Insect ATP-Binding Cassette (ABC) Transporters: Roles in Xenobiotic Detoxification and Bt Insecticidal Activity





Abstract
:1. Introduction
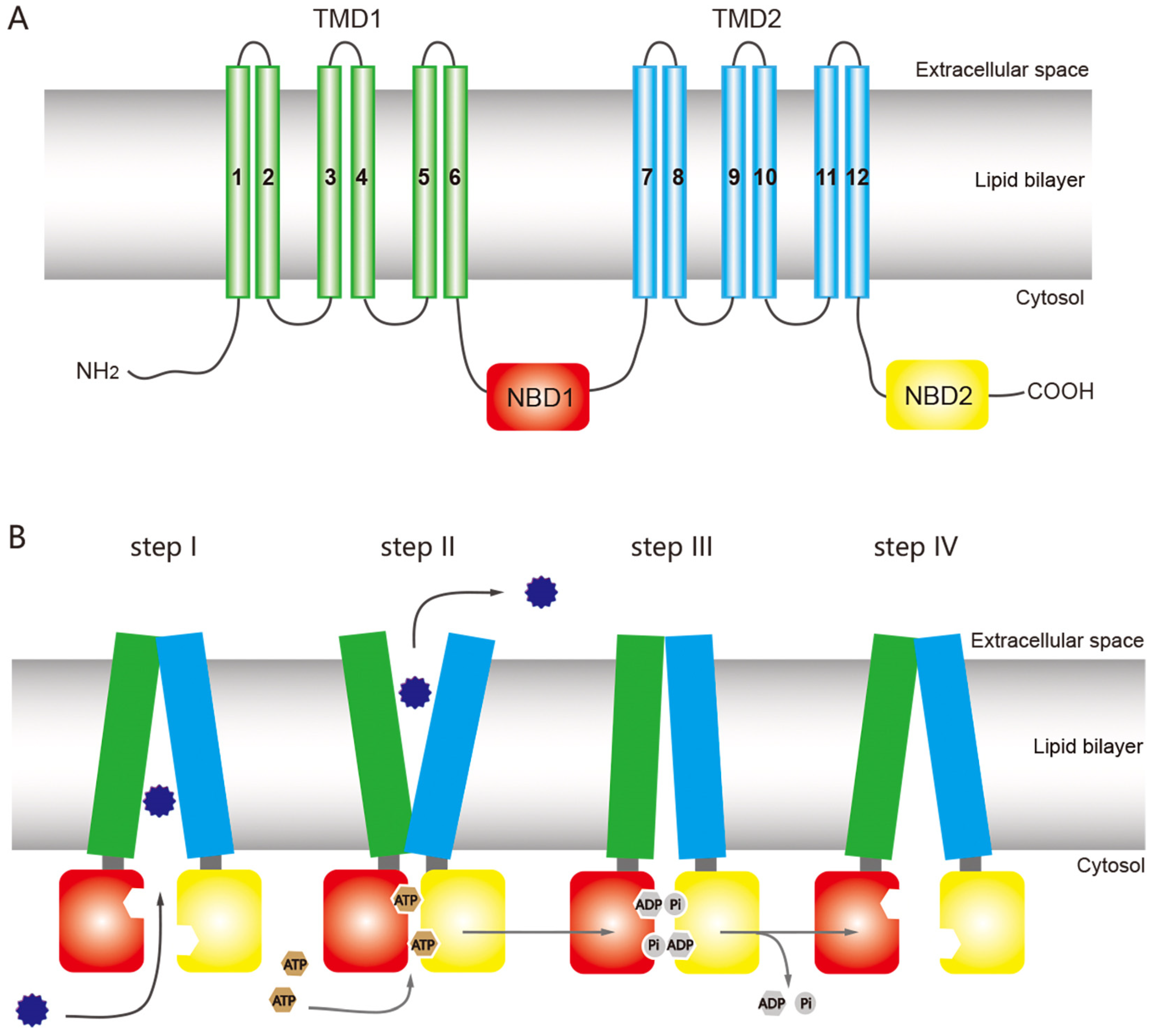
2. Structure and Mechanism of ABC Transporters
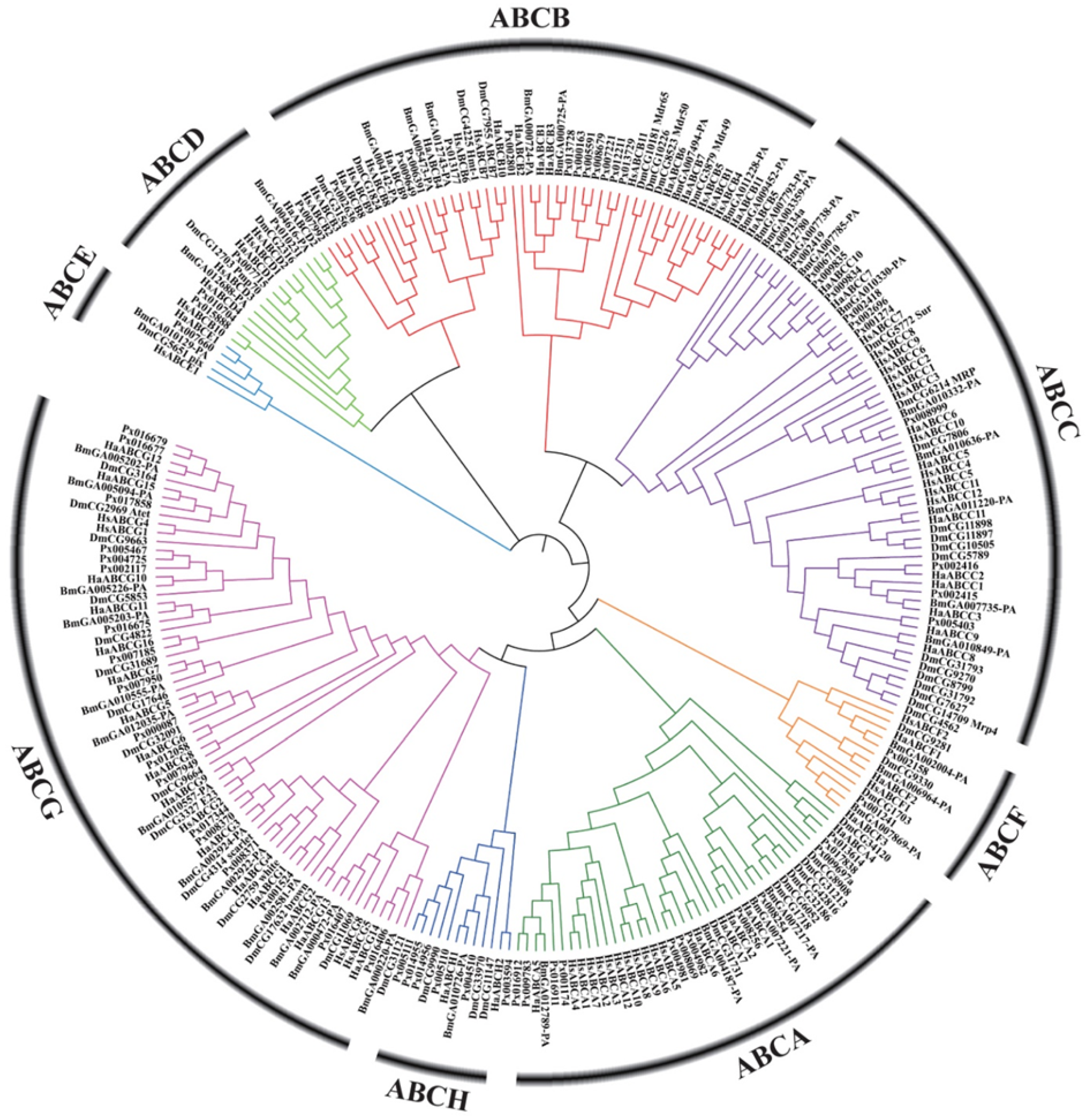
3. ABC Transporter Subfamilies in Insects
3.1. The ABCA Subfamily
3.2. The ABCB Subfamily
3.3. The ABCC Subfamily
3.4. The ABCD, ABCE and ABCF Subfamilies
3.5. The ABCG Subfamily
3.6. The ABCH Subfamily
4. ABC Transporters in Xenobiotic Detoxification by Insects
4.1. Insecticide Transport and Detoxification
4.2. Detoxification of Plant Secondary Metabolites
5. ABC Transporter Roles in Bt Insecticidal Activity
6. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Dean, M.; Rzhetsky, A.; Allikmets, R. The human ATP-binding cassette (ABC) transporter superfamily. Genome Res. 2001, 11, 1156–1166. [Google Scholar] [CrossRef] [PubMed]
- Dassa, E.; Bouige, P. The ABC of ABCS: A phylogenetic and functional classification of ABC systems in living organisms. Res. Microbiol. 2001, 152, 211–229. [Google Scholar] [CrossRef]
- Holland, I.B.; Cole, S.P.; Kuchler, K.; Higgins, C.F. ABC Proteins—From Bacteria to Man; Academic Press: Cambridge, MA, USA, 2003; p. 647. [Google Scholar]
- Berger, E.A. Different mechanisms of energy coupling for the active transport of proline and glutamine in Escherichia coli. Proc. Natl. Acad. Sci. USA 1973, 70, 1514–1518. [Google Scholar] [CrossRef] [PubMed]
- Berger, E.A.; Heppel, L.A. Different mechanisms of energy coupling for the shock-sensitive and shock-resistant amino acid permeases of Escherichia coli. J. Biol. Chem. 1974, 249, 7747–7755. [Google Scholar] [PubMed]
- Higgins, C.F.; Haag, P.D.; Nikaido, K.; Ardeshir, F.; Garcia, G.; Ames, G.F. Complete nucleotide sequence and identification of membrane components of the histidine transport operon of S. typhimurium. Nature 1982, 298, 723–727. [Google Scholar] [CrossRef] [PubMed]
- Gilson, E.; Higgins, C.F.; Hofnung, M.; Ames, F.L.; Nikaido, H. Extensive homology between membrane-associated components of histidine and maltose transport systems of Salmonella typhimurium and Escherichia coli. J. Biol. Chem. 1982, 257, 9915. [Google Scholar]
- Juliano, R.L.; Ling, V. A surface glycoprotein modulating drug permeability in Chinese hamster ovary cell mutants. Biochim. Biophys. Acta 1976, 455, 152–162. [Google Scholar] [CrossRef]
- Riordan, J.R.; Deuchars, K.; Kartner, N.; Alon, N.; Trent, J.; Ling, V. Amplification of P-glycoprotein genes in multidrug-resistant mammalian cell lines. Nature 1985, 316, 817–819. [Google Scholar] [CrossRef]
- Hyde, S.C.; Emsley, P.; Hartshorn, M.J.; Mimmack, M.M.; Gileadi, U.; Pearce, S.R.; Gallagher, M.P.; Gill, D.R.; Hubbard, R.E.; Higgins, C.F. Structural model of ATP-binding proteing associated with cystic fibrosis, multidrug resistance and bacterial transport. Nature 1990, 346, 362–365. [Google Scholar] [CrossRef]
- Merzendorfer, H. Chapter one–ABC transporters and their role in protecting insects from pesticides and their metabolites. Adv. Insect Physiol. 2014, 46, 1–72. [Google Scholar]
- Rees, D.C.; Johnson, E.; Lewinson, O. ABC transporters: The power to change. Nat. Rev. Mol. Cell Biol. 2009, 10, 218–227. [Google Scholar] [CrossRef] [PubMed]
- Higgins, C.F. ABC transporters: From microorganisms to man. Annu. Rev. Cell Biol. 1991, 8, 67–113. [Google Scholar] [CrossRef] [PubMed]
- Dermauw, W.; Van Leeuwen, T. The ABC gene family in arthropods: Comparative genomics and role in insecticide transport and resistance. Insect Biochem. Mol. Biol. 2014, 45, 89–110. [Google Scholar] [CrossRef] [PubMed]
- Hopfner, K.P. Invited review: Architectures and mechanisms of ATP binding cassette proteins. Biopolymers 2016, 105, 492–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kartner, N.; Evernden-Porelle, D.; Bradley, G.; Ling, V. Detection of P-glycoprotein in multidrug-resistant cell lines by monoclonal antibodies. Nature 1985, 316, 820–823. [Google Scholar] [CrossRef] [PubMed]
- Lage, H. ABC-transporters: Implications on drug resistance from microorganisms to human cancers. Int. J. Antimicrob. Agents 2003, 22, 188–199. [Google Scholar] [CrossRef]
- Ewart, G.D.; Howells, A.J. ABC transporters involved in transport of eye pigment precursors in Drosophila melanogaster. Methods Enzymol. 1998, 292, 213–224. [Google Scholar]
- Tatematsu, K.; Yamamoto, K.; Uchino, K.; Narukawa, J.; Iizuka, T.; Banno, Y.; Katsuma, S.; Shimada, T.; Tamura, T.; Sezutsu, H.; et al. Positional cloning of silkworm white egg 2 (w-2) locus shows functional conservation and diversification of ABC transporters for pigmentation in insects. Genes Cells 2011, 16, 331–342. [Google Scholar] [CrossRef]
- Broehan, G.; Kroeger, T.; Lorenzen, M.; Merzendorfer, H. Functional analysis of the ATP-binding cassette (ABC) transporter gene family of Tribolium castaneum. BMC Genom. 2013, 14, 6. [Google Scholar] [CrossRef]
- Bariami, V.; Jones, C.M.; Poupardin, R.; Vontas, J.; Ranson, H. Gene amplification, ABC transporters and cytochrome P450s: Unraveling the molecular basis of pyrethroid resistance in the dengue vector, Aedes aegypti. PLoS Neglect. Trop. Dis. 2012, 6, e1692. [Google Scholar] [CrossRef]
- Bonizzoni, M.; Afrane, Y.; Dunn, W.A.; Atieli, F.K.; Zhou, G.; Zhong, D.; Li, J.; Githeko, A.; Yan, G. Comparative transcriptome analyses of deltamethrin-resistant and -susceptible Anopheles gambiae mosquitoes from Kenya by RNA-Seq. PLoS ONE 2012, 7, e44607. [Google Scholar] [CrossRef] [PubMed]
- Mamidala, P.; Wijeratne, A.J.; Wijeratne, S.; Kornacker, K.; Sudhamalla, B.; Rivera-Vega, L.J.; Hoelmer, A.; Meulia, T.; Jones, S.C.; Mittapalli, O. RNA-Seq and molecular docking reveal multi-level pesticide resistance in the bed bug. BMC Genom. 2012, 13, 6. [Google Scholar] [CrossRef] [PubMed]
- Pardo-López, L.; Soberón, M.; Bravo, A. Bacillus thuringiensis insecticidal three-domain Cry toxins: Mode of action, insect resistance and consequences for crop protection. FEMS Microbiol. Rev. 2012, 37, 3–22. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Zhang, T.; Liu, C.; Heckel, D.G.; Li, X.; Tabashnik, B.E.; Wu, K. Mis-splicing of the ABCC2 gene linked with Bt toxin resistance in Helicoverpa armigera. Sci. Rep. 2014, 4, 6184. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Wei, J.Z.; Liu, C.; Zhang, W.N.; Wang, B.J.; Niu, L.L.; Liang, G.M. Specific binding protein ABCC1 Is associated with Cry2Ab toxicity in Helicoverpa armigera. Front. Physiol. 2018, 9, 745. [Google Scholar] [CrossRef] [PubMed]
- Heckel, D.G. Learning the ABCs of Bt: ABC transporters and insect resistance to Bacillus thuringiensis provide clues to a crucial step in toxin mode of action. Pestic. Biochem. Phys. 2012, 104, 103–110. [Google Scholar] [CrossRef]
- Gott, R.C.; Kunkel, G.R.; Zobel, E.S.; Lovett, B.R.; Hawthorne, D.J. Implicating ABC transporters in insecticide resistance: Research strategies and a decision framework. J. Econ. Entomol. 2017, 110, 667–677. [Google Scholar] [CrossRef]
- Chang, G.; Roth, C.B. Structure of MsbA from E. coli: A homolog of the multidrug resistance ATP binding cassette (ABC) transporters. Science 2001, 293, 1793–1800. [Google Scholar] [CrossRef]
- Dawson, R.J.; Locher, K.P. Structure of a bacterial multidrug ABC transporter. Nature 2006, 443, 180–185. [Google Scholar] [CrossRef]
- Hvorup, R.N.; Goetz, B.A.; Niederer, M.; Hollenstein, K.; Perozo, E.; Locher, K.P. Asymmetry in the structure of the ABC transporter-binding protein complex BtuCD-BtuF. Science 2007, 317, 1387–1390. [Google Scholar] [CrossRef]
- Ter Beek, J.; Guskov, A.; Slotboom, D.J. Structural diversity of ABC transporters. J. Gen. Physiol. 2014, 143, 419–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holland, I.B.; Blight, M.A. ABC-ATPases, adaptable energy generators fuelling transmembrane movement of a variety of molecules in organisms from bacteria to humans. J. Mol. Biol. 1999, 293, 381–399. [Google Scholar] [CrossRef] [PubMed]
- Erkens, G.B.; Berntsson, R.P.; Fulyani, F.; Majsnerowska, M.; Vujicic-Zagar, A.; Ter Beek, J.; Poolman, B.; Slotboom, D.J. The structural basis of modularity in ECF-type ABC transporters. Nat. Struct. Mol. Biol. 2011, 18, 755–760. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Zhang, M.; Zhao, Q.; Yu, F.; Guo, H.; Wang, C.; He, F.; Ding, J.; Zhang, P. Crystal structure of a folate energy-coupling factor transporter from Lactobacillus brevis. Nature 2013, 497, 268–271. [Google Scholar] [CrossRef] [PubMed]
- Eitinger, T.; Rodionov, D.A.; Grote, M.; Schneider, E. Canonical and ECF-type ATP-binding cassette importers in prokaryotes: Diversity in modular organization and cellular functions. FEMS Microbiol. Rev. 2015, 35, 3–67. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Fu, G.; Pan, X.; Wu, J.; Gong, X.; Wang, J.; Shi, Y. Structure of a bacterial energy-coupling factor transporter. Sci. Found. China 2014, 497, 272–276. [Google Scholar] [CrossRef] [PubMed]
- Gottesman, M.M.; Ambudkar, S.V. Overview: ABC transporters and human disease. J. Bioenerg. Biomembr. 2001, 33, 453–458. [Google Scholar] [CrossRef] [PubMed]
- Higgins, C.F.; Linton, K.J. The ATP switch model for ABC transporters. Nat. Struct Mol. Biol. 2004, 11, 918–926. [Google Scholar] [CrossRef] [PubMed]
- Senior, A.E.; al-Shawi, M.K.; Urbatsch, I.L. The catalytic cycle of P-glycoprotein. FEBS Lett. 1995, 377, 285–289. [Google Scholar] [Green Version]
- Sauna, Z.E.; Kim, I.W.; Nandigama, K.; Kopp, S.; Chiba, P.; Ambudkar, S.V. Catalytic cycle of ATP hydrolysis by P-glycoprotein: Evidence for formation of the E.S reaction intermediate with ATP-gamma-S, a nonhydrolyzable analogue of ATP. Biochemistry 2007, 46, 13787–13799. [Google Scholar] [CrossRef]
- Siarheyeva, A.; Liu, R.; Sharom, F.J. Characterization of an asymmetric occluded state of P-glycoprotein with two bound nucleotides: Implications for catalysis. J. Biol. Chem. 2010, 285, 7575–7586. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.C.; Han, L.; Zhao, Y. Thermodynamics of ABC transporters. Protein Cell 2016, 7, 17–27. [Google Scholar] [CrossRef] [PubMed]
- George, A.M.; Jones, P.M. Perspectives on the structure-function of ABC transporters: The switch and constant contact models. Prog. Biophys. Mol. Biol. 2012, 109, 95–107. [Google Scholar] [CrossRef] [PubMed]
- Linton, K.J.; Higgins, C.F. Structure and function of ABC transporters: The ATP switch provides flexible control. Pflugers Arch. 2007, 453, 555–567. [Google Scholar] [CrossRef] [PubMed]
- Deeley, R.G.; Westlake, C.; Cole, S.P. Transmembrane transport of endo- and xenobiotics by mammalian ATP-binding cassette multidrug resistance proteins. Physiol. Rev. 2006, 86, 849–899. [Google Scholar] [CrossRef] [PubMed]
- Roth, C.W.; Holm, I.; Graille, M.; Dehoux, P.; Rzhetsky, A.; Wincker, P.; Weissenbach, J.; Brey, P.T. Identification of the Anopheles gambiae ATP-binding cassette transporter superfamily genes. Mol. Cells 2003, 15, 150–158. [Google Scholar]
- Sturm, A.; Cunningham, P.; Dean, M. The ABC transporter gene family of Daphnia pulex. BMC Genom. 2009, 10, 170. [Google Scholar] [CrossRef]
- Lee, S.H.; Kang, J.S.; Min, J.S.; Yoon, K.S.; Strycharz, J.P.; Johnson, R.; Mittapalli, O.; Margam, V.M.; Sun, W.; Li, H.M.; et al. Decreased detoxification genes and genome size make the human body louse an efficient model to study xenobiotic metabolism. Insect Mol. Biol. 2010, 19, 599–615. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Zhou, S.; Tian, L.; Guo, E.; Luan, Y.; Zhang, J.; Li, S. Genome-wide identification and characterization of ATP-binding cassette transporters in the silkworm, Bombyx mori. BMC Genom. 2011, 12, 491. [Google Scholar] [CrossRef]
- Xie, X.; Cheng, T.; Wang, G.; Duan, J.; Niu, W.; Xia, Q. Genome-wide analysis of the ATP-binding cassette (ABC) transporter gene family in the silkworm, Bombyx mori. Mol. Biol Rep. 2012, 39, 7281–7291. [Google Scholar] [CrossRef]
- Pearce, S.; Clarke, D.; East, P.; Elfekih, S.; Gordon, K.; Jermiin, L.; McGaughran, A.; Oakeshott, J.; Papanikolaou, A.; Perera, O. Genomic innovations, transcriptional plasticity and gene loss underlying the evolution and divergence of two highly polyphagous and invasive Helicoverpa pest species. BMC Biol. 2017, 15, 63. [Google Scholar] [CrossRef] [PubMed]
- Dermauw, W.; Osborne, E.J.; Clark, R.M.; Grbic, M.; Tirry, L.; Van Leeuwen, T. A burst of ABC genes in the genome of the polyphagous spider mite Tetranychus urticae. BMC Genom. 2013, 14, 317. [Google Scholar] [CrossRef] [PubMed]
- Strauss, A.S.; Wang, D.; Stock, M.; Gretscher, R.R.; Groth, M.; Boland, W.; Burse, A. Tissue-specific transcript profiling for ABC transporters in the sequestering larvae of the phytophagous leaf beetle Chrysomela populi. PLoS ONE 2014, 9, e98637. [Google Scholar] [CrossRef] [PubMed]
- Hull, J.J.; Chaney, K.; Geib, S.M.; Fabrick, J.A.; Brent, C.S.; Walsh, D.; Lavine, L.C. Transcriptome-based identification of ABC transporters in the western tarnished plant bug Lygus hesperus. PLoS ONE 2014, 9, e113046. [Google Scholar] [CrossRef]
- Carmona-Antonanzas, G.; Carmichael, S.N.; Heumann, J.; Taggart, J.B.; Gharbi, K.; Bron, J.E.; Bekaert, M.; Sturm, A. A Survey of the ATP-binding cassette (ABC) gene superfamily in the salmon louse (Lepeophtheirus salmonis). PLoS ONE 2015, 10, e0137394. [Google Scholar] [CrossRef]
- Sun, H.; Pu, J.; Chen, F.; Wang, J.; Han, Z. Multiple ATP-binding cassette transporters are involved in insecticide resistance in the small brown planthopper, Laodelphax striatellus. Insect Mol. Biol. 2017, 26, 343–355. [Google Scholar] [CrossRef]
- Tian, L.; Song, T.; He, R.; Yang, Z.; Xie, W.; Wu, Q.; Wang, S.; Zhou, X.; Zhang, Y.J.B.G. Genome-wide analysis of ATP-binding cassette (ABC) transporters in the sweetpotato whitefly, Bemisia tabaci. BMC Genom. 2017, 18, 330. [Google Scholar] [CrossRef]
- Guo, Z.J.; Kang, S.; Zhu, X.; Xia, J.X.; Wu, Q.J.; Wang, S.L.; Xie, W.; Zhang, Y.J. The novel ABC transporter ABCH1 is a potential target for RNAi-based insect pest control and resistance management. Sci. Rep. 2015, 5, 13728. [Google Scholar] [CrossRef]
- Qi, W.; Ma, X.; He, W.; Chen, W.; Zou, M.; Gurr, G.M.; Vasseur, L.; You, M. Characterization and expression profiling of ATP-binding cassette transporter genes in the diamondback moth, Plutella xylostella (L.). BMC Genom. 2016, 17, 760. [Google Scholar] [CrossRef]
- Bretschneider, A.; Heckel, D.G.; Vogel, H. Know your ABCs: Characterization and gene expression dynamics of ABC transporters in the polyphagous herbivore Helicoverpa armigera. Insect Biochem. Mol. Biol. 2016, 72, 1–9. [Google Scholar] [CrossRef]
- Jin, M.; Liao, C.; Chakrabarty, S.; Zheng, W.; Wu, K.; Xiao, Y. Transcriptional response of ATP-binding cassette (ABC) transporters to insecticides in the cotton bollworm, Helicoverpa armigera. Pestic. Biochem. Phys. 2019, 154, 46–59. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.H.; Fang, X.D.; Yang, P.C.; Jiang, X.T.; Jiang, F.; Zhao, D.J.; Li, B.L.; Cui, F.; Wei, J.N.; Ma, C.A.; et al. The locust genome provides insight into swarm formation and long-distance flight. Nat. Commun. 2014, 5, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Xu, Y.; Cui, F. Phylogenetic analysis of the ATP-binding cassette transporter family in three mosquito species. Pestic. Biochem. Physiol. 2016, 132, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Quazi, F.; Molday, R.S. Lipid transport by mammalian ABC proteins. Essays Biochem. 2011, 50, 265–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Z.; Zera, A.J. Differential lipid biosynthesis underlies a tradeoff between reproduction and flight capability in a wing-polymorphic cricket. Proc. Natl. Acad. Sci. USA 2002, 99, 16829–16834. [Google Scholar] [CrossRef] [Green Version]
- Zera, A.J.; Zhao, Z. Effect of a juvenile hormone analogue on lipid metabolism in a wing-polymorphic cricket: Implications for the endocrine-biochemical bases of life-history trade-offs. Physiol. Biochem. Zool. 2004, 77, 255–266. [Google Scholar] [CrossRef]
- Tay, W.T.; Mahon, R.J.; Heckel, D.G.; Walsh, T.K.; Downes, S.; James, W.J.; Lee, S.F.; Reineke, A.; Williams, A.K.; Gordon, K.H.J. Insect resistance to Bacillus thuringiensis toxin Cry2Ab is conferred by mutations in an ABC transporter subfamily a protein. PLoS Genet. 2015, 11, e1005534. [Google Scholar] [CrossRef] [PubMed]
- Metzendorf, C.; Wu, W.; Lind, M.I. Overexpression of Drosophila mitoferrin in l(2)mbn cells results in dysregulation of Fer1HCH expression. Biochem. J. 2009, 421, 463–471. [Google Scholar] [CrossRef]
- Sooksa-Nguan, T.; Yakubov, B.; Kozlovskyy, V.I.; Barkume, C.M.; Howe, K.J.; Thannhauser, T.W.; Rutzke, M.A.; Hart, J.J.; Kochian, L.V.; Rea, P.A.; et al. Drosophila ABC transporter, DmHMT-1, confers tolerance to cadmium. DmHMT-1 and its yeast homolog, SpHMT-1, are not essential for vacuolar phytochelatin sequestration. J. Biol. Chem. 2009, 284, 354–362. [Google Scholar] [CrossRef]
- Telonis-Scott, M.; Hallas, R.; McKechnie, S.W.; Wee, C.W.; Hoffmann, A.A. Selection for cold resistance alters gene transcript levels in Drosophila melanogaster. J. Insect Physiol. 2009, 55, 549–555. [Google Scholar] [CrossRef]
- Petschenka, G.; Pick, C.; Wagschal, V.; Dobler, S. Functional evidence for physiological mechanisms to circumvent neurotoxicity of cardenolides in an adapted and a non-adapted hawk-moth species. Proc. Biol. Sci. 2013, 280, 20123089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wanders, R.J.; Visser, W.F.; van Roermund, C.W.; Kemp, S.; Waterham, H.R. The peroxisomal ABC transporter family. Pflug. Arch. 2007, 453, 719–734. [Google Scholar] [CrossRef] [PubMed]
- Morita, M.; Imanaka, T. Peroxisomal ABC transporters: Structure, function and role in disease. Biochim. Biophys. Acta 2012, 1822, 1387–1396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kerr, I.D. Sequence analysis of twin ATP binding cassette proteins involved in translational control, antibiotic resistance, and ribonuclease L inhibition. Biochem. Biophys. Res. Commun. 2004, 315, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Tyzack, J.K.; Wang, X.; Belsham, G.J.; Proud, C.G. ABC50 interacts with eukaryotic initiation factor 2 and associates with the ribosome in an ATP-dependent manner. J. Biol. Chem. 2000, 275, 34131–34139. [Google Scholar] [CrossRef] [PubMed]
- Andersen, D.S.; Leevers, S.J. The essential Drosophila ATP-binding cassette domain protein, pixie, binds the 40 S ribosome in an ATP-dependent manner and is required for translation initiation. J. Biol. Chem. 2007, 282, 14752–14760. [Google Scholar] [CrossRef]
- Barthelme, D.; Dinkelaker, S.; Albers, S.V.; Londei, P.; Ermler, U.; Tampe, R. Ribosome recycling depends on a mechanistic link between the FeS cluster domain and a conformational switch of the twin-ATPase ABCE1. Proc. Natl. Acad. Sci. USA 2011, 108, 3228–3233. [Google Scholar] [CrossRef] [Green Version]
- Bisbal, C.; Martinand, C.; Silhol, M.; Lebleu, B.; Salehzada, T. Cloning and characterization of a RNAse L inhibitor. A new component of the interferon-regulated 2-5A pathway. J. Biol. Chem. 1995, 270, 13308–13317. [Google Scholar] [CrossRef]
- Mackenzie, S.M.; Brooker, M.R.; Gill, T.R.; Cox, G.B.; Howells, A.J.; Ewart, G.D. Mutations in the white gene of Drosophila melanogaster affecting ABC transporters that determine eye colouration. BBA Biomembr. 1999, 1419, 173–185. [Google Scholar] [CrossRef]
- Wang, L.; Kiuchi, T.; Fujii, T.; Daimon, T.; Li, M.; Banno, Y.; Kikuta, S.; Kikawada, T.; Katsuma, S.; Shimada, T. Mutation of a novel ABC transporter gene is responsible for the failure to incorporate uric acid in the epidermis of ok mutants of the silkworm, Bombyx mori. Insect Biochem. Mol. Biol. 2013, 43, 562–571. [Google Scholar] [CrossRef]
- Kovalchuk, A.; Driessen, A.J. Phylogenetic analysis of fungal ABC transporters. BMC Genom. 2010, 11, 177. [Google Scholar] [CrossRef] [PubMed]
- Verrier, P.J.; Bird, D.; Burla, B.; Dassa, E.; Forestier, C.; Geisler, M.; Klein, M.; Kolukisaoglu, U.; Lee, Y.; Martinoia, E.; et al. Plant ABC proteins—A unified nomenclature and updated inventory. Trends Plant. Sci. 2008, 13, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Popovic, M.; Zaja, R.; Loncar, J.; Smital, T. A novel ABC transporter The first insight into zebrafish (Danio rerio) ABCH1. Mar. Environ. Res. 2010, 69, S11–S13. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Li, Q.; Liu, Z. Genome-wide identification, characterization and phylogenetic analysis of 50 catfish ATP-binding cassette (ABC) transporter genes. PLoS ONE 2013, 8, e63895. [Google Scholar] [CrossRef] [PubMed]
- Dean, M.; Annilo, T. Evolution of the ATP-binding cassette (ABC) transporter superfamily in vertebrates. Annu. Rev. Genomics Hum. Genet. 2005, 6, 123–142. [Google Scholar] [CrossRef] [PubMed]
- Jeong, C.B.; Kim, B.M.; Kang, H.M.; Choi, I.Y.; Rhee, J.S.; Lee, J.S. Marine medaka ATP-binding cassette (ABC) superfamily and new insight into teleost Abch nomenclature. Sci. Rep. 2015, 5, 15409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Z.; Wang, Y.; Zhao, X.; Liu, X.; Ma, E.; Moussian, B.; Zhang, J. The ABC transporter ABCH-9C is needed for cuticle barrier construction in Locusta migratoria. Insect Biochem. Mol. Biol. 2017, 87, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Zuber, R.; Norum, M.; Wang, Y.; Oehl, K.; Gehring, N.; Accardi, D.; Bartozsewski, S.; Berger, J.; Flotenmeyer, M.; Moussian, B. The ABC transporter Snu and the extracellular protein Snsl cooperate in the formation of the lipid-based inward and outward barrier in the skin of Drosophila. Eur. J. Cell Biol. 2018, 97, 90–101. [Google Scholar] [CrossRef]
- Strycharz, J.P.; Lao, A.; Li, H.M.; Qiu, X.H.; Lee, S.H.; Sun, W.L.; Yoon, K.S.; Doherty, J.J.; Pittendrigh, B.R.; Clark, J.M. Resistance in the highly DDT-resistant 91-R strain of Drosophila melanogaster involves decreased penetration, increased metabolism, and direct excretion. Pestic. Biochem. Phys. 2013, 107, 207–217. [Google Scholar] [CrossRef]
- Granada, Y.; Mejia-Jaramillo, A.M.; Strode, C.; Triana-Chavez, O. A point mutation V419L in the sodium channel gene from natural populations of Aedes aegypti is involved in resistance to lambda-cyhalothrin in Colombia. Insects 2018, 9, 23. [Google Scholar] [CrossRef]
- Li, X.; Schuler, M.A.; Berenbaum, M.R. Molecular mechanisms of metabolic resistance to synthetic and natural xenobiotics. Annu. Rev. Entomol. 2007, 52, 231–253. [Google Scholar] [CrossRef] [PubMed]
- Van, L.T.; Vontas, J.; Tsagkarakou, A.; Dermauw, W.; Tirry, L. Acaricide resistance mechanisms in the two-spotted spider mite Tetranychus urticae and other important Acari: A review. Insect Biochem. Mol. Biol. 2010, 40, 563–572. [Google Scholar]
- Buss, D.S.; Callaghan, A. Interaction of pesticides with p-glycoprotein and other ABC proteins: A survey of the possible importance to insecticide, herbicide and fungicide resistance. Pestic. Biochem. Phys. 2008, 90, 141–153. [Google Scholar] [CrossRef]
- Nakata, K.; Tanaka, Y.; Nakano, T.; Adachi, T.; Tanaka, H.; Kaminuma, T.; Ishikawa, T. Nuclear receptor-mediated transcriptional regulation in phase I, II, and III xenobiotic metabolizing systems. Drug Metab. Pharmacokinet. 2006, 21, 437–457. [Google Scholar] [CrossRef]
- Sykiotis, G.P.; Bohmann, D. Stress-activated cap’n’collar transcription factors in aging and human disease. Sci. Signal. 2010, 3, re3. [Google Scholar] [CrossRef] [PubMed]
- Kalsi, M.; Palli, S.R. Cap n collar transcription factor regulates multiple genes coding for proteins involved in insecticide detoxification in the red flour beetle, Tribolium castaneum. Insect Biochem. Mol. Biol. 2017, 90, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Sharom, F.J. The P-glycoprotein multidrug transporter. Essays Biochem. 2011, 50, 161–178. [Google Scholar] [CrossRef] [Green Version]
- Figueiramansur, J.; Ferreirapereira, A.; Mansur, J.F.; Franco, T.A.; Alvarenga, E.S.; Sorgine, M.H.; Neves, B.C.; Melo, A.C.; Leal, W.S.; Masuda, H. Silencing of P-glycoprotein increases mortality in temephos-treated Aedes aegypti larvae. Insect Mol. Biol. 2013, 22, 648–658. [Google Scholar] [CrossRef]
- Aurade, R.M.; Jayalakshmi, S.K.; Sreeramulu, K. P-glycoprotein ATPase from the resistant pest, Helicoverpa armigera: Purification, characterization and effect of various insecticides on its transport function. Biochim. Biophys. Acta 2010, 1798, 1135–1143. [Google Scholar] [CrossRef]
- Luo, L.; Sun, Y.J.; Wu, Y.J. Abamectin resistance in Drosophila is related to increased expression of P-glycoprotein via the dEGFR and dAkt pathways. Insect Biochem. Mol. Biol. 2013, 43, 627–634. [Google Scholar] [CrossRef]
- Mayer, F.; Mayer, N.; Chinn, L.; Pinsonneault, R.L.; Kroetz, D.; Bainton, R.J. Evolutionary conservation of vertebrate blood-brain barrier chemoprotective mechanisms in Drosophila. J. Neurosci. 2009, 29, 3538–3550. [Google Scholar] [CrossRef] [PubMed]
- Gellatly, K.J.; Yoon, K.S.; Doherty, J.J.; Sun, W.L.; Pittendrigh, B.R.; Clark, J.M. RNAi validation of resistance genes and their interactions in the highly DDT-resistant 91-R strain of Drosophila melanogaster. Pestic. Biochem. Phys. 2015, 121, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Seong, K.M.; Sun, W.; Clark, J.M.; Pittendrigh, B.R. Splice form variant and amino acid changes in MDR49 confers DDT resistance in transgenic Drosophila. Sci. Rep. 2016, 6, 23355. [Google Scholar] [CrossRef]
- Denecke, S.; Fusetto, R.; Batterham, P. Describing the role of Drosophila melanogaster ABC transporters in insecticide biology using CRISPR-Cas9 knockouts. Insect Biochem. Mol. Biol. 2017, 91, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Buchon, N.; Scott, J.G. Mdr65 decreases toxicity of multiple insecticides in Drosophila melanogaster. Insect Biochem. Mol. Biol. 2017, 89, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Labbe, R.; Caveney, S.; Donly, C. Expression of multidrug resistance proteins is localized principally to the Malpighian tubules in larvae of the cabbage looper moth, Trichoplusia ni. J. Exp. Biol. 2011, 214, 937–944. [Google Scholar] [CrossRef] [PubMed]
- Yang, N.; Xie, W.; Jones, C.; Bass, C.; Jiao, X.; Yang, X.; Liu, B.; Li, R.; Zhang, Y. Transcriptome profiling of the whitefly Bemisia tabaci reveals stage-specific gene expression signatures for thiamethoxam resistance. Insect Mol. Biol. 2013, 22, 485–496. [Google Scholar] [CrossRef]
- Yang, N.; Xie, W.; Yang, X.; Wang, S.L.; Wu, Q.J.; Li, R.M.; Pan, H.P.; Liu, B.M.; Shi, X.B.; Fang, Y.; et al. Transcriptomic and Proteomic Responses of Sweetpotato Whitefly, Bemisia tabaci, to Thiamethoxam. PLoS ONE 2013, 8, e61820. [Google Scholar] [CrossRef]
- He, W.; You, M.; Vasseur, L.; Yang, G.; Xie, M.; Cui, K.; Bai, J.; Liu, C.; Li, X.; Xu, X.; et al. Developmental and insecticide-resistant insights from the de novo assembled transcriptome of the diamondback moth, Plutella xylostella. Genomics 2012, 99, 169–177. [Google Scholar] [CrossRef]
- You, M.; Yue, Z.; He, W.; Yang, X.; Yang, G.; Xie, M.; Zhan, D.; Baxter, S.W.; Vasseur, L.; Gurr, G.M. A heterozygous moth genome provides insights into herbivory and detoxification. Nat. Genet. 2013, 45, 220–225. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Gellatly, K.J.; Lueke, B.; Kohler, M.; Nauen, R.; Murenzi, E.; Yoon, K.S.; Clark, J.M. Detoxification of ivermectin by ATP binding cassette transporter C4 and cytochrome P450 monooxygenase 6CJ1 in the human body louse, Pediculus humanus humanus. Insect Mol. Biol. 2018, 27, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Petschenka, G.; Agrawal, A.A. How herbivores coopt plant defenses: Natural selection, specialization, and sequestration. Curr. Opin. Insect Sci. 2016, 14, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Groen, S.C.; LaPlante, E.R.; Alexandre, N.M.; Agrawal, A.A.; Dobler, S.; Whiteman, N.K. Multidrug transporters and organic anion transporting polypeptides protect insects against the toxic effects of cardenolides. Insect Biochem. Mol. Biol. 2017, 81, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Strauss, A.S.; Peters, S.; Boland, W.; Burse, A. ABC transporter functions as a pacemaker for sequestration of plant glucosides in leaf beetles. Elife 2013, 2, e01096. [Google Scholar] [CrossRef]
- Aldonza, M.B.; Hong, J.Y.; Lee, S.K. Paclitaxel-resistant cancer cell-derived secretomes elicit ABCB1-associated docetaxel cross-resistance and escape from apoptosis through FOXO3a-driven glycolytic regulation. Exp. Mol. Med. 2017, 49, e286. [Google Scholar] [CrossRef] [PubMed]
- Koenig, C.; Bretschneider, A.; Heckel, D.G.; Grosse-Wilde, E.; Hansson, B.S.; Vogel, H. The plastic response of Manduca sexta to host and non-host plants. Insect Biochem. Mol. Biol. 2015, 63, 72–85. [Google Scholar] [CrossRef] [PubMed]
- Mathew, L.G.; Ponnuraj, J.; Mallappa, B.; Chowdary, L.R.; Zhang, J.; Tay, W.T.; Walsh, T.K.; Gordon, K.H.J.; Heckel, D.G.; Downes, S.; et al. ABC transporter mis-splicing associated with resistance to Bt toxin Cry2Ab in laboratory- and field-selected pink bollworm. Sci. Rep. 2018, 8, 13531. [Google Scholar] [CrossRef] [PubMed]
- Tabashnik, B.E. ABCs of insect resistance to Bt. PLoS Genet. 2015, 11, e1005646. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, H.; Liu, S.; Liu, L.; Tay, W.T.; Walsh, T.K.; Yang, Y.; Wu, Y. CRISPR/Cas9 mediated genome editing of Helicoverpa armigera with mutations of an ABC transporter gene HaABCA2 confers resistance to Bacillus thuringiensis Cry2A toxins. Insect Biochem. Mol. Biol. 2017, 87, 147–153. [Google Scholar] [CrossRef]
- Tanaka, S.; Endo, H.; Adegawa, S.; Iizuka, A.; Imamura, K.; Kikuta, S.; Sato, R. Bombyx mori ABC transporter C2 structures responsible for the receptor function of Bacillus thuringiensis Cry1Aa toxin. Insect Biochem. Mol. Biol. 2017, 91, 44–54. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Li, M.; Islam, I.; You, L.; Wang, Y.; Li, Z.; Ling, L.; Zeng, B.; Xu, J.; Huang, Y.; et al. Allelic-specific expression in relation to Bombyx mori resistance to Bt toxin. Insect Biochem Mol. Biol. 2014, 54, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Coates, B.S.; Wang, Y.; Wang, Y.; Bai, S.; Wang, Z.; He, K. Down-regulation of aminopeptidase N and ABC transporter subfamily G transcripts in Cry1Ab and Cry1Ac resistant Asian corn borer, Ostrinia furnacalis (Lepidoptera: Crambidae). Int. J. Biol. Sci. 2017, 13, 835–851. [Google Scholar] [CrossRef] [PubMed]
- Gahan, L.J.; Pauchet, Y.; Vogel, H.; Heckel, D.G. An ABC Transporter Mutation Is Correlated with Insect Resistance to Bacillus thuringiensis Cry1Ac Toxin. PLoS Genet. 2010, 6, e1001248. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Kang, S.; Chen, D.; Wu, Q.; Wang, S.; Xie, W.; Zhu, X.; Baxter, S.W.; Zhou, X.; Jurat-Fuentes, J.L.; et al. MAPK signaling pathway alters expression of midgut ALP and ABCC genes and causes resistance to Bacillus thuringiensis Cry1Ac toxin in diamondback moth. PLoS Genet. 2015, 11, e1005124. [Google Scholar] [CrossRef] [PubMed]
- Baxter, S.W.; Badenes-Perez, F.R.; Morrison, A.; Vogel, H.; Crickmore, N.; Kain, W.; Wang, P.; Heckel, D.G.; Jiggins, C.D. Parallel evolution of Bacillus thuringiensis toxin resistance in Lepidoptera. Genetics 2011, 189, 675–679. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.; Gonzalez-Martinez, R.M.; Navarro-Cerrillo, G.; Chakroun, M.; Kim, Y.; Ziarsolo, P.; Blanca, J.; Canizares, J.; Ferre, J.; Herrero, S. ABCC transporters mediate insect resistance to multiple Bt toxins revealed by bulk segregant analysis. BMC Biol. 2014, 12, 46. [Google Scholar] [CrossRef]
- Li, J.; Ma, Y.; Yuan, W.; Xiao, Y.; Liu, C.; Wang, J.; Peng, J.; Peng, R.; Soberon, M.; Bravo, A.; et al. FOXA transcriptional factor modulates insect susceptibility to Bacillus thuringiensis Cry1Ac toxin by regulating the expression of toxin-receptor ABCC2 and ABCC3 genes. Insect Biochem. Mol. Biol. 2017, 88, 1–11. [Google Scholar] [CrossRef]
- Coates, B.S.; Siegfried, B.D. Linkage of an ABCC transporter to a single QTL that controls Ostrinia nubilalis larval resistance to the Bacillus thuringiensis Cry1Fa toxin. Insect Biochem. Mol. Biol. 2015, 63, 86–96. [Google Scholar] [CrossRef]
- Pauchet, Y.; Bretschneider, A.; Augustin, S.; Heckel, D.G. A P-glycoprotein is linked to resistance to the Bacillus thuringiensis Cry3Aa toxin in a Leaf BEETLE. Toxins Basel 2016, 8, 362. [Google Scholar] [CrossRef]
- Flagel, L.E.; Swarup, S.; Chen, M.; Bauer, C.; Wanjugi, H.; Carroll, M.; Hill, P.; Tuscan, M.; Bansal, R.; Flannagan, R.; et al. Genetic markers for western corn rootworm resistance to Bt toxin. G3 Bethesda 2015, 5, 399–405. [Google Scholar] [CrossRef]
- Bretschneider, A.; Heckel, D.G.; Pauchet, Y. Three toxins, two receptors, one mechanism: Mode of action of Cry1A toxins from Bacillus thuringiensis in Heliothis virescens. Insect Biochem. Mol. Biol. 2016, 76, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Stevens, T.; Song, S.; Bruning, J.B.; Choo, A.; Baxter, S.W. Expressing a moth abcc2 gene in transgenic Drosophila causes susceptibility to Bt Cry1Ac without requiring a cadherin-like protein receptor. Insect Biochem. Mol. Biol. 2017, 80, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Atsumi, S.; Miyamoto, K.; Yamamoto, K.; Narukawa, J.; Kawai, S.; Sezutsu, H.; Kobayashi, I.; Uchino, K.; Tamura, T.; Mita, K.; et al. Single amino acid mutation in an ATP-binding cassette transporter gene causes resistance to Bt toxin Cry1Ab in the silkworm, Bombyx mori. Proc. Natl. Acad. Sci. USA 2012, 109, E1591–E1598. [Google Scholar] [CrossRef] [PubMed]
- Endo, H.; Tanaka, S.; Adegawa, S.; Ichino, F.; Tabunoki, H.; Kikuta, S.; Sato, R. Extracellular loop structures in silkworm ABCC transporters determine their specificities for Bacillus thuringiensis cry toxins. J. Biol. Chem. 2018, 293, 8569–8577. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.J.; Kang, S.; Zhu, X.; Xia, J.X.; Wu, Q.J.; Wang, S.L.; Xie, W.; Zhang, Y.J. Down-regulation of a novel ABC transporter gene (Pxwhite) is associated with Cry1Ac resistance in the diamondback moth, Plutella xylostella (L.). Insect Biochem. Mol. Biol. 2015, 59, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Locher, K.P. Mechanistic diversity in ATP-binding cassette (ABC) transporters. Nat. Struct. Mol. Biol. 2016, 23, 487–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Khan, S.A.; Hasse, C.; Ruf, S.; Heckel, D.G.; Bock, R. Pest control. Full crop protection from an insect pest by expression of long double-stranded RNAs in plastids. Science 2015, 347, 991–994. [Google Scholar] [CrossRef]
- Malik, H.J.; Raza, A.; Amin, I.; Scheffler, J.A.; Scheffler, B.E.; Brown, J.K.; Mansoor, S. RNAi-mediated mortality of the whitefly through transgenic expression of double-stranded RNA homologous to acetylcholinesterase and ecdysone receptor in tobacco plants. Sci. Rep. 2016, 6, 38469. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Organisms | A | B | C | D | E | F | G | H | Total | References |
|---|---|---|---|---|---|---|---|---|---|---|
| Homo sapiens | 12 | 11 | 12 | 4 | 1 | 3 | 5 | 0 | 48 | [1] |
| Drosophila melanogaster | 10 | 8 | 14 | 2 | 1 | 3 | 15 | 3 | 56 | [1] |
| Anopheles gambiae | 9 | 5 | 13 | 2 | 1 | 3 | 16 | 3 | 52 | [47] |
| Daphnia pulex | 4 | 7 | 7 | 3 | 1 | 4 | 24 | 15 | 65 | [48] |
| Pediculus humanus humanus | 2 | 6 | 5 | 2 | 1 | 3 | 13 | 6 | 40 a | [49] |
| Apis melifera | 3 | 5 | 9 | 2 | 1 | 3 | 15 | 3 | 41 | [50] |
| Bombyx mori | 7 | 9 | 11 | 2 | 1 | 3 | 16 | 2 | 51 b | [50,51,52] |
| Tribolium castaneum | 10 | 6 | 35 | 2 | 1 | 3 | 13 | 3 | 73 | [20] |
| Tetranychus urticae | 9 | 4 | 39 | 2 | 1 | 3 | 23 | 22 | 103 | [53] |
| Chrysomela populi | 5 | 8 | 29 | 2 | 1 | 3 | 14 | 3 | 65 | [54] |
| Lygus hesperus | 11 | 6 | 12 | 2 | 1 | 3 | 19 | 11 | 65 | [55] |
| Lepeophtheirus salmonis | 3 | 4 | 11 | 3 | 1 | 4 | 2 | 5 | 33 | [56] |
| Laodelphax striatellux | 2 | 6 | 5 | 2 | 1 | 2 | 14 | 8 | 40 | [57] |
| Bemisia tabaci | 8 | 3 | 6 | 2 | 1 | 3 | 23 | 9 | 55 | [58] |
| Plutella xylostella | 15 | 14 | 21 | 3 | 1 | 3 | 19 | 6 | 82 | [59,60] |
| Manduca sexta | 7 | 9 | 11 | 2 | 1 | 3 | 13 | 3 | 52 | [60] |
| Danaus plexippus | 8 | 16 | 12 | 3 | 1 | 3 | 16 | 3 | 62 | [60] |
| Heliconius melpomene | 10 | 11 | 15 | 2 | 1 | 3 | 17 | 3 | 62 | [60] |
| Helicoverpa armigera | 7 | 11 | 11 | 2 | 1 | 3 | 17 | 2 | 54 | [52,61,62] |
| Helicoverpa zea | 7 | 11 | 11 | 2 | 1 | 3 | 17 | 2 | 54 | [52] |
| Acyrthosiphon pisum | 11 | 9 | 16 | 2 | 1 | 4 | 19 | 9 | 71 | [63,64] |
| Bt toxin | Receptor | Target pest | Reference |
|---|---|---|---|
| Cry1Aa | ABCC2 | Bombyx mori | [121] |
| Cry1Ab | ABCC2 | Bombyx mori | [122] |
| ABCG1 | Ostrinia furnacalis | [123] | |
| Cry1Ac | ABCC2 | Helicoverpa armigera | [25] |
| ABCC2 | Heliothis virescens | [124] | |
| ABCG1 | Ostrinia furnacalis | [123] | |
| ABCG1 | Plutella xylostella | [125] | |
| ABCC2 | Plutella xylostella | [126] | |
| ABCC2 | Spodoptera exigua | [127] | |
| ABCC3 | Spodoptera frugiperda | [128] | |
| Cry1Ca | ABCC2 | Spodoptera exigua | [127] |
| Cry1Fa | ABCC2 | Ostrinia nubilalis | [129] |
| Cry2Aa | ABCA2 | Helicoverpa armigera | [120] |
| Cry2Ab | ABCA2 | Helicoverpa armigera | [68] |
| ABCA2 | Helicoverpa punctigera | [68] | |
| ABCA2 | Pectinophora gossypiella | [118] | |
| Cry3Aa | ABCB1 | Chrysomela tremuela | [130] |
| Cry3Bb1 | ABCB1 | Diabrotica virgifera virgifera | [131] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, C.; Chakrabarty, S.; Jin, M.; Liu, K.; Xiao, Y. Insect ATP-Binding Cassette (ABC) Transporters: Roles in Xenobiotic Detoxification and Bt Insecticidal Activity. Int. J. Mol. Sci. 2019, 20, 2829. https://doi.org/10.3390/ijms20112829
Wu C, Chakrabarty S, Jin M, Liu K, Xiao Y. Insect ATP-Binding Cassette (ABC) Transporters: Roles in Xenobiotic Detoxification and Bt Insecticidal Activity. International Journal of Molecular Sciences. 2019; 20(11):2829. https://doi.org/10.3390/ijms20112829
Chicago/Turabian StyleWu, Chao, Swapan Chakrabarty, Minghui Jin, Kaiyu Liu, and Yutao Xiao. 2019. "Insect ATP-Binding Cassette (ABC) Transporters: Roles in Xenobiotic Detoxification and Bt Insecticidal Activity" International Journal of Molecular Sciences 20, no. 11: 2829. https://doi.org/10.3390/ijms20112829
APA StyleWu, C., Chakrabarty, S., Jin, M., Liu, K., & Xiao, Y. (2019). Insect ATP-Binding Cassette (ABC) Transporters: Roles in Xenobiotic Detoxification and Bt Insecticidal Activity. International Journal of Molecular Sciences, 20(11), 2829. https://doi.org/10.3390/ijms20112829