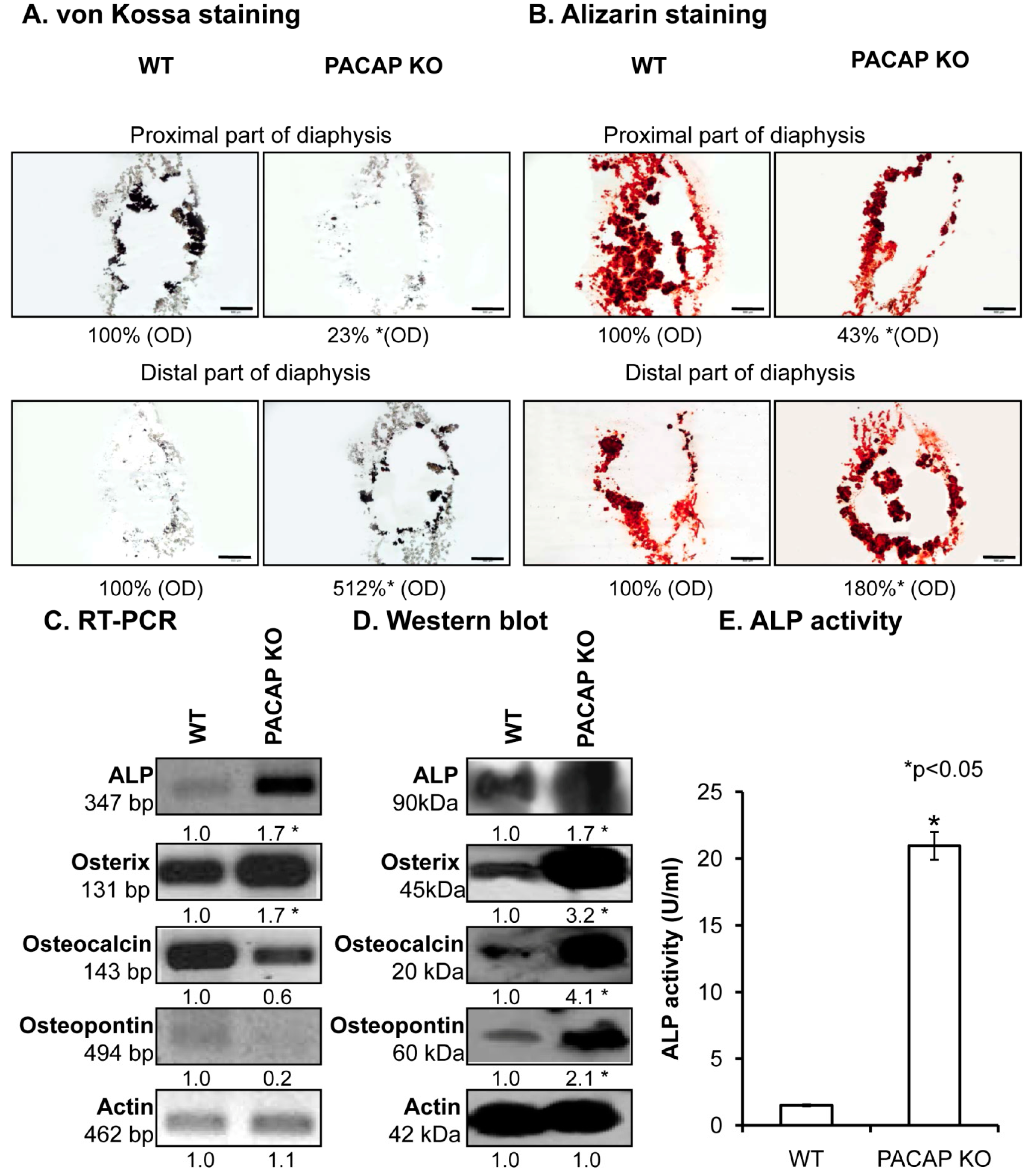
2.2. Inorganic Matrix Production Is Disturbed in PACAP KO Mice
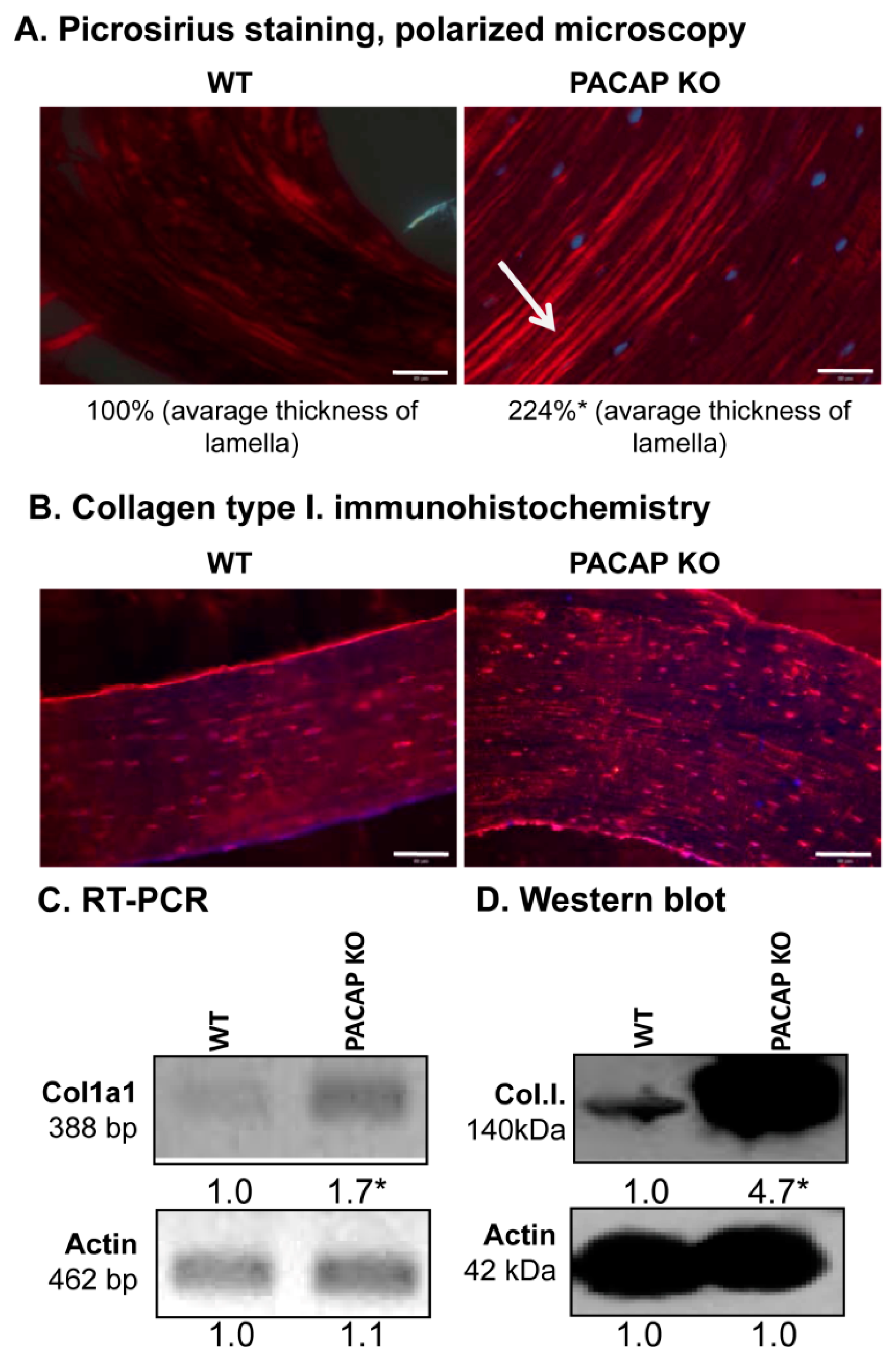
2.3. Augmented Collagen Type I Expression in PACAP KO Mice
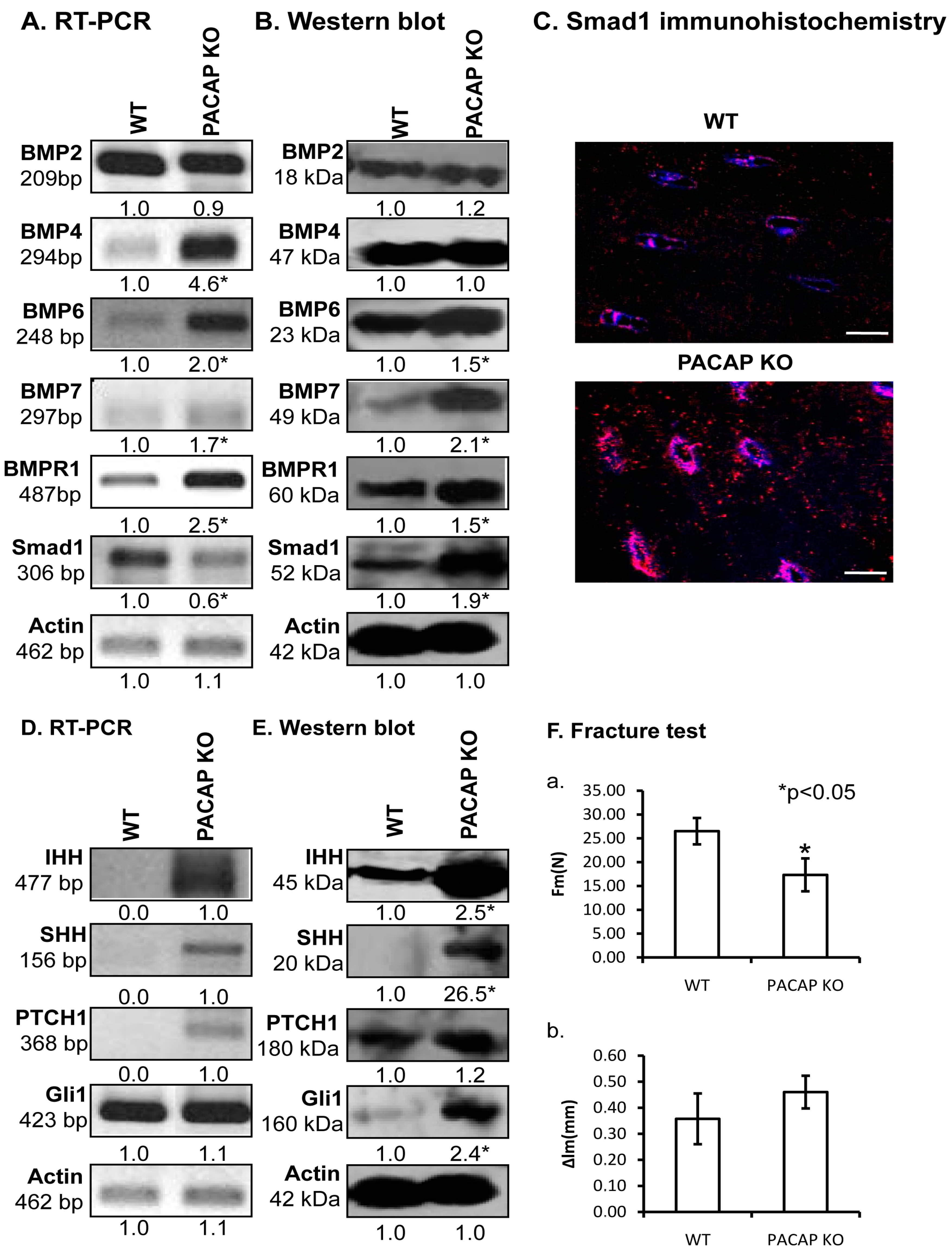
2.4. Alterations of PACAP Signalling Pathway in Cortical Bone
2.5. Alterations of Possible Signalling Crosstalks
2.6. Biomechanical Properties of Bones Are Altered in PACAP KO Mice
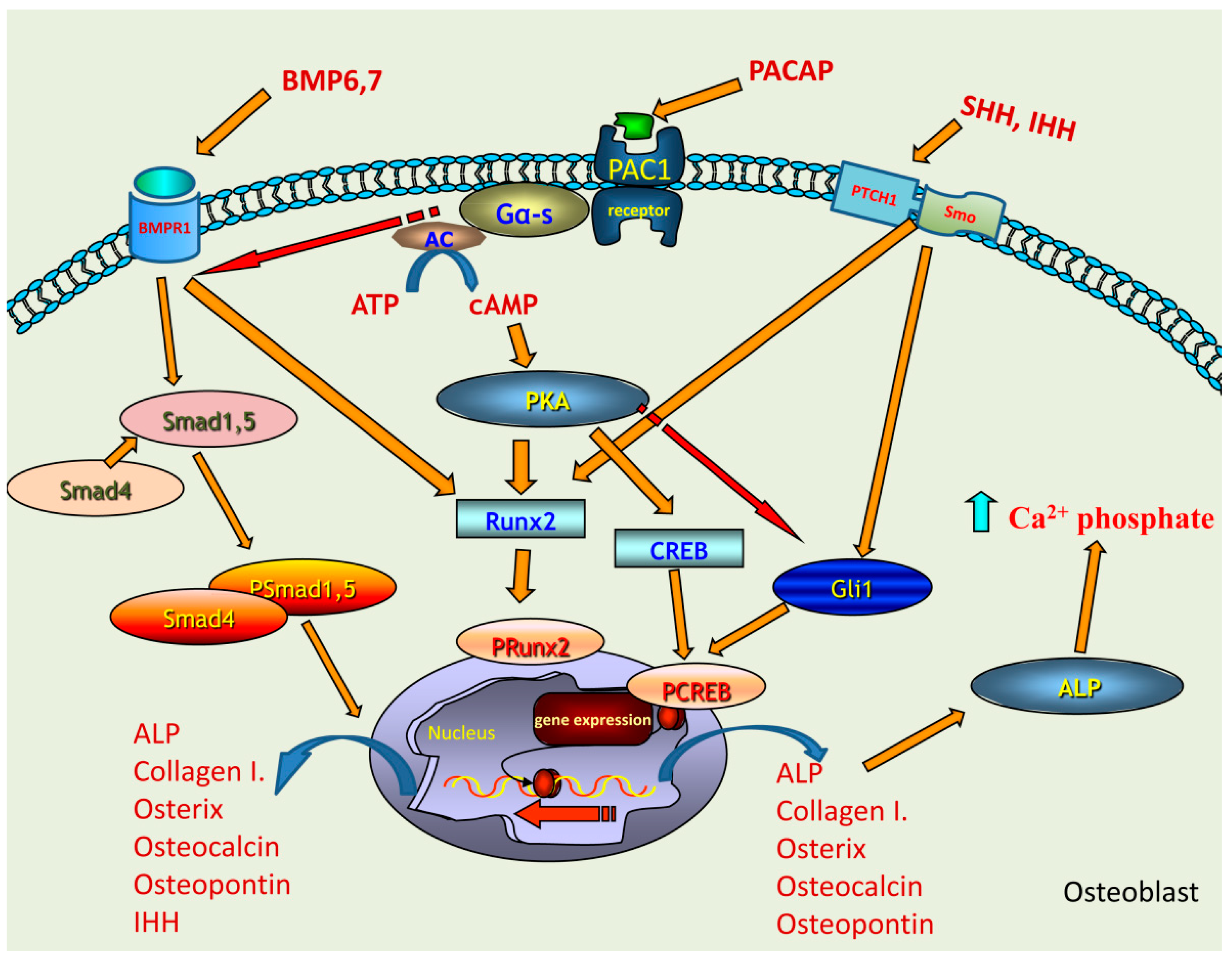
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Whole Limb Alizarin Staining
4.3. Staining Procedure of Calcificated Tissue
4.4. Staining Procedure of Decalcified Samples
4.5. Radiographic Analysis (Micro-CT Scan)
4.6. Measurement of Alkaline Phosphatase Activity
4.7. Immunohistochemistry
4.8. RT-PCR Analysis
4.9. Western Blot Analysis
4.10. Fracture Test
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ALP | Alkaline phosphatase |
| BMP | Bone morphogenetic protein |
| cAMP | Cyclic adenosine monophosphate |
| CNS | Central nervous system |
| CREB | cAMP response element-binding protein |
| DMEM | Dulbecco’s Modified Eagle’s Medium |
| dNTP | deoxynucleotide triphosphate |
| ECM | Extracellular matrix |
| EDTA | Ethylene diamine tetra-acetic acid |
| EGTA | Ethylene glycol-bis(β-aminoethyl ether)-N,N,N′,N′-tetra acetic acid |
| FBS | Foetal bovine serum |
| FGF | Fibroblast growth factor; Gli, glioma-associated oncogene |
| HH | hedgehog |
| HEPES | 4-(2-hydroxyethyl)-1-piperazine ethanesulfonic acid |
| hMSC | human mesenchymal stem cell |
| IHH | Indian Hedgehog |
| MAPK | Mitogen-activated protein kinase |
| PAC1 | Pituitary adenylate cyclase-activating polypeptide type I receptor |
| PACAP | Pituitary adenylate cyclase activating polypeptide, phosphate buffered saline |
| PBST | Phosphate buffered saline supplemented with 1% Tween-20 |
| PLC | Phospholipase C |
| PKA | Protein kinase A |
| PKC | Protein kinase C |
| PTCH | Patched |
| RT-PCR | Reverse transcription followed by polymerase chain reaction Runt-related transcription factor 2 |
| SHH | Sonic Hedgehog |
| TGFβ | Transforming growth factor-β |
| VIP | Vasoactive intestinal polypeptide |
| VPAC | Vasoactive intestinal polypeptide receptor |
| WNT | Wingless int1 |
References
- Majidinia, M.; Sadeghpour, A.; Yousefi, B. The roles of signaling pathways in bone repair and regeneration. J. Cell. Physiol. 2017, 233, 2937–2948. [Google Scholar] [CrossRef] [PubMed]
- Oftadeh, R.; Perez-Viloria, M.; Villa-Camacho, J.C.; Vaziri, A.; Nazarian, A. Biomechanics and mechanobiology of trabecular bone: A review. J. Biomech. Eng. 2015, 137, 010802. [Google Scholar] [CrossRef] [PubMed]
- Hegde, V.; Jo, J.E.; Andreopoulou, P.; Lane, J.M. Effect of osteoporosis medications on fracture healing. Osteoporos. Int. 2016, 27, 861–871. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Chen, G.; Li, Y.P. TGF-beta and BMP signaling in osteoblast, skeletal development, and bone formation, homeostasis and disease. Bone Res. 2016, 4, 16009. [Google Scholar] [CrossRef] [PubMed]
- Forlino, A.; Marini, J.C. Osteogenesis imperfecta. Lancet 2016, 387, 1657–1671. [Google Scholar] [CrossRef]
- Ascenzi, M.G.; Roe, A.K. The osteon: The micromechanical unit of compact bone. Front. Biosci. 2012, 17, 1551–1581. [Google Scholar] [CrossRef]
- Shahi, M.; Peymani, A.; Sahmani, M. Regulation of Bone Metabolism. Rep. Biochem. Mol. Biol. 2017, 5, 73–82. [Google Scholar] [PubMed]
- Zhou, W.; Yu, L.; Fan, J.; Wan, B.; Jiang, T.; Yin, J.; Huang, Y.; Li, Q.; Yin, G.; Hu, Z. Endogenous Parathyroid Hormone Promotes Fracture Healing by Increasing Expression of BMPR2 through cAMP/PKA/CREB Pathway in Mice. Cell. Physiol. Biochem. 2017, 42, 551–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Wang, W.; Xu, H.; Ning, Y.; Fang, W.; Liao, W.; Zou, J.; Yang, Y.; Shao, N. Effects of altered CXCL12/CXCR4 axis on BMP2/Smad/Runx2/Osterix axis and osteogenic gene expressions during osteogenic differentiation of MSCs. Am. J. Transl. Res. 2017, 9, 1680–1693. [Google Scholar] [PubMed]
- Vaudry, D.; Falluel-Morel, A.; Bourgault, S.; Basille, M.; Burel, D.; Wurtz, O.; Fournier, A.; Chow, B.K.; Hashimoto, H.; Galas, L.; et al. Pituitary adenylate cyclase-activating polypeptide and its receptors: 20 years after the discovery. Pharmacol. Rev. 2009, 61, 283–357. [Google Scholar] [CrossRef] [PubMed]
- Miyata, A.; Arimura, A.; Dahl, R.R.; Minamino, N.; Uehara, A.; Jiang, L.; Culler, M.D.; Coy, D.H. Isolation of a novel 38 residue-hypothalamic polypeptide which stimulates adenylate cyclase in pituitary cells. Biochem. Biophys. Res. Commun. 1989, 164, 567–574. [Google Scholar] [CrossRef]
- Barberi, M.; Di Paolo, V.; Latini, S.; Guglielmo, M.C.; Cecconi, S.; Canipari, R. Expression and functional activity of PACAP and its receptors on cumulus cells: Effects on oocyte maturation. Mol. Cell. Endocrinol. 2013, 375, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Heimesaat, M.M.; Dunay, I.R.; Schulze, S.; Fischer, A.; Grundmann, U.; Alutis, M.; Kuhl, A.A.; Tamas, A.; Toth, G.; Dunay, M.P.; et al. Pituitary adenylate cyclase-activating polypeptide ameliorates experimental acute ileitis and extra-intestinal sequelae. PLoS ONE 2014, 9, e108389. [Google Scholar] [CrossRef] [PubMed]
- Banki, E.; Kovacs, K.; Nagy, D.; Juhasz, T.; Degrell, P.; Csanaky, K.; Kiss, P.; Jancso, G.; Toth, G.; Tamas, A.; et al. Molecular mechanisms underlying the Nephroprotective effects of PACAP in diabetes. J. Mol. Neurosci. 2014, 54, 300–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Csanaky, K.; Banki, E.; Szabadfi, K.; Reglodi, D.; Tarcai, I.; Czegledi, L.; Helyes, Z.; Ertl, T.; Gyarmati, J.; Szanto, Z.; et al. Changes in PACAP immunoreactivity in human milk and presence of PAC1 receptor in mammary gland during lactation. J. Mol. Neurosci. 2012, 48, 631–637. [Google Scholar] [CrossRef] [PubMed]
- Juhasz, T.; Matta, C.; Katona, E.; Somogyi, C.; Takacs, R.; Gergely, P.; Csernoch, L.; Panyi, G.; Toth, G.; Reglodi, D.; et al. Pituitary adenylate cyclase activating polypeptide (PACAP) signalling exerts chondrogenesis promoting and protecting effects: Implication of calcineurin as a downstream target. PLoS ONE 2014, 9, e91541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mester, L.; Kovacs, K.; Racz, B.; Solti, I.; Atlasz, T.; Szabadfi, K.; Tamas, A.; Reglodi, D. Pituitary adenylate cyclase-activating polypeptide is protective against oxidative stress in human retinal pigment epithelial cells. J. Mol. Neurosci. 2011, 43, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Laszlo, E.; Varga, A.; Kovacs, K.; Jancso, G.; Kiss, P.; Tamas, A.; Szakaly, P.; Fulop, B.; Reglodi, D. Ischemia/reperfusion-induced Kidney Injury in Heterozygous PACAP-deficient Mice. Transplant. Proc. 2015, 47, 2210–2215. [Google Scholar] [CrossRef] [PubMed]
- Gomariz, R.P.; Juarranz, Y.; Abad, C.; Arranz, A.; Leceta, J.; Martinez, C. VIP-PACAP system in immunity: New insights for multitarget therapy. Ann. N. Y. Acad. Sci. 2006, 1070, 51–74. [Google Scholar] [CrossRef] [PubMed]
- Tsuchida, M.; Nakamachi, T.; Sugiyama, K.; Tsuchikawa, D.; Watanabe, J.; Hori, M.; Yoshikawa, A.; Imai, N.; Kagami, N.; Matkovits, A.; et al. PACAP stimulates functional recovery after spinal cord injury through axonal regeneration. J. Mol. Neurosci. 2014, 54, 380–387. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Nakamachi, T.; Endo, K.; Ito, K.; Machida, T.; Oka, T.; Hori, M.; Ishizaka, K.; Shioda, S. Distribution of pituitary adenylate cyclase-activating polypeptide (PACAP) in the human testis and in testicular germ cell tumors. Andrologia 2014, 46, 465–471. [Google Scholar] [CrossRef] [PubMed]
- Juhasz, T.; Helgadottir, S.L.; Tamas, A.; Reglodi, D.; Zakany, R. PACAP and VIP signaling in chondrogenesis and osteogenesis. Peptides 2015, 66, 51–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lundberg, P.; Lundgren, I.; Mukohyama, H.; Lehenkari, P.P.; Horton, M.A.; Lerner, U.H. Vasoactive intestinal peptide (VIP)/pituitary adenylate cyclase-activating peptide receptor subtypes in mouse calvarial osteoblasts: Presence of VIP-2 receptors and differentiation-induced expression of VIP-1 receptors. Endocrinology 2001, 142, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Juhasz, T.; Matta, C.; Katona, E.; Somogyi, C.; Takacs, R.; Hajdu, T.; Helgadottir, S.L.; Fodor, J.; Csernoch, L.; Toth, G.; et al. Pituitary adenylate cyclase-activating polypeptide (PACAP) signalling enhances osteogenesis in UMR-106 cell line. J. Mol. Neurosci. 2014, 54, 555–573. [Google Scholar] [CrossRef] [PubMed]
- Nagata, A.; Tanaka, T.; Minezawa, A.; Poyurovsky, M.; Mayama, T.; Suzuki, S.; Hashimoto, N.; Yoshida, T.; Suyama, K.; Miyata, A.; et al. cAMP activation by PACAP/VIP stimulates IL-6 release and inhibits osteoblastic differentiation through VPAC2 receptor in osteoblastic MC3T3 cells. J. Cell. Physiol. 2009, 221, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Strange-Vognsen, H.H.; Arnbjerg, J.; Hannibal, J. Immunocytochemical demonstration of pituitary adenylate cyclase activating polypeptide (PACAP) in the porcine epiphyseal cartilage canals. Neuropeptides 1997, 31, 137–141. [Google Scholar] [CrossRef]
- Juhasz, T.; Szentleleky, E.; Somogyi, C.S.; Takacs, R.; Dobrosi, N.; Engler, M.; Tamas, A.; Reglodi, D.; Zakany, R. Pituitary Adenylate Cyclase Activating Polypeptide (PACAP) Pathway Is Induced by Mechanical Load and Reduces the Activity of Hedgehog Signaling in Chondrogenic Micromass Cell Cultures. Int. J. Mol. Sci. 2015, 16, 17344–17367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- May, V.; Clason, T.A.; Buttolph, T.R.; Girard, B.M.; Parsons, R.L. Calcium influx, but not intracellular calcium release, supports PACAP-mediated ERK activation in HEK PAC1 receptor cells. J. Mol. Neurosci. 2014, 54, 342–350. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, J.; Nakamachi, T.; Matsuno, R.; Hayashi, D.; Nakamura, M.; Kikuyama, S.; Nakajo, S.; Shioda, S. Localization, characterization and function of pituitary adenylate cyclase-activating polypeptide during brain development. Peptides 2007, 28, 1713–1719. [Google Scholar] [CrossRef] [PubMed]
- Toriyama, M.; Mizuno, N.; Fukami, T.; Iguchi, T.; Toriyama, M.; Tago, K.; Itoh, H. Phosphorylation of doublecortin by protein kinase A orchestrates microtubule and actin dynamics to promote neuronal progenitor cell migration. J. Biol. Chem. 2012, 287, 12691–12702. [Google Scholar] [CrossRef] [PubMed]
- Farkas, J.; Sandor, B.; Tamas, A.; Kiss, P.; Hashimoto, H.; Nagy, A.D.; Fulop, B.D.; Juhasz, T.; Manavalan, S.; Reglodi, D. Early Neurobehavioral Development of Mice Lacking Endogenous PACAP. J. Mol. Neurosci. 2017, 61, 468–478. [Google Scholar] [CrossRef] [PubMed]
- Baumgartner, R.; Heeren, N.; Quast, D.; Babst, R.; Brunner, A. Is the cortical thickness index a valid parameter to assess bone mineral density in geriatric patients with hip fractures? Arch. Orthop. Trauma Surg. 2015, 135, 805–810. [Google Scholar] [CrossRef] [PubMed]
- Alam, I.; Alkhouli, M.; Gerard-O’Riley, R.L.; Wright, W.B.; Acton, D.; Gray, A.K.; Patel, B.; Reilly, A.M.; Lim, K.E.; Robling, A.G.; et al. Osteoblast-Specific Overexpression of Human WNT16 Increases Both Cortical and Trabecular Bone Mass and Structure in Mice. Endocrinology 2016, 157, 722–736. [Google Scholar] [CrossRef] [PubMed]
- Sinha, P.; Aarnisalo, P.; Chubb, R.; Poulton, I.J.; Guo, J.; Nachtrab, G.; Kimura, T.; Swami, S.; Saeed, H.; Chen, M.; et al. Loss of Gsalpha in the Postnatal Skeleton Leads to Low Bone Mass and a Blunted Response to Anabolic Parathyroid Hormone Therapy. J. Biol. Chem. 2016, 291, 1631–1642. [Google Scholar] [CrossRef] [PubMed]
- Zakany, R.; Bako, E.; Felszeghy, S.; Hollo, K.; Balazs, M.; Bardos, H.; Gergely, P.; Modis, L. Okadaic acid-induced inhibition of protein phosphatase 2A enhances chondrogenesis in chicken limb bud micromass cell cultures. Anat. Embryol. 2001, 203, 23–34. [Google Scholar] [PubMed]
- Zakany, R.; Szucs, K.; Bako, E.; Felszeghy, S.; Czifra, G.; Biro, T.; Modis, L.; Gergely, P. Protein phosphatase 2A is involved in the regulation of protein kinase A signaling pathway during in vitro chondrogenesis. Exp. Cell Res. 2002, 275, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Li, L.; Dong, Q.; Wang, Y.; Feng, Q.; Ou, X.; Zhou, P.; He, T.; Luo, J. Activation of PKA/CREB Signaling is Involved in BMP9-Induced Osteogenic Differentiation of Mesenchymal Stem Cells. Cell. Physiol. Biochem. 2015, 37, 548–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laurie, L.E.; Kokubo, H.; Nakamura, M.; Saga, Y.; Funato, N. The Transcription Factor Hand1 Is Involved in Runx2-Ihh-Regulated Endochondral Ossification. PLoS ONE 2016, 11, e0150263. [Google Scholar] [CrossRef] [PubMed]
- Poole, K.E.; Treece, G.M.; Mayhew, P.M.; Vaculik, J.; Dungl, P.; Horak, M.; Stepan, J.J.; Gee, A.H. Cortical thickness mapping to identify focal osteoporosis in patients with hip fracture. PLoS ONE 2012, 7, e38466. [Google Scholar] [CrossRef] [PubMed]
- Sandor, B.; Fintor, K.; Felszeghy, S.; Juhasz, T.; Reglodi, D.; Mark, L.; Kiss, P.; Jungling, A.; Fulop, B.D.; Nagy, A.D.; et al. Structural and morphometric comparison of the molar teeth in pre-eruptive developmental stage of PACAP-deficient and wild-type mice. J. Mol. Neurosci. 2014, 54, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Sharma, U.; Pal, D.; Prasad, R. Alkaline phosphatase: An overview. IndianJ. Clin. Biochem. 2014, 29, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Schaffler, M.B.; Choi, K.; Milgrom, C. Aging and matrix microdamage accumulation in human compact bone. Bone 1995, 17, 521–525. [Google Scholar] [CrossRef]
- Hu, L.; Su, P.; Yin, C.; Zhang, Y.; Li, R.; Yan, K.; Chen, Z.; Li, D.; Zhang, G.; Wang, L.; et al. Microtubule actin crosslinking factor 1 promotes osteoblast differentiation by promoting beta-catenin/TCF1/Runx2 signaling axis. J. Cell. Physiol. 2017, 233, 1574–1584. [Google Scholar] [CrossRef] [PubMed]
- Shen, B.; Mu, J.X.; Pei, F.X. Relationship among bone mineral density, collagen composition, and biomechanical properties of callus in the healing of osteoporotic fracture. Chin. J. Traumatol. 2007, 10, 360–365. [Google Scholar] [PubMed]
- Caetano-Lopes, J.; Nery, A.M.; Canhao, H.; Duarte, J.; Cascao, R.; Rodrigues, A.; Perpetuo, I.P.; Abdulghani, S.; Amaral, P.M.; Sakaguchi, S.; et al. Chronic arthritis leads to disturbances in the bone collagen network. Arthritis Res. Ther. 2010, 12, R9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tou, L.; Quibria, N.; Alexander, J.M. Transcriptional regulation of the human Runx2/Cbfa1 gene promoter by bone morphogenetic protein-7. Mol. Cell. Endocrinol. 2003, 205, 121–129. [Google Scholar] [CrossRef]
- Li, J.; Hao, L.; Wu, J.; Zhang, J.; Su, J. Linarin promotes osteogenic differentiation by activating the BMP-2/RUNX2 pathway via protein kinase A signaling. Int. J. Mol. Med. 2016, 37, 901–910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bais, M.V.; Wigner, N.; Young, M.; Toholka, R.; Graves, D.T.; Morgan, E.F.; Gerstenfeld, L.C.; Einhorn, T.A. BMP2 is essential for post natal osteogenesis but not for recruitment of osteogenic stem cells. Bone 2009, 45, 254–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadjicharalambous, C.; Kozlova, D.; Sokolova, V.; Epple, M.; Chatzinikolaidou, M. Calcium phosphate nanoparticles carrying BMP-7 plasmid DNA induce an osteogenic response in MC3T3-E1 pre-osteoblasts. J. Biomed. Mater. Res. 2015, 103, 3834–3842. [Google Scholar] [CrossRef] [PubMed]
- Friedman, M.S.; Long, M.W.; Hankenson, K.D. Osteogenic differentiation of human mesenchymal stem cells is regulated by bone morphogenetic protein-6. J. Cell. Biochem. 2006, 98, 538–554. [Google Scholar] [CrossRef] [PubMed]
- Nam, J.; Perera, P.; Rath, B.; Agarwal, S. Dynamic regulation of bone morphogenetic proteins in engineered osteochondral constructs by biomechanical stimulation. Tissue Eng. 2013, 19, 783–792. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Du, J.; Yin, X.; Shan, Z.; Ma, Y.; Ma, P.; Du, J.; Fan, Z. Shh signaling, negatively regulated by BMP signaling, inhibits the osteo/dentinogenic differentiation potentials of mesenchymal stem cells from apical papilla. Mol. Cell. Biochem. 2013, 383, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Niewiadomski, P.; Zhujiang, A.; Youssef, M.; Waschek, J.A. Interaction of PACAP with Sonic hedgehog reveals complex regulation of the hedgehog pathway by PKA. Cell. Signal. 2013, 25, 2222–2230. [Google Scholar] [CrossRef] [PubMed]
- Seki, K.; Hata, A. Indian hedgehog gene is a target of the bone morphogenetic protein signaling pathway. J. Biol. Chem. 2004, 279, 18544–18549. [Google Scholar] [CrossRef] [PubMed]
- Duench, K.; Franz-Odendaal, T.A. BMP and Hedgehog signaling during the development of scleral ossicles. Dev. Biol. 2012, 365, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, H.; Shintani, N.; Tanaka, K.; Mori, W.; Hirose, M.; Matsuda, T.; Sakaue, M.; Miyazaki, J.; Niwa, H.; Tashiro, F.; et al. Altered psychomotor behaviors in mice lacking pituitary adenylate cyclase-activating polypeptide (PACAP). Proc. Natl. Acad. Sci. USA 2001, 98, 13355–13360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashimoto, H.; Hashimoto, R.; Shintani, N.; Tanaka, K.; Yamamoto, A.; Hatanaka, M.; Guo, X.; Morita, Y.; Tanida, M.; Nagai, K.; et al. Depression-like behavior in the forced swimming test in PACAP-deficient mice: Amelioration by the atypical antipsychotic risperidone. J. Neurochem. 2009, 110, 595–602. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Nucleotide Sequence (5′→3′) | GenBank ID | Annealing Temperature | Amplimer Size (bp) |
|---|---|---|---|---|---|
| Alkaline phosphatase (Alpl) | sense | GAA GTC CGT GGG CAT CGT (474–491) | NM013059 | 59 °C | 347 |
| antisense | CAG TGC GGT TCC AGA CAT AG (801–820) | ||||
| BMP2 (Bmp2) | sense | AAG CCA GGT GTC TCC AAG (697–714) | NM017178.1 | 53 °C | 209 |
| antisense | AAG TCC ACA TAC AAA GGG TG (886–905) | ||||
| BMP4 (Bmp4) | sense | TAG TCC CAA GCA TCA CCC (876–893) | NM012827.2 | 53 °C | 294 |
| antisense | TCG TAC TCG TCC AGA TAC AAC (1149–1169) | ||||
| BMP6 (Bmp6) | sense | CCC AGA TTC CTG AGG GTG A (936–954) | NM013107.1 | 56 °C | 248 |
| antisense | CAT GTT GTG CTG CGG TGT (1166–1183) | ||||
| BMP7 (Bmp7) | sense | AGG GAG TCC GAC CTC TTC T (607–625) | NM001191856.1 | 54 °C | 297 |
| antisense | GTT CTG GCT GCG TTG TTT (886–903) | ||||
| BMPR1 (Bmpr1a) | sense | CCA TTG CTT TGC CAT TAT (240–257) | NM009758.4 | 47 °C | 487 |
| antisense | TTT ACC AAC CTG CCG AAC (709–726) | ||||
| Collagen type I (Col1a1) | sense | GGG CGA GTG CTG TGC TTT (348–365) | NM007742.3 | 60 °C | 388 |
| antisense | GGG ACC CAT TGG ACC TGA A (717–735) | ||||
| CREB (Creb1) | sense | AGA TTG CCA CAT TAG CCC (95–112) | NM031017.1 | 52 °C | 441 |
| antisense | GCT GTA TTG CTC CTC CCT (518–535) | ||||
| Actin (Actb) | sense | GCC AAC CGT GAA AAG ATG A (419–437) | NM007393.5 | 54 °C | 462 |
| antisense | CAA GAA GGA AGG CTG GAA AA (861–880) | ||||
| Gli1 (Gli1) | sense | CCA CCC TAC CTC TGT CTA TTC G (2201–2222) | NM010296.2 | 49 °C | 423 |
| antisense | CAC CCT TGT TCT GGT TTT ACC (2603–2623) | ||||
| IHH (Ihh) | sense | CCA ACT ACA ATC CCG ACA TCA (248–268) | NM053384.1 | 58 °C | 477 |
| antisense | GTC TTC ATC CCA GCC TTC C (390–408) | ||||
| Osterix (Sp7) | sense | GCC TAC TTA CCC GTC TGA CTT T (525–543) | NM001037632.1 | 56 °C | 131 |
| antisense | GCC CAC TAT TGC CAA CTG C (634–652) | ||||
| Osteocalcin (Bglap2) | sense | TAA GGT GGT GAA TAG ACT CCG (123–143) | NM013414.1 | 56 °C | 143 |
| antisense | CCT GGA AGC CAA TGT GGT (248–265) | ||||
| PAC1 (ADCYAP1R1) | sense | CTA CGC CCT TTA CTA CCC AG (210–229) | NM016989.2 | 49 °C | 247 |
| antisense | GTA TTT CTT GAC AGC CAT TTG T (435–456) | ||||
| PKA (Prkaca) | sense | GCA AAG GCT ACA ACA AGG C (847–865) | NM008854 | 53 °C | 280 |
| antisense | ATG GCA ATC CAG TCA ATC G (1109–1126) | ||||
| Osteopontin (Spp1) | sense | GCT GAA GCC TGA CCC ATC T (126–144) | X51834 | 59 °C | 494 |
| antisense | TCC CGT TGC TGT CCT GAT (602–619) | ||||
| PTCH1 (Ptch1) | sense | GGA ACT TAT CAC GGA GAC AG (579–578) | NM053566.1 | 56 °C | 368 |
| antisense | AAC CTT GAC ATC CAC CAT T (928–946) | ||||
| Runx2 (Runx2) | sense | GGA CGA GGC AAG AGT TTC A (598–616) | NM001278483.1 | 55 °C | 249 |
| antisense | TGG TGC AGA GTT CAG GGA G (828–846) | ||||
| SHH (Shh) | sense | TCG TGC TAC GCA GTC ATC G (1042–1060) | NM017221.1 | 56 °C | 156 |
| antisense | CCT CGC TTC CGC TAC AGA (1180–1197) | ||||
| Smad1 (Smad1) | sense | AGC ACC TAC CCT CAC TCC C (935–953) | NM013130.2 | 56 °C | 306 |
| antisense | GAA ACC ATC CAC CAA CAC G (1222–1240) | ||||
| VPAC1 (VIPR1) | sense | GTT CTA TGG CAC GGT CAA (376–393) | NM001097523 | 52 °C | 216 |
| antisense | AGC AAT GTT CGG GTT CTC (573–590) | ||||
| VPAC2 (VIPR2) | sense | TCG GAA CTA CAT CCA TCT (477–497) | NM001014970 | 48 °C | 177 |
| antisense | TTT GCC ATA ACA CCA TAC (636–653) |
| Antibody | Host Animal | Dilution | Distributor |
|---|---|---|---|
| Anti-PAC1 | rabbit, polyclonal, | 1:600 | Sigma-Aldrich, St. Louis, MO, USA |
| Anti-VPAC1 | rabbit, polyclonal, | 1:800 | Alomone Labs., Jerusalem, Israel |
| Anti-VPAC2 | rabbit, polyclonal, | 1:600 | Abcam, Camridge, UK |
| Anti-Coll. I. | mouse, monoclonal, | 1:1000 | Sigma-Aldrich, St. Louis, MO, USA |
| Anti-CREB | rabbit, polyclonal, | 1:800 | Millipore, Billerica, MA, USA |
| Anti-P-CREB | rabbit, polyclonal, | 1:800 | Millipore, Billerica, MA, USA |
| Anti-Osterix | rabbit, polyclonal, | 1:200 | SantaCruz Biotechnology Inc., Santa Cruz, CA, USA |
| Anti-Osteocalcin | rabbit, polyclonal, | 1:600 | Abcam, Camridge, UK |
| Anti-Osteopontin | rabbit, polyclonal, | 1:500 | Abcam, Camridge, UK |
| Anti-ALP | rabbit, polyclonal, | 1:500 | Abcam, Camridge, UK |
| Anti-SHH | rabbit, polyclonal, | 1:600 | Cell Signaling, Danvers, MA, USA |
| Anti-IHH | rabbit, polyclonal, | 1:600 | Millipore, Billerica, MA, USA |
| Anti-PKA | rabbit, polyclonal, | 1:800 | Cell Signaling, Danvers, MA, USA |
| Anti-Runx2 | rabbit, polyclonal, | 1:500 | Cell Signaling, Danvers, MA, USA |
| Anti-Gli1 | rabbit, polyclonal, | 1:600 | Cell Signaling, Danvers, MA, USA |
| Anti-BMP2 | mouse, monoclonal, | 1:500 | Abcam, Camridge, UK |
| Anti-BMP4 | rabbit, polyclonal, | 1:600 | Cell Signaling, Danvers, MA, USA |
| Anti-BMP6 | rabbit, polyclonal, | 1:200 | SantaCruz Biotechnology Inc., Santa Cruz, CA, USA |
| Anti-BMP7 | rabbit, polyclonal, | 1:600 | Abcam, Camridge, UK |
| Anti-BMPR1 | mouse, monoclonal, | 1:600 | Abcam, Camridge, UK |
| Anti-Smad1 | rabbit, polyclonal, | 1:600 | Cell Signaling, Danvers, MA, USA |
| Anti-PTCH1 | rabbit, polyclonal, | 1:800 | Cell Signaling, Danvers, MA, USA |
| Anti-Actin | mouse, monoclonal, | 1:10,000 | Sigma-Aldrich, St. Louis, MO, USA |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Józsa, G.; Szegeczki, V.; Pálfi, A.; Kiss, T.; Helyes, Z.; Fülöp, B.; Cserháti, C.; Daróczi, L.; Tamás, A.; Zákány, R.; et al. Signalling Alterations in Bones of Pituitary Adenylate Cyclase Activating Polypeptide (PACAP) Gene Deficient Mice. Int. J. Mol. Sci. 2018, 19, 2538. https://doi.org/10.3390/ijms19092538
Józsa G, Szegeczki V, Pálfi A, Kiss T, Helyes Z, Fülöp B, Cserháti C, Daróczi L, Tamás A, Zákány R, et al. Signalling Alterations in Bones of Pituitary Adenylate Cyclase Activating Polypeptide (PACAP) Gene Deficient Mice. International Journal of Molecular Sciences. 2018; 19(9):2538. https://doi.org/10.3390/ijms19092538
Chicago/Turabian StyleJózsa, Gergő, Vince Szegeczki, Andrea Pálfi, Tamás Kiss, Zsuzsanna Helyes, Balázs Fülöp, Csaba Cserháti, Lajos Daróczi, Andrea Tamás, Róza Zákány, and et al. 2018. "Signalling Alterations in Bones of Pituitary Adenylate Cyclase Activating Polypeptide (PACAP) Gene Deficient Mice" International Journal of Molecular Sciences 19, no. 9: 2538. https://doi.org/10.3390/ijms19092538
APA StyleJózsa, G., Szegeczki, V., Pálfi, A., Kiss, T., Helyes, Z., Fülöp, B., Cserháti, C., Daróczi, L., Tamás, A., Zákány, R., Reglődi, D., & Juhász, T. (2018). Signalling Alterations in Bones of Pituitary Adenylate Cyclase Activating Polypeptide (PACAP) Gene Deficient Mice. International Journal of Molecular Sciences, 19(9), 2538. https://doi.org/10.3390/ijms19092538