Increased Chromosome Aberrations in Cells Exposed Simultaneously to Simulated Microgravity and Radiation

 ,
, 
 ,
, 


Abstract
:1. Introduction
2. Results
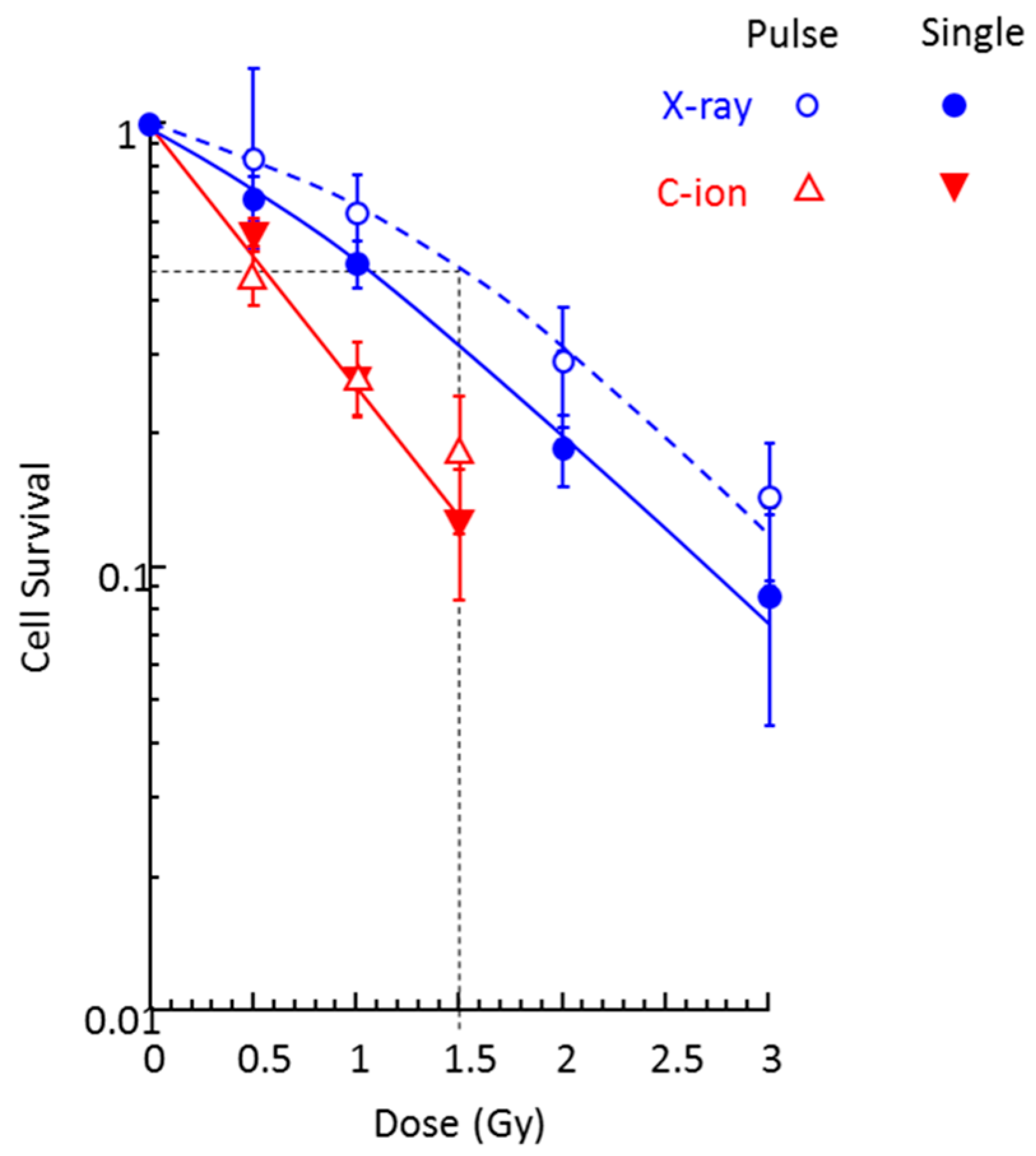
2.1. Cell Survival
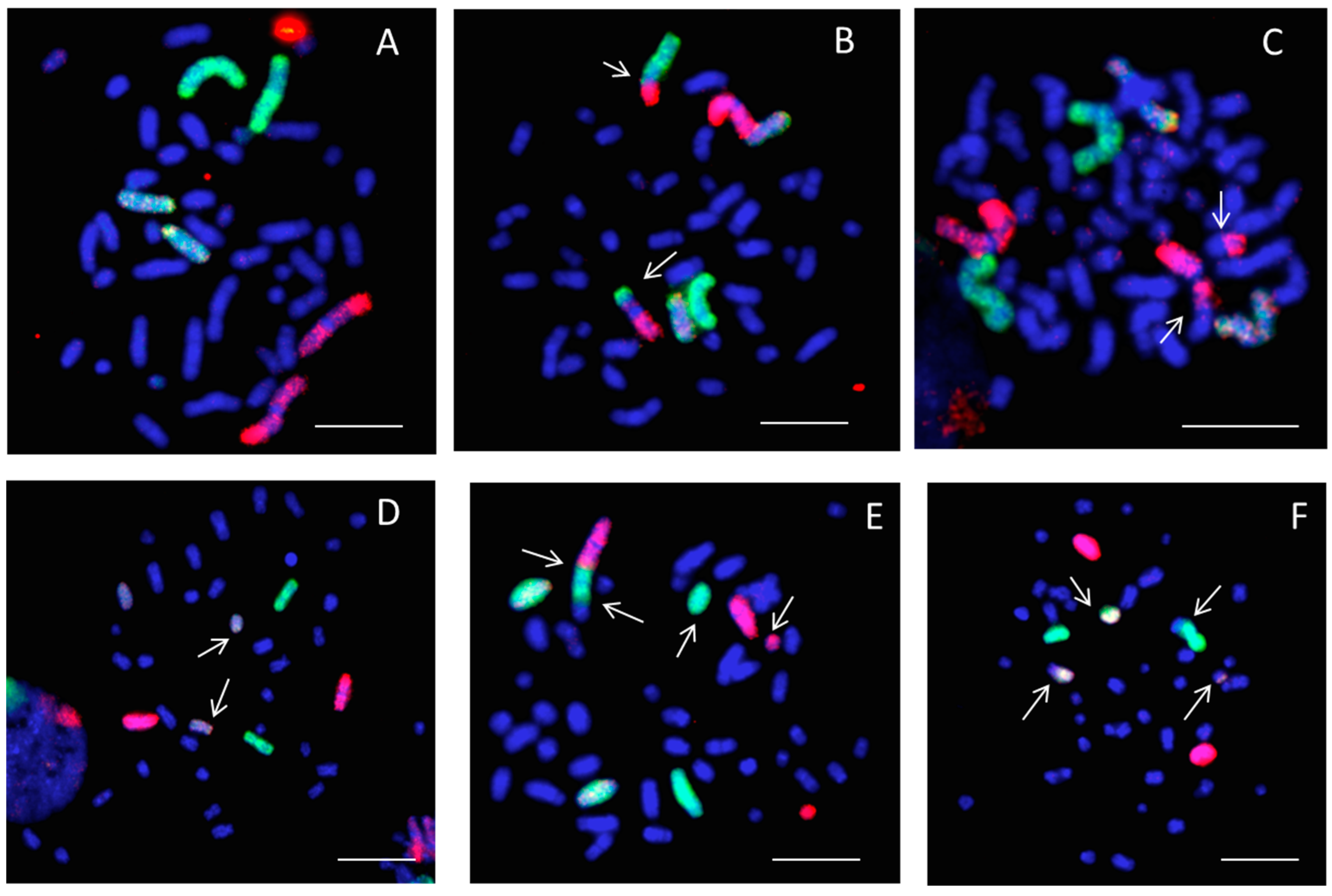
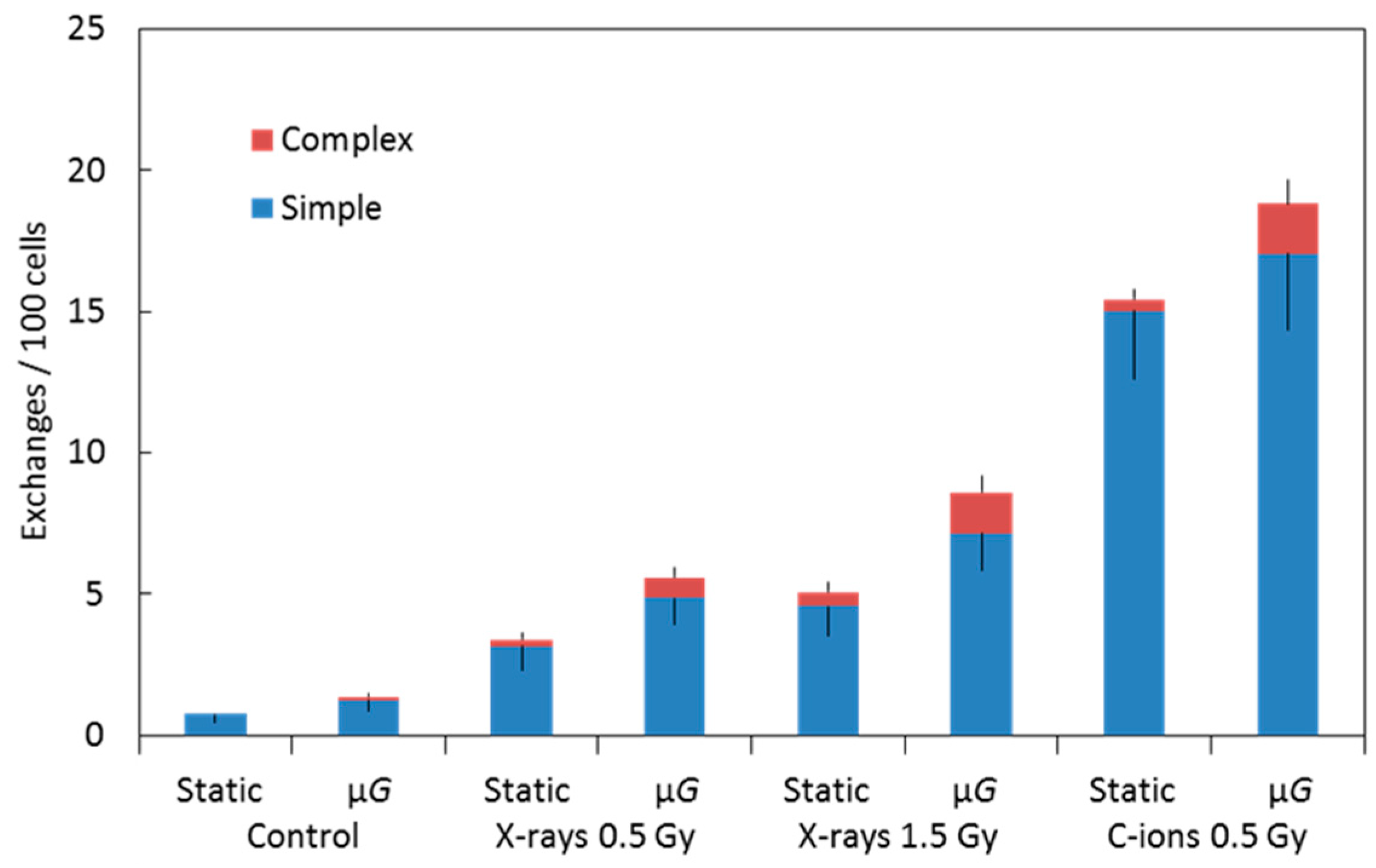
2.2. Chromosome Aberrations
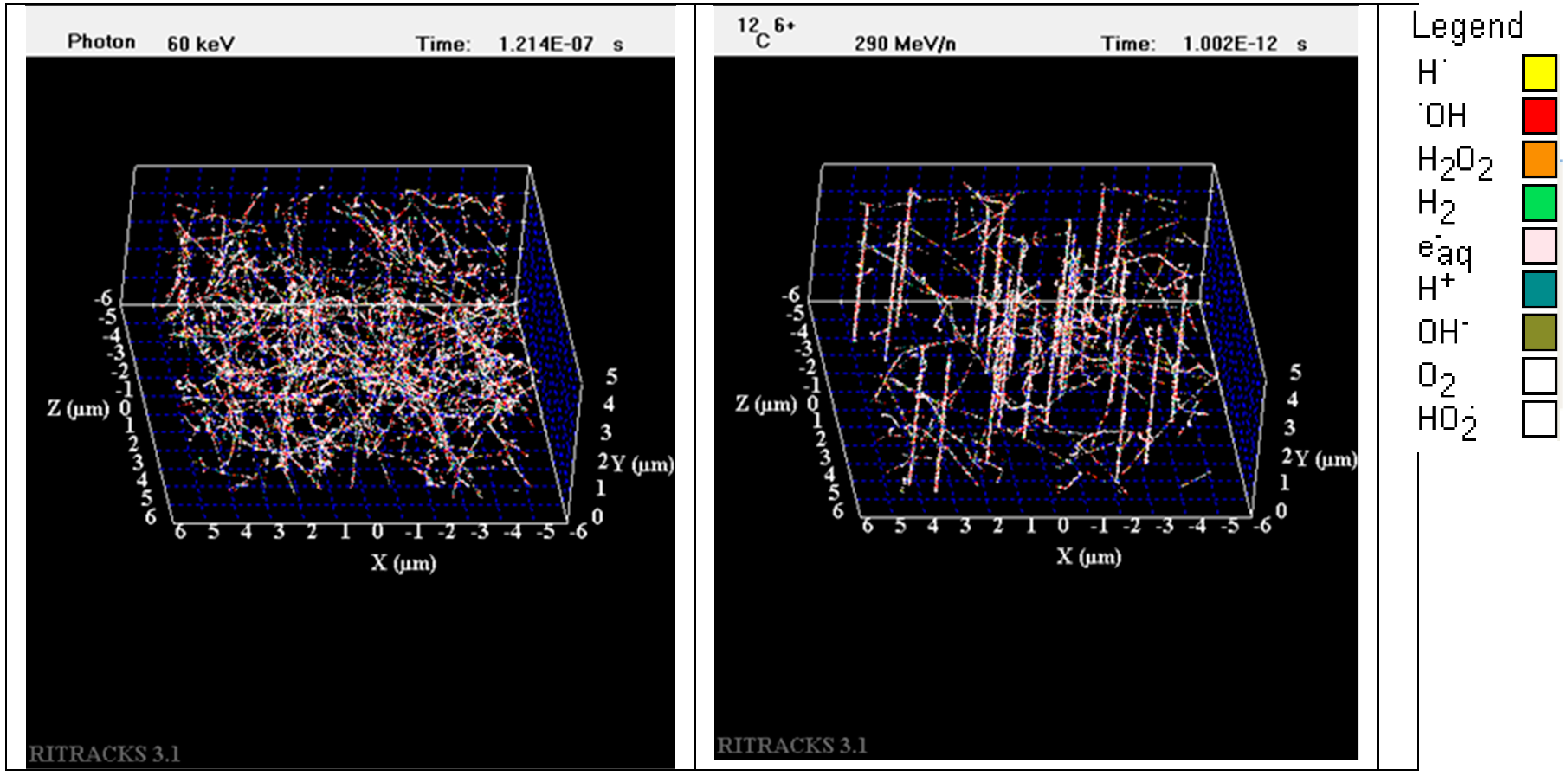
2.3. Simulation of Track Structure of Irradiation
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Synchronized Irradiation Systems under Simulated μG or 1G
4.3. Cell Survival Colony Formation Assay
4.4. Premature Chromosome Condensation (PCC)
4.5. Fluorescence In Situ Hybridization (FISH)
4.6. Simulations of Radiation Tracks
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 3D | 3-Dimensional |
| CA | Chromosome aberrations |
| CCD | Charge-coupled device |
| COI | CO2-independent medium |
| DSB | Double-strand breaks |
| FISH | Fluorescence in situ hybridization |
| GCR | Galactic cosmic rays |
| GHMC | Gunma University Heavy Ion Medical Center |
| HZE | High atomic number and energy |
| ISS | International Space Station |
| LD50 | Lethal dose 50 |
| LET | Linear Energy Transfer |
| MeV/n | Mega electron Volt per nucleon |
| PCC | Premature chromosome condensation |
| SEP | Solar energetic particle |
| SE | Standard error |
| SPE | Solar particle events |
| SOBP | Spread-out Bragg peak |
| TAMU | Texas A&M University |
| µG | Microgravity |
References
- West, J.B. Physiology in microgravity. J. Appl. Physiol. 2000, 89, 379–384. [Google Scholar] [CrossRef]
- Williams, D.; Kuipers, A.; Mukai, C.; Thirsk, R. Acclimation during space flight: Effects on human physiology. CMAJ 2009, 180, 1317–1323. [Google Scholar] [CrossRef]
- Hargens, A.R.; Vico, L. Long-duration bed rest as an analog to microgravity. J. Appl. Physiol. 2016, 120, 891–903. [Google Scholar] [CrossRef]
- Maier, J.A.M.; Cialdai, F.; Monici, M.; Morbidelli, L. The impact of microgravity and hypergravity on endothelial cells. BioMed Res. Int. 2015, 434803. [Google Scholar] [CrossRef]
- Louis, F.; Deroanne, C.; Nusgens, B.; Vico, L.; Guignandon, A. RhoGTPases as key player in mammalian cell adaptation to microgravity. BioMed Res. Int. 2015, 747693. [Google Scholar] [CrossRef]
- Rudimov, E.G.; Buravkova, L.B. Gravisensitivity of endothelial cells: The role of cytoskeleton and adhesion molecules. Hum. Physiol. 2016, 42, 687–693. [Google Scholar] [CrossRef]
- Mains, R.; Reynolds, S.; Baker, T.; Sato, K. A Researcher’s Guide to: Cellular Biology; NASA: Washington, DC, USA, 2015.
- Cucinotta, F.A.; Durante, M. Cancer risk from exposure to galactic cosmic rays: Implications for space exploration by human beings. Lancet Oncol. 2006, 7, 431–435. [Google Scholar] [CrossRef]
- Huff, J.; Carnell, L.; Blattnig, S.; Chappell, L.; George, K.; Lumpkins, S.; Simonsen, L.; Slaba, T.; Werneth, C. Evidence Report: Risk of Radiation Carcinogenesis; Johnson Space Center: Houston, TX, USA, 2016. [Google Scholar]
- Cucinotta, F.A.; Kim, M.H.; Ren, L. Evaluating shielding effectiveness for reducing space radiation cancer risks. Radiat. Meas. 2006, 41, 1173–1185. [Google Scholar] [CrossRef] [Green Version]
- Zeitlin, C.; Hassler, D.M.; Cucinotta, F.A.; Ehresmann, B.; Wimmer-Schweingruber, R.F.; Brinza, D.E.; Kang, S.; Weigle, G.; Böttcher, S.; Böhm, E.; et al. Measurements of energetic particle radiation in transit to Mars on the Mars Science Laboratory. Science 2013, 340, 1080–1084. [Google Scholar] [CrossRef]
- Cucinotta, F.A. Space radiation risk for astronauts on multiple International Space Station missions. PLoS ONE 2014, 9, e96099. [Google Scholar] [CrossRef]
- Cucinotta, F.A. Review of NASA approach to space radiation risk assessments for Mars exploration. Health Phys. 2015, 108, 131–142. [Google Scholar] [CrossRef]
- Cucinotta, F.A.; Schimmerling, W.; Wilson, J.W.; Peterson, L.E.; Saganti, P.; Badhwar, G.D.; Dicello, J.F. Space radiation cancer risks and uncertainties for Mars missions. Radiat. Res. 2001, 156, 682–688. [Google Scholar] [CrossRef]
- Jones, J.A.; McCarten, M.; Manuel, K.; Djojonegoro, B.; Murray, J.; Feiversen, A.; Wear, M. Cataract formation mechanism and risk in aviation and space crews. Aviat. Space Environ. Med. 2007, 78, 56–66. [Google Scholar]
- Cucinotta, F.A.; Alp, M.; Sulzman, F.M.; Wang, M. Space radiation risk to the central nervous system. Life Sci. Space Res. 2014, 2, 54–69. [Google Scholar] [CrossRef]
- Moreno-Villanueva, M.; Wong, M.; Lu, T.; Zhang, Y.; Wu, H. Interplay of space radiation and microgravity in DNA damage and DNA damage response. NPJ Microgravity 2017, 3, 14. [Google Scholar] [CrossRef]
- Horneck, G.; Rittberg, P.; Kozubek, S.; Baumstark-Khan, C.; Rink, H.; Schäfer, M.; Schmitz, C. The influence of microgravity on repair of radiation-induced DNA damage in bacteria and human fibroblasts. Radiat. Res. 1997, 147, 376–384. [Google Scholar] [CrossRef]
- Pross, H.D.; Casares, A.; Kiefer, J. Induction and repair of DNA double-strand breaks under irradiation and microgravity. Radiat. Res. 2000, 153, 521–525. [Google Scholar] [CrossRef]
- Takahashi, A.; Ohnishi, K.; Takahashi, S.; Masukawa, M.; Sekikawa, K.; Amano, T.; Nakano, T.; Nagaoka, S.; Ohnishi, T. The effects of microgravity on ligase activity in the repair of DNA double-strand breaks. Int. J. Radiat. Biol. 2000, 76, 783–788. [Google Scholar] [CrossRef]
- Bücker, H.; Horneck, G.; Reitz, G. Embryogenesis and organogenesis of Curausius morosus under spaceflight conditions. Naturwissenschaften 1986, 73, 433–434. [Google Scholar] [CrossRef]
- Gao, Y.; Xu, D.; Zhao, L.; Zhang, M.; Sun, Y. Effects of microgravity on DNA damage response in Caenorhabditis elegans during Shenzhou-8 spaceflight. Int. J. Radiat. Biol. 2015, 91, 531–539. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Kikuchi, M.; Nagaoka, S.; Watanabe, H. Recovery of Deinococcus radiodurans from radiation damage was enhanced under microgravity. Biol. Sci. Space 1996, 10, 97–101. [Google Scholar] [CrossRef]
- Wuest, S.L.; Richard, S.; Kopp, S.; Grimm, D.; Egli, M. Simulated microgravity: Critical review on the use of random positioning machines for mammalian cell culture. BioMed Res. Int. 2015, 971474. [Google Scholar] [CrossRef]
- Indo, H.P.; Tomiyoshi, T.; Suenaga, S.; Tomita, K.; Suzuki, H.; Masuda, D.; Terada, M.; Ishioka, N.; Gusev, O.; Cornette, R.; et al. MnSOD downregulation induced by extremely low 0.1 mGy single and fractionated X-rays and microgravity treatment in human neuroblastoma cell line, NB-1. J. Clin. Biochem. Nutr. 2015, 57, 98–104. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Sun, Q.; Xu, W.; Li, F.; Li, H.; Lu, J.; Wu, L.; Wu, Y.; Liu, M.; Bian, P. Modulation of modeled microgravity on radiation-induced bystander effects in Arabidopsis thaliana. Mutat. Res. 2015, 773, 27–36. [Google Scholar] [CrossRef]
- Ikeda, H.; Souda, H.; Puspitasari, A.; Held, K.D.; Hidema, J.; Nikawa, T.; Yoshida, Y.; Kanai, T.; Takahashi, A. Development and performance evaluation of a three-dimensional clinostat synchronized heavy-ion irradiation system. Life Sci. Space Res. 2017, 12, 51–60. [Google Scholar] [CrossRef]
- Tashiro, M.; Ishii, T.; Koya, J.; Okada, R.; Kurosawa, Y.; Arai, K.; Abe, S.; Ohashi, Y.; Shimada, H.; Yusa, K.; et al. Technical approach to individualized respiratory-gated carbon-ion therapy for mobile organs. Radiol. Phys. Technol. 2013, 6, 356–366. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, H.; Souda, H.; Puspitasari, A.; Held, K.D.; Hidema, J.; Nikawa, T.; Yoshida, Y.; Kanai, T.; Takahashi, A. A new system for three-dimensional clinostat synchronized X-irradiation with high-speed shutter for space radiation research. Biol. Sci. Space 2016, 30, 8–16. [Google Scholar] [CrossRef]
- Harada-Sukeno, A.; Kohno, S.; Nakano, R.; Hirasaka, K.; Higashibata, A.; Yamazaki, T.; Ishioka, N.; Suzuki, H.; Shimazu, T.; Maita, A.; et al. Myo Lab: A JAXA cell biology experiment in “Kibo (JEM)” of the International Space Station. Biol. Sci. Space 2009, 23, 189–193. [Google Scholar] [CrossRef]
- Tanigawa, N.; Yano, S.; Higashibata, A.; Tsuchiya, Y.; Tanii, H.; Ando, N.; Kubota, K.; Nagase, M. Development and validation of a closed chamber for cell culture experiments in space. Biol. Sci. Space 2014, 28, 1–5. [Google Scholar] [CrossRef]
- George, K.; Durante, M.; Wu, H.; Willingham, V.; Badhwar, G.; Cucinotta, F.A. Chromosome aberrations in the blood lymphocytes of astronauts after space flight. Radiat. Res. 2001, 156, 731–738. [Google Scholar] [CrossRef]
- George, K.; Willingham, V.; Cucinotta, F.A. Stability of chromosome aberrations in the blood lymphocytes of astronauts measured after space flight by FISH chromosome painting. Radiat. Res. 2005, 164, 474–480. [Google Scholar] [CrossRef]
- George, K.; Rhone, J.; Beitman, A.; Cucinotta, F.A. Cytogenetic damage in the blood lymphocytes of astronauts: Effect of repeat long-duration space missions. Mutat. Res. 2013, 756, 165–169. [Google Scholar] [CrossRef]
- Alpen, E.L.; Powers-Risius, P.; Curtis, S.B.; DeGuzman, R. Tumorigenic potential of high-Z, high-LET charged particle radiations. Radiat. Res. 1993, 88, 132–143. [Google Scholar] [CrossRef]
- Bonassi, S.; Norppa, H.; Ceppi, M.; Strömberg, U.; Vermeulen, R.; Znaor, A.; Cebulska-Wasilewska, A.; Fabianova, E.; Fucic, A.; Gundy, S.; et al. Chromosomal aberration frequency in lymphocytes predicts the risk of cancer: Results from a pooled cohort study of 22358 subjects in 11 countries. Carcinogenesis 2008, 29, 1178–1183. [Google Scholar] [CrossRef]
- Weil, M.M.; Bedford, J.S.; Bielefeldt-Ohmann, H.; Ray, A.F.; Gernik, P.C.; Ehrhart, E.J.; Fallgren, C.M.; Hailu, F.; Battaglia, C.L.; Charles, B.; et al. Incidence of acute myeloid leukemia and hepatocellular carcinoma in mice irradiated with 1 GeV/nucleon 56Fe ions. Radiat. Res. 2009, 172, 213–219. [Google Scholar] [CrossRef]
- Cucinotta, F.A.; Chappell, L.J. Non-targeted effects and the dose response for heavy ion tumor formation. Mutat. Res. 2010, 687, 49–53. [Google Scholar] [CrossRef]
- George, K.; Wu, H.; Willingham, V.; Furusawa, Y.; Kawata, T.; Cucinotta, F.A. High- and low-LET induced chromosome damage in human lymphocytes; a time course of aberrations in metaphase and interphase. Int. J. Radiat. Biol. 2001, 77, 175–183. [Google Scholar] [CrossRef]
- George, K.; Cucinotta, F.A. The influence of shielding on the biological effectiveness of accelerated particles for the induction of chromosome damage. Adv. Space Res. 2007, 39, 1076–1081. [Google Scholar] [CrossRef]
- Loucas, B.D.; Durante, M.; Bailey, S.M.; Cornforth, M.N. Chromosome damage in human cells by c rays, a particles and heavy ions: Track interactions in basic dose-response relationships. Radiat. Res. 2013, 179, 9–20. [Google Scholar] [CrossRef]
- George, K.; Willingham, V.; Wu, H.; Gridley, D.; Nelson, G.; Cucinotta, F.A. Chromosome aberrations in human lymphocytes induced by 250 MeV protons: Effects of dose, dose rate and shielding. Adv. Space Res. 2002, 30, 891–899. [Google Scholar] [CrossRef]
- George, K.; Durante, M.; Willingham, V.; Wu, H.; Yang, T.C.; Cucinotta, F.A. Biological effectiveness of accelerated particles for the induction of chromosome damage measured in metaphase and interphase in human lymphocytes. Radiat. Res. 2003, 160, 425–435. [Google Scholar] [CrossRef]
- Durante, M.; Cucinotta, F.A. Heavy ion carcinogenesis and human space exploration. Nat. Rev. Cancer 2008, 8, 465–472. [Google Scholar] [CrossRef] [Green Version]
- Manti, L.; Durante, M.; Cirrone, G.A.P.; Grossi, G.; Lattuada, M.; Pugliese, M.; Sabini, M.G.; Scampoli, P.; Valastro, L.; Gialanella, G. Modeled microgravity does not modify the yield of chromosome aberrations induced by high-energy protons in human lymphocytes. Int. J. Radiat. Biol. 2005, 81, 147–155. [Google Scholar] [CrossRef]
- Mosesso, P.; Schuber, M.; Seibt, D.; Schmitz, C.; Fiore, M.; Schinoppi, A.; Penna, S.; Palitti, F. X-ray-induced chromosome aberrations in human lymphocytes in vitro are potentiated under simulated microgravity conditions (Clinostat). Phys. Med. 2001, 17, 264–266. [Google Scholar]
- Canova, S.; Fiorasi, F.; Mognato, M.; Glifalconi, M.; Reddi, E.; Russo, A.; Celotti, L. “Modeled microgravity” affects cell response to ionizing radiation and increases genomic damage. Radiat. Res. 2005, 163, 191–199. [Google Scholar] [CrossRef]
- Mognato, M.; Celotti, L. Modeled microgravity affects cell survival and HPRT mutant frequency, but not the expression of DNA repair genes in human lymphocytes irradiated with ionizing radiation. Mutat. Res. 2005, 578, 417–429. [Google Scholar] [CrossRef]
- Lobrich, M.; Cooper, P.K.; Rydberg, B. Non-random distribution of DNA double-strand breaks induced by particle irradiation. Int. J. Radiat. Biol. 1996, 70, 493–503. [Google Scholar] [CrossRef]
- Hada, M.; Georgakilas, A.G. Formation of clustered DNA damage after high-LET irradiations: A review. J. Radiat. Res. 2008, 49, 203–210. [Google Scholar] [CrossRef]
- Singleton, B.K.; Griffin, C.S.; Thacker, J. Clustered DNA damage leads to complex genetic changes in irradiated human cells. Cancer Res. 2002, 62, 6263–6269. [Google Scholar]
- Tsao, D.; Kalogerinis, P.; Tabrizi, I.; Dingfelder, M.; Stewart, R.D.; Georgakilas, A.G. Induction and processing of oxidative clustered DNA lesions in 56Fe-ion-irradiated human monocytes. Radiat. Res. 2007, 168, 87–97. [Google Scholar] [CrossRef]
- Hair, J.M.; Terzoudi, G.I.; Hatzi, V.I.; Lehockey, K.A.; Srivastava, D.; Wang, W.; Pantelias, G.E.; Georgakilas, A.G. BRCA1 role in the mitigation of radiotoxicity and chromosomal instability through repair of clustered DNA lesions. Chem. Biol. Interact. 2010, 188, 350–358. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Hada, M.; Wu, H. Interphase chromosome conformation and chromatin-chromatin interactions in human epithelial cells cultured under different gravity conditions. In Proceedings of the 30th Annual Meeting of the American Society for Gravitational and Space Research, Pasadena, CA, USA, 23–26 October 2014. [Google Scholar]
- Takata, H.; Hanafusa, T.; Mori, T.; Shimura, M.; Iida, Y.; Ishikawa, K.; Yoshikawa, K.; Yoshikawa, Y.; Maeshima, K. Chromatin compaction protects genomic DNA from radiation damage. PLoS ONE 2013, 8, e75622. [Google Scholar] [CrossRef] [PubMed]
- Cogoli, A.; Iversen, T.H.; Johnsson, D.; Mesland, D.; Oser, H. Cell biology. In Life Sciences Research in Space (ESTEC, ESA SP-1105); Oser, H., Battrick, B., Eds.; ESA Publication Division: Noordwijk, The Netherlands, 1989; pp. 49–64. [Google Scholar]
- Moore, D.; Cogoli, A. Gravitational and space biology at the cellular level. In Biological and Medical Research in Space; Moore, D., Bie, P., Oser, H., Eds.; Springer: Berlin/Heidelberg, Germany, 1996; pp. 1–106. [Google Scholar]
- Forgacs, G.; Yook, S.H.; Janmey, P.A.; Jeong, H.; Christopher, G.; Burd, C.G. Role of the cytoskeleton in signaling networks. J. Cell Sci. 2004, 117, 2769–2775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohashi, K.; Fujiwara, S.; Mizuno, K. Roles of the cytoskeleton, cell adhesion and rho signalling in mechanosensing and mechanotransduction. J. Biochem. 2017, 161, 245–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamazaki, Y.; Tsuruga, M.; Zhou, D.; Fujita, Y.; Shang, X.; Dang, Y.; Kawasaki, K.; Oka, S. Cytoskeletal disruption accelerates Caspase-3 activation and alters the intracellular membrane reorganization in DNA damage-induced apoptosis. Exp. Cell Res. 2000, 259, 64–78. [Google Scholar] [CrossRef] [PubMed]
- Yaffe, M.; Hwang, M.-K.; Gordonov, S.; Ivaska, J.; Lauffenburger, D.; Gertler, F. Phosphoproteomic analysis of DNA damage checkpoint kinase signaling reveals unexpected links to actin cytoskeletal remodeling, cell migration, and chemoresistance. FASAB J. 2017, 31, 114. [Google Scholar]
- Herraiz, C.; Calvo, F.; Pandya, P.; Cantelli, G.; Rodriguez-Hernandez, I.; Orgaz, J.L.; Kang, N.; Chu, T.; Sahai, E.; Sanz-Moreno, V. Reactivation of p53 by a cytoskeletal sensor to control the balance between DNA damage and tumor dissemination. J. Nat. Cancer Inst. 2016, 108, djv289. [Google Scholar] [CrossRef]
- Hada, M.; Huff, J.L.; Patel, Z.S.; Kawata, T.; Pluth, J.M.; George, K.A.; Cucinotta, F.A. AT cells are not radiosensitive for chromosomal exchanges at low dose. Mutat. Res. 2011, 716, 76–83. [Google Scholar] [CrossRef]
- Kumari, R.; Singh, K.P.; Dumond, J.W., Jr. Simulated microgravity decreases DNA repair capacity and induces DNA damage in human lymphocytes. J. Cell Biochem. 2009, 107, 723–731. [Google Scholar] [CrossRef]
- Mognato, M.; Girardi, C.; Fabris, S.; Celotti, L. DNA repair in modeled microgravity: Double strand break rejoining activity in human lymphocytes irradiated with gamma-rays. Mutat. Res. 2009, 663, 32–39. [Google Scholar] [CrossRef]
- Butts, J.J.; Katz, R. Theory of RBE for heavy ion bombardment. Radiat. Res. 1967, 30, 855–871. [Google Scholar] [CrossRef] [PubMed]
- Plante, I.; Cucinotta, F.A. Monte-Carlo simulation of ionizing radiation tracks. In Application of Monte Carlo Methods in Biology, Medicine and Other Fields of Science; Mode, C.J., Ed.; InTech: Rijeka, Croatia, 2011. [Google Scholar]
- Ohno, T.; Kanai, T.; Yamada, S.; Yusa, K.; Tashiro, M.; Shimada, H.; Torikai, K.; Yoshida, Y.; Kitada, Y.; Katoh, H.; et al. Carbon ion radio-therapy at the Gunma University Heavy Ion Medical Center: New facility set-up. Cancers 2011, 3, 4046–4060. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, A.; Ikeda, H.; Souda, H.; Puspitasari, A.; Held, K.D.; Hidema, J.; Nikawa, T.; Yoshida, Y.; Kanai, T. Performance evaluation of new devices to synchronize heavy-ion or X-ray irradiation and simulated microgravity for space radiation research—Improvement of a control stage. In Proceedings of the 2017 International Symposium on Space Technology and Science, Matsuyama, Japan, 3–9 June 2017. [Google Scholar]
- George, K.A.; Hada, M.; Elliott, T.; Kawata, T.; Pluth, J.M.; Cucinotta, F.A. Dose response of γ-rays and iron nuclei induction of chromosomal aberrations in normal and repair deficient cell lines. Radiat. Res. 2009, 171, 752–763. [Google Scholar] [CrossRef]
- Hada, M.; Chappell, L.J.; Wang, M.; George, K.A.; Cucinotta, F.A. Induction of chromosomal aberrations at fluences of less than one HZE particle per cell nucleus. Radiat. Res. 2014, 182, 368–379. [Google Scholar] [CrossRef]
- Hada, M.; Meador, J.A.; Cucinotta, F.A.; Gonda, S.R.; Wu, H. Chromosome aberrations induced by dual exposure of protons and iron ions. Radiat. Environ. Biophys. 2007, 46, 125–129. [Google Scholar] [CrossRef] [PubMed]
- Luomahaara, S.; Lindholm, C.; Mustonen, R.; Salomaa, S. Distribution of radiation-induced exchange aberrations in human chromosomes 1, 2 and 4. Int. J. Radiat. Biol. 1999, 75, 1551–1556. [Google Scholar] [CrossRef] [PubMed]
- Lucas, J.N.; Poggensee, M.; Straume, T. The persistence of chromosome translocations in a radiation worker accidentally exposed to tritium. Cytogenet. Cell Genet. 1992, 60, 255–256. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Radiation | Static or μG | Total Spreads Scored | No. of Aberrant Spreads | Simple Exchanges | Complex Exchanges | Total Exchanges |
|---|---|---|---|---|---|---|
| Control (0 Gy) | Static | 2025 | 13 | 0.73 ± 0.30 | 0 | 0.73 ± 0.30 |
| μG | 1994 | 19 | 1.24 ± 0.39 | 0.12 ± 0.12 | 1.37 ± 0.41 | |
| X-ray (0.5 Gy) | Static | 1024 | 23 | 3.15 ± 0.87 | 0.24 ± 0.24 | 3.39 ± 0.91 |
| μG | 1376 | 12 | 4.86 ± 0.94 | 0.72 ± 0.36 | 5.58 ± 1.00 | |
| X-ray (1.5 Gy) | Static | 1025 | 29 | 4.58 ± 1.05 | 0.48 ± 0.34 | 5.06 ± 1.10 |
| μG | 1038 | 38 | 7.16 ± 1.31 | 1.43 ± 0.58 | 8.59 ± 1.43 | |
| C-ions (0.5 Gy) | Static | 643 | 50 | 15.03 ± 2.41 | 0.39 ± 0.39 | 15.42 ± 2.44 |
| μG | 566 | 48 | 17.07 ± 2.73 | 1.75 ± 0.88 | 18.83 ± 2.87 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hada, M.; Ikeda, H.; Rhone, J.R.; Beitman, A.J.; Plante, I.; Souda, H.; Yoshida, Y.; Held, K.D.; Fujiwara, K.; Saganti, P.B.; et al. Increased Chromosome Aberrations in Cells Exposed Simultaneously to Simulated Microgravity and Radiation. Int. J. Mol. Sci. 2019, 20, 43. https://doi.org/10.3390/ijms20010043
Hada M, Ikeda H, Rhone JR, Beitman AJ, Plante I, Souda H, Yoshida Y, Held KD, Fujiwara K, Saganti PB, et al. Increased Chromosome Aberrations in Cells Exposed Simultaneously to Simulated Microgravity and Radiation. International Journal of Molecular Sciences. 2019; 20(1):43. https://doi.org/10.3390/ijms20010043
Chicago/Turabian StyleHada, Megumi, Hiroko Ikeda, Jordan R. Rhone, Andrew J. Beitman, Ianik Plante, Hikaru Souda, Yukari Yoshida, Kathryn D. Held, Keigi Fujiwara, Premkumar B. Saganti, and et al. 2019. "Increased Chromosome Aberrations in Cells Exposed Simultaneously to Simulated Microgravity and Radiation" International Journal of Molecular Sciences 20, no. 1: 43. https://doi.org/10.3390/ijms20010043
APA StyleHada, M., Ikeda, H., Rhone, J. R., Beitman, A. J., Plante, I., Souda, H., Yoshida, Y., Held, K. D., Fujiwara, K., Saganti, P. B., & Takahashi, A. (2019). Increased Chromosome Aberrations in Cells Exposed Simultaneously to Simulated Microgravity and Radiation. International Journal of Molecular Sciences, 20(1), 43. https://doi.org/10.3390/ijms20010043