Fine Mapping of the Wheat Leaf Rust Resistance Gene Lr42

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Genetic Analysis of Lr42 in Ae. tauschii
2.2. Phenotypic Evaluation of KS93U50/Morocco RIL Population
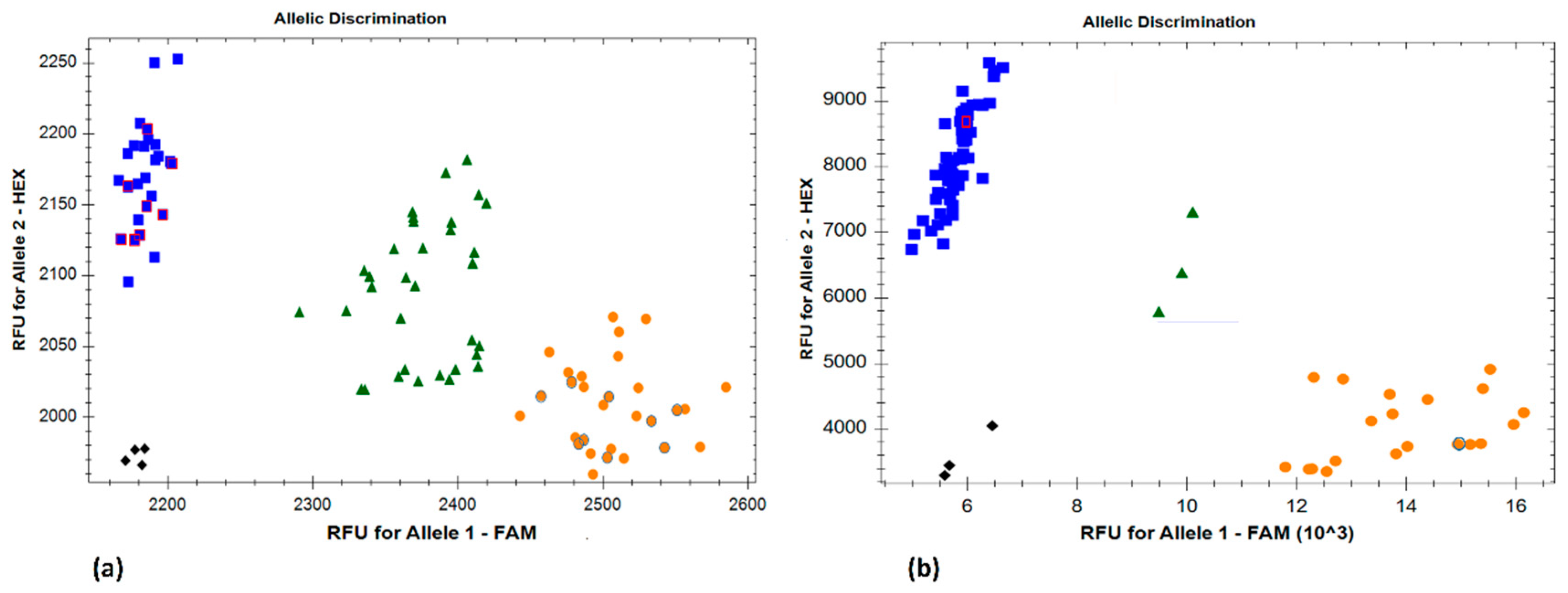
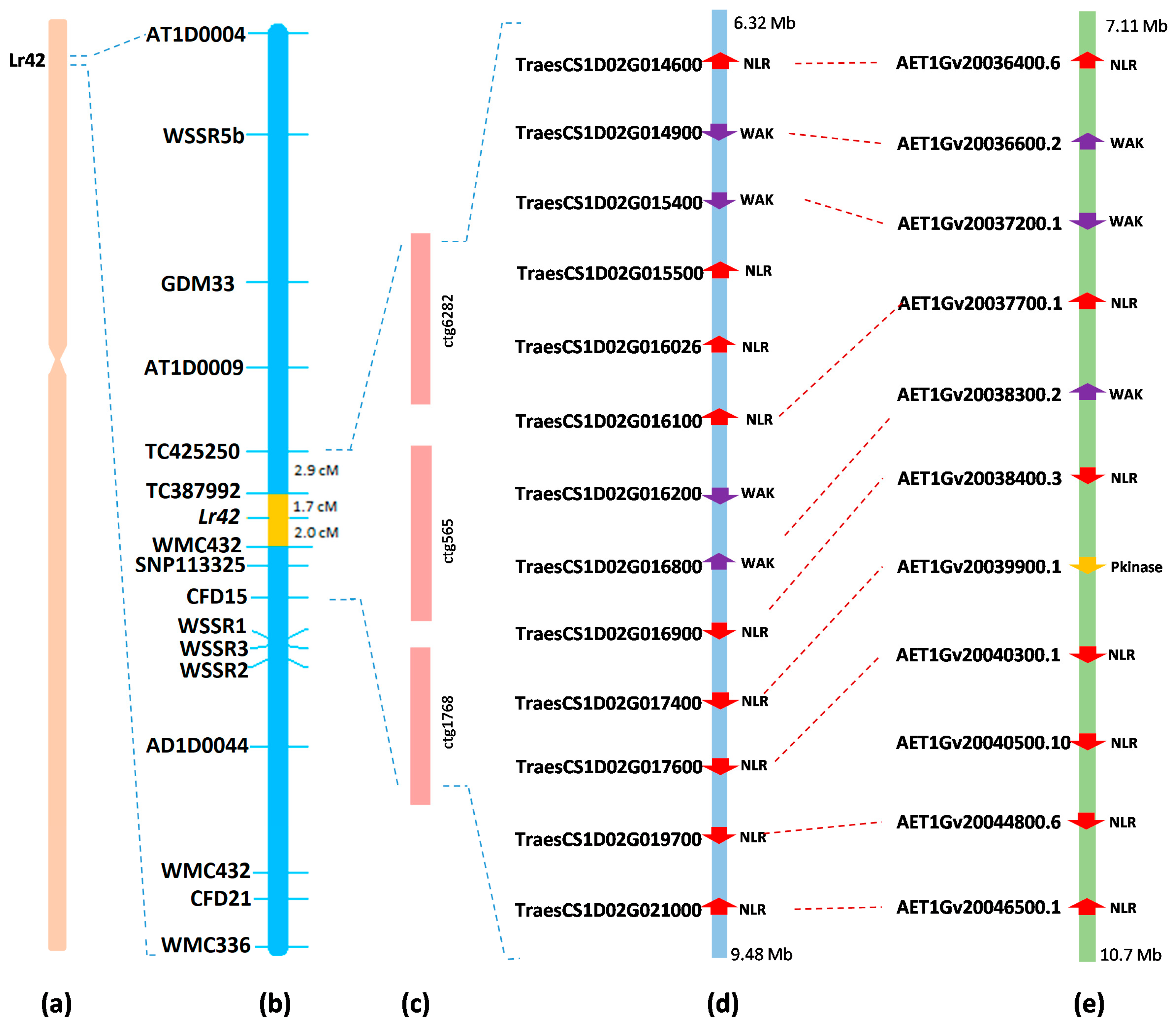
2.3. Marker Discovery and Molecular Mapping
2.4. Candidate Genes in the Lr42 Region in Wheat and Ae. tauschii
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Rust Resistance Evaluation
4.2. Marker Discovery and Saturation in the Candidate Region
4.3. DNA Extraction and Genotyping of the RIL Population
4.4. Statistical Analysis and Genetic Mapping
4.5. Physical Mapping of the Lr42 Region on Chromosome 1DS
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| MAS | Marker-assisted selection |
| KASP | Kompetitive allele-specific PCR |
| BAC | Bacterial artificial chromosome |
| EST | Expressed sequence tag |
| SSR | Simple sequence repeat |
References
- Shiferaw, B.; Smale, M.; Braun, H.J.; Duveiller, E.; Reynolds, M.; Muricho, G. Crops that feed the world 10. Past successes and future challenges to the role played by wheat in global food security. Food Sec. 2013, 5, 291–317. [Google Scholar] [CrossRef] [Green Version]
- Bolton, M.D.; Kolmer, J.A.; Garvin, D.F. Wheat Leaf Rust Caused by Puccinia Triticina. Mol. Plant Pathol. 2008, 9, 563–575. [Google Scholar] [CrossRef] [PubMed]
- Appel, J.A.; Dewolf, E.; Todd, T.; Bockus, W.W. Preliminary 2015 Kansas Wheat Disease Loss Estimates; Kansas Cooperative Plant Disease Survey Report; Kansas Department of Agriculture: Manhattan, KS, USA, 2015. Available online: http://agriculture.ks.gov/docs/default-source/PP-Disease-Reports-2014/2014-ks-wheat-disease-loss-estimates.pdf (accessed on 18 March 2019).
- Marasas, C.N.; Melinda, S.; Singh, R.P. The Economic Impact in Developing Countries of Leaf Rust Resistance Breeding in CIMMYT-Related Spring Bread Wheat; CIMMYT (International Maize and Wheat Improvement Center): Texcoco, Mexico, 2004. [Google Scholar]
- Chester, K.S. The Nature and Prevention of the Cereal Rusts as Exemplified in the Leaf Rust of Wheat; Chronica Botanica Company: Walthan, MA, USA, 1946; p. 269. [Google Scholar]
- Kolmer, J.A.; Hughes, M.E. Physiologic Specialization of Puccinia Triticina on Wheat in the United States in 2012. Plant Dis. 2014, 98, 1145–1150. [Google Scholar] [CrossRef]
- Kolmer, J.A.; Chen, X.; Jin, Y. Diseases Which Challenge Global Wheat Production the Wheat Rusts. In Wheat: Science and Trade; Carver, B.F., Ed.; Wiley-Blackwell: Ames, IA, USA, 2008. [Google Scholar]
- Qureshi, N.; Bariana, H.; Kumran, V.V.; Muruga, S.; Forrest, K.L.; Hayden, M.J.; Bansal, U. A New Leaf Rust Resistance Gene Lr79 Mapped in Chromosome 3BL from the Durum Wheat Landrace Aus26582. Theor. Appl. Genet. 2018, 131, 1091–1098. [Google Scholar] [CrossRef] [PubMed]
- Gill, B.S.; Raupp, W.J.; Sharma, H.C.; Browder, L.E.; Hatchett, J.H.; Harvey, T.L.; Moseman, J.G.; Waines, J.G. Resistance in Aegilops Squarrosa to Wheat Leaf Rust, Wheat Powdery Mildew, Greenbug, and Hessian Fly. Plant Dis. 1986, 70, 553–556. [Google Scholar] [CrossRef]
- Dhaliwal, H.S.; Singh, H.; Gupta, S.; Bagga, P.S.; Gill, K.S. Evaluation of Aegilops and wild Triticum species for resistance to leaf rust (Puccinia recondita f. Sp. Tritici) of Wheat. Int. J. Trop. Agric. 1991, 9, 118–122. [Google Scholar]
- Okamoto, Y.; Nguyen, A.T.; Yoshioka, M.; Iehisa, J.C.M.; Takumi, S. Identification of Quantitative Trait Loci Controlling Grain Size and Shape in the D Genome of Synthetic Hexaploid Wheat Lines. Breed. Sci. 2013, 63, 423–429. [Google Scholar] [CrossRef]
- Gill, B.S.; Friebe, B.; Raupp, W.J.; Wilson, D.L.; Cox, T.S.; Sears, R.G.; Brown-Guedira, G.L.; Fritz, A.K. Wheat genetics resource center: The first 25 years. Adv. Agron. 2006, 1, 73–136. [Google Scholar] [CrossRef]
- Bacon, R.K.; Kelly, J.T.; Milus, E.A.; Parsons, C.E. Registration of Soft Wheat Germplasm AR93005 Resistant to Leaf Rust. Crop Sci. 2006, 46, 1398. [Google Scholar] [CrossRef]
- Singh, R.P.; Huerta-Espino, J.; Sharma, R.; Joshi, A.K.; Trethowan, R. High Yielding Spring Bread Wheat Germplasm for Global Irrigated and Rainfed Production Systems. Euphytica 2007, 157, 351–363. [Google Scholar] [CrossRef]
- Martin, J.N.; Carver, B.F.; Hunger, R.M.; Cox, T.S. Contributions of Leaf Rust Resistance and Awns to Agronomic and Grain Quality Performance in Winter Wheat. Crop Sci. 2003, 43, 1712. [Google Scholar] [CrossRef] [Green Version]
- Cox, T.S.; Raupp, W.J.; Gill, B.S. Leaf Rust-Resistance Genes Lr41, Lr42, and Lr43 Transferred from Triticum Tauschii to Common Wheat. Crop Sci. 1994, 34, 339. [Google Scholar] [CrossRef]
- Czembor, P.C.; Radecka-Janusik, M.; Pietrusińska, A.; Czembor, H.J. Proceedings of the 11th International Wheat Genetics Symposium, Brisbane, QLD, Australia, 24–29 August 2008; Appel, R., Eastwood, R., Lagudah, E., Langridge, P., Mackay, M., McIntyre, L., Sharp, P., Eds.; Sydney University Press: Sidney, Australia, 2008; pp. 739–740. [Google Scholar]
- Sun, X.; Bai, G.; Carver, B.F.; Bowden, R. Molecular mapping of wheat leaf rust resistance gene Lr42. Crop Sci. 2010, 50, 59–66. [Google Scholar] [CrossRef]
- Liu, Z.; Bowden, R.L.; Bai, G. Molecular markers for leaf rust resistance gene Lr42 in Wheat. Crop Sci. 2013, 53, 1566–1570. [Google Scholar] [CrossRef]
- International Barley Genome Sequencing Consortium; Mayer, K.F.; Waugh, R.; Brown, J.W.; Schulman, A.; Langridge, P.; Platzer, M.; Fincher, G.B.; Muehlbauer, G.J.; Sato, K.; et al. A physical, genetic and functional sequence assembly of the barley genome. Nature 2012, 491, 711–716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, M.-C.; Gu, Y.Q.; You, F.M.; Deal, K.R.; Ma, Y.; Hu, Y.; Huo, N.; Wang, Y.; Wang, J.; Chen, S.; et al. A 4-gigabase physical map unlocks the structure and evolution of the complex genome of Aegilops tauschii, the wheat D-genome progenitor. Proc. Natl. Acad. Sci. USA 2013, 110, 7940–7945. [Google Scholar] [CrossRef]
- International Wheat Genome Sequencing Consortium (IWGSC); IWGSC RefSeq principal investigators; Appels, R.; Eversole, K.; Feuillet, C.; Keller, B.; Rogers, J.; Stein, N.; IWGSC whole-genome assembly principal investigators; Pozniak, C.J.; et al. Shifting the Limits in Wheat Research and Breeding Using a Fully Annotated Reference Genome. Science 2018, 361, eaar7191. [Google Scholar] [CrossRef]
- El-Gebali, S.; Mistry, J.; Bateman, A.; Eddy, S.R.; Luciani, A.; Potter, S.C.; Qureshi, M.; Richardson, L.J.; Salazar, G.A.; Smart, A.; et al. The Pfam Protein Families Database in 2019. Nucleic Acids Res. 2019, 47, D427–D432. [Google Scholar] [CrossRef] [PubMed]
- Kolmer, J.; Phytopathology, P.D. Gene Expression in the Triticum Aestivum-Puccinia Recondita f. Sp. tritici gene-for-gene system. Phytopathology 1994, 84, 437–440. [Google Scholar] [CrossRef]
- Feuillet, C.; Travella, S.; Stein, N.; Albar, L.; Nublat, A.; Keller, B. Map-Based Isolation of the Leaf Rust Disease Resistance Gene Lr10 from the Hexaploid Wheat (Triticum aestivum, L.) Genome. Proc. Natl. Acad. Sci. USA 2003, 100, 15253–15258. [Google Scholar] [CrossRef]
- Huang, L.; Brooks, S.A.; Li, W.; Fellers, J.P.; Trick, H.N.; Gill, B.S. Map-Based cloning of leaf rust resistance gene lr21 from the large and polyploid genome of bread wheat. Genetics 2003, 164, 655–664. [Google Scholar] [PubMed]
- Cloutier, S.; McCallum, B.D.; Loutre, C.; Banks, T.W.; Wicker, T.; Feuillet, C.; Keller, B.; Jordan, M.C. Leaf rust resistance gene Lr1, isolated from bread wheat (Triticum aestivum, L.) is a member of the large psr567 gene family. Plant Mol. Biol. 2007, 65, 93–106. [Google Scholar] [CrossRef] [PubMed]
- Thind, A.K.; Wicker, T.; Šimková, H.; Fossati, D.; Moullet, O.; Brabant, C.; Vrána, J.; Doležel, J.; Krattinger, S.G. Rapid Cloning of Genes in Hexaploid Wheat Using Cultivar-Specific Long-Range Chromosome Assembly. Nat. Biotechnol. 2017, 35, 793–796. [Google Scholar] [CrossRef] [PubMed]
- Periyannan, S.; Moore, J.; Ayliffe, M.; Bansal, U.; Wang, X.; Huang, L.; Deal, K.; Luo, M.; Kong, X.; Bariana, H.; et al. the gene Sr33, an ortholog of barley mla genes, encodes resistance to wheat stem rust race Ug99. Science 2013, 341, 786–788. [Google Scholar] [CrossRef]
- Saintenac, C.; Zhang, W.; Salcedo, A.; Rouse, M.N.; Trick, H.N.; Akhunov, E.; Dubcovsky, J. Identification of wheat gene Sr35 that confers resistance to Ug99 stem rust race group. Science 2013, 341, 783–786. [Google Scholar] [CrossRef]
- Steuernagel, B.; Periyannan, S.K.; Hernández-Pinzón, I.; Witek, K.; Rouse, M.N.; Yu, G.; Hatta, A.; Ayliffe, M.; Bariana, H.; Jones, J.D.G.; et al. Rapid cloning of disease-resistance genes in plants using mutagenesis and sequence capture. Nat. Biotechnol. 2016, 34, 652–655. [Google Scholar] [CrossRef]
- Liu, W.; Frick, M.; Huel, R.; Nykiforuk, C.L.; Wang, X.; Gaudet, D.A.; Eudes, F.; Conner, R.L.; Kuzyk, A.; Chen, Q.; et al. The stripe rust resistance gene Yr10 encodes an evolutionary-conserved and unique CC–NBS–LRR sequence in wheat. Mol. Plant 2014, 7, 1740–1755. [Google Scholar] [CrossRef]
- Shi, G.; Zhang, Z.; Friesen, T.L.; Raats, D.; Fahima, T.; Brueggeman, R.S.; Lu, S.; Trick, H.N.; Liu, Z.; Chao, W.; et al. The hijacking of a receptor kinase–driven pathway by a wheat fungal pathogen leads to disease. Sci. Adv. 2016, 2, e1600822. [Google Scholar] [CrossRef] [PubMed]
- Saintenac, C.; Lee, W.-S.; Cambon, F.; Rudd, J.J.; King, R.C.; Marande, W.; Powers, S.J.; Bergès, H.; Phillips, A.L.; Uauy, C.; et al. Wheat receptor-kinase-like protein stb6 controls gene-for-gene resistance to fungal pathogen zymoseptoria tritici. Nat. Genet. 2018, 50, 368–374. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Martín, J.; Steuernagel, B.; Ghosh, S.; Herren, G.; Hurni, S.; Adamski, N.; Vrána, J.; Kubaláková, M.; Krattinger, S.G.; Wicker, T.; et al. Rapid gene isolation in barley and wheat by mutant chromosome sequencing. Genome Biol. 2016, 17, 221. [Google Scholar] [CrossRef]
- Huang, X.-Q.; Röder, M.S. High-density genetic and physical bin mapping of wheat chromosome 1D reveals that the powdery mildew resistance gene Pm24 is located in a highly recombinogenic region. Genetica 2011, 139, 1179–1187. [Google Scholar] [CrossRef]
- Gill, K.S.; Gill, B.S.; Endo, T.R.; Taylor, T. Identification and high-density mapping of gene-rich regions in chromosome group 1 of wheat. Genetics 1996, 144, 1001–1012. [Google Scholar]
- Stakman, E.C.; Stewart, D.M.; Loegering, W.Q. Identification of physiologic races of Puccinia graminis var. tritici. Can. J. Plant Pathol. 1981, 3, 33–39. [Google Scholar]
- Roelfs, A.P.; Bushnell, W.R.; San, O.; New, D.; London, Y.; Montreal, T.; Tokyo, S. The Cereal Rusts: Diseases, Distribution, Epidemiology, and Control; Elsevier: Amsterdam, The Netherlands, 1985. [Google Scholar]
- Saghai-Maroof, M.A.; Soliman, K.M.; Jorgensen, R.A.; Allard, R.W. Ribosomal DNA Spacer-Length Polymorphisms in Barley: Mendelian Inheritance, Chromosomal Location, and Population Dynamics. Proc. Natl. Acad. Sci. USA 1984, 81, 8014–8018. [Google Scholar] [CrossRef]
- The R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017; Available online: https://www.R-project.org (accessed on 18 March 2019).
- de Givry, S.; Bouchez, M.; Chabrier, P.; Milan, D.; Schiex, T. CARHTA GENE: Multipopulation integrated genetic and radiation hybrid mapping. Bioinformatics 2005, 21, 1703–1704. [Google Scholar] [CrossRef] [PubMed]
- Kosambi, D.D. The estimation of map distances from recombination values. Ann. Eugen. 1943, 12, 172–175. [Google Scholar] [CrossRef]
- Voorrips, R.E. MapChart: Software for the graphical presentation of linkage maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef]
- Luo, M.-C.; Gu, Y.Q.; Puiu, D.; Wang, H.; Twardziok, S.O.; Deal, K.R.; Huo, N.; Zhu, T.; Wang, L.; Wang, Y.; et al. Genome sequence of the progenitor of the wheat d genome aegilops tauschii. Nature 2017, 551, 498. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Sr No. | Species | Population | Generation | Lines Evaluated | Observed Ratio | Expected Ratio | χ2 | p-Value* |
|---|---|---|---|---|---|---|---|---|
| 1 | Ae. tauschii | TA2450/TA2433 | F2 | 66 | 50:16 | 3:1 | 0.20 | 0.89 |
| F2:3 | 100 | 27:54:19 | 1:2:1 | 1.92 | 0.38 | |||
| 2 | Ae. tauschii | TA2450/TA10132 (AL8/78) | F2 | 67 | 53:14 | 3:1 | 0.60 | 0.44 |
| F2:3 | 100 | 33:53:14 | 1:2:1 | 7.58 | 0.02 | |||
| 3 | T. aestivum | KS93U50/Morocco | F5 RIL | 234 | 99:135 | 1:1 | 5.54 | 0.02 |
| Sr. No. | Primer Name | Assay Type | Sequence(s) |
|---|---|---|---|
| 1 | WSSR1 | SSR | ACGACGTTGTAAAACGACTGGAGACAGACGAACGCATA |
| TGCATGCATACACACACCAG | |||
| 2 | WSSR2 | SSR | ACGACGTTGTAAAACGACAGCAATGCAGTTGCAAAGAG |
| GCAAAGATGGACAGATGGCT | |||
| 3 | WSSR3 | SSR | ACGACGTTGTAAAACGACAAGATCAGCTCCGACAGCTC |
| CGAAGTCAGCACAAACCAAA | |||
| 4 | WSSR5 | SSR | ACGACGTTGTAAAACGACTGGTGAATCTTGCACCACAT |
| CTGGACACCGTTCGTTAGGT | |||
| 5 | AT1D004 | KASP | GGTACCATGTTGTTTCGCATGTCTAT |
| GTACCATGTTGTTTCGCATGTCTAC | |||
| GGAGGCAGAGACAATAAGTTTATGTTACAA | |||
| 6 | AT1D0009 | KASP | GGAGATCTTTATATTTGTGGTTTGCCA |
| GAGATCTTTATATTTGTGGTTTGCCG | |||
| CCAGGTCACAGGCTGTGATGTTTAA | |||
| 7 | TC425250 | KASP | GCACTACTTTTATTGATGTTGTGTAACC |
| AAGCACTACTTTTATTGATGTTGTGTAACT | |||
| CAGAGGGAAGAAAACAACACTGAACAAAA | |||
| 8 | TC387992 | KASP | TTGGATCTGCATTCCTTCTCCCA |
| GGATCTGCATTCCTTCTCCCG | |||
| CTTTGGGATGTTGCTGCTGGAGAT | |||
| 9 | SNP113325 | KASP | GGTGTTTGGCAGCATCATCACG |
| GGTGTTTGGCAGCATCATCACC | |||
| GACAACTTGAGACACTAGATATCAGAGAT |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gill, H.S.; Li, C.; Sidhu, J.S.; Liu, W.; Wilson, D.; Bai, G.; Gill, B.S.; Sehgal, S.K. Fine Mapping of the Wheat Leaf Rust Resistance Gene Lr42. Int. J. Mol. Sci. 2019, 20, 2445. https://doi.org/10.3390/ijms20102445
Gill HS, Li C, Sidhu JS, Liu W, Wilson D, Bai G, Gill BS, Sehgal SK. Fine Mapping of the Wheat Leaf Rust Resistance Gene Lr42. International Journal of Molecular Sciences. 2019; 20(10):2445. https://doi.org/10.3390/ijms20102445
Chicago/Turabian StyleGill, Harsimardeep S., Chunxin Li, Jagdeep S. Sidhu, Wenxuan Liu, Duane Wilson, Guihua Bai, Bikram S. Gill, and Sunish K. Sehgal. 2019. "Fine Mapping of the Wheat Leaf Rust Resistance Gene Lr42" International Journal of Molecular Sciences 20, no. 10: 2445. https://doi.org/10.3390/ijms20102445
APA StyleGill, H. S., Li, C., Sidhu, J. S., Liu, W., Wilson, D., Bai, G., Gill, B. S., & Sehgal, S. K. (2019). Fine Mapping of the Wheat Leaf Rust Resistance Gene Lr42. International Journal of Molecular Sciences, 20(10), 2445. https://doi.org/10.3390/ijms20102445