Physiological Analysis and Proteome Quantification of Alligator Weed Stems in Response to Potassium Deficiency Stress

Abstract
:1. Introduction
2. Results
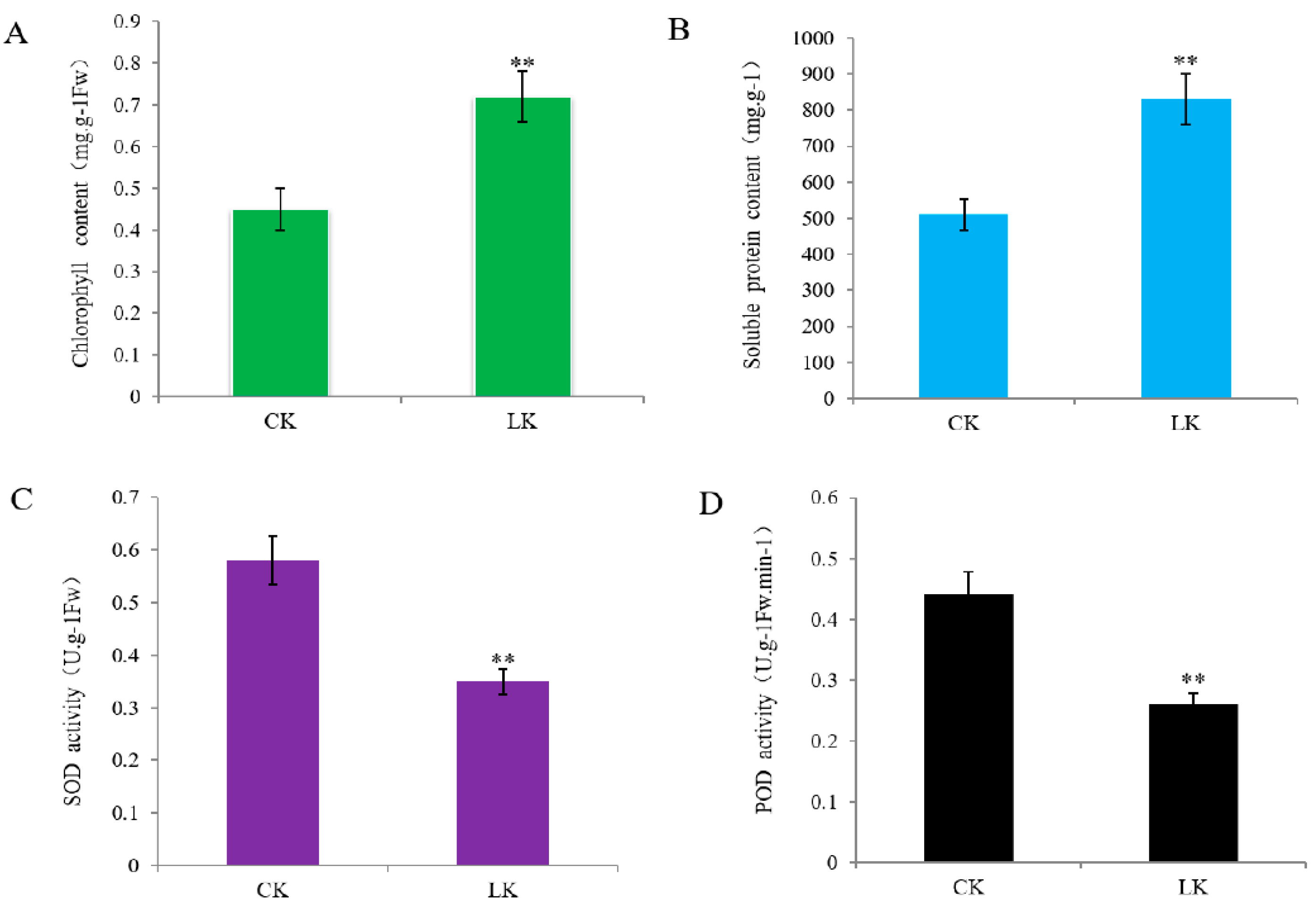
2.1. Effect of LK Stress on the Physiology of Alligator Weed Stems
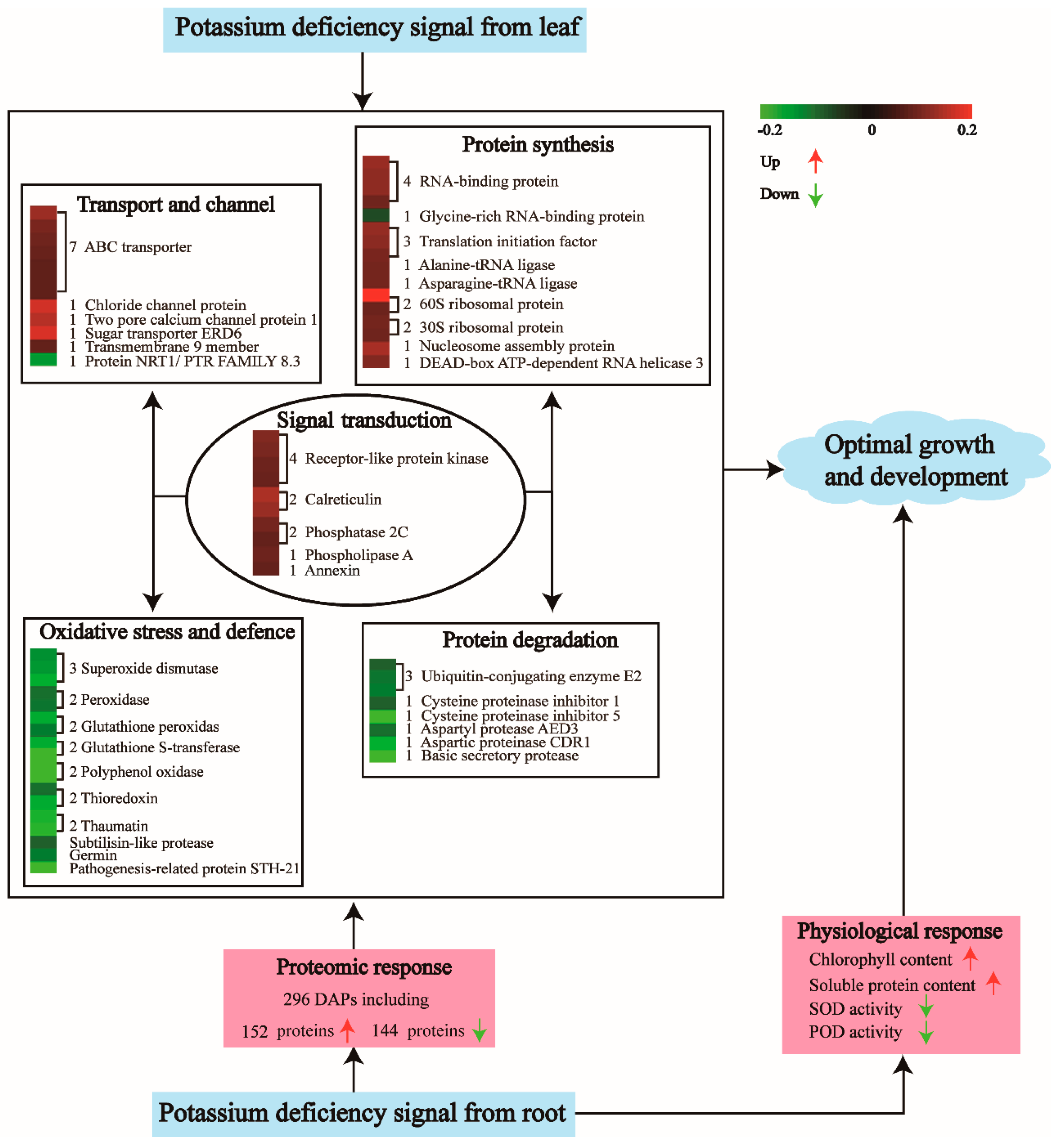
2.2. Protein Responses to LK Stress Revealed by the Proteomic Analysis
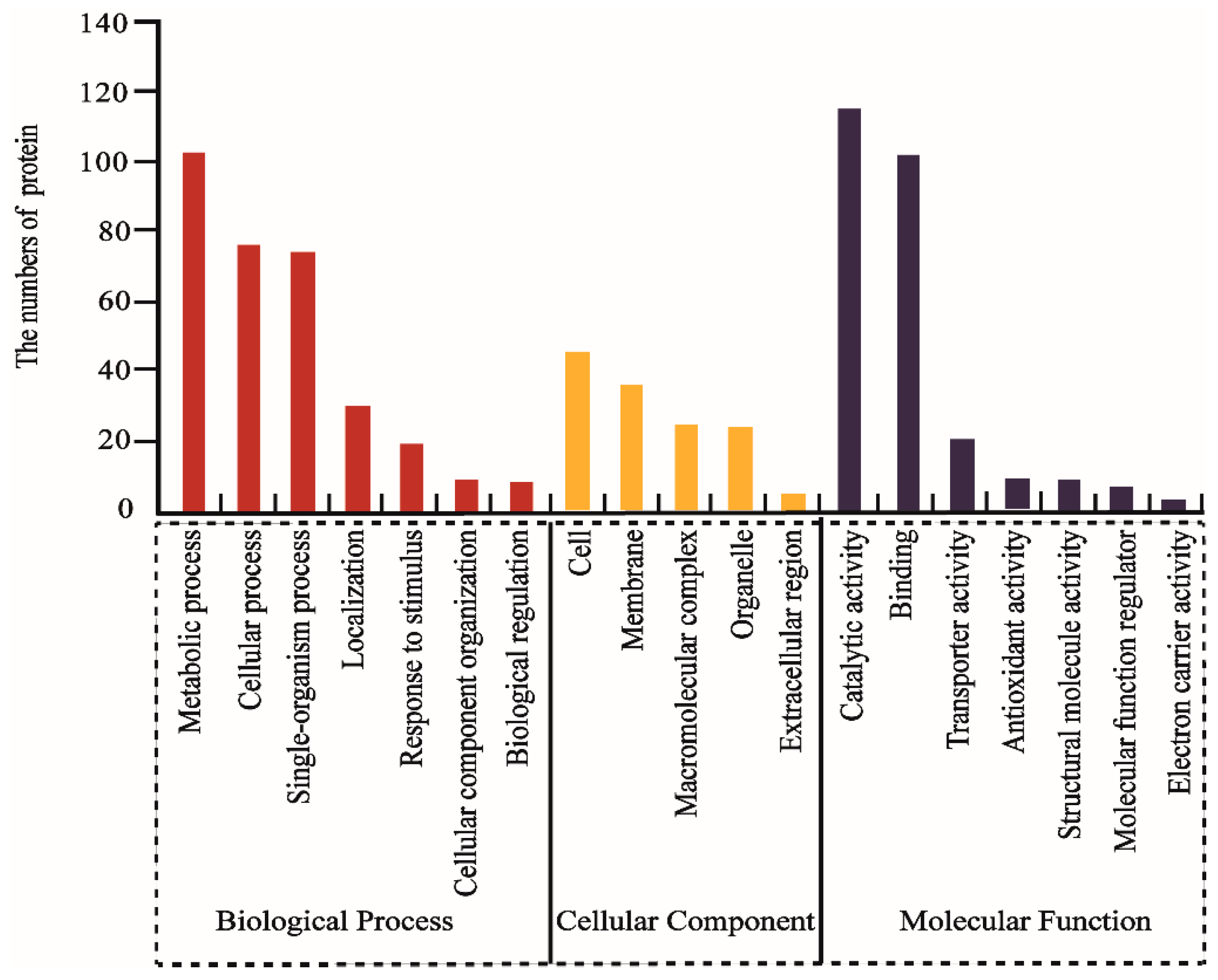
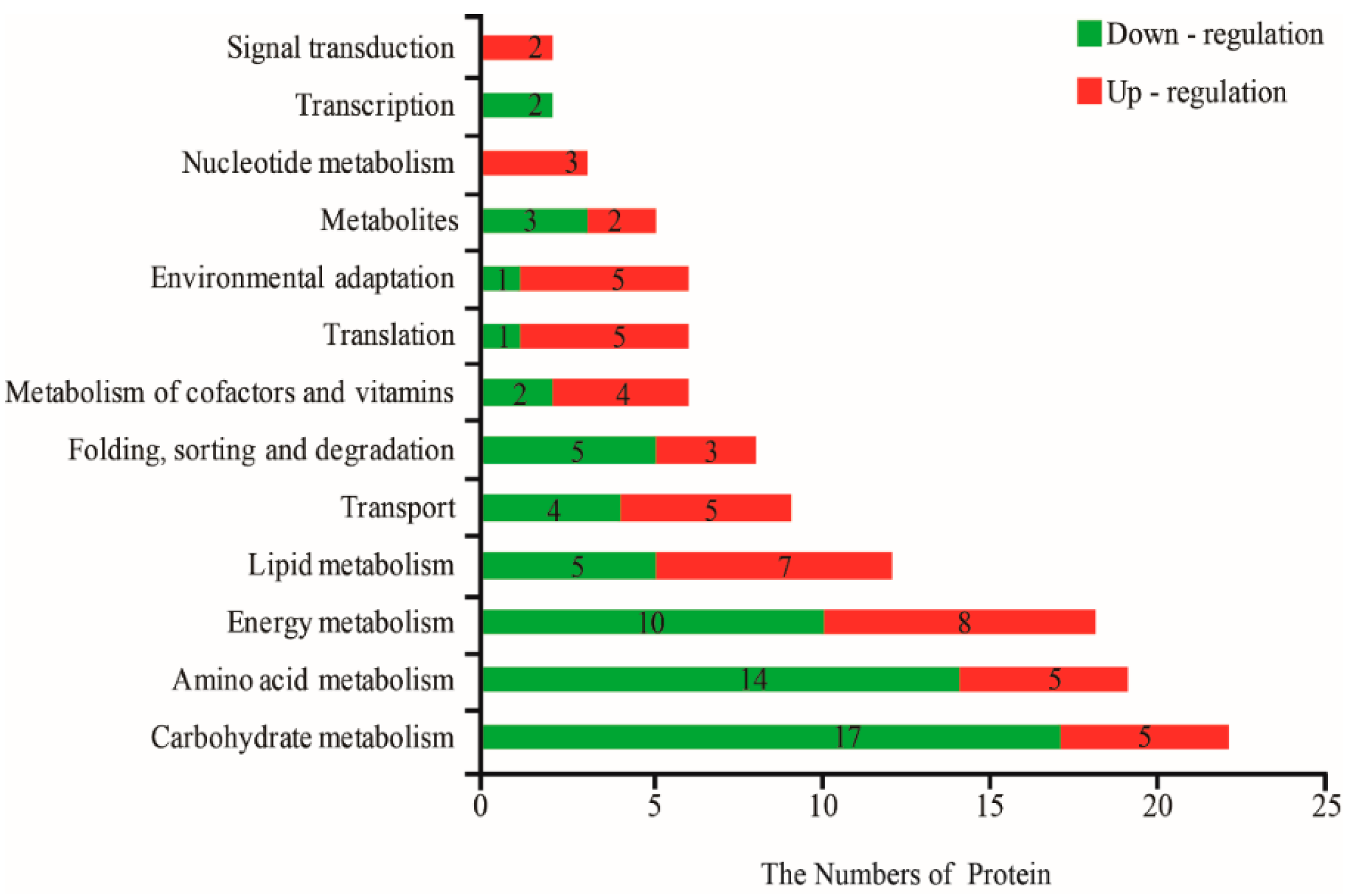
2.3. GO and KEGG Analysis of DAPs
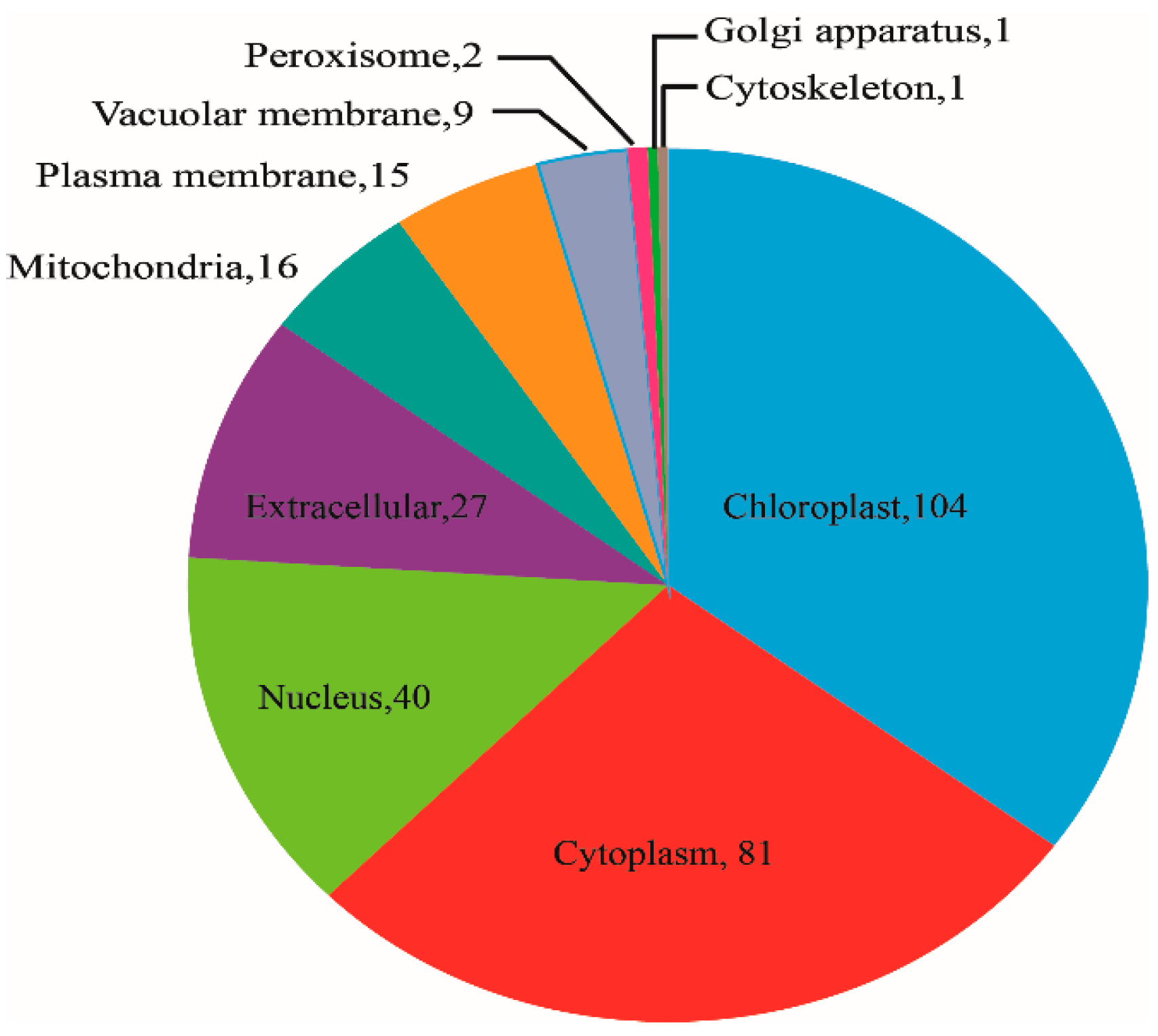
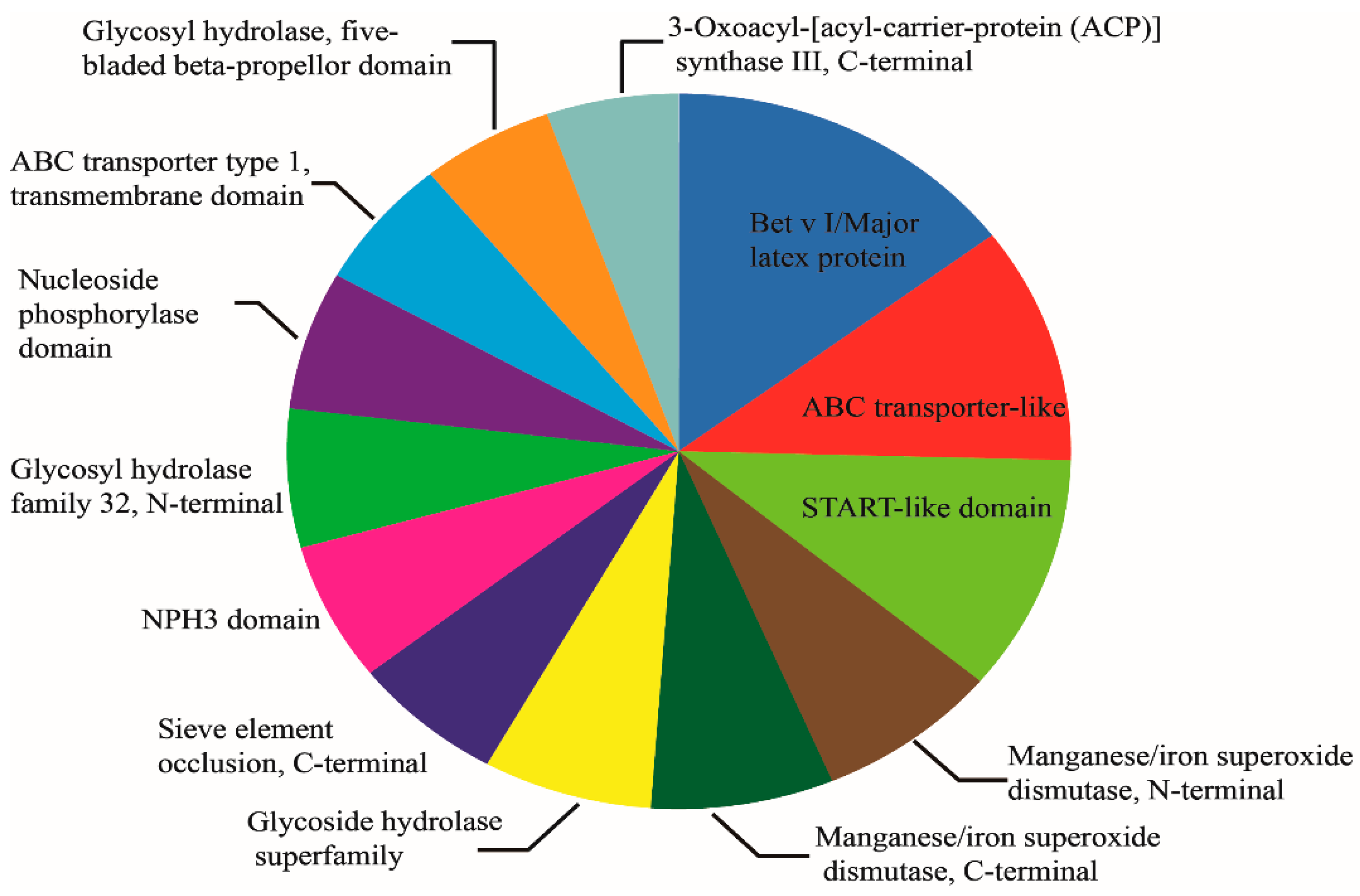
2.4. Subcellular Location and Domain Analysis of DAPs
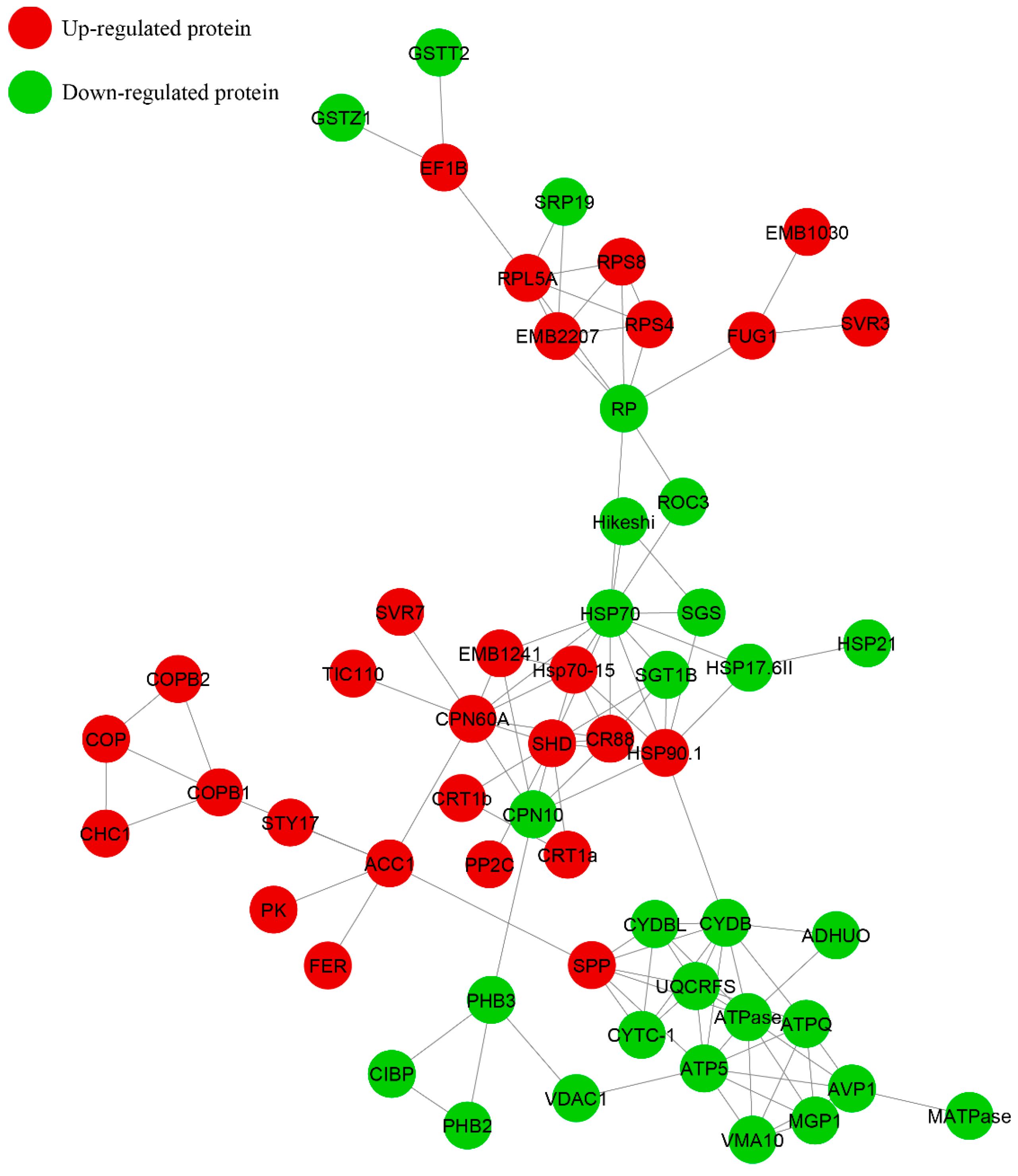
2.5. Interaction Network Analysis of the DAPs
2.6. PRM-MS Quantification of DAPs
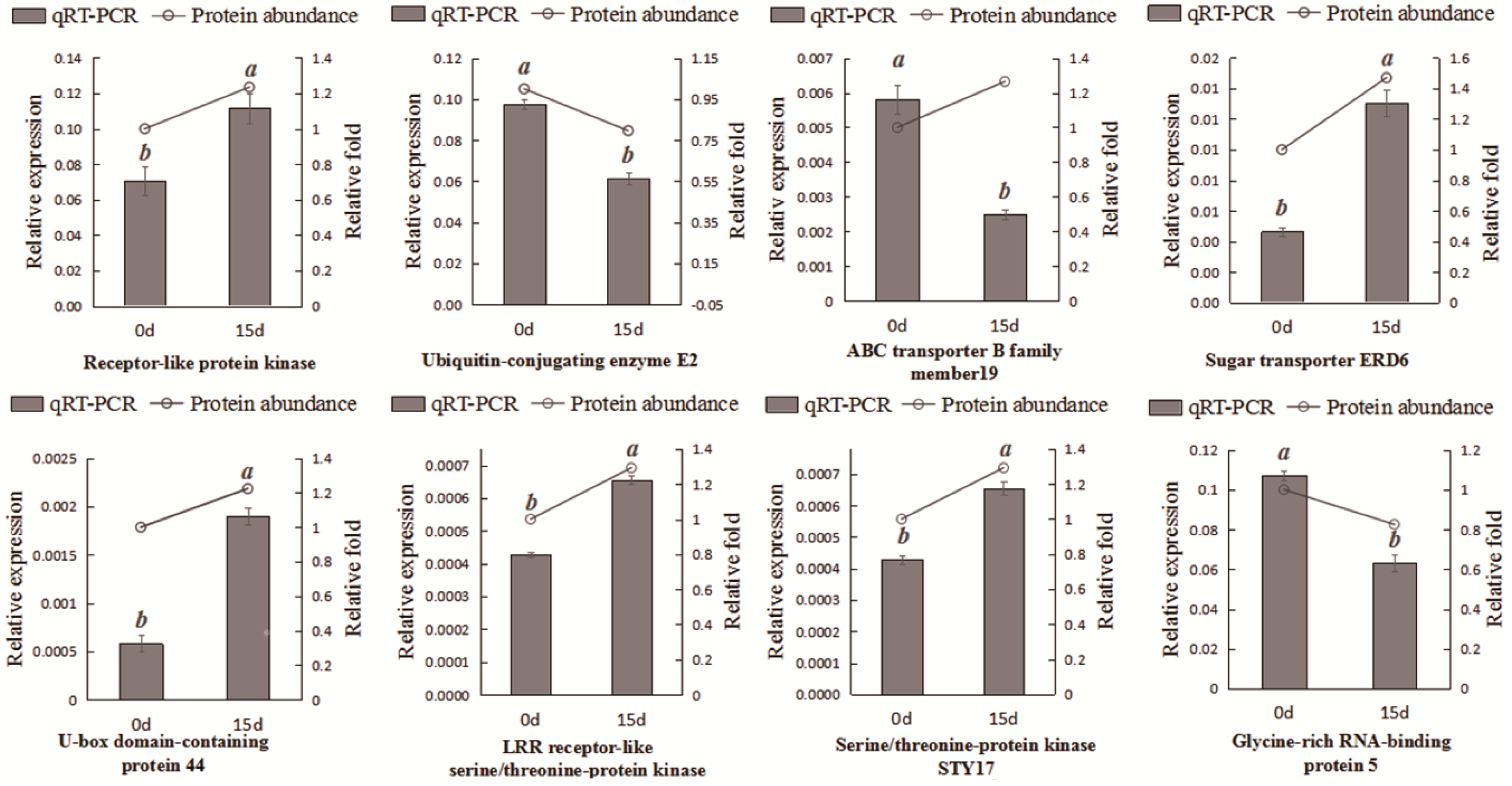
2.7. Complementation of the Proteomic Results via qRT-PCR
3. Discussion
3.1. LK Affected DAPs Involved in Transport Physiological Process
3.2. LK Affected DAPs Related to Carbohydrate and Energy Metabolism
3.3. LK Affected DAPs Related to Photosynthesis
3.4. LK Affected DAPs Related to Common Stress Responses
3.5. Comparative Analysis of Low-K+ Responses between Alligator Weed and Arabidopsis
4. Materials and Methods
4.1. Physiological Experiments of Alternanthera philoxeroides Stem
4.2. Protein Extraction
4.3. Trypsin Digestion and TMT Labelling
4.4. HPLC Fractionation
4.5. LC-MS/MS Analysis
4.6. Database Search
4.7. DAPs Functional Analysis
4.8. Parallel Reaction Monitoring PRM-MS Analysis
4.9. Quantitative Reverse Transcription PCR (qRT-PCR) Analysis
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Kanai, S.; Ohkura, K.; Adu-Gyamfi, J.J.; Mohapatra, P.K.; Nguyen, N.T.; Saneoka, H.; Fujita, K. Depression of sink activity precedes the inhibition of biomass production in tomato plants subjected to potassium deficiency stress. J. Exp. Bot. 2007, 58, 2917–2928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maathuis, F.J. Physiological functions of mineral macronutrients. Curr. Opin. Plant Biol. 2009, 12, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wu, W.H. Potassium transport and signaling in higher plants. Annu. Rev. Plant Biol. 2013, 64, 451–476. [Google Scholar] [CrossRef] [PubMed]
- Chérei, I.; Lefoulon, C.; Boeglin, M.; Sentenac, H. Molecular mechanisms involved in plant adaptation to low K+ availability. J. Exp. Bot. 2014, 65, 833–848. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.X.; Geng, Y.P.; Li, B.; Chen, J.K.; Yang, J. Genome-wide DNA methylation alterations of Alternanthera philoxeroidesin natural and manipulated habitats: Implications for epigenetic regulation of rapid responses to environmental fluctuation and phenotypic variation. Plant Cell Environ. 2010, 33, 1820–1827. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.Z.; Su, Y.H. Distinctive potassium-accumulation capability of alligator weed (Alternanthera philoxeroides) links to high-affinity potassium transport facilitated by K+-uptake systems. Weed Sci. 2013, 6, 77–84. [Google Scholar] [CrossRef]
- Li, L.Q.; Xu, L.; Wang, X.Y.; Pan, G.; Lu, L.M. De novo characterization of the alligator weed (Alternanthera philoxeroides) transcriptome illuminates gene expression under potassium deprivation. J. Genet. 2015, 94, 95–104. [Google Scholar] [CrossRef]
- Tomanek, L. Proteomics to study adaptations in marine organisms to environmental stress. J. Proteom. 2014, 105, 92–106. [Google Scholar] [CrossRef]
- Savitski, M.M.; Mathieson, T.; Zinn, N.; Sweetman, G.; Doce, C.; Becher, I.; Pachl, F.; Kuster, B.; Bantscheff, M. Measuring and managing ratio compression for accurate iTRAQ/TMT quantification. J. Proteome Res. 2013, 12, 3586–3598. [Google Scholar] [CrossRef]
- Zeng, J.B.; He, X.Y.; Quan, X.Y.; Cai, S.G.; Han, Y.; Nadira, U.A.; Zhang, G.P. Identification of the proteins associated with low potassium tolerance in cultivated and Tibetan wild barley. J. Proteom. 2015, 126, 1–11. [Google Scholar] [CrossRef]
- Zhang, Z.; Chao, M.; Wang, S.; Bu, J.; Tang, J.; Li, F. Proteome quantification of cotton xylem sap suggests the mechanisms of potassium-deficiency-induced changes in plant resistance to environmental stresses. Sci. Rep. 2016, 6, 21060–21075. [Google Scholar] [CrossRef]
- Li, G.Z.; Wu, Y.F.; Liu, G.Y.; Xiao, X.H.; Wang, P.F.; Gao, T. A Large-scale proteomics combined with transgenic experiments demonstrates an important role of jasmonic acid in potassium deficiency response in wheat and rice. Mol. Cell. Proteom. 2017, 16, 1889–1905. [Google Scholar] [CrossRef]
- Li, L.Q.; Liu, L.; Zhuo, W.; Chen, Q.; Hu, S.; Peng, S.; Wang, X.Y.; Lu, Y.F.; Lu, L.M. Physiological and quantitative proteomic analyses unraveling potassium deficiency stress response in alligator weed (Alternanthera philoxeroides L.) root. Plant Mol. Biol. 2018, 97, 265–278. [Google Scholar] [CrossRef] [PubMed]
- Theodoulou, F.L. Plant ABC transporters. Biochim. Biophys. Acta 2000, 1465, 79–103. [Google Scholar] [CrossRef] [Green Version]
- Ji, W.; Cong, R.; Li, S.; Li, R.; Qin, Z.W.; Li, Y.J.; Zhou, X.L.; Chen, S.; Li, J. Comparative proteomic analysis of soybean leaves and roots by iTRAQ provides insights into response mechanisms to short-term salt stress. Front. Plant Sci. 2016, 7, 573. [Google Scholar] [CrossRef] [PubMed]
- Ofori, P.A.; Geisler, M.; di Donato, M.; Pengchao, H.; Otagaki, S.; Matsumoto, S.; Shiratake, K. Tomato ATP-binding cassette transporter SlABCB4 is involved in auxin transport in the developing fruit. Plants 2018, 7, 65. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.X.; Tang, H.; Wang, E.T.; Chen, W.X. An ABC transporter is required for alkaline stress and potassium transport regulation in Sinorhizobium meliloti. FEMS Microbiol. 2009, 293, 35–41. [Google Scholar] [CrossRef]
- Wang, C.; Chen, H.F.; Hao, Q.N.; Sha, A.H.; Shan, Z.H.; Chen, L.M.; Zhou, R.; Zhi, H.J.; Zhou, X.A. Transcript profile of the response of two soybean genotypes to potassium deficiency. PLoS ONE 2012, 7, e39856. [Google Scholar] [CrossRef]
- Zhou, H.P.; Wang, C.W.; Tan, T.H.; Cai, J.Q.; He, J.X.; Lin, H.H. Patellin1 negatively modulates salt tolerance by regulating PM Na+/H+ antiport activity and cellular redox homeostasis in Arabidopsis. Plant Cell Physiol. 2018, 59, 1630–1642. [Google Scholar] [CrossRef]
- Zhang, B.; Tolstikov, V.; Turnbull, C.; Hicks, L.M.; Fiehn, O. Divergent metabolism and proteome suggest functional independence of dual phloem transport systems in cucurbits. PNAS 2010, 107, 13532–13537. [Google Scholar] [CrossRef]
- Ernst, A.M.; Jekat, S.B.; Zielonka, S.; Müller, B.; Neumann, U.; Rüping, B.; Twyman, R.M.; Krzyzanek, V.; Prüfer, D.; Noll, G.A. Sieve element occlusion (SEO) genes encode structural phloem proteins involved in wound sealing of the phloem. PNAS 2012, 109, 11084–11085. [Google Scholar] [CrossRef] [PubMed]
- Choi, W.G.; Masatsugu Toyota, M.; Kim, S.H.; Hilleary, R.; Gilroy, S. Salt stress-induced Ca2+ waves are associated with rapid, long-distance root-to-shoot signaling in plants. PNAS 2014, 111, 6497–6502. [Google Scholar] [CrossRef] [PubMed]
- Laohavisit, A.; Shang, Z.L.; Rubio, L.; Cuin, T.A.; Véry, A.A.; Wang, A.H.; Mortimer, J.C.; Macpherson, N.; Coxon, K.M.; Battey, N.H.; et al. Arabidopsis annexin1 mediates the radical-activated plasma membrane Ca2+- and K+-permeable conductance in root cells. Plant Cell 2012, 24, 1522–1533. [Google Scholar] [CrossRef]
- Wang, P.; Sun, Y.; Pei, Y.K.; Li, X.C.; Zhang, X.Y.; Li, F.G.; Hou, Y.X. GhSNAP33, a t-SNARE protein from Gossypium hirsutum, mediates resistance to Verticillium dahlia infection and tolerance to drought stress. Front. Plant Sci. 2018, 9, 896. [Google Scholar] [CrossRef] [PubMed]
- Zhou, A.M.; Ma, H.P.; Feng, S.; Gong, S.F.; Wang, J.G. A novel sugar transporter from Dianthus spiculifolius, DsSWEET12, affects sugar metabolism and confers osmotic and oxidative stress tolerance in Arabidopsis. Int. J. Mol. Sci. 2018, 19, 497. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.J.; Sun, M.H.; Kang, H.; Lu, J.; You, C.X.; Hao, Y.J. A CIPK protein kinase targets sucrose transporter MdSUT2.2 at Ser 254 for phosphorylation to enhance salt tolerance. Plant Cell Environ. 2018. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.J.; Sun, M.H.; Lu, J.; Kang, H.; You, C.X.; Hao, Y.J. An apple sucrose transporter MdSUT2.2 is a phosphorylation target for protein kinase MdCIPK22 in response to drought. Plant Biotechnol. J. 2018. [Google Scholar] [CrossRef] [PubMed]
- Drechsler, N.; Zheng, Y.; Bohner, A.; Nobmann, B.; Wirén, N.; Kunze, R.; Rausch, C. Nitrate-dependent control of shoot K homeostasis by the nitrate transporter1/peptide transporter family member NPF7.3/NRT1.5 and the stelar K+ outward rectifier SKOR in Arabidopsis. Plant Physiol. 2015, 169, 2832–2847. [Google Scholar]
- Li, H.; Yu, M.; Du, X.Q.; Wang, Z.F.; Wu, W.H.; Quintero, F.J.; Jin, X.H.; Li, H.D.; Wang, Y. NRT1.5/NPF7.3 functions as a proton-coupled H+/K+ antiporter for K+ loading into the xylem in Arabidopsis. Plant Cell 2017, 29, 2016–2026. [Google Scholar] [CrossRef]
- Lin, S.H.; Kuo, H.F.; Canivenc, G.; Lin, C.S.; Lepetit, M.; Hsu, P.K.; Tillard, P.; Lin, H.L.; Wang, Y.Y.; Tsai, C.B.; et al. Mutation of the Arabidopsis NRT1.5 nitrate transporter causes defective root-to-shoot nitrate transport. Plant Cell 2008, 20, 2514–2528. [Google Scholar] [CrossRef]
- Zhang, B.; Karnik, R.; Wang, Y.; Wallmeroth, N.; Blatt, M.R.; Grefen, C. The Arabidopsis R-SNARE VAMP721 interacts with KAT1 and KC1 K+ channels to moderate K+ current at the plasma membrane. Plant Cell 2015, 27, 1697–1717. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Yadav, N.S.; Tiwari, V.; Agarwal, P.K.; Jha, B. A SNARE-like superfamily protein SbSLSP from the Halophyte Salicornia brachiata confers salt and drought tolerance by maintaining membrane stability, K+/Na+ ratio, and antioxidant machinery. Front. Plant Sci. 2016, 7, 737. [Google Scholar] [CrossRef] [PubMed]
- Pittman, J.K. Multiple transport pathways for mediating intracellular pH homeostasis: The contribution of H+/ion exchangers. Front. Plant Sci. 2012, 3, 11. [Google Scholar] [CrossRef] [PubMed]
- Welchen, E.; Daniel, H.G. Cytochrome c, a hub linking energy, redox, stress and signaling pathways in mitochondria and other cell compartments. Physiol. Plant. 2016, 157, 310–321. [Google Scholar] [CrossRef]
- Chivasa, S.; Tome, D.F.; Hamilton, J.M.; Slabas, A.R. Proteomic analysis of extracellular ATP-regulated proteins identifies ATP synthase beta-subunit as a novel plant cell death regulator. Mol. Cell. Proteom. 2011, 10, 1074–1087. [Google Scholar] [CrossRef] [PubMed]
- Armengaud, P.; Sulpice, R.; Miller, A.J.; Stitt, M.; Amtmann, A.; Gibon, Y. Multilevel analysis of primary metabolism provides new insights into the role of potassium nutrition for glycolysis and nitrogen assimilation in Arabidopsis roots. Plant Physiol. 2009, 150, 772–785. [Google Scholar] [CrossRef] [PubMed]
- Guevara, D.R.; El-Kereamy, A.; Yaish, M.W.; Mei-Bi, Y.; Rothstein, S.J. Functional characterization of the rice UDP-glucose 4-epimerase 1, OsUGE1: A potential role in cell wall carbohydrate partitioning during limiting nitrogen conditions. PLoS ONE 2014, 9, e96158. [Google Scholar] [CrossRef] [PubMed]
- Reyes, T.H.; Scartazza, A.; Pompeiano, A.; Ciurli, A.; Lu, Y.; Guglielminetti, L.; Yamaguchi, J. Nitrate reductase modulation in response to changes in C/N balance and nitrogen source in Arabidopsis. Plant Cell Physiol. 2018, 59, 1248–1254. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Wang, H.; Yang, J.J.; Liu, X.; Liu, Z.R. Pyruvate kinase M2 regulates gene transcription by acting as a protein kinase. Mol. Cell 2012, 45, 598–609. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.G.; Pyo, Y.J.; Cho, J.W.; Cho, M.H. Comparative proteome analysis of differentially expressed proteins induced by K+ deficiency in Arabidopsis thaliana. Proteomics 2004, 4, 3549–3559. [Google Scholar] [CrossRef]
- Du, S.Y.; Zhang, X.F.; Lu, Z.; Xin, Q.; Wu, Z.; Jiang, T.; Lu, Y.; Wang, X.F.; Zhang, D.P. Roles of the different components of magnesium chelatase in abscisic acid signal transduction. Plant Mol. Biol. 2012, 80, 519–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, S.; Lu, K.; Wu, Z.; Jiang, S.C.; Yu, Y.T.; Bi, C.; Xin, Q.; Wang, X.F.; Zhang, D.P. A link between magnesium-chelatase H subunit and sucrose nonfermenting 1(SNF1)-related protein kinase SnRK2.6/OST1 in Arabidopsis guard cell signaling in response to abscisic acid. J. Exp. Bot. 2015, 66, 6355–6369. [Google Scholar] [CrossRef] [PubMed]
- Cai, B.B.; Li, Q.; Liu, F.J.; Bi, H.G.; Ai, X.Z. Decreasing fructose-1,6-bisphosphate aldolase activity reduces plant growth and tolerance to chilling stress in tomato seedlings. Physiol. Plant. 2018, 163, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Ciani, S.; Schachtman, D.P. A peroxidase contributes to ROS production during Arabidopsis root response to potassium deficiency. Mol. Plant 2010, 3, 420–427. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.Y.; Choi, Y.H.; Lee, J.I.; Kim, I.H.; Nam, T.J. Antioxidant activity of oxygen evolving enhancer protein 1 purified from Capsosiphon fulvescens. J. Food Sci. 2015, 80, H1412–H1417. [Google Scholar] [CrossRef] [PubMed]
- Piao, W.L.; Han, S.H.; Sakuraba, Y.; Paek, N.C. Rice 7-hydroxymethyl chlorophyll a reductase is involved in the promotion of chlorophyll degradation and modulates cell death signaling. Mol. Cells 2017, 40, 773–786. [Google Scholar]
- Pattanayak, G.K.; Tripathy, B.C. Overexpression of protochlorophyllide oxidoreductase C regulates oxidative stress in Arabidopsis. PLoS ONE 2011, 6, e26532. [Google Scholar] [CrossRef]
- Chen, J.Y.; Dai, X.F. Cloning and characterization of the Gossypium hirsutum major latex protein gene and functional analysis in Arabidopsis thaliana. Planta 2010, 231, 861–873. [Google Scholar] [CrossRef]
- Wang, Y.P.; Yang, L.; Chen, X.; Ye, T.T.; Zhong, B.; Liu, R.J.; Wu, Y.; Chan, Z.L. Major latex protein-like protein 43 (MLP43) functions as a positive regulator during abscisic acid responses and confers drought tolerance in Arabidopsis thaliana. J. Exp. Bot. 2016, 67, 421–434. [Google Scholar] [CrossRef]
- Zhang, C.M.; Shi, S.L. Physiological and proteomic responses of contrasting alfalfa (Medicago sativa L.) varieties to PEG-induced osmotic stress. Front. Plant Sci. 2018, 9, 242. [Google Scholar] [CrossRef]
- Kim, S.R.; Yang, J.; An, G. OsCpn60α1, encoding the plastid chaperonin 60α subunit, is essential for folding of rbcL. Mol. Cells 2013, 35, 402–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, Y.J.; Melencion, S.M.; Lee, E.S.; Park, J.H.; Alinapon, C.V.; Oh, H.T.; Yun, D.J.; Chi, Y.H.; Lee, S.Y. Universal stress protein exhibits a redox-dependent chaperone function in Arabidopsis and enhances plant tolerance to heat shock and oxidative Stress. Front. Plant Sci. 2015, 6, 1141. [Google Scholar] [CrossRef] [PubMed]
- Lan, W.Z.; Lee, S.C.; Che, Y.F.; Jiang, Y.Q.; Luan, S. Mechanistic analysis of AKT1 regulation by the CBL–CIPK–PP2CA interactions. Mol. Plant 2011, 4, 527–536. [Google Scholar] [CrossRef] [PubMed]
- Rashotte, A.M.; DeLong, A.; Muday, G.K. Genetic and chemical reductions in protein phosphatase activity alter auxin transport, gravity response, and lateral root growth. Plant Cell 2001, 13, 1683–1697. [Google Scholar] [CrossRef]
- Kim, J.H.; Nguyen, N.H.; Nguyen, N.T.; Hong, S.W.; Lee, H. Loss of all three calreticulins, CRT1, CRT2 and CRT3, causes enhanced sensitivity to water stress in Arabidopsis. Plant Cell Rep. 2013, 32, 1843–1853. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Choi, Y.J.; Kwon, C.; Yun, H.S. Endoplasmic reticulum stress-induced accumulation of VAMP721/722 requires CALRETICULIN 1 and CALRETICULIN 2 in Arabidopsis. J. Integr. Plant Biol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Karnik, R.; Waghmare, S.; Donald, N.; Blatt, M.R. VAMP721 conformations unmask an extended motif for K+ channel binding and gating control. Plant Physiol. 2017, 173, 536–551. [Google Scholar] [CrossRef] [PubMed]
- Morillo, S.A.; Tax, F.E. Functional analysis of receptor-like kinases in monocots and dicots. Curr. Opin. Plant Biol. 2006, 9, 460–469. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, J.S.; Chaudhuri, S.; Dey, N.; Pal, A. Functional characterization of a serine-threonine protein kinase from Bambusa balcooa that implicates in cellulose overproduction and superior quality fiber formation. BMC Plant Biol. 2013, 13, 128–145. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Zhang, G.; Wu, M. CLE peptide signaling and crosstalk with phytohormones and environmental stimuli. Front. Plant Sci. 2016, 6, 1211. [Google Scholar] [CrossRef] [PubMed]
- Bita, C.; Gerats, T. Plant tolerance to high temperature in a changing environment: Scientific fundamentals and production of heat stress-tolerant crops. Front Plant Sci. 2013, 4, 273. [Google Scholar] [CrossRef] [PubMed]
- Ciuzan, O.; Hancock, J.; Pamfil, D.; Wilson, I.; Ladomery, M. The evolutionarily conserved multifunctional glycine-rich RNA binding proteins play key roles in development and stress adaptation. Physiol. Plant. 2015, 153, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Lorkovic, Z.J. Role of plant RNA-binding proteins in development, stress response and genome organization. Trends Plant Sci. 2009, 14, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Kim, W.Y.; Kwak, K.J.; Oh, S.H.; Han, Y.S.; Kang, H. Glycine-rich RNA-binding proteins are functionally conserved in Arabidopsis thaliana and Oryza sativa during cold adaptation process. J. Exp. Bot. 2010, 61, 2317–2325. [Google Scholar] [CrossRef] [PubMed]
- Teng, K.; Tan, P.H.; Xiao, G.Z.; Han, L.B.; Chang, Z.H.; Chao, Y.H. Heterologous expression of a novel Zoysia japonica salt induced glycine-rich RNA-binding protein gene, ZjGRP, caused salt sensitivity in Arabidopsis. Plant Cell Rep. 2017, 36, 179–191. [Google Scholar] [CrossRef] [PubMed]
- Stone, S.L. The role of ubiquitin and the 26S proteasome in plant abiotic stress signaling. Front. Plant Sci. 2014, 5, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.A.; Chang, R.Z.; Qiu, L.J. Overexpression of soybean ubiquitin-conjugating enzyme gene GmUBC2 confers enhanced drought and salt tolerance through modulating abiotic stress-responsive gene expression in Arabidopsis. Plant Mol. Biol. 2010, 72, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Ahn, M.Y.; Oh, T.R.; Seo, D.H.; Kim, J.H.; Cho, N.H.; Kim, W.T. Arabidopsis group XIV ubiquitin-conjugating enzymes AtUBC32, AtUBC33, and AtUBC34 play negative roles in drought stress response. J. Plant Physiol. 2018, 230, 73–79. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Accession Number | Protein Annotation | Protein Score | Percentage of Protein Sequence Coverage % | Peptides Count | Number of Unique Peptides | Fold Change | p-Value |
|---|---|---|---|---|---|---|---|
| Gene.10143 | ABC transporter F family member 5 | 52.24 | 8.1 | 5 | 5 | 1.34 | 0.0425 |
| Gene.15326 | ABC transporter B family member 19 | 91.58 | 10 | 11 | 8 | 1.27 | 0.00104 |
| Gene.11683 | ABC transporter F family member 4 | 45.75 | 6 | 3 | 3 | 1.25 | 0.0222 |
| Gene.22137 | ABC transporter B family member 1 | 72.34 | 9.9 | 11 | 3 | 1.23 | 0.00932 |
| Gene.37843 | ABC transporter C family member 10 | 7.88 | 3.8 | 1 | 1 | 1.21 | 0.0478 |
| Gene.39686 | ABC transporter D family member 2 | 30.70 | 3.1 | 2 | 2 | 1.21 | 0.0289 |
| Gene.14866 | ABC transporter B family member 2 | 107.13 | 11.8 | 12 | 10 | 1.21 | 0.0108 |
| Gene.22512 | Protein sieve element occlusion | 30.17 | 5.9 | 4 | 4 | 1.29 | 0.00538 |
| Gene.1036 | Protein sieve element occlusion | 19.51 | 26.9 | 3 | 3 | 1.29 | 0.00456 |
| Gene.16899 | Protein sieve element occlusion | 323.31 | 35.3 | 31 | 29 | 1.27 | 0.000702 |
| Gene.29767 | Patellin-3 | 286.67 | 34.9 | 24 | 10 | 1.39 | 0.0226 |
| Gene.42322 | Patellin-3 | 25.04 | 25.7 | 4 | 4 | 1.34 | 0.0464 |
| Gene.25481 | Patellin-4 | 7.54 | 2.1 | 1 | 1 | 1.29 | 0.0105 |
| Gene.42892 | Patellin-5 | 12.17 | 8.9 | 3 | 2 | 1.85 | 0.0142 |
| Gene.22040 | Sugar transporter ERD6-like 6 | 46.34 | 3.8 | 1 | 1 | 1.47 | 0.000758 |
| Gene.19069 | Chloride channel protein CLC-b | 13.79 | 2.4 | 2 | 2 | 1.46 | 0.042 |
| Gene.44796 | Two pore calcium channel protein 1 | 80.37 | 7.5 | 5 | 5 | 1.39 | 0.0196 |
| Gene.41564 | Vesicle-associated membrane protein | 32.77 | 41.5 | 4 | 2 | 1.26 | 0.0172 |
| Gene.23148 | Transmembrane 9 superfamily member | 14.87 | 8.8 | 6 | 2 | 1.21 | 0.0113 |
| Gene.36797 | Kinesin-like protein KIN-UA | 9.03 | 1.8 | 1 | 1 | 1.21 | 0.0246 |
| Gene.1641 | Vacuolar membrane proton pump | 25.00 | 12.9 | 9 | 1 | 0.81 | 0.0441 |
| Gene.51508 | Chloroplastic lipocalin | 41.09 | 17.7 | 5 | 5 | 0.78 | 0.784 |
| Gene.38083 | Stem-specific protein TSJT1 | 48.56 | 18.5 | 4 | 4 | 0.78 | 0.0021 |
| Gene.14846 | Protein NRT1/PTR FAMILY 8.3 | 34.84 | 6.5 | 4 | 2 | 0.74 | 0.0466 |
| Gene.1247 | Syntaxin-61 | 6.42 | 4.5 | 1 | 1 | 0.78 | 0.0021 |
| Gene.1250 | Syntaxin-61 | 6.32 | 3.7 | 1 | 1 | 0.67 | 0.0248 |
| Gene.11391 | V-type proton ATPase subunit G | 155.18 | 62.7 | 8 | 8 | 0.7 | 0.0448 |
| Description | Change in TMT | p-Value in TMT | Change in RPM | p-Value in RPM |
|---|---|---|---|---|
| Sieve element occlusion | 1.27 | 0.000702 | 1.28 | 0.005 |
| Patellin-3 | 1.39 | 0.0226 | 1.49 | 0.03 |
| ATP synthase | 0.83 | 0.00086 | 0.74 | 0.0015 |
| NAD(P)H dehydrogenase | 0.81 | 0.00398 | 0.88 | 0.01 |
| Glycine-rich RNA-binding protein 5 | 0.83 | 0.0457 | 0.67 | 0.002 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, L.-Q.; Lyu, C.-C.; Li, J.-H.; Tong, Z.; Lu, Y.-F.; Wang, X.-Y.; Ni, S.; Yang, S.-M.; Zeng, F.-C.; Lu, L.-M. Physiological Analysis and Proteome Quantification of Alligator Weed Stems in Response to Potassium Deficiency Stress. Int. J. Mol. Sci. 2019, 20, 221. https://doi.org/10.3390/ijms20010221
Li L-Q, Lyu C-C, Li J-H, Tong Z, Lu Y-F, Wang X-Y, Ni S, Yang S-M, Zeng F-C, Lu L-M. Physiological Analysis and Proteome Quantification of Alligator Weed Stems in Response to Potassium Deficiency Stress. International Journal of Molecular Sciences. 2019; 20(1):221. https://doi.org/10.3390/ijms20010221
Chicago/Turabian StyleLi, Li-Qin, Cheng-Cheng Lyu, Jia-Hao Li, Zhu Tong, Yi-Fei Lu, Xi-Yao Wang, Su Ni, Shi-Min Yang, Fu-Chun Zeng, and Li-Ming Lu. 2019. "Physiological Analysis and Proteome Quantification of Alligator Weed Stems in Response to Potassium Deficiency Stress" International Journal of Molecular Sciences 20, no. 1: 221. https://doi.org/10.3390/ijms20010221
APA StyleLi, L.-Q., Lyu, C.-C., Li, J.-H., Tong, Z., Lu, Y.-F., Wang, X.-Y., Ni, S., Yang, S.-M., Zeng, F.-C., & Lu, L.-M. (2019). Physiological Analysis and Proteome Quantification of Alligator Weed Stems in Response to Potassium Deficiency Stress. International Journal of Molecular Sciences, 20(1), 221. https://doi.org/10.3390/ijms20010221