1. Introduction
The fur color of mammals mainly depends on melanin deposition, and melanogenesis is mainly regulated by melanocytes. The production of different types of melanin by melanocytes, together with different distribution of these pigments, result in a variety of hair colors in mammals [
1]. Related genes, such as
TYR,
TYRP1,
ASIP,
MITF, and
CREB1, have been found to regulate melanin deposition [
2,
3]. Previously, by using the transcriptome sequencing (RNA-Seq), a significant difference was found in the expression of the
Slc7a11 gene in the skin of rabbits with different fur colors. It was speculated to be involved in fur pigmentation [
4].
In the melanogenesis pathway, both eumelanin and pheomelanin are derived from a common precursor named dopaquinone [
5]. Cystine or glutathione is required for the production of pheomelanin and it is xCT, the protein encoded by the
Slc7a11 gene, that acts as a vector to transport extracellular cystine into the cell and maintain normal intracellular glutathione levels. Pheomelanin and eumelanin together form a mixed pigment that determines the skin and fur color of animals [
6,
7,
8]. In the hair of
Slc7a11 gene-mutated mice (sut), the level of pheomelanin was significantly decreased, while the eumelanin level was substantially unchanged, so that the wild-type mice with yellow background appeared gray [
9]. The sut mutation results in a huge deletion in the
Slc7a11 gene, but similar deletions could not be found in this region of Rex Rabbits with six different fur colors, including black (BL), chinchilla (CH), white (WH), brown (BR), protein yellow (PY), and protein chinchilla (PC). SNPs in the exon region of
Slc7a11 were also scanned, but no mutation site was found. This indicates that
Slc7a11 is highly conserved in the population (data not shown). Currently, studies on the functions of
Slc7a11 mainly focus on its important roles in cell proliferation [
10], oxidative stress response [
11], and Alzheimer’s disease target treatment [
12]. Research studying its regulatory mechanisms is focused on microRNAs affecting cancer development and apoptosis by targeting and regulating
Slc7a11 [
10,
13]. Few studies regarding melanin deposition have been reported.
To explore the molecular regulation mechanism of Slc7a11 in the melanin deposition of Rex rabbit fur, rabbit melanocytes were isolated and identified. The expression pattern of Slc7a11 in Rex rabbits with different fur colors was analyzed. Furthermore, we verified that POU2F1 has an important regulatory role in the transcriptional activation of the Slc7a11 gene promoter. This result provides a theoretical basis for further analysis of the deposition mechanism of the fur pigmentation as well as for the transformation of fur color in animals.
3. Discussion
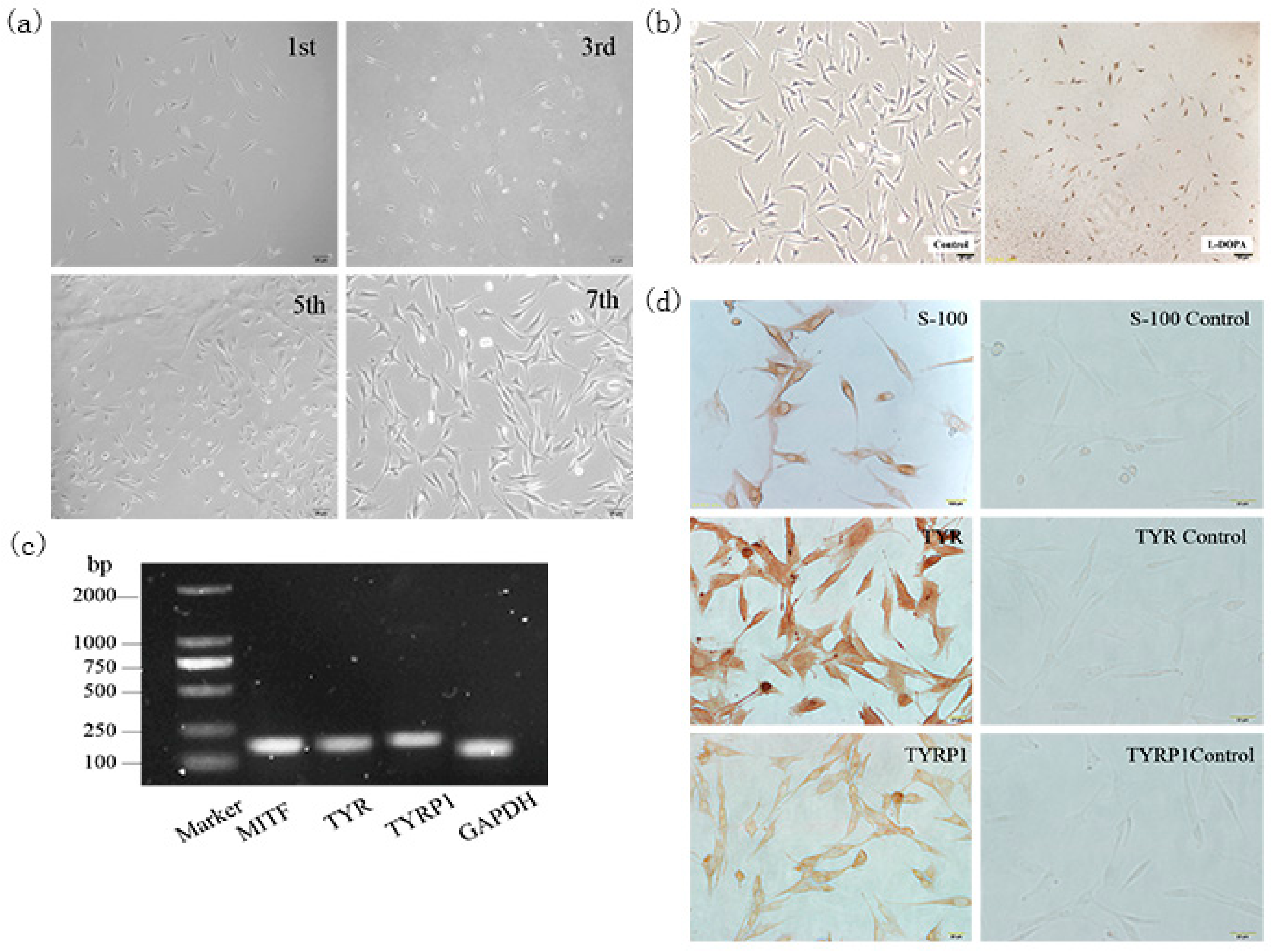
Skin melanocytes are located on the basement membrane between the epidermis and the dermis. Skin melanocytes have been isolated from humans, pigs, and other animals using 0.25% trypsin and successfully cultured [
14,
15]. In a preliminary study, the back skin of rabbits was digested with trypsin, but the results were not satisfactory, indicating that the use of trypsin alone did not result in sufficient digestion or separation of melanocytes from the skin of hairy animals. However, the use of Dispase II enzyme digestion, allowed us to obtain pure melanocytes for the first time, which laid the foundation for our later work.
Fur color is controlled by different genes in the process of pigment biosynthesis. The differences in color are mainly due to the different ratios between pheomelanin (red and yellow) and eumelanin (black) [
16,
17,
18]. The main model currently proposed is that the ratio of eumelanin/pheomelanin in mammalian pigments is solely or indirectly regulated by the activity of tyrosinase—the rate-limiting enzyme of melanin synthesis [
19]. This model inferred that in the presence of low concentrations of tyrosinase, dopaquinone reacts with cysteine to produce cysteine dopa, thereby increasing the level of pheomelanin [
20]. However, this pattern is not yet fully understood.
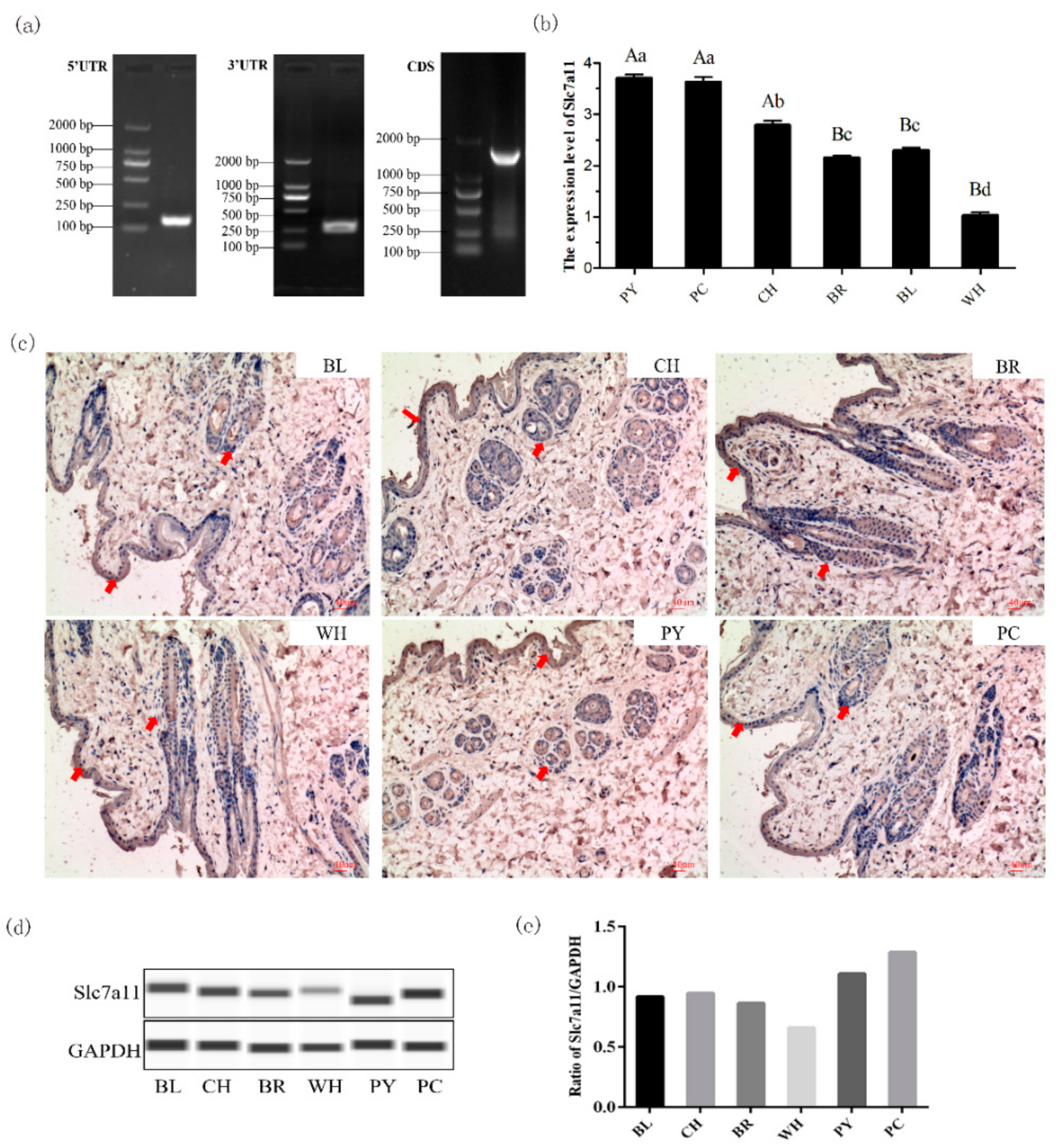
Studies have confirmed that the xCT transporter encoded by the
Slc7a11 gene is crucial for the regulation of pigments and can directly affect the increase of pheomelanin [
9]. Based on previous studies, Rex Rabbits with a variety of natural fur colors were used to explore the expression pattern of
Slc7a11 gene in dorsal skin tissues with different fur colors. xCT was expressed in the epidermis, hair bulb, and hair root-sheaths of the skin tissues examined by immunohistochemistry. It is known that melanocytes in the skin are often distributed in different regions when matured and that the only place where melanin can be supplied to the hair shaft is the hair bulb [
21,
22,
23]. In this study, it was found that the expression sites of the
Slc7a11 gene were consistent with the distribution of melanin, suggesting that the protein was related to the formation of Rex Rabbit fur color. Moreover, the
Slc7a11 gene had the highest expression level in protein yellow coat, and the lowest level in white coat, by real-time quantitative and Wes system analyses. It is speculated that
Slc7a11 affects the formation of cystine, which is reduced to cysteine, and thus alters the production of pheomelanin. This is consistent with studies on alpaca and sheep [
24,
25].
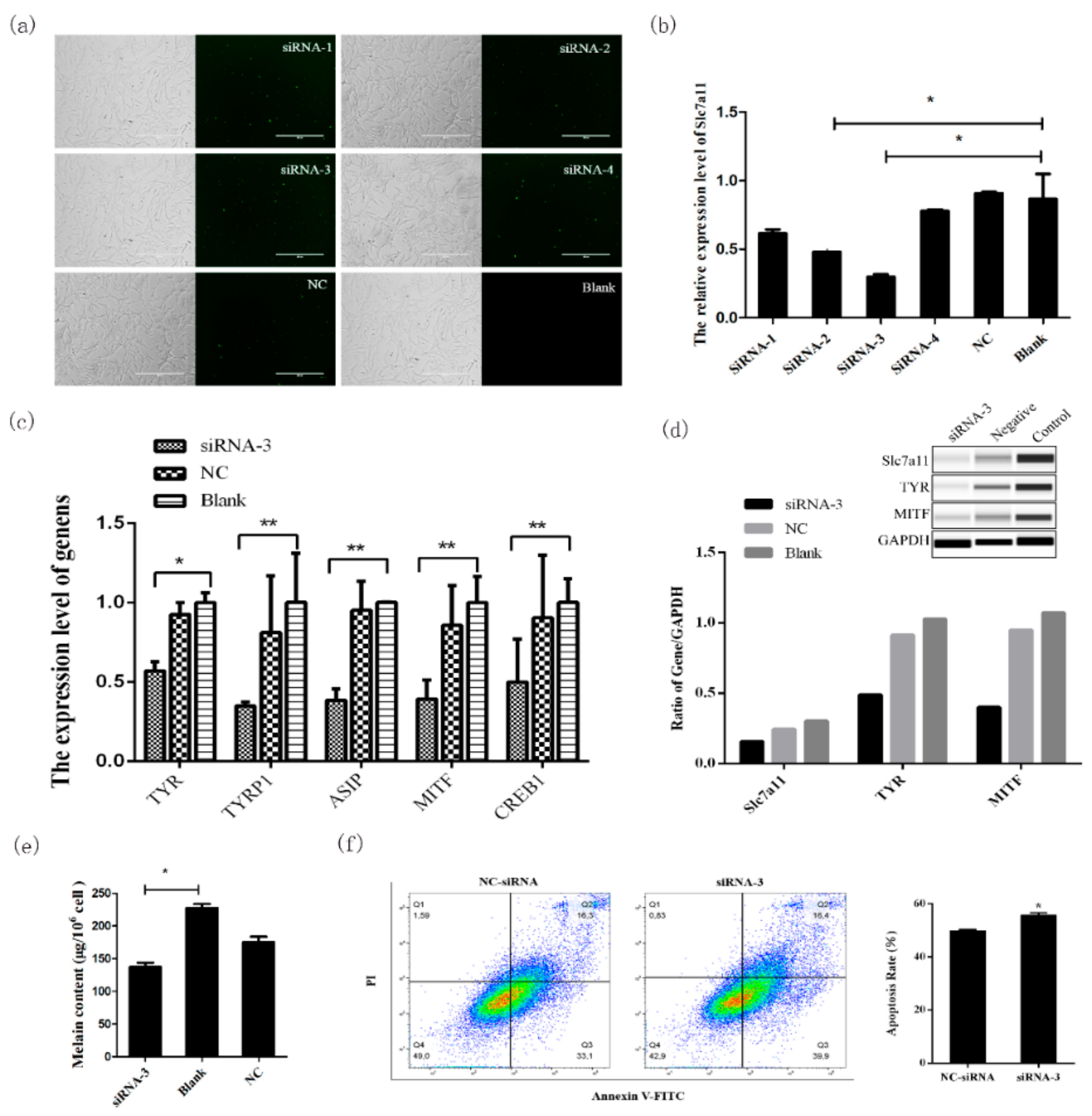
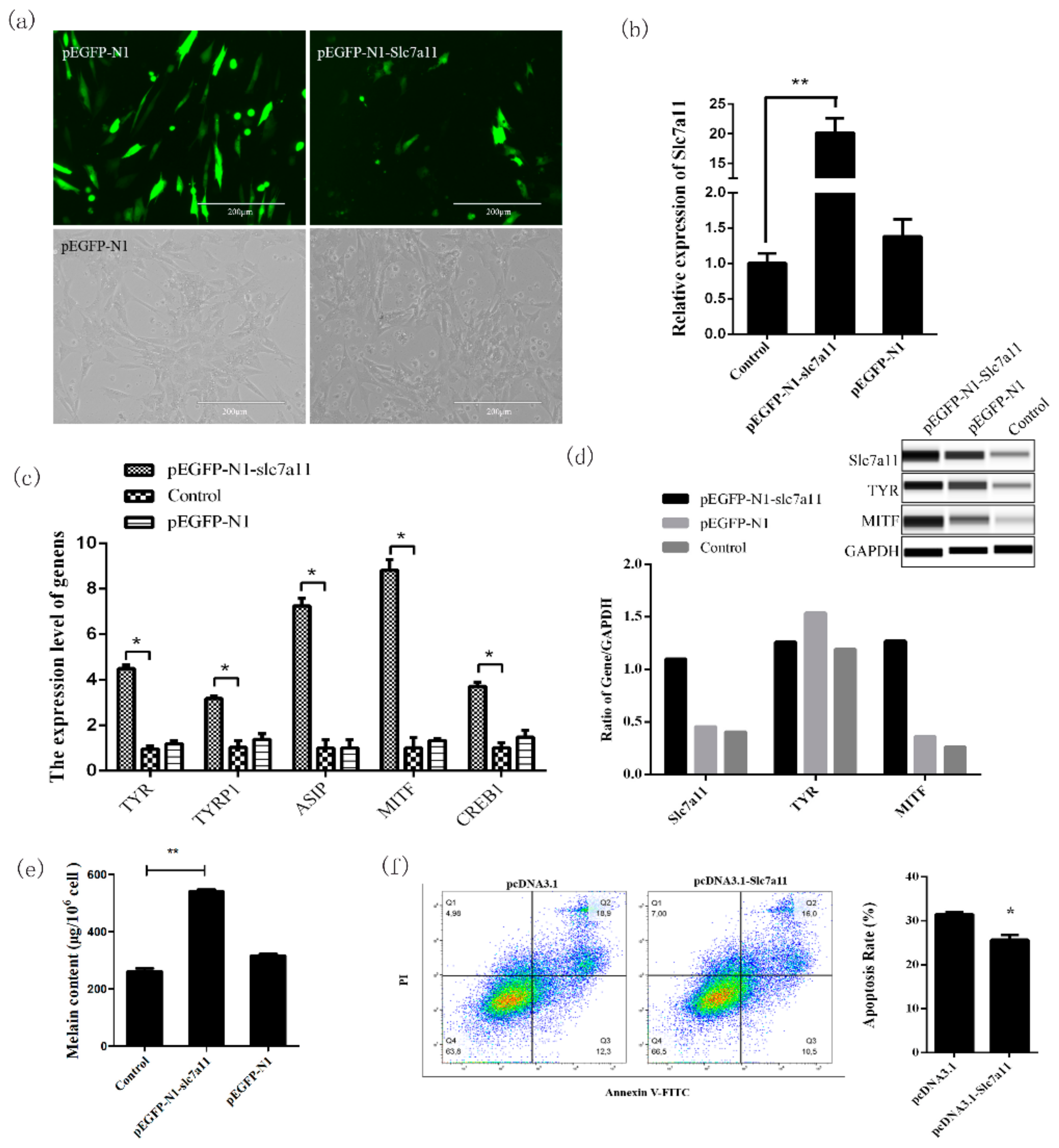
Knockdown and overexpression analyses of
Slc7a11 confirmed that this gene can affect the expression of
TYR,
MITF,
TYRP1, and
ASIP in the melanogenesis pathway [
3,
26].
TYR is a key enzyme in melanin formation and its expression level and activity determine the rate and yield of melanin production. Upon activation,
TYR catalyzes the hydroxylation of tyrosine to L-3,4-phenylalanine (DOPA), which is rapidly oxidized to form dopaquinone [
27].
MITF regulates the expression of the tyrosine gene family and participates in melanin production [
28]. Studies have shown that the expression of
TYRP1,
ASIP and
CREB1 determines skin melanin deposition [
29,
30,
31]. The
Slc7a11 gene affects the transcription of genes involved in melanogenesis, which is closely related to the formation of fur color. Moreover,
Slc7a11 decreases melanocyte apoptosis and further affects melanogenesis of melanocytes. These results confirmed that
Slc7a11 is closely related to the formation of Rex rabbit fur color. The regulatory factors required for such expression patterns would be the next research objective.
Chintala et al. found that the mouse light gray (sut) mutation was due to the inhibition of phaeomelanin production. It was caused by a large deletion of the
Slc7a11 gene starting from the 11th intron crossing the 12th exon into the region adjacent to the Pcdh18 gene. This resulted in a change in the 3′ end of
Slc7a11 transcription [
9]. Based on these results, the identification of similar deletions in the natural populations of Rex Rabbits with six fur colors was carried out. Unfortunately, no such large fragment deletions were found in similar areas. Exon scanning did not show any SNPs sites, indicating that
Slc7a11 is relatively conserved in the population. Similar results have been seen in humans and sheep [
25,
32,
33]. The regulatory mechanism of
Slc7a11 is unknown.
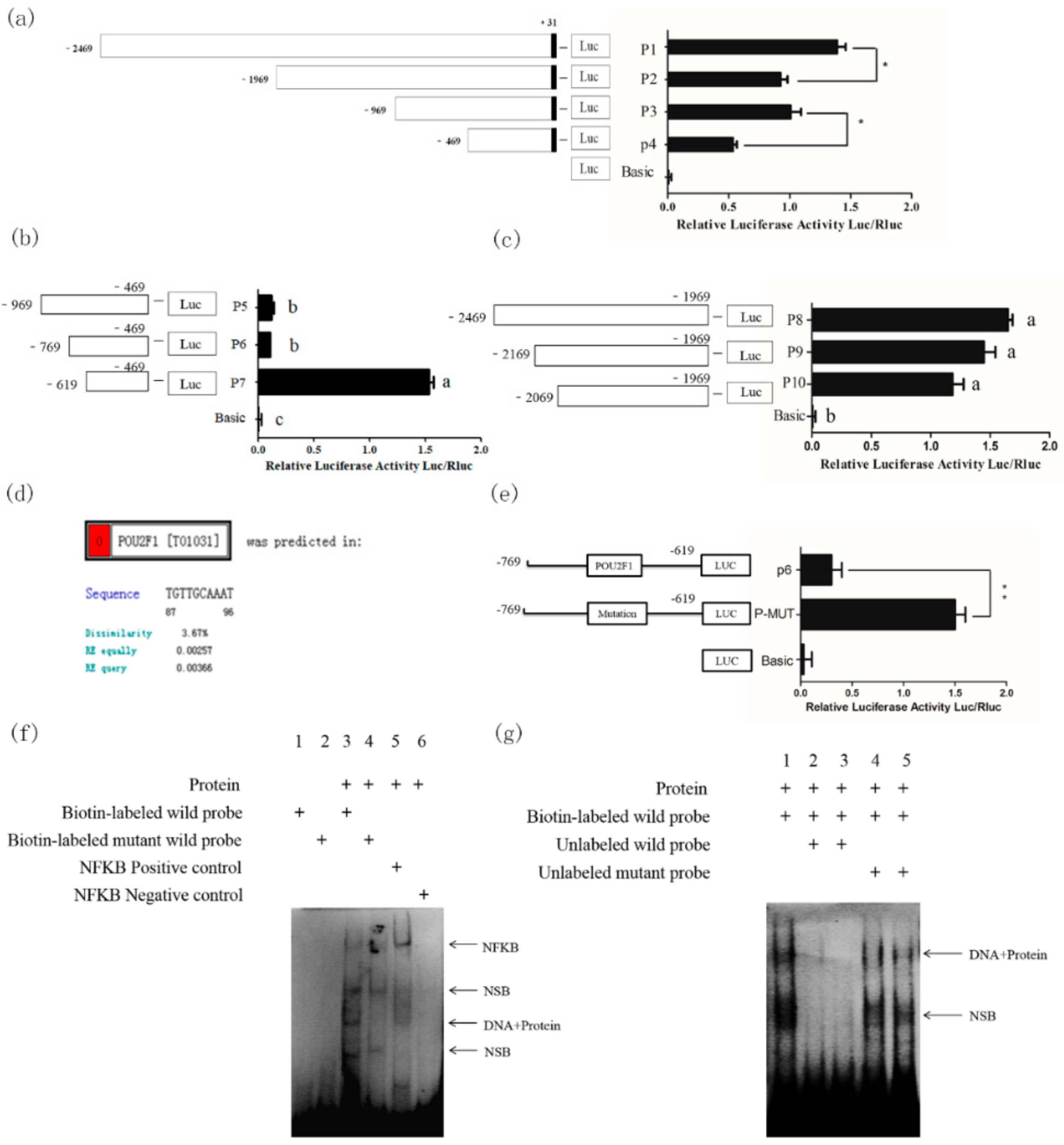
Furthermore, the promoter series deletion vector dual-luciferase was used to search for the −769 to −619 bp transcription core region of
Slc7a11. We also confirmed that POU2F1 protein binds to the −713 to −703 bp region of the
Slc7a11 promoter to inhibit its activity.
POU2F1, also known as Octamer Transcription factor-1 (Oct-1), is a widely expressed POU protein factor. Recent studies suggest that it can regulate target genes associated with processes such as oxidation, anti-cytotoxicity, stem cell function, and cancer development, etc. [
34,
35]. Ethanol has been reported to increase the expression of
Slc7a11 by reducing the binding of
POU2F1 to the
Slc7a11 gene promoter [
36]. In this study,
POU2F1 was found to specifically bind to the
Slc7a11 promoter and inhibit its transcription. Together with the previous finding that
Slc7a11 promotes melanin cytochrome deposition,
POU2F1 can be used as a target for artificial modification of animal fur colors.
4. Materials and Methods
4.1. Primary Separation and Culture of Rabbit Melanocytes
Rabbits were injected with anesthetic on the back, and a piece of the back skin (1.5 cm × 1.5 cm) was dissected. Any particles on the skin surface and subcutaneous connective tissue were removed. After tissue collection, the wounds were treated with iodine tincture. After the experiment, rabbits were anesthetized by an intraperitoneal injection of sodium pentobarbital (50 mg/kg).
The skin sample was digested with 0.25% DispaseII enzyme digestion solution (Sigma, Darmstadt, Germany) for 14–16 h at 4 °C. The epidermis was gently peeled off from the dermis, cut into small pieces, and digested with 0.25% trypsin (Gibco, Carlsbad, CA, USA) for 8 min at 37 °C. The sample was then filtered through a 200 mesh filter and the supernatant discarded. The cells were resuspended in M254 medium (Gibco, Carlsbad, CA, USA) and incubated at 37 °C in a 5% CO2 incubator. The cells were digested with 1 mL of 0.25% trypsin and subcultured.
This study was carried out in accordance with the recommendations of Animal Care and Use Committee at Yangzhou University. The protocol was approved by the Animal Care and Use Committee at Yangzhou University (Yangzhou, China, 24 October 2017, No. 201710001).
4.2. DOPA Staining
Melanocytes in the logarithmic growth phase were used to prepare sterile cell culture slides. The inoculated 24-well plates were cultured for 3 days and treated with 1 mL of 4% paraformaldehyde fixative (Solarbio, Beijing, Tongzhou, China) for 30 min at 4 °C. The plates were washed 3 times with pre-cooled PBS prior to the addition of L-DOPA (Sigma). After incubation in L-DOPA for 1 day, the incubation solution was renewed and the plates were further incubated at 37 °C for 12 h, with constant observation once every 30 min. The plates were washed with PBS once the staining was complete and observed under a microscope [
37].
4.3. Immunostaining
Melanocytes in the logarithmic growth phase or skin tissues from Rex rabbits of 6 different fur colors were used to prepare slides. The slides were incubated with primary antibodies Slc7a11 (1:500 rabbit polyclonal, Abcam, Cambridge, UK), S-100 (1:500 mouse monoclonal, Boster, Wuhan, China), TYRP1 (1:250 rabbit polyclonal, Abcam), TYR (1:1000 rabbit polyclonal, Abcam) overnight at 4 °C, with PBS as a negative control. The slides were subsequently incubated with IgG secondary antibody (1:2500 goat polyclonal, Abcam) at 37 °C for 20 min and developed for 3–5 min at room temperature in the dark with freshly prepared DAB solution (Boster). The slides were observed under a microscope.
4.4. RACE and Cloning of Slc7a11 Gene
Three specific 5′ RACE primers and two 3′ RACE primers were designed according to the Race kit instructions (Invitrogen & Clontech, Carlsbad, CA, USA) (
Table 1). The full-length cDNA sequence of the
Slc7a11 gene was assembled based on known sequences and 5′ and 3′ RACE results, and submitted to NCBI (Accession no.: KY971639.1). The
Slc7a11 cDNA was reconstructed into the pEGFP-N1 vector with restriction enzymes HindIII and SacII.
4.5. Knockdown of Slc7a11 by siRNA
Fluorescently labeled siRNAs (with 5′ FAM modification) and Negative Control siRNAs were purchased from Shanghai GenePharma Co., Ltd (
Table 2). When melanocyte confluence reached about 65%, the siRNA oligo/Lipofectamine™ 2000 (Invitrogen, Carlsbad, CA, USA) complex at a ratio of 1:2 was prepared for transfection. After 24 h, the transfection efficiency was examined by fluorescence microscopy.
4.6. Real-Time PCR
Real-time PCR was carried out using SYBR
® Premix Ex Taq™ II (TaKaRa, Dalian, China) on an Applied Bio systems 7500 Real-Time PCR System with the following program: 1 cycle at 95 °C for 30 s, followed by 40 cycles of 95 °C for 5 s, and 60 °C for 30 s. The primers used for detection of gene expression are shown in
Table 3. Each sample was measured three times, and the results were normalized to GAPDH. The relative expression of the target gene was calculated by the ΔΔCt method; namely, the fold difference between the target gene and the reference gene (experimental group)/the fold difference between the target gene and the reference gene (control group)
.
4.7. Simple Western Analysis
Pre-cooled RIPA lysis buffer (Sigma) was mixed with PMSF (with a final concentration of 1 mM) and added to the tissue or cell samples, which were centrifuged at 10,000 rpm for 5 min at 4 °C. The supernatant was discarded and the total protein obtained. Simple Western analysis was performed using the Wes Simple Western (Protein Simple) system. This assay uses an automated Western system—no gels, no transfer devices, no blots, no film and no manual analysis, resulting in a digital result. The test results were analyzed using the Compass program.
4.8. Apoptosis Assay
The apoptosis rate was measured with an Annexin V-FITC Apoptosis Detection Kit (Vazyme, Nanjing, China), according to the manufacturer’s instructions. After transfection, the cells were collected and washed twice with PBS, then centrifuged at 300 g, 4 °C for 5 min. The cells were resuspended in 100 μL 1× Binding Buffer and mixed gently followed by the addition of 5 μL Annexin V-FITC and 5 μL PI Staining Solution, which were allowed to incubate at room temperature for 10 min in the dark. Finally, cells were added to 400 μL 1×Binding Buffer and mixed gently. Cells were sorted by fluorescence-activated cell sorting using the Flow cytometer FACSAria SORP (Becton Dickinson, Franklin Lakes, NJ, USA). The apoptosis rate was calculated using the following equation: total number of cells is composed of number of cells in the right upper quadrant and number of cells in the right lower quadrant.
4.9. Determination of Melanin Level
The cells were lysed with 1 mL 0.2 mol/L NaOH. The cell lysate was collected and incubated at 37 °C for 2 h. Wavelength measurement was performed at 475 nm using a microplate reader. The standard curve was plotted using the Melanin synthetic standard (Sigma). Each group was repeated 3 times, from which the melanin level was calculated.
4.10. Luciferase Vector Construction and Reporter Assays
Promoter-specific primers were designed using O1igo7 (
Table 4), and the
Slc7a11 promoter region was analyzed using PROMO (
http://www.cbs.dtu.dk/services/Promoter/) to obtain possible transcription factor binding sites. The
Slc7a11 promoter was reconstructed into the pGL3-basic vector with restriction enzymes KpnI and BgIII. The internal reference plasmid pRL-TK and the recombinant plasmid were co-transfected into RAB-9 cells (ATCC), with the pGL3-basic plasmid and pRL-TK plasmid co-transfected cells as the negative control group and the cells with no substance transfected as the blank group. The transfected cells were collected and analyzed using the Dual-Luciferase Reporter Assay System (Promega, Madison, WI, USA).
4.11. Electrophoresis Mobility Shift Assay (EMSA)
The nuclear proteins of melanocytes were extracted and the concentrations determined. Based on the binding sequence of
POU2F1, normal and mutant probes were designed (
Table 5) and biotinylated at the 5′ end. The EMSA reaction system was formulated as shown in
Table 6. The cold-competitive EMSA reaction system is shown in
Table 7. The samples were analyzed by Native-PAGE, transferred, and UV cross-linked prior to carrying out the chemiluminescence reaction, development, and photographing.
4.12. Statistical Analysis
Each experiment was repeated at least three times. Statistical analysis Q-Q plot was used to check the normality of the data. Statistical significance between experimental and control groups was analyzed by Independent-Sample Test and one-way ANOVA. The results are presented as mean ± standard deviation (SD) at two levels of significance, p < 0.05 and p < 0.01.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}