Involvement of G-Protein-Coupled Receptor 40 in the Inhibitory Effects of Docosahexaenoic Acid on SREBP1-Mediated Lipogenic Enzyme Expression in Primary Hepatocytes


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
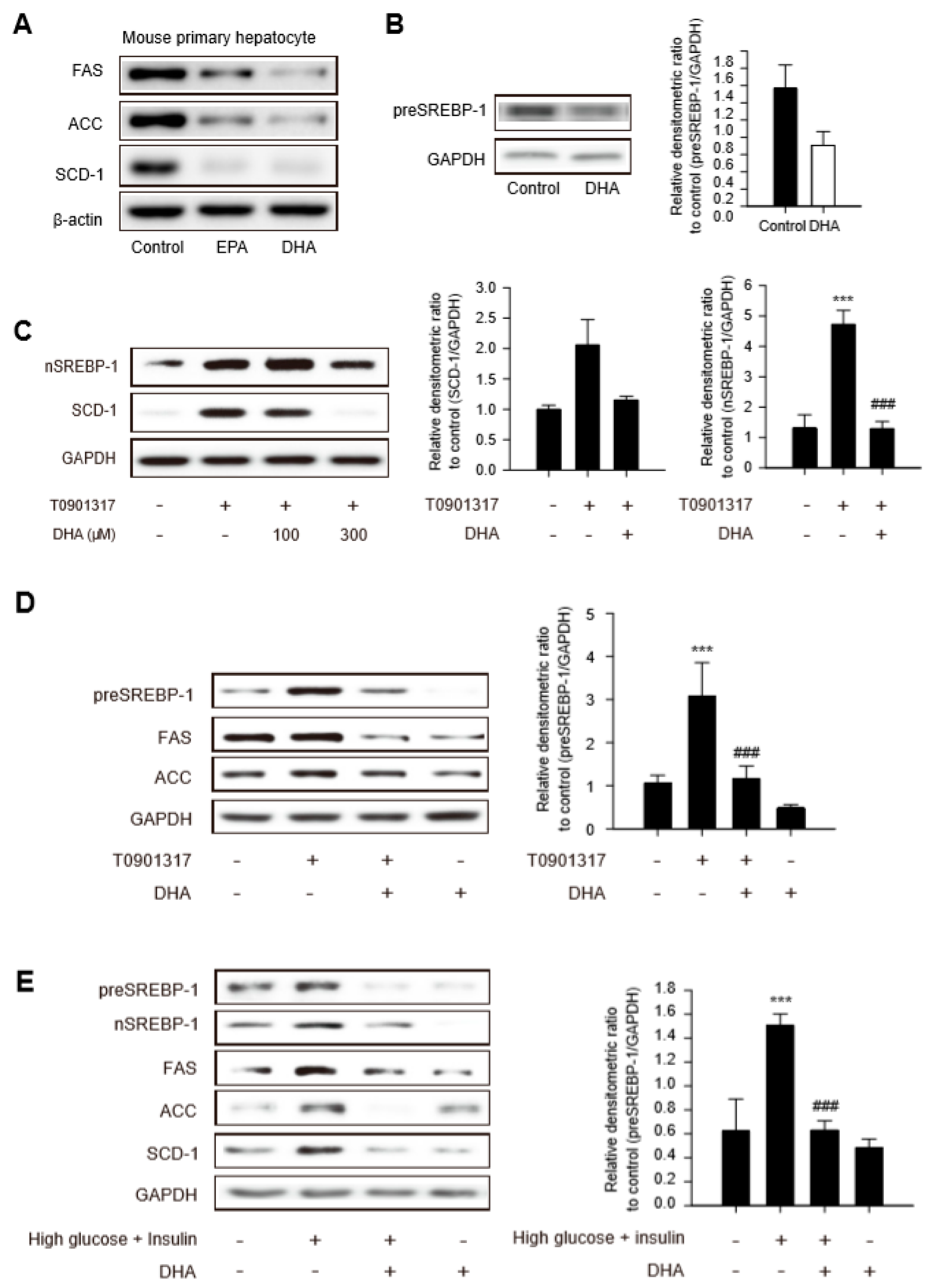
2.1. Downregulation of Sterol Regulatory Element Binding Protein (SREBP)-1-Dependent Lipogenic Enzyme Expression by DHA in Primary Hepatocytes
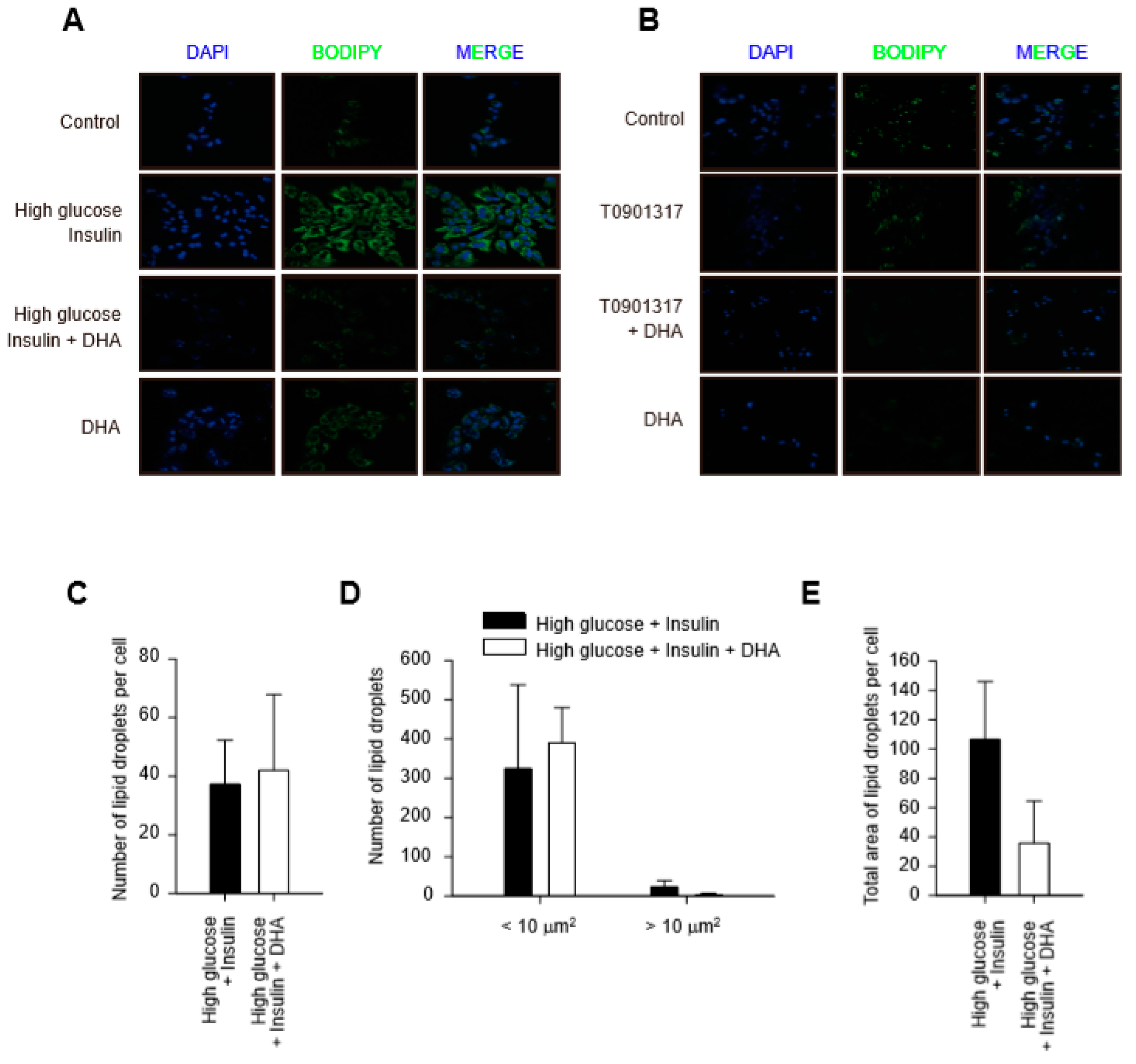
2.2. DHA-Induced Reduction of Total Area of Lipid Droplets in Hepatocytes
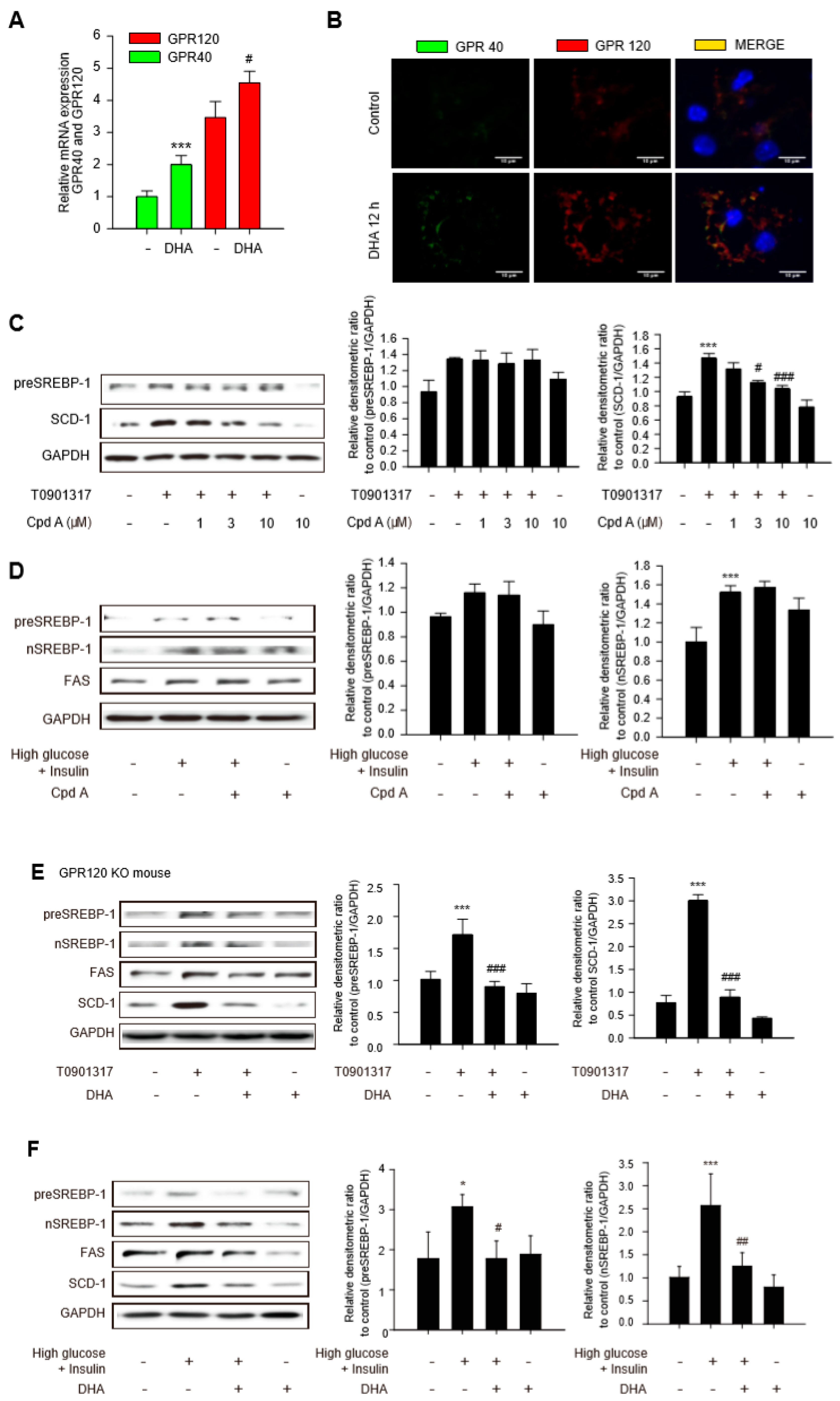
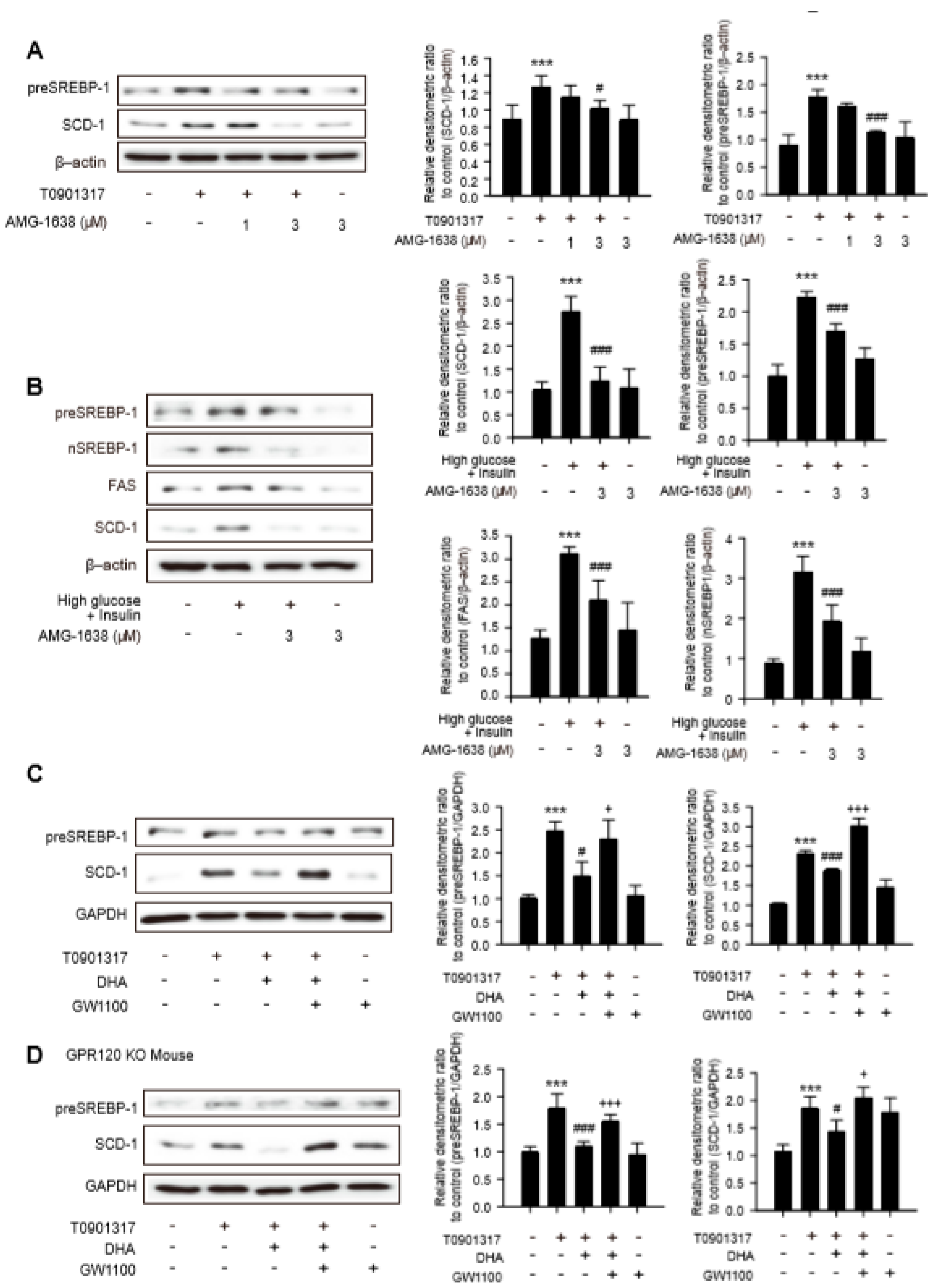
2.3. Limited Role of GPR 120 in Antilipogenic Effects of DHA in Hepatocytes
2.4. Involvement of GPR40 in Antilipogenesis Effect of DHA
3. Discussion
4. Materials and Methods
4.1. Reagent and Antibodies
4.2. Cell Culture
4.3. Animals
4.4. Isolation of Primary Hepatocytes
4.6. Immunoblot Analysis
4.7. Hematoxylin and Eosin Staining
4.8. Detection of Lipid Droplets (LDs) by Confocal Fluorescence Scanning Microscopy
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
| ACC | Acetyl-Coenzyme A carboxylase |
| ALT | Alanine transaminase |
| AMPK | Adenosine Monophosphate-activated protein kinase |
| AST | Aspartate transaminase |
| CpdA | Compound A |
| DHA | Docosahexaenoic acid |
| FAS | Fatty acid synthase |
| FFA | Free fatty acid |
| GPR120 | G-protein-coupled receptor 120 |
| GPR40 | G-protein-coupled receptor 40 |
| GADPH | Glyceraldehyde-3-phosphate dehydrogenase |
| GO | Gene Ontology |
| LDs | Lipid droplets |
| LXR | Liver X receptor |
| NAFLD | Nonalcoholic fatty liver disease |
| NASH | Nonalcoholic steatohepatitis |
| ND | Not detected |
| SCD-1 | Stearoyl-Coenzyme A desaturase-1 |
| SREBP-1 | Sterol regulatory element binding protein-1 |
| TG | Triglyceride |
| T090 | T0901317 |
References
- Perumpail, B.J.; Khan, M.A.; Yoo, E.R.; Cholankeril, G.; Kim, D.; Ahmed, A. Clinical epidemiology and disease burden of nonalcoholic fatty liver disease. World J. Gastroenterol 2017, 23, 8263–8276. [Google Scholar] [CrossRef] [PubMed]
- Bedogni, G.; Nobili, V.; Tiribelli, C. Epidemiology of fatty liver: An update. World J. Gastroenterol 2014, 20, 9050–9054. [Google Scholar] [PubMed]
- Fan, J.G.; Kim, S.U.; Wong, V.W. New trends on obesity and nafld in asia. J. Hepatol. 2017, 67, 862–873. [Google Scholar] [CrossRef]
- Calzadilla Bertot, L.; Adams, L.A. The natural course of non-alcoholic fatty liver disease. Int. J. Mol. Sci. 2016, 17. [Google Scholar] [CrossRef]
- Chalasani, N.; Younossi, Z.; Lavine, J.E.; Charlton, M.; Cusi, K.; Rinella, M.; Harrison, S.A.; Brunt, E.M.; Sanyal, A.J. The diagnosis and management of nonalcoholic fatty liver disease: Practice guidance from the american association for the study of liver diseases. Hepatology 2018, 67, 328–357. [Google Scholar] [CrossRef]
- Clarke, T.C.; Black, L.I.; Stussman, B.J.; Barnes, P.M.; Nahin, R.L. Trends in the use of complementary health approaches among adults: United states, 2002–2012. Natl. Health Stat. Rep. 2015, 10, 1–16. [Google Scholar]
- Backes, J.; Anzalone, D.; Hilleman, D.; Catini, J. The clinical relevance of omega-3 fatty acids in the management of hypertriglyceridemia. Lipids Health Dis. 2016, 15, 118. [Google Scholar] [CrossRef]
- Parker, H.M.; Johnson, N.A.; Burdon, C.A.; Cohn, J.S.; O’Connor, H.T.; George, J. Omega-3 supplementation and non-alcoholic fatty liver disease: A systematic review and meta-analysis. J. Hepatol. 2012, 56, 944–951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Briscoe, C.P.; Tadayyon, M.; Andrews, J.L.; Benson, W.G.; Chambers, J.K.; Eilert, M.M.; Ellis, C.; Elshourbagy, N.A.; Goetz, A.S.; Minnick, D.T. The orphan g protein-coupled receptor gpr40 is activated by medium and long chain fatty acids. J. Biol. Chem. 2003, 278, 11303–11311. [Google Scholar] [CrossRef]
- Oh, D.Y.; Talukdar, S.; Bae, E.J.; Imamura, T.; Morinaga, H.; Fan, W.; Li, P.; Lu, W.J.; Watkins, S.M.; Olefsky, J.M. Gpr120 is an omega-3 fatty acid receptor mediating potent anti-inflammatory and insulin-sensitizing effects. Cell 2010, 142, 687–698. [Google Scholar] [CrossRef]
- Oh, D.Y.; Walenta, E.; Akiyama, T.E.; Lagakos, W.S.; Lackey, D.; Pessentheiner, A.R.; Sasik, R.; Hah, N.; Chi, T.J.; Cox, J.M.; et al. A gpr120-selective agonist improves insulin resistance and chronic inflammation in obese mice. Nat. Med. 2014, 20, 942–947. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Meng, X.; Xu, J.; Huang, X.; Li, H.; Li, G.; Wang, S.; Man, Y.; Tang, W.; Li, J. Gpr40 agonist ameliorates liver x receptor-induced lipid accumulation in liver by activating ampk pathway. Sci. Rep. 2016, 6, 25237. [Google Scholar] [CrossRef] [PubMed]
- Eberle, D.; Hegarty, B.; Bossard, P.; Ferre, P.; Foufelle, F. Srebp transcription factors: Master regulators of lipid homeostasis. Biochimie 2004, 86, 839–848. [Google Scholar] [CrossRef] [PubMed]
- Gluchowski, N.L.; Becuwe, M.; Walther, T.C.; Farese, R.V., Jr. Lipid droplets and liver disease: From basic biology to clinical implications. Nat. Rev. Gastroenterol Hepatol. 2017, 14, 343–355. [Google Scholar] [CrossRef]
- Shi, X.; Li, J.; Zou, X.; Greggain, J.; Rødkær, S.V.; Færgeman, N.J.; Liang, B.; Watts, J.L. Regulation of lipid droplet size and phospholipid composition by stearoyl-coa desaturase. J. Lipid Res. 2013, 54, 2504–2514. [Google Scholar] [CrossRef]
- Steneberg, P.; Rubins, N.; Bartoov-Shifman, R.; Walker, M.D.; Edlund, H. The ffa receptor gpr40 links hyperinsulinemia, hepatic steatosis, and impaired glucose homeostasis in mouse. Cell Metab. 2005, 1, 245–258. [Google Scholar] [CrossRef]
- Huang, G.-M.; Jiang, Q.-H.; Cai, C.; Qu, M.; Shen, W. Scd1 negatively regulates autophagy-induced cell death in human hepatocellular carcinoma through inactivation of the ampk signaling pathway. Cancer Lett. 2015, 358, 180–190. [Google Scholar] [CrossRef]
- Paton, C.M.; Ntambi, J.M. Biochemical and physiological function of stearoyl-coa desaturase. Am. J. Physiol. Endocrinol. Metab. 2009, 297, E28–E37. [Google Scholar] [CrossRef]
- Wang, Y.; Kurdi-Haidar, B.; Oram, J.F. Lxr-mediated activation of macrophage stearoyl-coa desaturase generates unsaturated fatty acids that destabilize abca1. J. Lipid Res. 2004, 45, 972–980. [Google Scholar] [CrossRef]
- Brown, S.P.; Dransfield, P.J.; Vimolratana, M.; Jiao, X.; Zhu, L.; Pattaropong, V.; Sun, Y.; Liu, J.; Luo, J.; Zhang, J.; et al. Discovery of am-1638: A potent and orally bioavailable gpr40/ffa1 full agonist. Acs. Med. Chem. Lett. 2012, 3, 726–730. [Google Scholar] [CrossRef]
- Jacobson, T.A.; Glickstein, S.B.; Rowe, J.D.; Soni, P.N. Effects of eicosapentaenoic acid and docosahexaenoic acid on low-density lipoprotein cholesterol and other lipids: A review. J. Clin. Lipidol. 2012, 6, 5–18. [Google Scholar] [CrossRef]
- Wei, M.Y.; Jacobson, T.A. Effects of eicosapentaenoic acid versus docosahexaenoic acid on serum lipids: A systematic review and meta-analysis. Curr. Atheroscler. Rep. 2011, 13, 474–483. [Google Scholar] [CrossRef]
- Gotoh, C.; Hong, Y.H.; Iga, T.; Hishikawa, D.; Suzuki, Y.; Song, S.H.; Choi, K.C.; Adachi, T.; Hirasawa, A.; Tsujimoto, G.; et al. The regulation of adipogenesis through gpr120. Biochem. Biophys. Res. Commun. 2007, 354, 591–597. [Google Scholar] [CrossRef]
- Hirasawa, A.; Tsumaya, K.; Awaji, T.; Katsuma, S.; Adachi, T.; Yamada, M.; Sugimoto, Y.; Miyazaki, S.; Tsujimoto, G. Free fatty acids regulate gut incretin glucagon-like peptide-1 secretion through gpr120. Nat. Med. 2005, 11, 90–94. [Google Scholar] [CrossRef]
- Nobili, V.; Carpino, G.; Alisi, A.; De Vito, R.; Franchitto, A.; Alpini, G.; Onori, P.; Gaudio, E. Role of docosahexaenoic acid treatment in improving liver histology in pediatric nonalcoholic fatty liver disease. Plos ONE 2014, 9, e88005. [Google Scholar] [CrossRef]
- Abaraviciene, S.M.; Muhammed, S.J.; Amisten, S.; Lundquist, I.; Salehi, A. Gpr40 protein levels are crucial to the regulation of stimulated hormone secretion in pancreatic islets. Lessons from spontaneous obesity-prone and non-obese type 2 diabetes in rats. Mol. Cell. Endocrinol. 2013, 381, 150–159. [Google Scholar] [CrossRef]
- Hudson, B.D.; Shimpukade, B.; Mackenzie, A.E.; Butcher, A.J.; Pediani, J.D.; Christiansen, E.; Heathcote, H.; Tobin, A.B.; Ulven, T.; Milligan, G. The pharmacology of tug-891, a potent and selective agonist of the free fatty acid receptor 4 (ffa4/gpr120), demonstrates both potential opportunity and possible challenges to therapeutic agonism. Mol. Pharm. 2013, 84, 710–725. [Google Scholar] [CrossRef]
- Kang, S.; Huang, J.; Lee, B.-K.; Jung, Y.-S.; Im, E.; Koh, J.-M.; Im, D.-S. Omega-3 polyunsaturated fatty acids protect human hepatoma cells from developing steatosis through FFA4 (GPR120). Biochim. Biophys. Acta Mol. Cell Biol. Lipids. 2018, 1863, 105–116. [Google Scholar] [CrossRef]
- Itoh, Y.; Kawamata, Y.; Harada, M.; Kobayashi, M.; Fujii, R.; Fukusumi, S.; Ogi, K.; Hosoya, M.; Tanaka, Y.; Uejima, H. Free fatty acids regulate insulin secretion from pancreatic β cells through gpr40. Nature 2003, 422, 173. [Google Scholar] [CrossRef]
- Feng, X.T.; Leng, J.; Xie, Z.; Li, S.L.; Zhao, W.; Tang, Q.L. Gpr40: A therapeutic target for mediating insulin secretion (review). Int. J. Mol. Med. 2012, 30, 1261–1266. [Google Scholar] [CrossRef]
- Syed, I.; Lee, J.; Moraes-Vieira, P.M.; Donaldson, C.J.; Sontheimer, A.; Aryal, P.; Wellenstein, K.; Kolar, M.J.; Nelson, A.T.; Siegel, D.; et al. Palmitic acid hydroxystearic acids activate gpr40, which is involved in their beneficial effects on glucose homeostasis. Cell Metab. 2018, 27, 419–427.e414. [Google Scholar] [CrossRef] [PubMed]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
On, S.; Kim, H.Y.; Kim, H.S.; Park, J.; Kang, K.W. Involvement of G-Protein-Coupled Receptor 40 in the Inhibitory Effects of Docosahexaenoic Acid on SREBP1-Mediated Lipogenic Enzyme Expression in Primary Hepatocytes. Int. J. Mol. Sci. 2019, 20, 2625. https://doi.org/10.3390/ijms20112625
On S, Kim HY, Kim HS, Park J, Kang KW. Involvement of G-Protein-Coupled Receptor 40 in the Inhibitory Effects of Docosahexaenoic Acid on SREBP1-Mediated Lipogenic Enzyme Expression in Primary Hepatocytes. International Journal of Molecular Sciences. 2019; 20(11):2625. https://doi.org/10.3390/ijms20112625
Chicago/Turabian StyleOn, Seungtae, Hyun Young Kim, Hyo Seon Kim, Jeongwoo Park, and Keon Wook Kang. 2019. "Involvement of G-Protein-Coupled Receptor 40 in the Inhibitory Effects of Docosahexaenoic Acid on SREBP1-Mediated Lipogenic Enzyme Expression in Primary Hepatocytes" International Journal of Molecular Sciences 20, no. 11: 2625. https://doi.org/10.3390/ijms20112625
APA StyleOn, S., Kim, H. Y., Kim, H. S., Park, J., & Kang, K. W. (2019). Involvement of G-Protein-Coupled Receptor 40 in the Inhibitory Effects of Docosahexaenoic Acid on SREBP1-Mediated Lipogenic Enzyme Expression in Primary Hepatocytes. International Journal of Molecular Sciences, 20(11), 2625. https://doi.org/10.3390/ijms20112625