Development of a Transcriptional Amplification System Based on the PEG3 Promoter to Target Androgen Receptor-Positive and -Negative Prostate Cancer Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
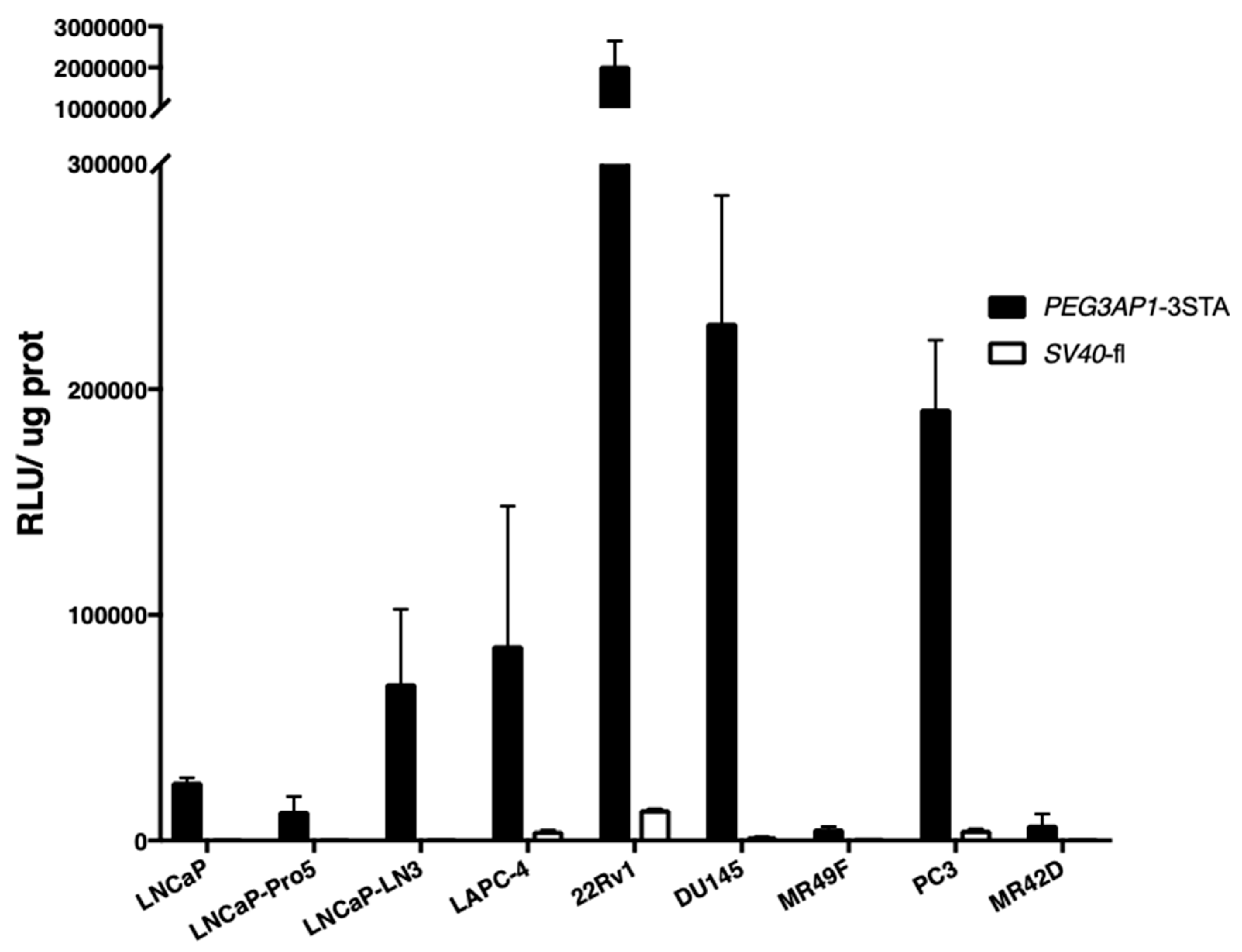
2.1. PEG3AP1-3STA Is a Highly Sensitive Amplification System
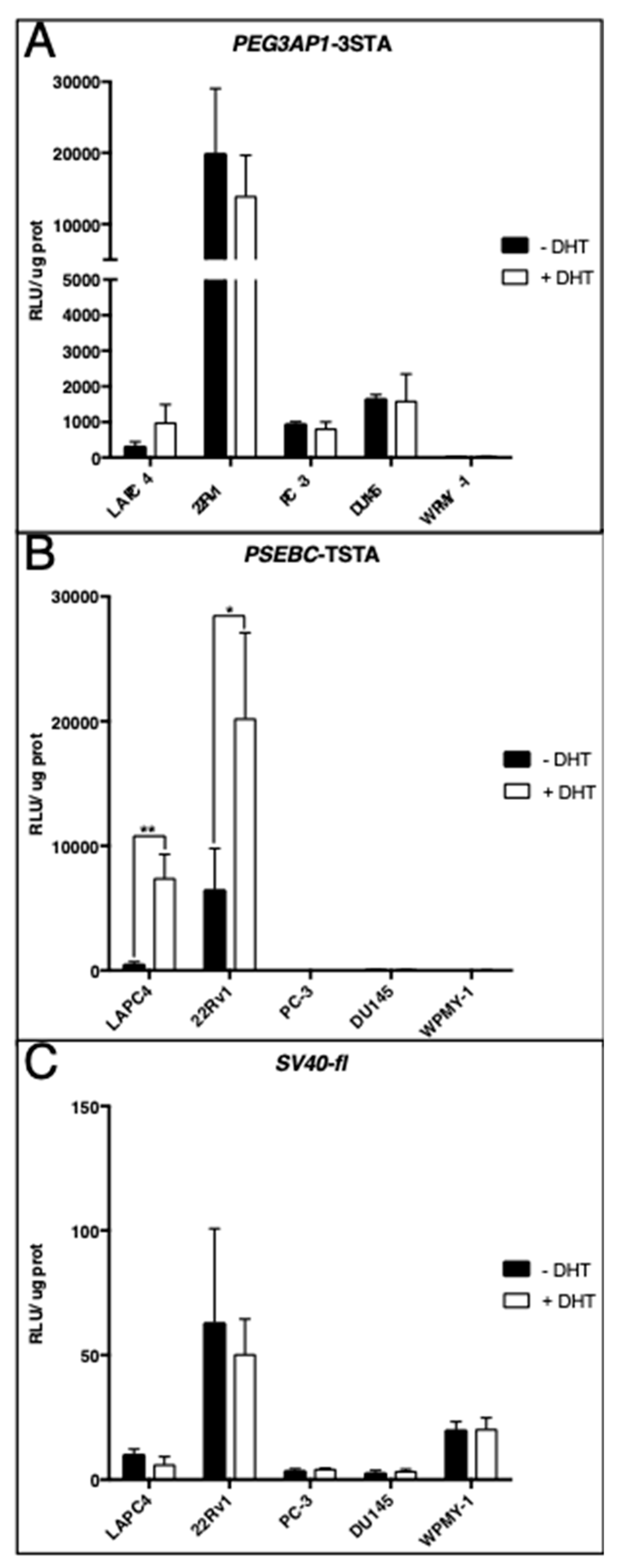
2.2. PEG3AP1-3STA Is an Androgen-Independent System
2.3. PEG3AP1-3STA Is Active in Both Adenocarcinoma and Neuroendocrine-Like Cell Lines
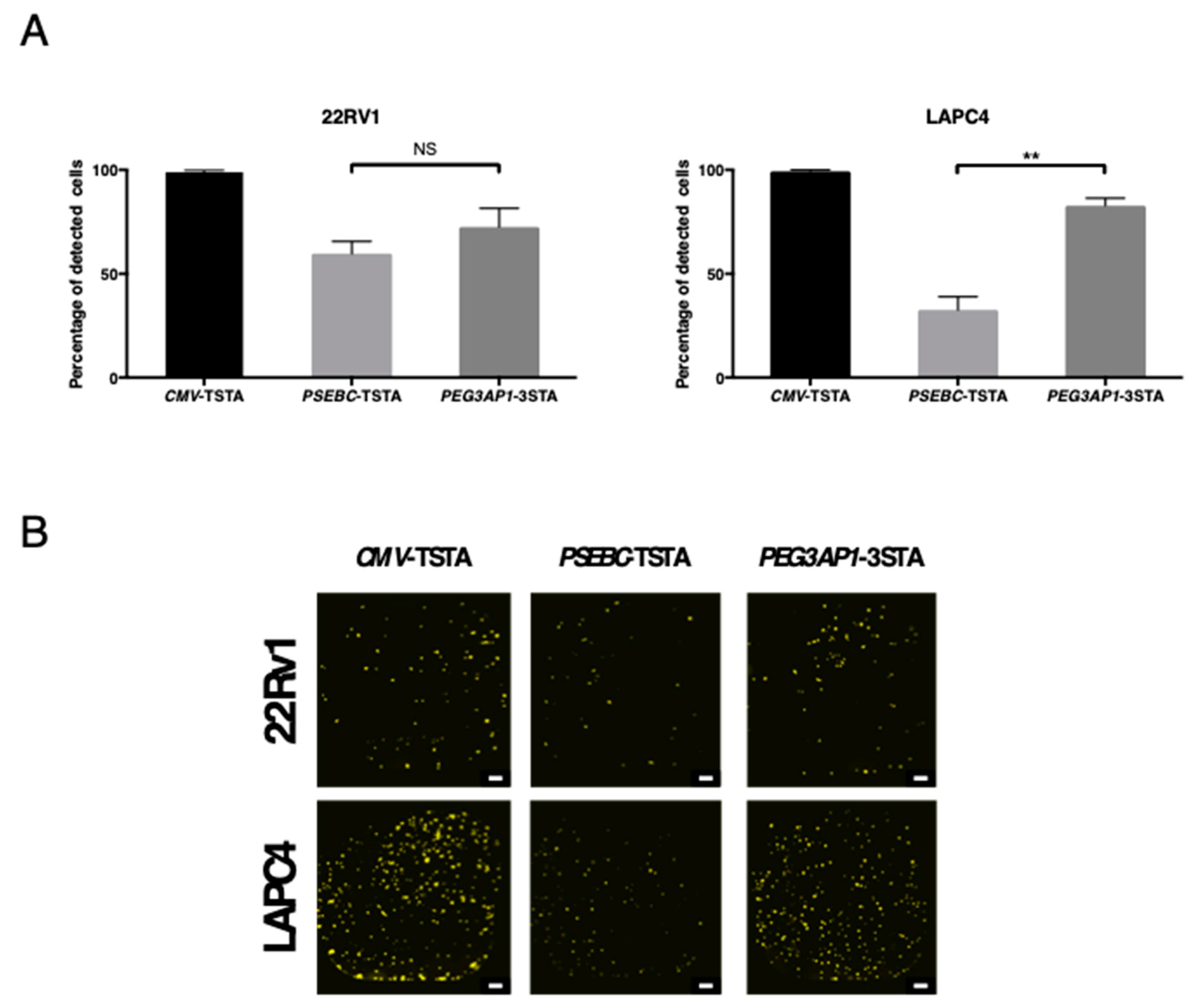
2.4. PEG3AP1-3STA Is Active in Heterogeneous PCa Cell Populations within AR+ Cell Lines
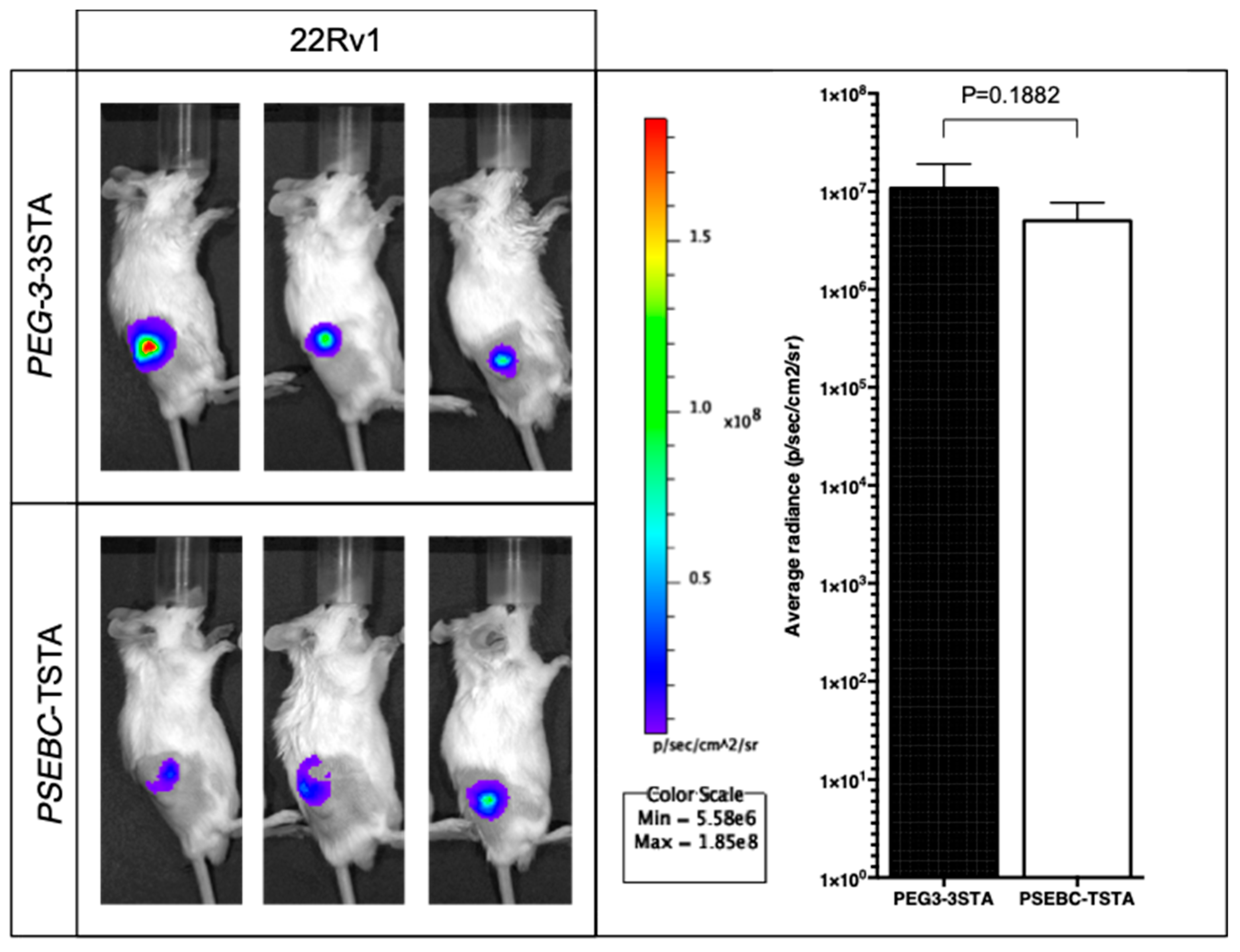
2.5. PEG3AP1-3STA Provides a Quantifiable Signal In Vivo That Is Higher Than That of Positron Emission Tomography (PET) Imageable PSEBC-TSTA
3. Discussion
4. Materials and Methods
4.1. Cloning and Virus Production
4.2. Cell Culture
4.3. Bioluminescence Quantification In Vitro
4.4. Bioluminescence Microscopy Detection
4.5. Bioluminescence Detection in Prostate Cancer Xenografts
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 3STA | Three-step amplification system |
| ADT | Androgen-deprivation therapy |
| AR | Androgen receptor |
| CRPC | Castration-resistant prostate cancer |
| DHT | Dihydrotestosterone |
| FBS | Fetal bovine serum |
| fl | Firefly luciferase |
| ivp | Infectious viral particules |
| MOI | Multiplicity of infection |
| NEPC | Neuroendocrine prostate cancer |
| PBS | Phosphate buffered saline |
| PCa | Prostate cancer |
| PEG3 | Progression elevated gene-3 |
| PET | Positron emission tomography |
| PSA | Prostate-specific antigen |
| RLU | Relative luciferase activity |
| TSTA | Two-step transcriptional amplification system |
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer Statistics, 2017. CA 2017, 67, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Resnick, M.J.; Koyama, T.; Fan, K.H.; Albertsen, P.C.; Goodman, M.; Hamilton, A.S.; Hoffman, R.M.; Potosky, A.L.; Stanford, J.L.; Stroup, A.M.; et al. Long-term functional outcomes after treatment for localized prostate cancer. N. Engl. J. Med. 2013, 368, 436–445. [Google Scholar] [CrossRef] [PubMed]
- Loblaw, D.A.; Virgo, K.S.; Nam, R.; Somerfield, M.R.; Ben-Josef, E.; Mendelson, D.S.; Middleton, R.; Sharp, S.A.; Smith, T.J.; Talcott, J.; et al. Initial hormonal management of androgen-sensitive metastatic, recurrent, or progressive prostate cancer: 2006 update of an American Society of Clinical Oncology practice guideline. J. Clin. Oncol. 2007, 25, 1596–1605. [Google Scholar] [CrossRef] [PubMed]
- Perlmutter, M.A.; Lepor, H. Androgen deprivation therapy in the treatment of advanced prostate cancer. Rev. Urol. 2007, 9 (Suppl. 1), S3–S8. [Google Scholar]
- Feldman, B.J.; Feldman, D. The development of androgen-independent prostate cancer. Nat. Rev. Cancer 2001, 1, 34–45. [Google Scholar] [CrossRef] [PubMed]
- Gleave, M.; Tolcher, A.; Miyake, H.; Nelson, C.; Brown, B.; Beraldi, E.; Goldie, J. Progression to androgen independence is delayed by adjuvant treatment with antisense Bcl-2 oligodeoxynucleotides after castration in the LNCaP prostate tumor model. Clin. Cancer Res. 1999, 5, 2891–2898. [Google Scholar] [PubMed]
- Zou, M.; Toivanen, R.; Mitrofanova, A.; Floch, N.; Hayati, S.; Sun, Y.; Magnen, C.L.; Chester, D.; Mostaghel, E.A.; Califano, A.; et al. Transdifferentiation as a mechanism of treatment resistance in a mouse model of castration-resistant prostate cancer. Cancer Discov. 2017. [Google Scholar] [CrossRef]
- Beltran, H.; Rickman, D.S.; Park, K.; Chae, S.S.; Sboner, A.; MacDonald, T.Y.; Wang, Y.; Sheikh, K.L.; Terry, S.; Tagawa, S.T.; et al. Molecular characterization of neuroendocrine prostate cancer and identification of new drug targets. Cancer Discov. 2011, 1, 487–495. [Google Scholar] [CrossRef]
- Clermont, P.L.; Lin, D.; Crea, F.; Wu, R.; Xue, H.; Wang, Y.; Thu, K.L.; Lam, W.L.; Collins, C.C.; Wang, Y.; et al. Polycomb-mediated silencing in neuroendocrine prostate cancer. Clin. Epigenetics 2015, 7, 40. [Google Scholar] [CrossRef] [Green Version]
- Beauregard, J.M.; Pouliot, F. New developments in the imaging of metastatic prostate cancer. Curr. Opin. Support Palliat. Care 2014, 8, 265–270. [Google Scholar] [CrossRef]
- Drake, C.G. Prostate cancer as a model for tumour immunotherapy. Nat. Rev. Immunol. 2010, 10, 580–593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pouliot, F.; Karanikolas, B.D.; Johnson, M.; Sato, M.; Priceman, S.J.; Stout, D.; Sohn, J.; Satyamurthy, N.; deKernion, J.B.; Wu, L. In vivo imaging of intraprostatic-specific gene transcription by PET. J. Nucl Med. 2011, 52, 784–791. [Google Scholar] [CrossRef] [PubMed]
- Keu, K.V.; Witney, T.H.; Yaghoubi, S.; Rosenberg, J.; Kurien, A.; Magnusson, R.; Williams, J.; Habte, F.; Wagner, J.R.; Forman, S.; et al. Reporter gene imaging of targeted T cell immunotherapy in recurrent glioma. Sci. Transl. Med. 2017, 9. [Google Scholar] [CrossRef] [PubMed]
- Yaghoubi, S.; Barrio, J.R.; Dahlbom, M.; Iyer, M.; Namavari, M.; Satyamurthy, N.; Goldman, R.; Herschman, H.R.; Phelps, M.E.; Gambhir, S.S. Human pharmacokinetic and dosimetry studies of [(18)F]FHBG: A reporter probe for imaging herpes simplex virus type-1 thymidine kinase reporter gene expression. J. Nucl Med. 2001, 42, 1225–1234. [Google Scholar]
- Neveu, B.; Jain, P.; Tetu, B.; Wu, L.; Fradet, Y.; Pouliot, F. A PCA3 gene-based transcriptional amplification system targeting primary prostate cancer. Oncotarget 2016, 7, 1300–1310. [Google Scholar] [CrossRef] [PubMed]
- Bhang, H.E.; Gabrielson, K.L.; Laterra, J.; Fisher, P.B.; Pomper, M.G. Tumor-specific imaging through progression elevated gene-3 promoter-driven gene expression. Nat. Med. 2011, 17, 123–129. [Google Scholar] [CrossRef]
- Su, Z.Z.; Sarkar, D.; Emdad, L.; Duigou, G.J.; Young, C.S.; Ware, J.; Randolph, A.; Valerie, K.; Fisher, P.B. Targeting gene expression selectively in cancer cells by using the progression-elevated gene-3 promoter. Proc. Natl. Acad. Sci. USA 2005, 102, 1059–1064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, I.; Lebedeva, I.V.; Su, Z.Z.; Sarkar, D.; Valerie, K.; Fisher, P.B. Progression elevated gene-3 promoter (PEG-Prom) confers cancer cell selectivity to human polynucleotide phosphorylase (hPNPase(old-35))-mediated growth suppression. J. Cell. Physiol. 2008, 215, 401–409. [Google Scholar] [CrossRef]
- Pouliot, F.; Johnson, M.; Wu, L. Non-invasive molecular imaging of prostate cancer lymph node metastasis. Trends Mol. Med. 2009, 15, 254–262. [Google Scholar] [CrossRef] [Green Version]
- Kajanne, R.; Miettinen, P.; Tenhunen, M.; Leppa, S. Transcription factor AP-1 promotes growth and radioresistance in prostate cancer cells. Int. J. Oncol. 2009, 35, 1175–1182. [Google Scholar] [CrossRef]
- Jain, P.; Neveu, B.; Velot, L.; Wu, L.; Fradet, Y.; Pouliot, F. Bioluminescence microscopy as a method to measure single cell androgen receptor activity heterogeneous responses to antiandrogens. Sci. Rep. 2016, 6, 33968. [Google Scholar] [CrossRef] [PubMed]
- Bishop, J.L.; Thaper, D.; Vahid, S.; Davies, A.; Ketola, K.; Kuruma, H.; Jama, R.; Nip, K.M.; Angeles, A.; Johnson, F.; et al. The master neural transcription factor BRN2 is an androgen receptor-suppressed driver of neuroendocrine differentiation in prostate cancer. Cancer Discov. 2017, 7, 54–71. [Google Scholar] [CrossRef] [PubMed]
- Kuruma, H.; Matsumoto, H.; Shiota, M.; Bishop, J.; Lamoureux, F.; Thomas, C.; Briere, D.; Los, G.; Gleave, M.; Fanjul, A.; et al. A novel antiandrogen, Compound 30, suppresses castration-resistant and MDV3100-resistant prostate cancer growth in vitro and in vivo. Mol. Cancer Ther. 2013, 12, 567–576. [Google Scholar] [CrossRef] [PubMed]
- Tai, S.; Sun, Y.; Squires, J.M.; Zhang, H.; Oh, W.K.; Liang, C.Z.; Huang, J. PC3 is a cell line characteristic of prostatic small cell carcinoma. Prostate 2011, 71, 1668–1679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarkar, D.; Lebedeva, I.V.; Su, Z.Z.; Park, E.S.; Chatman, L.; Vozhilla, N.; Dent, P.; Curiel, D.T.; Fisher, P.B. Eradication of therapy-resistant human prostate tumors using a cancer terminator virus. Cancer Res. 2007, 67, 5434–5442. [Google Scholar] [CrossRef] [PubMed]
- Saad, F.; Chi, K.N.; Finelli, A.; Hotte, S.J.; Izawa, J.; Kapoor, A.; Kassouf, W.; Loblaw, A.; North, S.; Rendon, R.; et al. The 2015 CUA-CUOG Guidelines for the management of castration-resistant prostate cancer (CRPC). Can. Urol. Assoc. J. 2015, 9, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Pezaro, C.J.; Omlin, A.; Lorente, D.; Nava Rodrigues, D.; Ferraldeschi, R.; Bianchini, D.; Mukherji, D.; Riisnaes, R.; Altavilla, A.; Crespo, M.; et al. Visceral disease in castration-resistant prostate cancer. Eur. Urol. 2014, 65, 270–273. [Google Scholar] [CrossRef]
- Devaud, C.; John, L.B.; Westwood, J.A.; Darcy, P.K.; Kershaw, M.H. Immune modulation of the tumor microenvironment for enhancing cancer immunotherapy. Oncoimmunology 2013, 2, e25961. [Google Scholar] [CrossRef]
- Gambhir, S.S.; Bauer, E.; Black, M.E.; Liang, Q.; Kokoris, M.S.; Barrio, J.R.; Iyer, M.; Namavari, M.; Phelps, M.E.; Herschman, H.R. A mutant herpes simplex virus type 1 thymidine kinase reporter gene shows improved sensitivity for imaging reporter gene expression with positron emission tomography. Proc. Natl. Acad. Sci. USA 2000, 97, 2785–2790. [Google Scholar] [CrossRef] [Green Version]
- Li, L.Q.; Shen, F.; Xu, X.Y.; Zhang, H.; Yang, X.F.; Liu, W.G. Gene therapy with HSV1-sr39TK/GCV exhibits a stronger therapeutic efficacy than HSV1-TK/GCV in rat C6 glioma cells. Sci. World J. 2013, 2013, 951343. [Google Scholar] [CrossRef]
- Gorin, M.A.; Verdone, J.E.; van der Toom, E.; Bivalacqua, T.J.; Allaf, M.E.; Pienta, K.J. Circulating tumour cells as biomarkers of prostate, bladder, and kidney cancer. Nat. Rev. Urol. 2017, 14, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Heller, G.; McCormack, R.; Kheoh, T.; Molina, A.; Smith, M.R.; Dreicer, R.; Saad, F.; de Wit, R.; Aftab, D.T.; Hirmand, M.; et al. Circulating tumor cell number as a response measure of prolonged survival for metastatic castration-resistant prostate cancer: A comparison with prostate-specific antigen across five randomized phase III clinical trials. J. Clin. Oncol. 2018, 36, 572–580. [Google Scholar] [CrossRef] [PubMed]
- Scher, H.I.; Graf, R.P.; Schreiber, N.A.; McLaughlin, B.; Jendrisak, A.; Wang, Y.; Lee, J.; Greene, S.; Krupa, R.; Lu, D.; et al. Phenotypic heterogeneity of circulating tumor cells informs clinical decisions between AR signaling inhibitors and taxanes in metastatic prostate cancer. Cancer Res. 2017, 77, 5687–5698. [Google Scholar] [CrossRef]
- Hwang, J.E.; Joung, J.Y.; Shin, S.P.; Choi, M.K.; Kim, J.E.; Kim, Y.H.; Park, W.S.; Lee, S.J.; Lee, K.H. Ad5/35E1aPSESE4: A novel approach to marking circulating prostate tumor cells with a replication competent adenovirus controlled by PSA/PSMA transcription regulatory elements. Cancer Lett. 2016, 372, 57–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fougeroux, C.; Holst, P.J. Future prospects for the development of cost-effective adenovirus vaccines. Int. J. Mol. Sci. 2017, 18, 686. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Matherly, J.; Smallwood, A.; Adams, J.Y.; Billick, E.; Belldegrun, A.; Carey, M. Chimeric PSA enhancers exhibit augmented activity in prostate cancer gene therapy vectors. Gene Ther. 2001, 8, 1416–1426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iyer, M.; Wu, L.; Carey, M.; Wang, Y.; Smallwood, A.; Gambhir, S.S. Two-step transcriptional amplification as a method for imaging reporter gene expression using weak promoters. Proc. Natl. Acad. Sci. USA 2001, 98, 14595–14600. [Google Scholar] [CrossRef] [Green Version]
- Bishop, J.L.; Sio, A.; Angeles, A.; Roberts, M.E.; Azad, A.A.; Chi, K.N.; Zoubeidi, A. PD-L1 is highly expressed in Enzalutamide resistant prostate cancer. Oncotarget 2015, 6, 234–242. [Google Scholar] [CrossRef] [PubMed]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jain, P.; Clermont, P.-L.; Desmeules, F.; Zoubeidi, A.; Neveu, B.; Pouliot, F. Development of a Transcriptional Amplification System Based on the PEG3 Promoter to Target Androgen Receptor-Positive and -Negative Prostate Cancer Cells. Int. J. Mol. Sci. 2019, 20, 216. https://doi.org/10.3390/ijms20010216
Jain P, Clermont P-L, Desmeules F, Zoubeidi A, Neveu B, Pouliot F. Development of a Transcriptional Amplification System Based on the PEG3 Promoter to Target Androgen Receptor-Positive and -Negative Prostate Cancer Cells. International Journal of Molecular Sciences. 2019; 20(1):216. https://doi.org/10.3390/ijms20010216
Chicago/Turabian StyleJain, Pallavi, Pier-Luc Clermont, Francis Desmeules, Amina Zoubeidi, Bertrand Neveu, and Frédéric Pouliot. 2019. "Development of a Transcriptional Amplification System Based on the PEG3 Promoter to Target Androgen Receptor-Positive and -Negative Prostate Cancer Cells" International Journal of Molecular Sciences 20, no. 1: 216. https://doi.org/10.3390/ijms20010216
APA StyleJain, P., Clermont, P. -L., Desmeules, F., Zoubeidi, A., Neveu, B., & Pouliot, F. (2019). Development of a Transcriptional Amplification System Based on the PEG3 Promoter to Target Androgen Receptor-Positive and -Negative Prostate Cancer Cells. International Journal of Molecular Sciences, 20(1), 216. https://doi.org/10.3390/ijms20010216