Pulmonary Vascular Platform Models the Effects of Flow and Pressure on Endothelial Dysfunction in BMPR2 Associated Pulmonary Arterial Hypertension

 , and
, and
Abstract
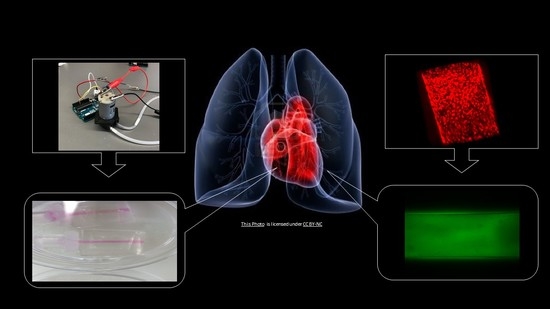
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Engineering an Artificial Arteriole
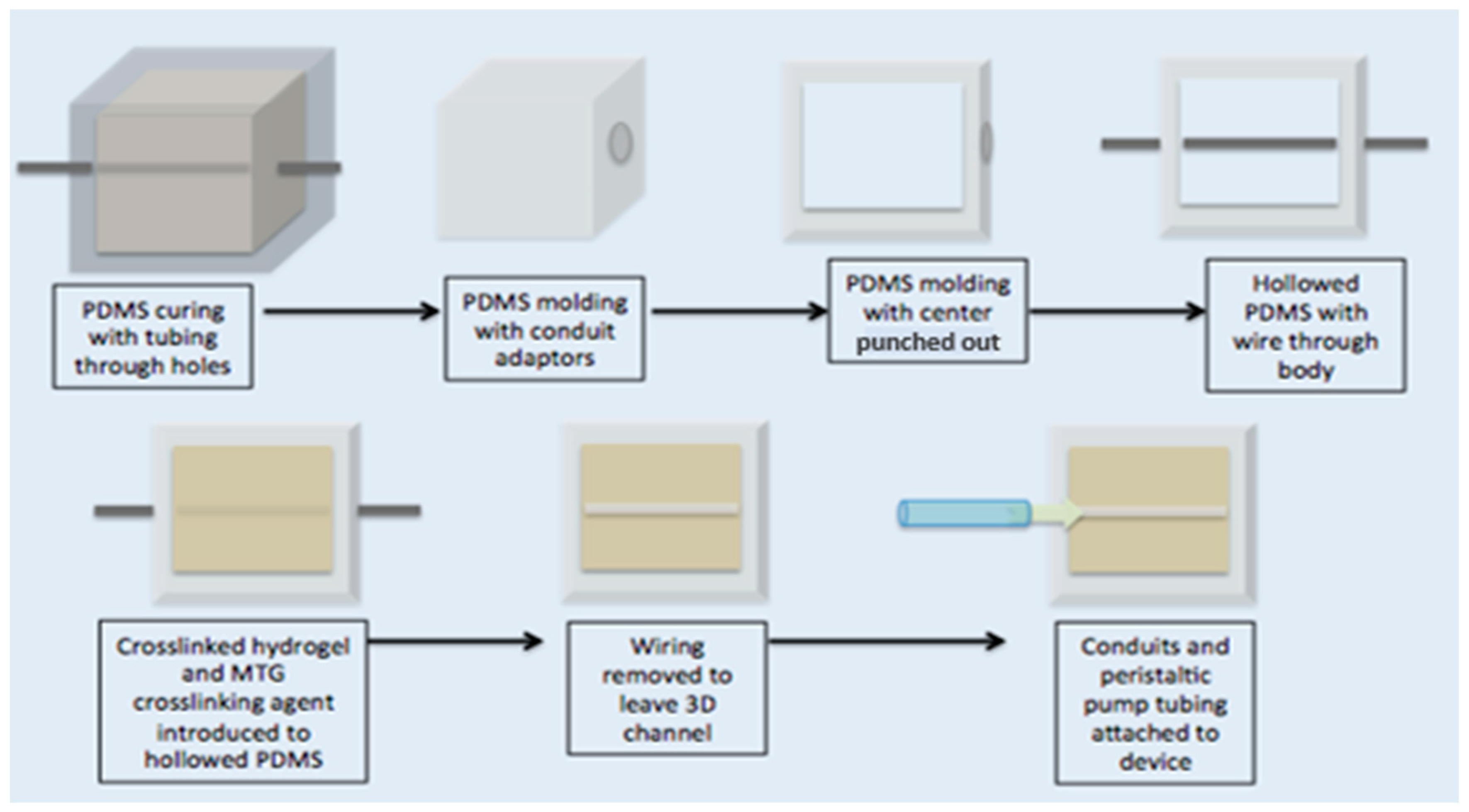
2.2. Artificial Pulmonary Arteriole Validation.
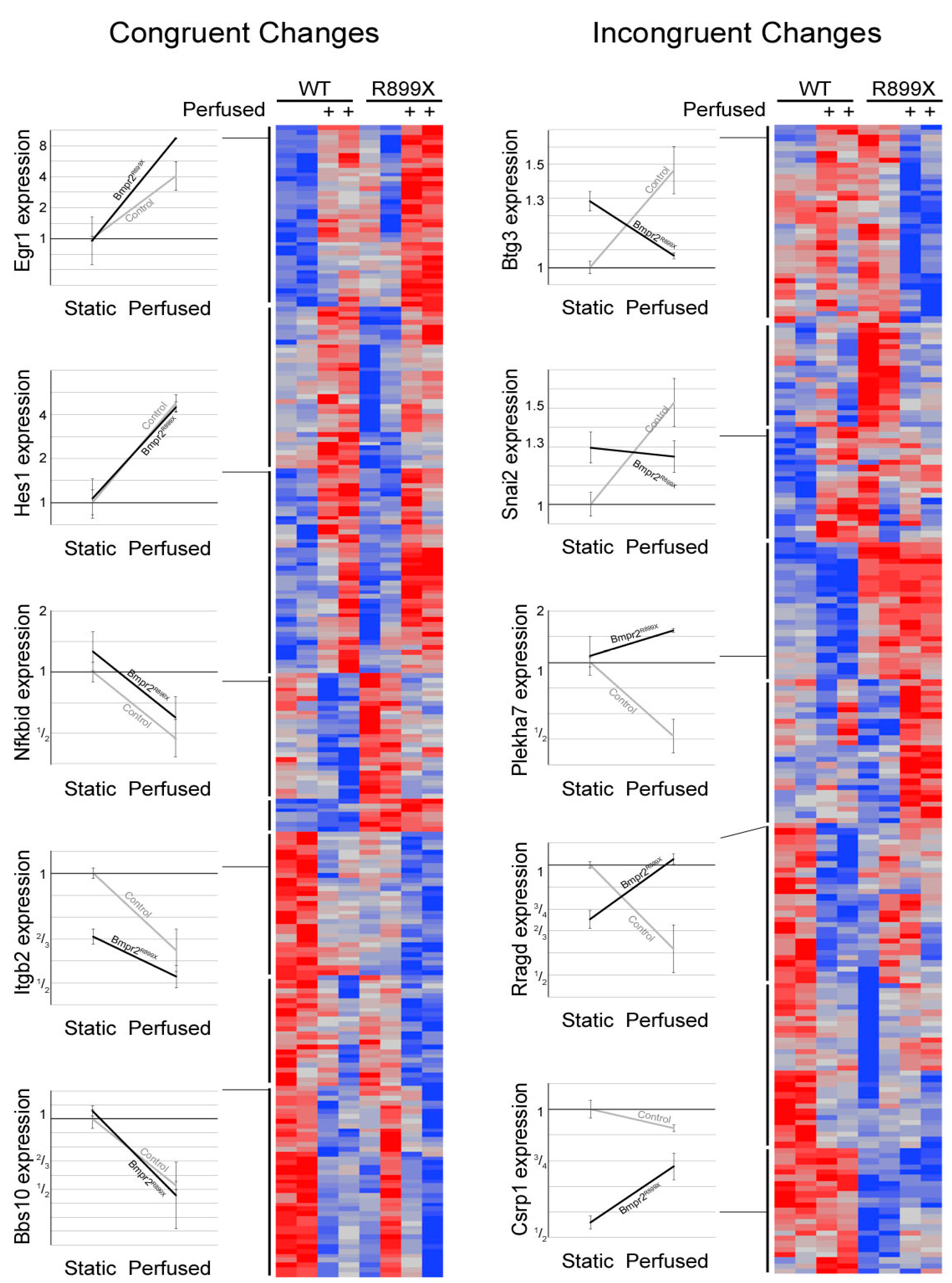
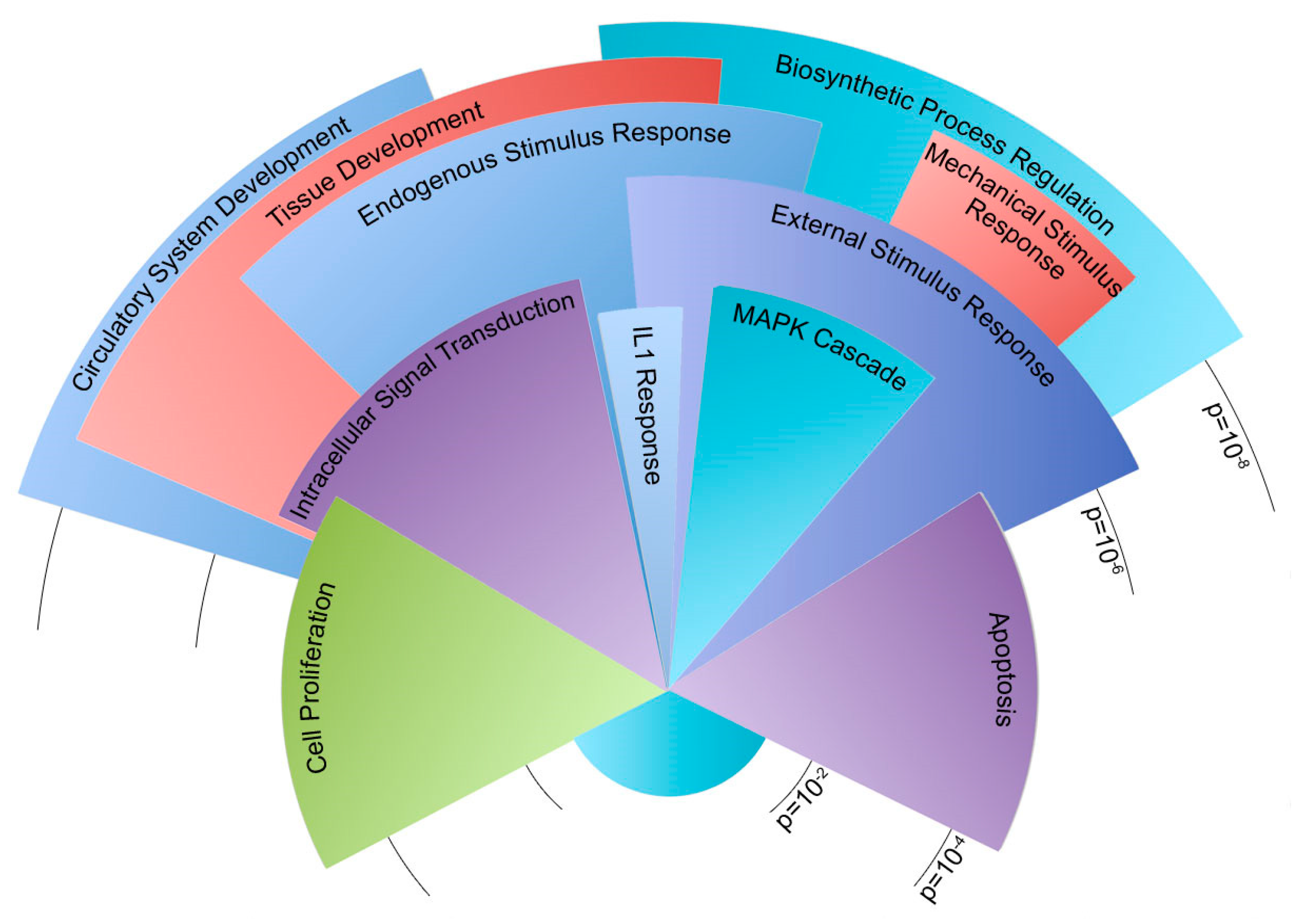
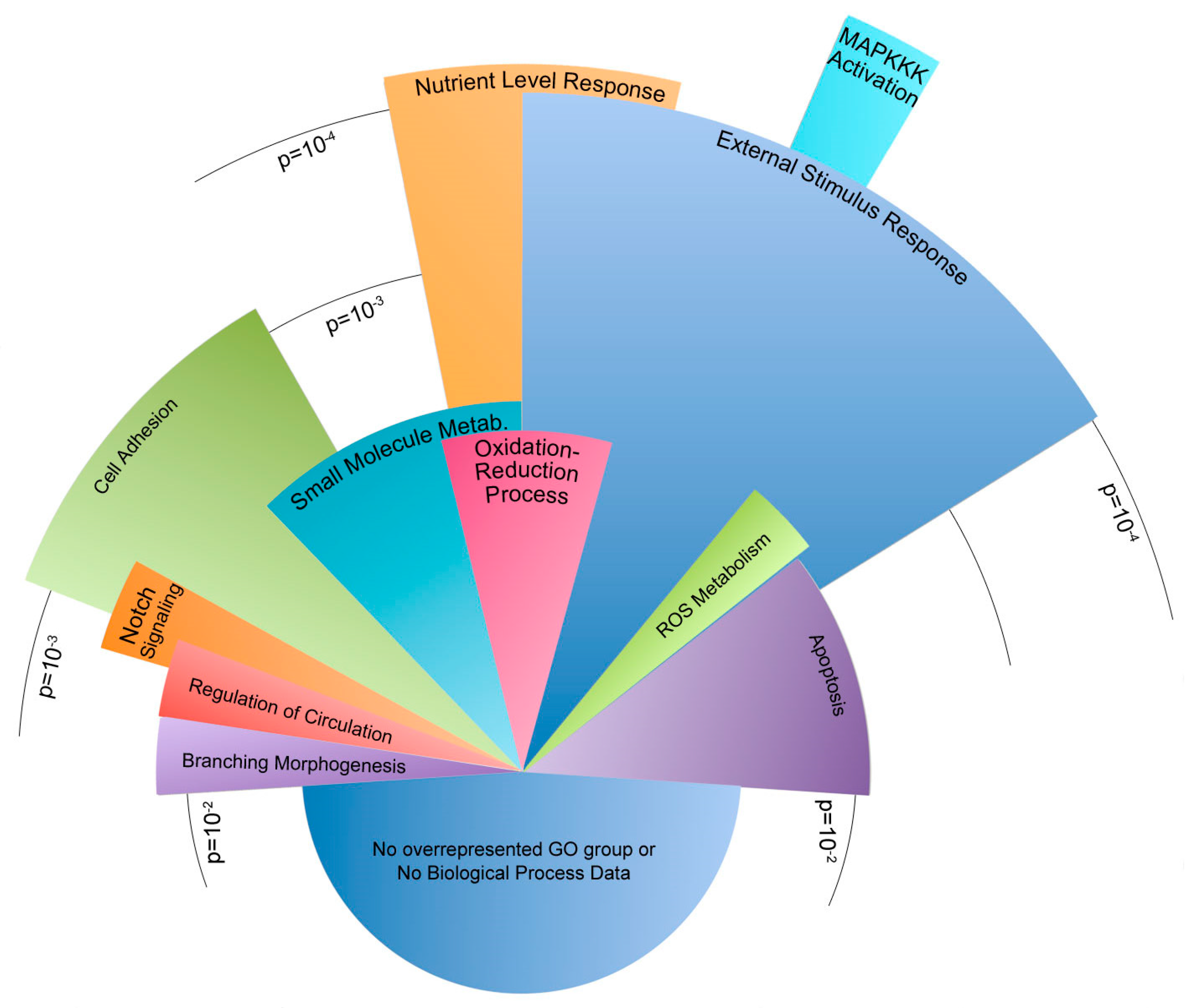
2.3. Oscillatory Flow Drives Genome-Wide Changes in WT and Bmpr2R899X Pulmonary Endothelial Cells
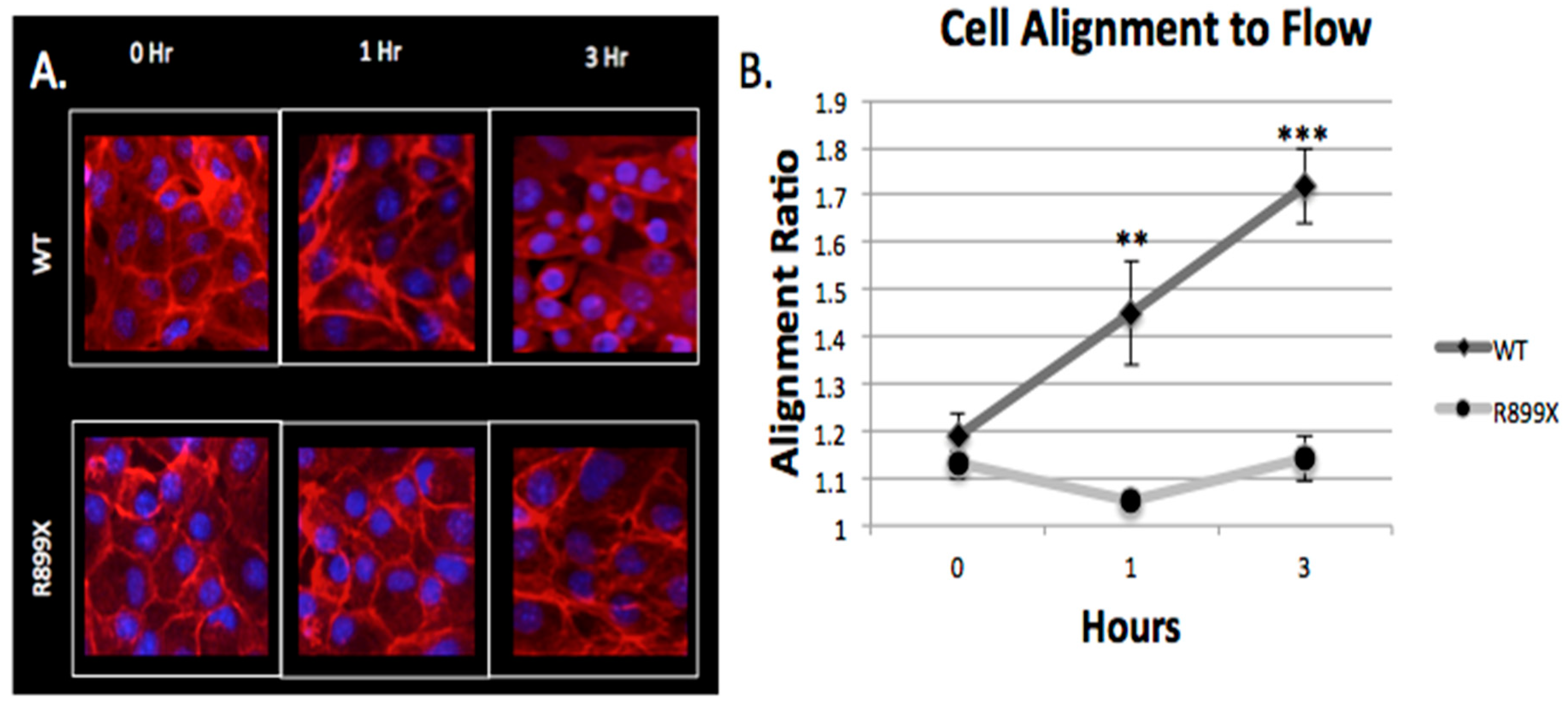
2.4. Bmpr2R899X Pulmonary Endothelial Cells Do Not Change Morphology in Response to Flow
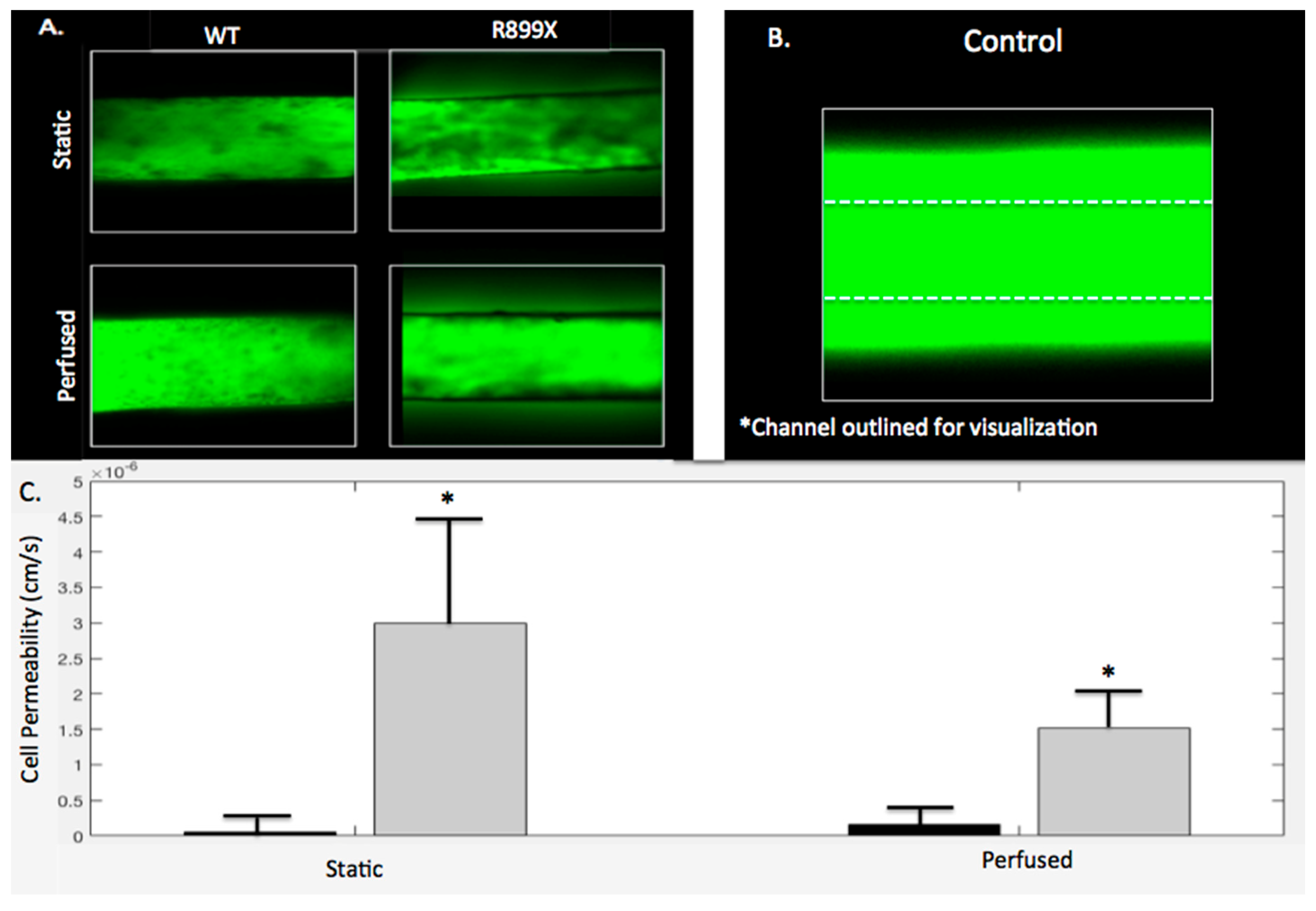
2.5. Bmpr2R899X Pulmonary Endothelial Cells Demonstrate Significant Barrier Dysfunction In-Vitro
3. Discussion
4. Materials and Methods
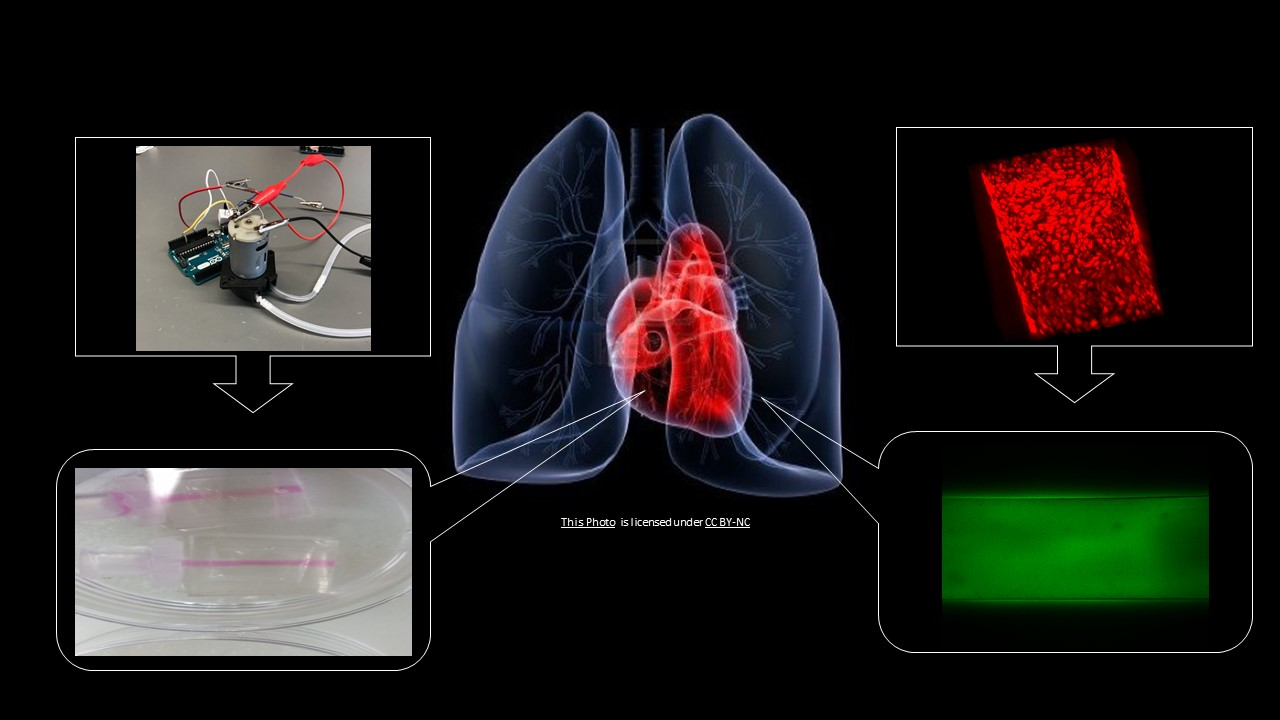
4.1. Device Construction
4.2. Bmpr2R899X and WT Cells
4.3. Cell Seeding
4.4. Platform Set-Up
4.5. Assessment of Artificial Lumen
4.6. RNA Sequencing
4.7. Endothelial Alignment
4.8. Barrier Function
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BMPR2 | Bone Morphogenetic Protein Type II Receptor |
| MPMVEC | Murine Pulmonary Microvascular Endothelial Cells |
| MTG | Microbial transglutaminase |
| PAH | Pulmonary Arterial Hypertension |
| WT | Wild type |
References
- Cool, C.D.; Stewart, J.S. Three-dimensional reconstruction of pulmonary arteries in plexiform pulmonary hypertension using cell-specific markers. Evidence for a dynamic and heterogeneous process of pulmonary endothelial cell growth. Am. J. Pathol. 1999, 155, 411–419. [Google Scholar] [CrossRef]
- Austin, E.D.; Loyd, J.E. The genetics of pulmonary arterial hypertension. Circ. Res. 2014, 115, 189–202. [Google Scholar] [CrossRef] [PubMed]
- Austin, E.D.; West, J. Molecular Medicine of Pulmonary Arterial Hypertension: From Population Genetics to Precision Medicine and Gene Editing. Am. J. Resp. Crit. Care Med. 2016, 195, 23–31. [Google Scholar] [CrossRef] [PubMed]
- De Jesus Perez, V.A. Molecular pathogenesis and current pathology of pulmonary hypertension. Heart Fail Rev. 2016, 21, 239–257. [Google Scholar] [CrossRef] [PubMed]
- Humbert, M.; Sitbon, O. Survival in patients with idiopathic, familial, and anorexigen-associated pulmonary arterial hypertension in the modern management era. Circulation 2010, 122, 156–163. [Google Scholar] [CrossRef] [PubMed]
- Rubin, L.J. Pulmonary arterial hypertension. Proc. Am. Thorac. Soc. 2006, 3, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Runo, J.R.; Loyd, J.E. Primary pulmonary hypertension. Lancet 2003, 361, 1533–1544. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.A.; Hemnes, A.R. Cytoskeletal defects in Bmpr2-associated pulmonary arterial hypertension. Am. J. Physiol. Lung Cell Mol. Physiol. 2012, 302, L474–L484. [Google Scholar] [CrossRef] [PubMed]
- Geraci, M.W.; Moore, M. Gene expression patterns in the lungs of patients with primary pulmonary hypertension: A gene microarray analysis. Circ. Res. 2001, 88, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Rajkumar, R.; Konishi, K. Genomewide RNA expression profiling in lung identifies distinct signatures in idiopathic pulmonary arterial hypertension and secondary pulmonary hypertension. Am. J. Physiol. Heart Circ. Physiol. 2010, 298, H1235–H1248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayless, K.J.; Davis, G.E. The Cdc42 and Rac1 GTPases are required for capillary lumen formation in three-dimensional extracellular matrices. J. Cell Sci. 2002, 115, 1123–1136. [Google Scholar] [PubMed]
- Connolly, J.O.; Simpson, N. Rac regulates endothelial morphogenesis and capillary assembly. Mol. Biol. Cell 2002, 13, 2193–2557. [Google Scholar] [CrossRef] [PubMed]
- Koh, W.; Mahan, R.D. Cdc42-and Rac1-mediated endothelial lumen formation requires Pak2, Pak4 and Par3, and PKC-dependent signaling. J. Cell Sci. 2008, 121, 989–1001. [Google Scholar] [CrossRef] [PubMed]
- Lovren, F.; Pan, Y. Angiotensin converting enzyme-2 confers endothelial protection and attenuates atherosclerosis. Am. J. Physiol. Heart Circ. Physiol. 2008, 295, H1377–H1384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Resnick, N.; Gimbrone, M.A. Hemodynamic forces are complex regulators of endothelial gene expression. FASEB J. 1995, 9, 874–882. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.P.; Li, Y.S. DNA microarray analysis of gene expression in endothelial cells in response to 24-h shear stress. Physiol. Genom. 2001, 7, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Blank, R.H.; Muller, W.H. Changes in pulmonary vascular lesions after restoring normal pulmonary artery pressure. Surg. Forum. 1958, 9, 356–359. [Google Scholar] [PubMed]
- Dammann, J.F.; McEachen, J.A. The regression of pulmonary vascular disease after the creation of pulmonary stenosis. J. Thorac. Cardiovasc. Surg. 1961, 42, 722–734. [Google Scholar] [PubMed]
- Ferguson, D.J.; Berkas, E.M. Process of healing in experimental pulmonary arteriosclerosis. Proc. Soc. Exp. Biol. Med. 1955, 89, 492–494. [Google Scholar] [CrossRef] [PubMed]
- Heath, D.; Helmholz, H.F. Relation between structural change in the small pulmonary arteries and the immediate reversibility of pulmonary hypertension following closure of ventricular and atrial septal defects. Circulation 1958, 18, 1167–1174. [Google Scholar] [CrossRef] [PubMed]
- Geer, J.C.; Glass, B.A. The Morphogenesis and Reversibility of Experimental Hyperkinetic Pulmonary Vascular Lesions in the Dog. Exp. Mol. Pathol. 1965, 4, 399–415. [Google Scholar] [CrossRef]
- Bertero, T.; Oldham, W.M. Vascular stiffness mechanoactivates YAP/TAZ-dependent glutaminolysis to drive pulmonary hypertension. J. Clin. Investig. 2016, 126, 3313–3335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, D.; Tan, Y. High pulsatility flow stimulates smooth muscle cell hypertrophy and contractile protein expression. Am. J. Physiol. Lung Cell Mol. Physiol. 2013, 304, L70–L81. [Google Scholar] [CrossRef] [PubMed]
- Wedgewood, S.; Lakshminrusimha, S. Cyclic stretch stimulates mitochondrial reactive oxygen species and Nox4 signaling in pulmonary artery smooth muscle cells. Am. J. Physiol. Lung Cell Mol. Physiol. 2015, 309, L196–L203. [Google Scholar] [CrossRef] [PubMed]
- Davies, P.F. Flow-mediated endothelial mechanotransduction. Physiol. Rev. 1995, 75, 519–560. [Google Scholar] [CrossRef] [PubMed]
- Ingber, D.E. Mechanical signaling and the cellular response to extracellular matrix in angiogenesis and cardiovascular physiology. Circ. Res. 2002, 91, 877–887. [Google Scholar] [CrossRef] [PubMed]
- Intengan, H.D.; Schiffrin, E.L. Structure and mechanical properties of resistance arteries in hypertension: Role of adhesion molecules and extracellular matrix determinants. Hypertension 2000, 36, 312–318. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.M.; Tsai, K.Y. Extracellular matrix and pulmonary hypertension: Control of vascular smooth muscle cell contractility. Am. J. Physiol. Heart Circ. Physiol. 1998, 274, H76–H82. [Google Scholar] [CrossRef]
- Negrini, D.; Gonano, C. Microvascular pressure profile in intact in situ lung. J. Appl. Physiol. 1992, 72, 332–339. [Google Scholar] [CrossRef] [PubMed]
- De Lee, G.J.; DuBois, A.B. Pulmonary capillary blood flow in man. J. Clin. Investig. 1955, 34, 1380–1390. [Google Scholar] [CrossRef] [PubMed]
- Haber, S.; Clark, A. Blood Flow in Capillaries of the Human Lung. J. Biomech. Eng. 2013, 135, 101006. [Google Scholar] [CrossRef] [PubMed]
- Levesque, M.J.; Nerem, R.M. The elongation and orientation of cultured endothelial cells in response to shear stress. J. Biomech. Eng. 1985, 107, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Dewey, C.F.; Bussolari, S.R. The dynamic response of vascular endothelial cells to fluid shear stress. J. Biomech. Eng. 1981, 103, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Tzima, E.; del Pozo, M.A. Activation of integrins in endothelial cells by fluid shear stress mediates Rho-dependent cytoskeletal alignment. EMBO J. 2001, 20, 4639–4647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gimbrone, M.A.; Topper, J.N. Endothelial dysfunction, hemodynamic forces, and atherogenesis. Ann. N. Y. Acad. Sci. 2000, 902, 230–240. [Google Scholar] [CrossRef] [PubMed]
- Blackman, B.R.; Guillermo, G.C. A new in vitro model to evaluate differential responses of endothelial cells to simulated arterial shear stress waveforms. J. Biomech. Eng. 2002, 124, 397–407. [Google Scholar] [CrossRef] [PubMed]
- Balcells, M.; Suárez, M.F. Cells in fluidic environments are sensitive to flow frequency. J. Cell Physiol. 2005, 204, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Scott, D.E. High pulsatility flow induces adhesion molecule and cytokine mRNA expression in distal pulmonary artery endothelial cells. Ann. Biomed. Eng. 2009, 37, 1082–1092. [Google Scholar] [CrossRef] [PubMed]
- Foletta, V.C.; Lim, M.A. Direct signaling by the BMP type II receptor via the cytoskeletal regulator LIMK1. J. Cell. Biol. 2003, 162, 1089–1098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machado, R.D.; Rudarakanchana, N. Functional interaction between BMPR-II and Tctex-1, a light chain of Dynein, is isoform-specific and disrupted by mutations underlying primary pulmonary hypertension. Hum. Mol. Genet. 2003, 12, 3277–3286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, W.K.; Knowles, J.A. Bone morphogenetic protein receptor type II C-terminus interacts with c-Src: Implication for a role in pulmonary arterial hypertension. Am. J. Respir. Cell Mol. Biol. 2005, 33, 438–446. [Google Scholar] [CrossRef] [PubMed]
- Budhiraja, R.; Rubin, M.T. Endothelial dysfunction in pulmonary hypertension. Circulation 2004, 109, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Humbert, M.; Morrell, N.W. Cellular and molecular pathobiology of pulmonary arterial hypertension. J. Am. Coll. Cardiol. 2004, 43, 13S–24S. [Google Scholar] [CrossRef] [PubMed]
- Morrell, N.W.; Adnot, S. Cellular and molecular basis of pulmonary arterial hypertension. J. Am. Coll. Cardiol. 2009, 54, 20S–31S. [Google Scholar] [CrossRef] [PubMed]
- Lo, C.M.; Wang, H.B. Cell movement is guided by the rigidity of the substrate. Biophys. J. 2000, 79, 144–152. [Google Scholar] [CrossRef]
- Bellan, L.M.; Pearsall, M. A 3D interconnected microchannel network formed in gelatin by sacrificial shellac microfibers. Adv. Mater. 2012, 24, 5187–5191. [Google Scholar] [CrossRef] [PubMed]
- Jat, P.S.; Noble, M.D. Direct derivation of conditionally immortal cell lines from an H-2Kb-tsA58 transgenic mouse. Proc. Natl. Acad. Sci. USA 1991, 88, 5096–5100. [Google Scholar] [CrossRef] [PubMed]
- Pozzi, A.; Moberg, P.E. Elevated matrix metalloprotease and angiostatin levels in integrin alpha 1 knockout mice cause reduced tumor vascularization. Proc. Natl. Acad. Sci. USA 2000, 97, 2202–2207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Duncan, D. WEB-based gene set analysis toolkit (WebGestalt): Update 2013. Nucle. Aci. Res. 2013, 41, W77–W83. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Stefan, K. WebGestalt: An integrated system for exploring gene sets in various biological contexts. Nucleic Acids Res. 2005, 33, W741–W748. [Google Scholar] [CrossRef] [PubMed]
- Arrio-Dupont, M.; Cribier, S. Diffusion of fluorescently labeled macromolecules in cultured muscle cells. Biophys. J. 1996, 70, 2327–2332. [Google Scholar] [CrossRef] [Green Version]
- Braeckmans, K.; Peeters, L. Three-dimensional fluorescence recovery after photobleaching with the confocal scanning laser microscope. Biophys. J. 2003, 85, P2240–P2252. [Google Scholar] [CrossRef]
- Kang, M.; Day, C.A. Simplified equation to extract diffusion coefficients from confocal FRAP data. Traffic 2012, 13, 1589–1600. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]







© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
D’Amico, R.W.; Faley, S.; Shim, H.-n.; Prosser, J.R.; Agrawal, V.; Bellan, L.M.; West, J.D. Pulmonary Vascular Platform Models the Effects of Flow and Pressure on Endothelial Dysfunction in BMPR2 Associated Pulmonary Arterial Hypertension. Int. J. Mol. Sci. 2018, 19, 2561. https://doi.org/10.3390/ijms19092561
D’Amico RW, Faley S, Shim H-n, Prosser JR, Agrawal V, Bellan LM, West JD. Pulmonary Vascular Platform Models the Effects of Flow and Pressure on Endothelial Dysfunction in BMPR2 Associated Pulmonary Arterial Hypertension. International Journal of Molecular Sciences. 2018; 19(9):2561. https://doi.org/10.3390/ijms19092561
Chicago/Turabian StyleD’Amico, Reid W., Shannon Faley, Ha-na Shim, Joanna R. Prosser, Vineet Agrawal, Leon M. Bellan, and James D. West. 2018. "Pulmonary Vascular Platform Models the Effects of Flow and Pressure on Endothelial Dysfunction in BMPR2 Associated Pulmonary Arterial Hypertension" International Journal of Molecular Sciences 19, no. 9: 2561. https://doi.org/10.3390/ijms19092561
APA StyleD’Amico, R. W., Faley, S., Shim, H.-n., Prosser, J. R., Agrawal, V., Bellan, L. M., & West, J. D. (2018). Pulmonary Vascular Platform Models the Effects of Flow and Pressure on Endothelial Dysfunction in BMPR2 Associated Pulmonary Arterial Hypertension. International Journal of Molecular Sciences, 19(9), 2561. https://doi.org/10.3390/ijms19092561