Integration Analysis of Small RNA and Degradome Sequencing Reveals MicroRNAs Responsive to Dickeya zeae in Resistant Rice

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Overview of miRNA Sequencing
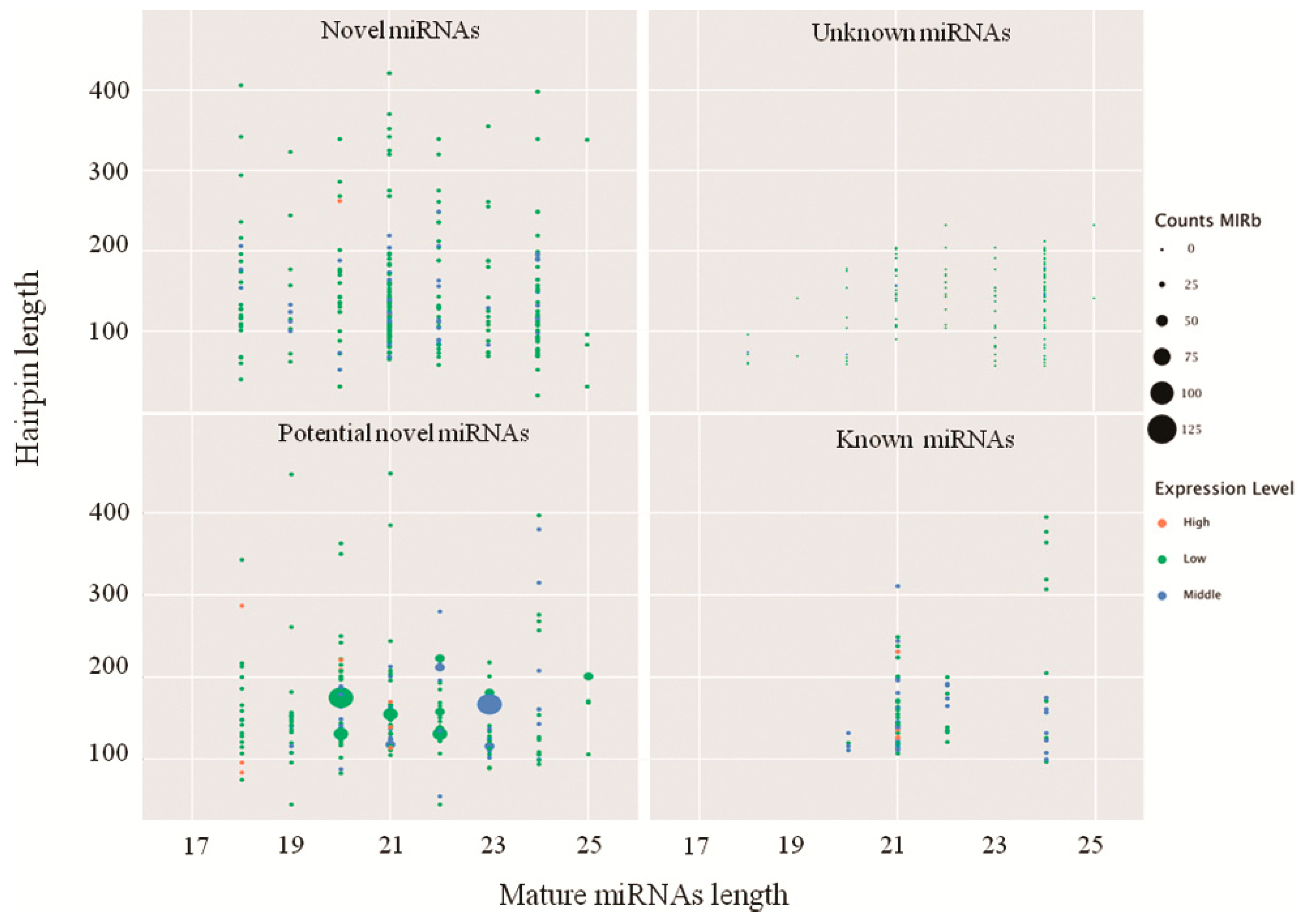
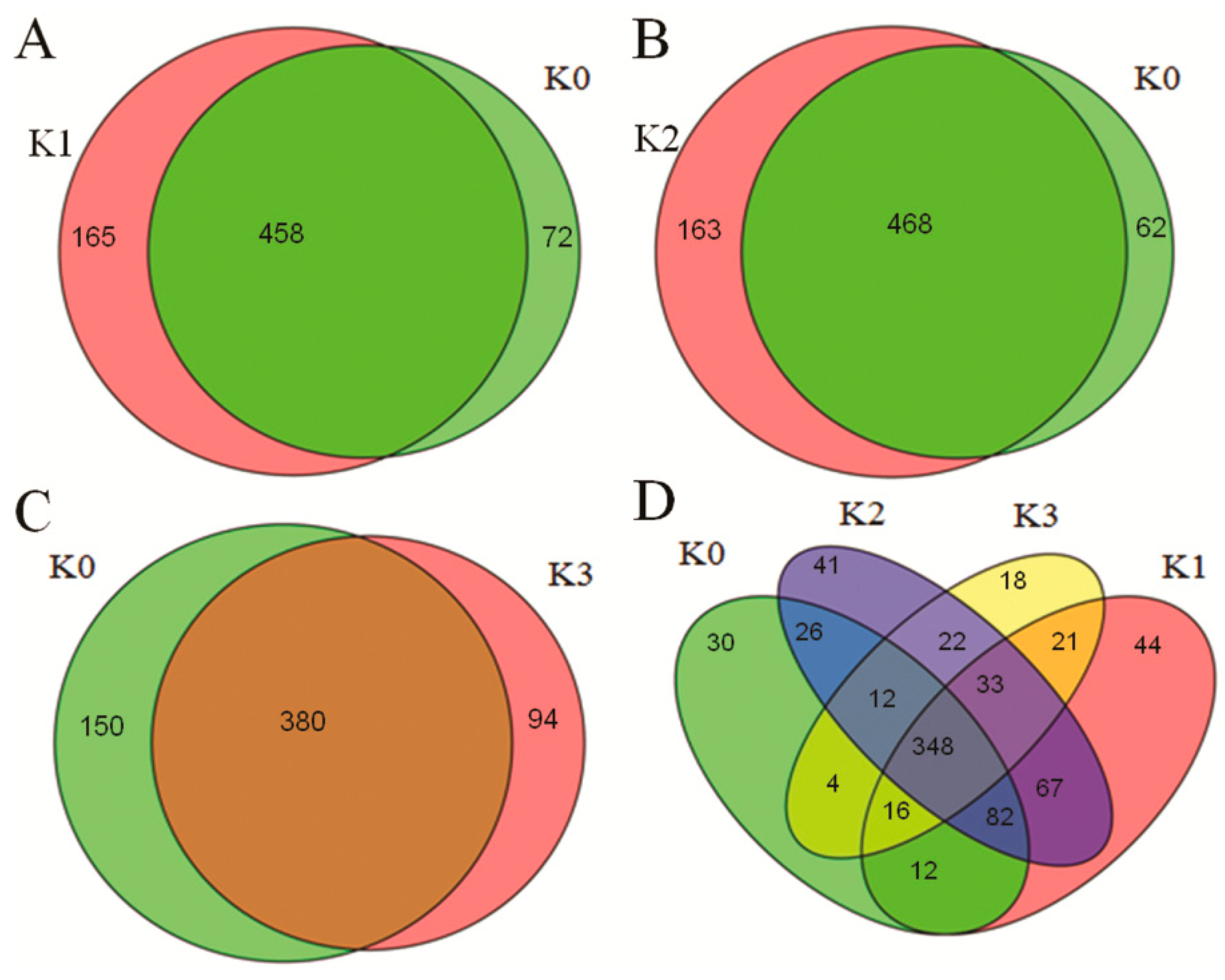
2.2. Identification of Known and Novel miRNAs
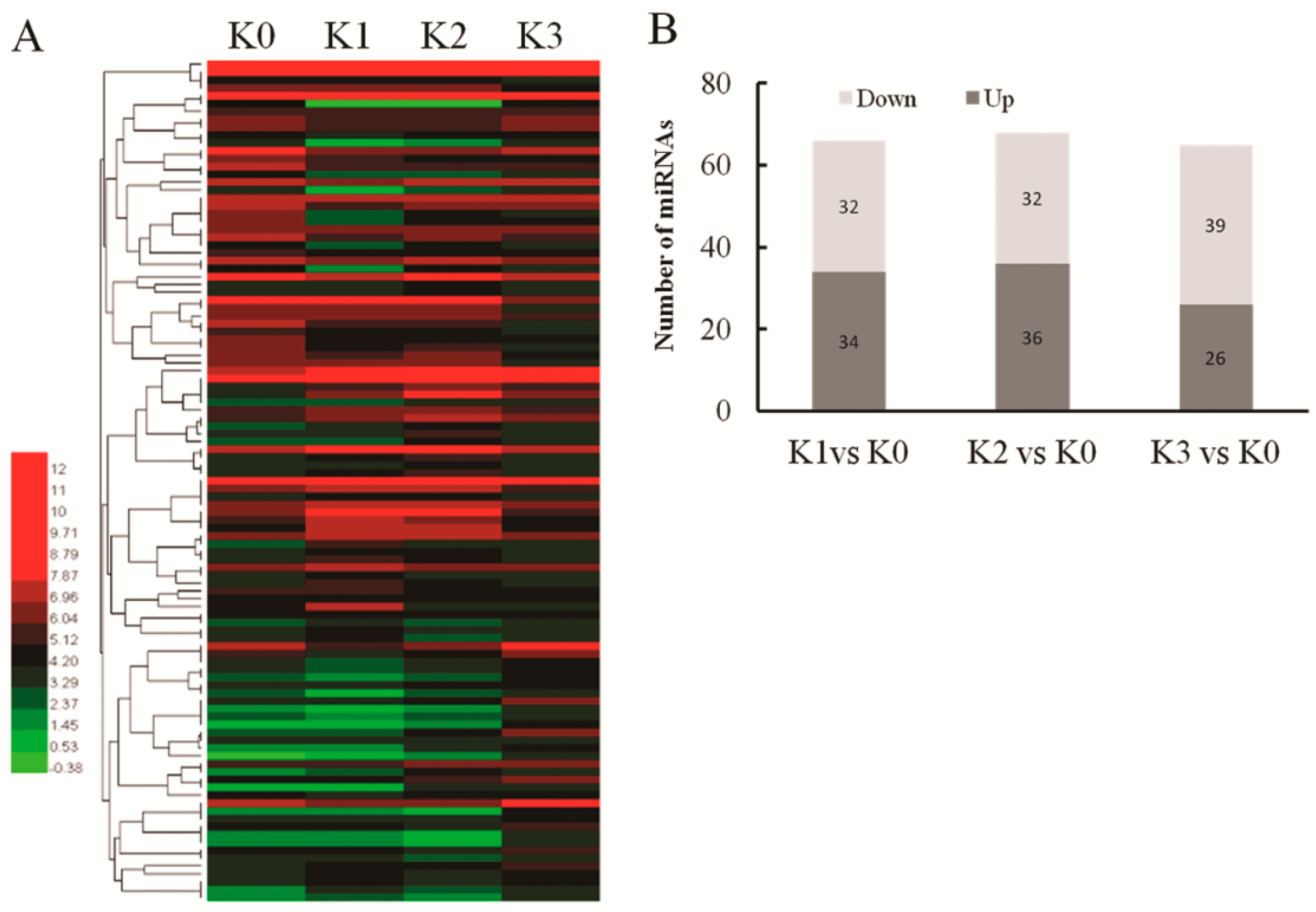
2.3. D. zeae-Responsive miRNAs in Resistant Rice
2.4. Target Prediction of miRNAs by Degradome Sequencing and Analysis
2.5. Functional Classification of Predicted Targets
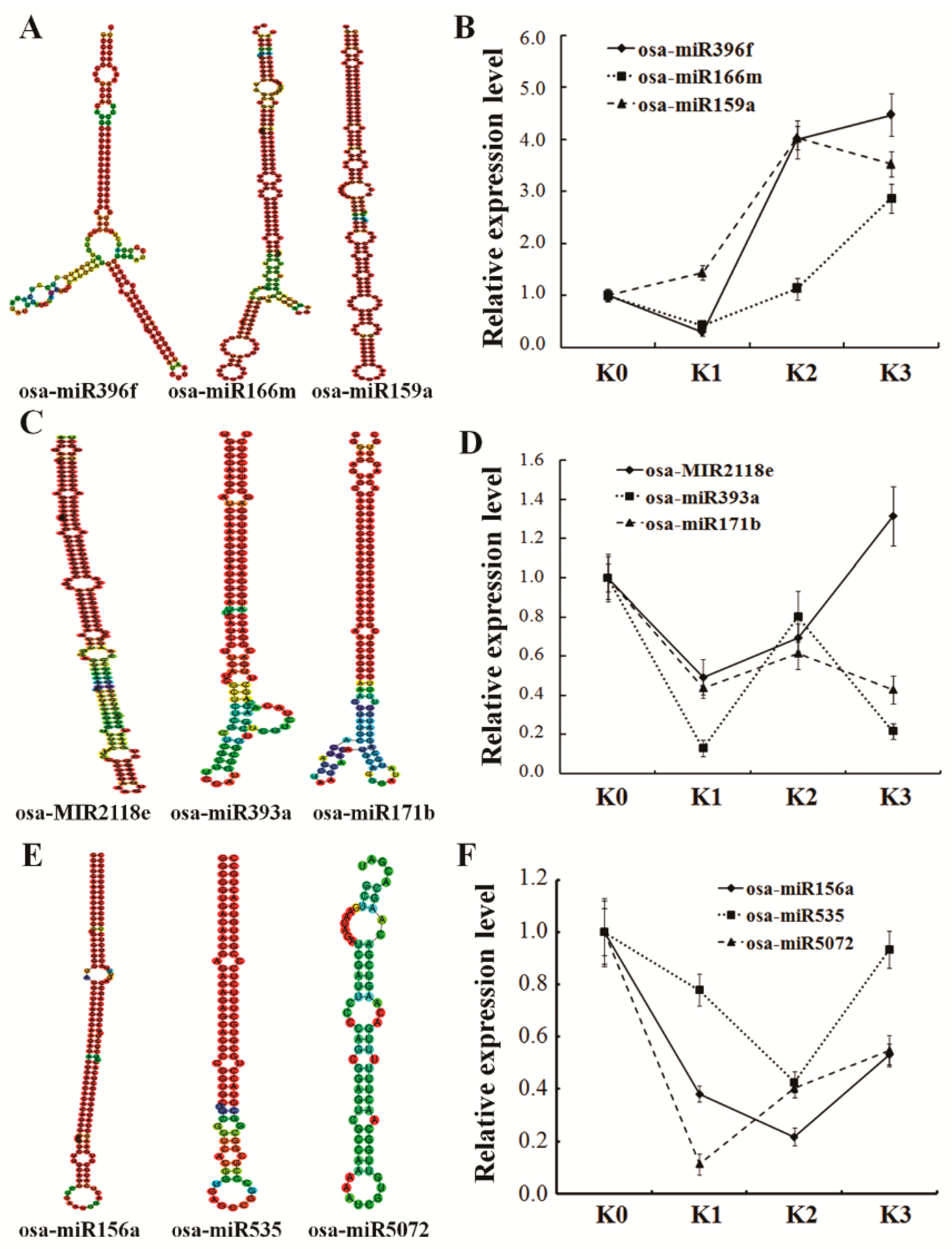
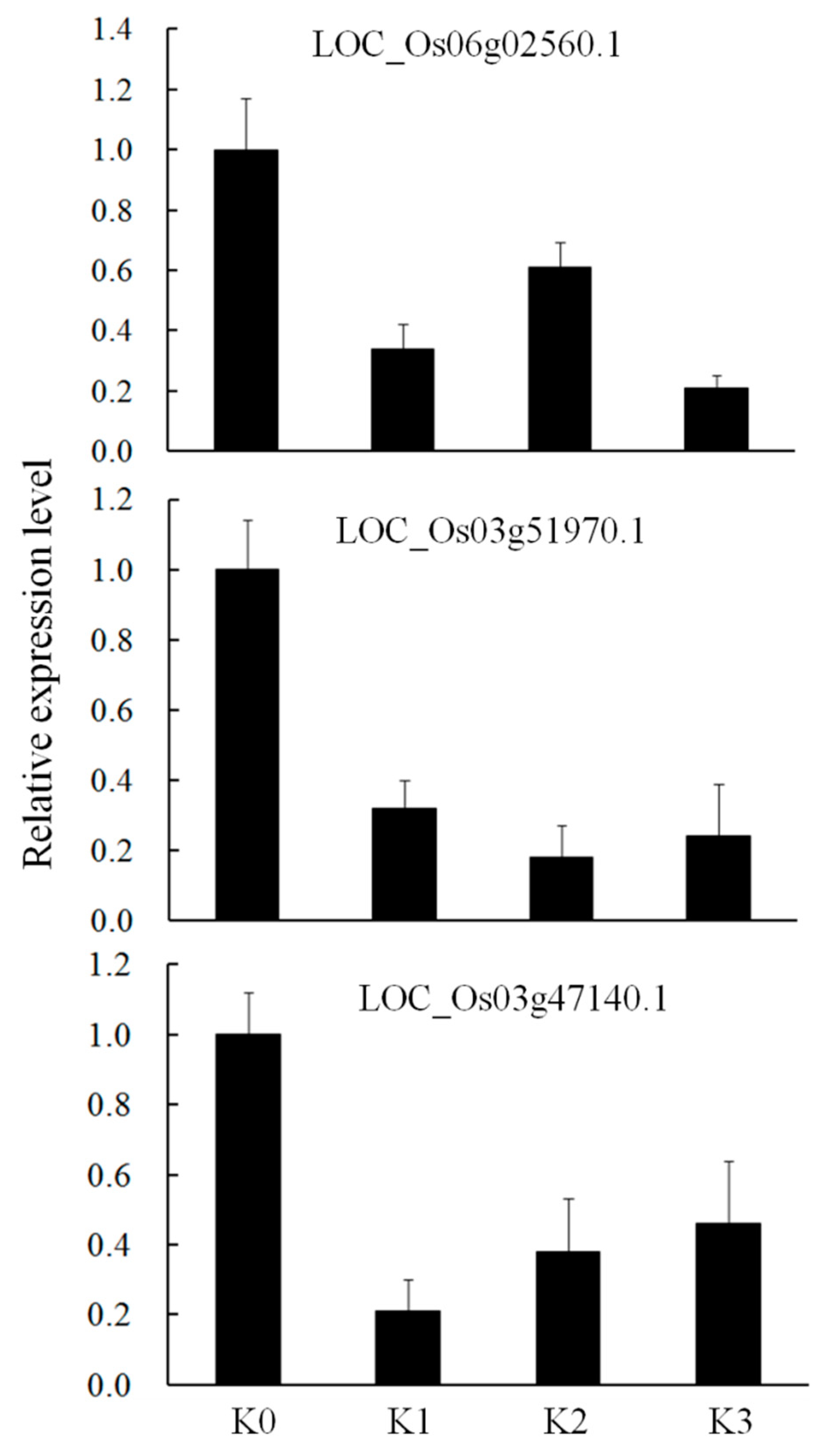
2.6. Co-expression Analysis and Validation of miRNA-Targets by qRT-PCR
2.7. Overexpression of Osa-miR396f Precursor Enhanced Rice Resistance to D. zeae
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Measurement of Resistance Reactions to D. zeae
4.2. Construction of Small RNA Library, Sequencing and Data Analysis
4.3. Identification and Function Analysis of Targets of miRNAs
4.4. Validation of Differentially Expressed miRNAs and Targets by qRT-PCR
4.5. Vector Construction and Rice Transformation
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Navarro, L.; Dunoyer, P.; Jay, F.; Arnold, B.; Dharmasiri, N.; Estelle, M.; Voinnet, O.; Jones, J.D. A plant miRNA contributes to antibacterial resistance by repressing auxin signaling. Science 2006, 312, 436–439. [Google Scholar] [CrossRef] [PubMed]
- Omidbakhshfard, M.A.; Proost, S.; Fujikura, U.; Mueller-Roeber, B. Growth-Regulating Factors (GRFs): A small transcription factor family with important functions in plant biology. Mol. Plant 2015, 16, 998–1010. [Google Scholar] [CrossRef] [PubMed]
- Baldrich, P.; San, S.B. MicroRNAs in rice innate immunity. Rice 2016, 9, 6. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.D.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellincampi, D.; Cervone, F.; Lionetti, V. Plant cell wall dynamics and wall-related susceptibility in plant-pathogen interactions. Front Plant Sci. 2014, 5, 228. [Google Scholar] [CrossRef]
- Guo, W.; Wang, C.; Zuo, Z.L.; Qiu, J.L. The roles of anion channels in Arabidopsis immunity. Plant Signal Behav. 2014, 9, e29230. [Google Scholar] [CrossRef]
- Liu, W.D.; Liu, J.; Triplett, L.; Leach, J.E.; Wang, G.L. Novel insights into rice innate immunity against bacterial and fungal pathogens. Annu. Rev. Phytopathol. 2014, 52, 213–241. [Google Scholar] [CrossRef]
- Gao, Q.M.; Zhu, S.; Kachroo, P.; Kachroo, A. Signal regulators of systemic acquired resistance. Front Plant Sci. 2015, 6, 228. [Google Scholar] [CrossRef]
- Li, F.; Pignatta, D.; Bendix, C.; Brunkard, J.O.; Cohn, M.M.; Tung, J.; Sun, H.; Kumar, P.; Baker, B. MicroRNA regulation of plant innate immune receptors. Proc. Natl. Acad. Sci. USA 2012, 109, 1790–1795. [Google Scholar] [CrossRef] [Green Version]
- Jones-Rhoades, M.W.; Bartel, D.P. Computational identification of plant microRNAs and their targets, including a stress-induced miRNA. Mol. Cell 2004, 14, 787–799. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, Q.Q.; Zhang, J.G.; Wu, L.; Qi, Y.J.; Zhou, J.M. Identification of microRNAs involved in pathogen-associated molecular pattern-triggered plant innate immunity. Plant Physiol. 2010, 152, 2222–2231. [Google Scholar] [CrossRef] [PubMed]
- Pumplin, N.; Voinnet, O. RNA silencing suppression by plant pathogens: Defence, counter-defence and counter-counter-defence. Nat. Rev. Microbiol. 2013, 11, 745–760. [Google Scholar] [CrossRef] [PubMed]
- Chow, H.T.; Ng, D.W. Regulation of miR163 and its targets in defense against Pseudomonas syringae in Arabidopsis thaliana. Sci. Rep. 2017, 7, 46433. [Google Scholar] [CrossRef] [PubMed]
- Eckardt, N.A. A microRNA cascade in plant defense. Plant Cell 2012, 24, 840. [Google Scholar] [CrossRef] [PubMed]
- Maudet, C.; Mano, M.; Eulalio, A. MicroRNAs in the interaction between host and bacterial pathogens. FEBS Lett. 2014, 588, 4140–4147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Huang, H. Roles of small RNAs in plant disease resistance. J. Integr. Plant Biol. 2014, 56, 962–970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, R.M.; He, L.Y.; He, J.Y.; Qin, P.G.; Wang, Y.R.; Deng, Q.M.; Yang, X.T.; Li, S.C.; Wang, S.Q.; Wang, W.M.; et al. Comprehensive analysis of microRNA-Seq and target mRNAs of rice sheath blight pathogen provides new insights into pathogenic regulatory mechanisms. DNA Res. 2016, 23, 415–425. [Google Scholar] [CrossRef] [PubMed]
- Campo, S.; Peris-Peris, C.; Siré, C.; Moreno, A.B.; Donaire, L.; Zytnicki, M.; Notredame, C.; Llave, C.; San, S.B. Identification of a novel microRNA (miRNA) from rice that targets an alternatively spliced transcript of the Nramp6 (Natural resistance-associated macrophage protein 6) gene involved in pathogen resistance. New Phytol. 2013, 199, 212–227. [Google Scholar] [CrossRef]
- Zhang, W.X.; Gao, S.; Zhou, X.; Chellappan, P.; Chen, Z.; Zhou, X.F.; Zhang, X.M.; Fromuth, N.; Coutino, G.; Coffey, M.; et al. Bacteria-responsive microRNAs regulate plant innate immunity by modulating plant hormone networks. Plant Mol. Biol. 2011, 75, 93–105. [Google Scholar] [CrossRef]
- Hong, H.M.; Liu, Y.Y.; Zhang, H.T.; Xiao, J.H.; Li, X.H.; Wang, S.P. Small RNAs and gene network in a durable disease resistance gene-mediated defense responses in rice. PLoS ONE 2015, 10, e0137360. [Google Scholar] [CrossRef]
- Li, Y.; Lu, Y.G.; Shi, Y.; Wu, L.; Xu, Y.J.; Huang, F.; Guo, X.Y.; Zhang, Y.; Fan, J.; Zhao, J.Q.; et al. Multiple rice microRNAs are involved in immunity against the blast fungus Magnaporthe oryzae. Plant Physiol. 2014, 164, 1077–1092. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Mei, J.; Wang, T.; Wang, C.C.; Zhang, W.L.; Yang, L. Identification and expression analysis of OsmiR1861k in rice leaves in response to Xanthomonas oryzae pv. oryzae. J. Gen. Plant Pathol. 2015, 81, 108–117. [Google Scholar] [CrossRef]
- Li, Y.; Zhao, S.L.; Li, J.L.; Hu, X.H.; Wang, H.; Cao, X.L.; Xu, Y.J.; Zhao, Z.X.; Xiao, Z.Y.; Yang, N.; et al. Osa-miR169 negatively regulates rice immunity against the blast fungus Magnaporthe oryzae. Front. Plant Sci. 2017, 17, 2. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.G.; Zhang, Q.; Wei, C.D. Advances in research of rice bacterial foot rot. Sci. Agric. Sin. 2013, 46, 2923–2931. [Google Scholar]
- Goto, M. Bacterial foot rot of rice caused by a strain of Erwinia chrysanthemi. Phytopathology 1979, 69, 213–216. [Google Scholar] [CrossRef]
- Yang, S.; Peng, Q.; Francisco, M.S.; Wang, Y.; Zeng, Q.; Yang, C.H. Type III secretion system genes of Dickeya dadantii 3937 are induced by plant phenolic acids. PLoS ONE 2008, 3, e2973. [Google Scholar] [CrossRef]
- Yang, S.; Peng, Q.; Zhang, Q.; Yi, X.; JaeChoi, C.; Reedy, R.M.; Charkowski, A.O.; Yang, C.H.; et al. Dynamic regulation of GacA in type III secretion, pectinase gene expression, pellicle formation, and pathogenicity of Dickeya dadantii (Erwinia chrysanthemi 3937). Mol. Plant-Microbe Interact. 2008, 21, 133–142. [Google Scholar] [CrossRef]
- Zhou, J.N.; Zhang, H.B.; Wu, J.; Liu, Q.G.; Xi, P.G.; Lee, J.; Liao, J.L.; Jiang, Z.D.; Zhang, L.H.; et al. A novel multidomain polyketide synthase is essential for zeamine antibiotics production and the virulence of Dickeya zeae. Mol. Plant-Microbe Interact. 2011, 24, 1156–1164. [Google Scholar] [CrossRef]
- Chen, X.F.; Wei, C.D.; Zhang, Q.; Liu, Q.G. Functional analysis of HrpX/HrpY in Dickeya zeae virulence. Sci. Agric. Sin. 2014, 47, 675–684. [Google Scholar]
- Li, W.Q.; Wang, J.; Fan, F.J.; Zhu, J.Y.; Wang, F.Q.; Yang, J.; Zhong, W.G. The isolation of rice bacterial foot rot disease pathogen in Jiangsu Province and identification of resistant rice varieties. Jiangsu Agric. Sci. 2014, 42, 139–142. [Google Scholar]
- Padmanabhan, C.; Zhang, X.M.; Jin, H.L. Host small RNAs are big contributors to plant innate immunity. Curr. Opin. Plant Biol. 2009, 12, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Sunkar, R.; Girke, T.; Jain, P.K.; Zhu, J.K. Cloning and characterization of microRNAs from rice. Plant Cell 2005, 17, 1397–1411. [Google Scholar] [CrossRef] [PubMed]
- Li, S.J.; Castillo-González, C.; Yu, B.; Zhang, X.R. The functions of plant small RNAs in development and in stress responses. Plant J. 2017, 90, 654–670. [Google Scholar] [CrossRef] [PubMed]
- Xin, M.M.; Wang, Y.; Yao, Y.Y.; Xie, C.J.; Peng, H.R.; Ni, Z.F.; Sun, Q.X. Diverse set of microRNAs are responsive to powdery mildew infection and heat stress in wheat (Triticum aestivum L.). BMC Plant Biol. 2010, 10, 123. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Ren, Y.Y.; Zhang, Y.Y.; Xu, J.C.; Zhang, Z.Y.; Wang, Y.W. Genome-wide profiling of novel and conserved Populus microRNAs involved in pathogen stress response by deep sequencing. Planta 2012, 235, 873–883. [Google Scholar] [CrossRef]
- Gupta, O.P.; Permar, V.; Koundal, V.; Singh, U.D.; Praveen, S. MicroRNA regulated defense responses in Triticum aestivum L. during Puccinia graminis f.sp. tritici infection. Mol. Biol. Rep. 2012, 39, 817–824. [Google Scholar] [CrossRef]
- Yin, Z.J.; Li, Y.; Han, X.L.; Shen, F.F. Genome-wide profiling of miRNAs and other small non-coding RNAs in the Verticillium dahliae-inoculated cotton roots. PLoS ONE 2012, 7, e35765. [Google Scholar] [CrossRef]
- Xu, D.G.; Mou, G.P.; Wang, K.; Zhou, G.H. MicroRNAs responding to southern rice black-streaked dwarf virus infection and their target genes associated with symptom development in rice. Virus Res. 2014, 190, 60–68. [Google Scholar] [CrossRef]
- Choi, D.; Kim, J.H.; Kende, H. Whole genome analysis of the Os-GRF gene family encoding plant-specific putative transcription activators in rice (Oryza sativa L.). Plant Cell Physiol. 2004, 45, 897–904. [Google Scholar] [CrossRef]
- Duan, P.G.; Ni, S.; Wang, J.M.; Zhang, B.L.; Xu, R.; Wang, Y.X.; Chen, H.Q.; Zhu, X.D.; Li, Y.H. Regulation of OsGRF4 by OsmiR396 controls grain size and yield in rice. Nat. Plants 2015, 2, 15203. [Google Scholar] [CrossRef]
- Gao, F.; Wang, K.; Liu, Y.; Chen, Y.P.; Chen, P.; Shi, Z.Y.; Luo, J.; Jiang, D.Q.; Fan, F.F.; Zhu, Y.G.; et al. Blocking miR396 increases rice yield by shaping inflorescence architecture. Nat. Plants 2015, 2, 15196. [Google Scholar] [CrossRef]
- Hu, J.; Wang, Y.X.; Fang, Y.X.; Zeng, L.J.; Xu, J.; Yu, H.P.; Shi, Z.Y.; Pan, J.J.; Zhang, D.; Kang, S.J.; et al. A rare allele of GS2 enhances grain size and grain yield in rice. Mol. Plant 2015, 8, 1455–1465. [Google Scholar] [CrossRef] [PubMed]
- Hewezi, T.; Maier, T.R.; Nettleton, D.; Baum, T.J. The Arabidopsis microRNA396-GRF1/GRF3 regulatory module acts as a developmental regulator in the reprogramming of root cells during cyst nematode infection. Plant Physiol. 2012, 159, 321–335. [Google Scholar] [CrossRef]
- Debernardi, J.M.; Rodriguez, R.E.; Mecchia, M.A.; Palatnik, J.F. Functional specialization of the plant miR396 regulatory network through distinct microRNA-target interactions. PLoS Genet. 2012, 8, e1002419. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.H.; Cao, Y.L.; Huang, L.L.; Zhao, J.; Xu, C.G.; Li, X.H.; Wang, S.P. Activation of the indole-3-acetic acid-amido synthetase GH3-8 suppresses expansin expression and promotes salicylate- and jasmonate-independent basal immunity in rice. Plant Cell 2008, 20, 228–240. [Google Scholar] [CrossRef] [PubMed]
- Li, W.Q.; Wang, F.Q.; Wang, J.; Fan, F.J.; Zhu, J.Y.; Yang, J.; Liu, F.Q.; Zhong, W.G. Overexpressing CYP71Z2 enhances resistance to bacterial blight by suppressing auxin biosynthesis in rice. PLoS ONE 2015, 10, e0119867. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.Y.; Rice, J.H.; Chen, N.N.; Baum, T.J.; Hewezi, T. Synchronization of developmental processes and defense signaling by growth regulating transcription factors. PLoS ONE 2014, 9, e98477. [Google Scholar] [CrossRef] [PubMed]
- Li, W.T.; Zhu, Z.W.; Chern, M.S.; Yin, J.J.; Yang, C.; Ran, L.; Cheng, M.P.; Wang, K.; Wang, J.; Zhou, X.G.; et al. A natural allele of a transcription factor in rice broad-spectrum blast resistance. Cell 2017, 170, 114–126. [Google Scholar] [CrossRef]
- Pu, X.M.; Zhou, J.N.; Lin, B.R.; Shen, H.F. First report of bacterial foot rot of rice caused by a Dickeya zeae in China. Plant Dis. 2012, 96, 1818. [Google Scholar] [CrossRef]
- Wu, L.; Zhang, Q.Q.; Zhou, H.Y.; Ni, F.R.; Wu, X.Y.; Qi, Y.J. Rice microRNA effector complexes and targets. Plant Cell 2009, 213, 3421–3435. [Google Scholar] [CrossRef]
- Taylor, R.S.; Tarver, J.E.; Hiscock, S.J.; Donoghue, P.C. Evolutionary history of plant microRNAs. Trends Plant Sci. 2014, 19, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.F.; Zheng, Y.; Addo-Quaye, C.; Zhang, L.; Saini, A.; Jagadeeswaran, G.; Axtell, M.J.; Zhang, W.; Sunkar, R. Transcriptome-wide identification of microRNA targets in rice. Plant J. 2010, 62, 742–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Y.; Li, Y.F.; Sunkar, R.; Zhang, W.X. SeqTar: An effective method for identifying microRNA guided cleavage sites from degradome of polyadenylated transcripts in plants. Nucleic Acids Res. 2012, 40, e28. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Gu, L.F.; Li, P.C.; Song, X.W.; Wei, L.Y.; Chen, Z.Y.; Cao, X.F. Degradome sequencing reveals endogenous small RNA targets in rice (Oryza sativa L. ssp. indica). Front. Biol. 2010, 5, 67–90. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sequence Type | K0 | K1 | K2 | K3 | ||||
|---|---|---|---|---|---|---|---|---|
| Total Counts | Unique Counts | Total Counts | Unique Counts | Total Counts | Unique Counts | Total Counts | Unique Counts | |
| Raw reads | 6,166,878 | 2,434,380 | 8,949,370 | 2,363,366 | 7,312,871 | 2,301,987 | 5,565,701 | 1,511,820 |
| 3ADT&length filter | 1,909,403 | 1,024,342 | 2,458,847 | 982,458 | 2,232,137 | 922,159 | 2,525,190 | 803,851 |
| Junk reads | 18,778 | 13,765 | 21,772 | 14,658 | 25,064 | 16,132 | 12,782 | 9,015 |
| Non-coding RNA | 559,796 | 169,225 | 600,364 | 129,071 | 625,544 | 132,521 | 359,131 | 91,856 |
| Rfam | 888,552 | 201,522 | 1,146,506 | 162,399 | 1,071,523 | 164,209 | 637,768 | 115,524 |
| mRNA | 1,279,886 | 170,257 | 2,684,763 | 234,424 | 1,778,770 | 204,532 | 1,146,100 | 122,153 |
| Repeats | 13,479 | 3,241 | 19,257 | 2,241 | 14,299 | 2,895 | 9,875 | 1,869 |
| rRNA | 641,748 | 127,477 | 857,965 | 102,851 | 749,376 | 98,155 | 491,401 | 73,325 |
| tRNA | 176,213 | 57,003 | 173,576 | 39,418 | 228,124 | 45,153 | 91,910 | 27,900 |
| snoRNA | 12,162 | 4,408 | 33,516 | 6,862 | 21,275 | 5,829 | 10,543 | 3,854 |
| snRNA | 9,586 | 2,845 | 24,758 | 3,638 | 14,329 | 3,441 | 7,713 | 2,389 |
| Other Rfam RNA | 48,843 | 9,789 | 56,691 | 9,630 | 58,419 | 11,631 | 36,201 | 8,056 |
| Clean reads | 2,385,536 | 1,053,550 | 3,164,367 | 1,000,514 | 2,637,057 | 1,023,748 | 1,512,623 | 483,076 |
| No. | Targets | Target Annotation | UP/Down | Cleavage Site |
|---|---|---|---|---|
| 1 | LOC_Os08g40900.1 | Auxin response factor 6 | up | 3466 |
| 2 | LOC_Os07g40290.1 | Auxin-responsive GH3 family protein | up | 2091 |
| 3 | LOC_Os06g39590.1 | AUX/IAA transcriptional regulator | up | 502 |
| 4 | LOC_Os03g51970.1 | Growth-regulating factor 1 | up | 430 |
| 5 | LOC_Os11g35030.1 | Growth-regulating factor 2 | up | 880 |
| 6 | LOC_Os02g53690.1 | Growth-regulating factor 5 | up | 581 |
| 7 | LOC_Os04g04330.1 | Leucine-rich repeat kinase | down | 3509 |
| 8 | LOC_Os10g33940.1 | Auxin response factor 16 | down | 2332 |
| 9 | LOC_Os12g10740.1 | Leucine-rich repeat protein kinase | down | 1219 |
| 10 | LOC_Os11g38440.1 | NB-ARC disease resistance protein | down | 3382 |
| 11 | LOC_Os04g59430.1 | Auxin response factor 16 | down | 1345 |
| 12 | LOC_Os04g38720.1 | NAC domain containing protein 80 | down | 811 |
| 13 | LOC_Os08g10080.1 | NAC domain containing protein 1 | down | 783 |
| 14 | LOC_Os12g40900.1 | AUX/IAA transcriptional regulator | down | 536 |
| 15 | LOC_Os07g42510.1 | Ethylene-responsive binding protein | down | 721 |
| 16 | LOC_Os03g08050.1 | GTP binding Elongation factor Tu protein | down | 969 |
| 17 | LOC_Os03g08020.1 | GTP binding Elongation factor Tu protein | down | 969 |
| 18 | LOC_Os03g08010.1 | GTP binding Elongation factor Tu protein | down | 969 |
| 19 | LOC_Os06g01620.1 | GRAS family transcription factor | down | 464 |
| 20 | LOC_Os02g44360.1 | GRAS family transcription factor | down | 1362 |
| 21 | LOC_Os02g09060.1 | Zinc finger family protein | down | 401 |
| 22 | LOC_Os08g41290.1 | Auxin-responsive family protein | down | 178 |
| 23 | LOC_Os04g51560.1 | WRKY DNA-binding protein 11 | down | 623 |
| 24 | LOC_Os10g01100.1 | Lectin protein kinase family protein | down | 2638 |
| 25 | LOC_Os02g50960.1 | Auxin efflux carrier family protein | down | 2109 |
| 26 | LOC_Os12g37760.1 | NB-ARC disease resistance protein | down | 3101 |
| 27 | LOC_Os07g33480.1 | Cytochrome P450 family 716 | down | 1606 |
| 28 | LOC_Os02g48080.1 | S-locus lectin protein kinase | down | 1135 |
| 29 | LOC_Os02g11980.1 | Leucine-rich repeat protein kinase | down | 3002 |
| 30 | LOC_Os12g43640.1 | Leucine-rich receptor-like protein kinase | down | 3203 |
| 31 | LOC_Os01g11340.1 | Cytochrome P450, family 710 | down | 1694 |
| 32 | LOC_Os02g14120.1 | Leucine-rich repeat protein kinase | down | 2050 |
| 33 | LOC_Os12g01510.1 | Leucine-rich repeat protein kinase | down | 2123 |
| 34 | LOC_Os12g37980.1 | Leucine-rich repeat transmembrane kinase | down | 2614 |
| No. | miR_name | Targets | Target Annotation | T0 h | T6–48 h | Up/Down | Cleavage Site |
|---|---|---|---|---|---|---|---|
| 1 | Osa-miR2118e- | LOC_Os03g06680.1 | Protein of unknown function 506 | 91.48 | 0 | down | 1608 |
| p5_1ss13TA | LOC_Os02g50960.1 | Auxin efflux carrier protein | 91.48 | 0 | down | 2109 | |
| 2 | Osa-miR393a | LOC_Os03g36080.1 | – | 1829.51 | 0 | down | 741 |
| LOC_Os05g05800.1 | F-box/RNI-like protein | 182.95 | 460.85 | up | 1708 | ||
| LOC_Os04g32460.1 | Auxin signaling F-box 2 | 1234.92 | 172.82 | down | 2235 | ||
| 3 | Osa-miR396f | LOC_Os02g53690.1 | Growth-regulating factor 5 | 0 | 460.85 | up | 581 |
| LOC_Os03g51970.1 | Growth-regulating factor 1 | 0 | 115.21 | up | 430 | ||
| LOC_Os11g35030.1 | Growth-regulating factor 2 | 0 | 115.21 | up | 880 | ||
| 4 | Osa-miR166m_R-1 | LOC_Os12g41860.1 | Leucine zipper family protein | 640.33 | 76.81 | down | 888 |
| LOC_Os03g43930.1 | Leucine zipper family protein | 548.85 | 76.81 | down | 966 | ||
| LOC_Os10g33960.1 | Leucine zipper family protein | 922.38 | 249.63 | down | 935 | ||
| LOC_Os03g01890.1 | Leucine zipper family protein | 899.51 | 249.63 | down | 1100 | ||
| 5 | Osa-miR171b | LOC_Os02g44370.1 | GRAS family transcription factor | 457.38 | 0 | down | 1537 |
| LOC_Os02g44360.1 | GRAS family transcription factor | 274.43 | 0 | down | 1362 | ||
| LOC_Os10g40390.1 | GRAS family transcription factor | 182.95 | 0 | down | 179 | ||
| LOC_Os06g01620.1 | GRAS family transcription factor | 1189.18 | 460.85 | down | 467 | ||
| LOC_Os04g46860.1 | GRAS family transcription factor | 2378.36 | 691.27 | down | 1337 | ||
| 6 | Osa-miR156a | LOC_Os08g41940.1 | SBP transcription factor | 457.38 | 0 | down | 1064 |
| LOC_Os01g69830.1 | SBP transcription factor | 182.95 | 115.21 | down | 1163 | ||
| LOC_Os02g04680.1 | SBP transcription factor | 457.38 | 57.61 | down | 1975 | ||
| LOC_Os11g30370.1 | SBP transcription factor | 91.48 | 0 | down | 1101 | ||
| LOC_Os09g31438.1 | SBP transcription factor | 91.48 | 0 | down | 819 | ||
| LOC_Os02g07780.1 | SBP transcription factor | 45.74 | 0 | down | 997 | ||
| LOC_Os06g49010.1 | SBP transcription factor | 45.74 | 0 | down | 1696 | ||
| LOC_Os06g45310.1 | SBP transcription factor | 274.43 | 115.21 | down | 864 | ||
| 7 | Osa-miR535-5p | LOC_Os06g45310.1 | Squamosa promoter-like 11 | 91.48 | 0 | down | 863 |
| 8 | Osa-miR159a_1R-3 | LOC_Os01g59660.1 | Myb domain protein 33 | 411.64 | 288.03 | down | 135 |
| LOC_Os10g05230.1 | RING/U-box superfamily protein | 274.43 | 0 | down | 1271 | ||
| LOC_Os12g10740.1 | Leucine-rich repeat protein kinase | 182.95 | 0 | down | 346 | ||
| LOC_Os06g40330.1 | Myb domain protein 65 | 182.95 | 0 | down | 1219 | ||
| LOC_Os01g47530.1 | MAP kinase 20 | 68.61 | 28.80 | down | 2333 | ||
| 9 | Osa-miR5072 _L-4 | LOC_Os05g07050.1 | Pre-mRNA processing splicing factor | 45.74 | 0 | down | 5043 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, W.; Jia, Y.; Liu, F.; Wang, F.; Fan, F.; Wang, J.; Zhu, J.; Xu, Y.; Zhong, W.; Yang, J. Integration Analysis of Small RNA and Degradome Sequencing Reveals MicroRNAs Responsive to Dickeya zeae in Resistant Rice. Int. J. Mol. Sci. 2019, 20, 222. https://doi.org/10.3390/ijms20010222
Li W, Jia Y, Liu F, Wang F, Fan F, Wang J, Zhu J, Xu Y, Zhong W, Yang J. Integration Analysis of Small RNA and Degradome Sequencing Reveals MicroRNAs Responsive to Dickeya zeae in Resistant Rice. International Journal of Molecular Sciences. 2019; 20(1):222. https://doi.org/10.3390/ijms20010222
Chicago/Turabian StyleLi, Wenqi, Yulin Jia, Fengquan Liu, Fangquan Wang, Fangjun Fan, Jun Wang, Jinyan Zhu, Yang Xu, Weigong Zhong, and Jie Yang. 2019. "Integration Analysis of Small RNA and Degradome Sequencing Reveals MicroRNAs Responsive to Dickeya zeae in Resistant Rice" International Journal of Molecular Sciences 20, no. 1: 222. https://doi.org/10.3390/ijms20010222
APA StyleLi, W., Jia, Y., Liu, F., Wang, F., Fan, F., Wang, J., Zhu, J., Xu, Y., Zhong, W., & Yang, J. (2019). Integration Analysis of Small RNA and Degradome Sequencing Reveals MicroRNAs Responsive to Dickeya zeae in Resistant Rice. International Journal of Molecular Sciences, 20(1), 222. https://doi.org/10.3390/ijms20010222