Corticosterone Upregulates Gene and Protein Expression of Catecholamine Markers in Organotypic Brainstem Cultures

 , ,
, , 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
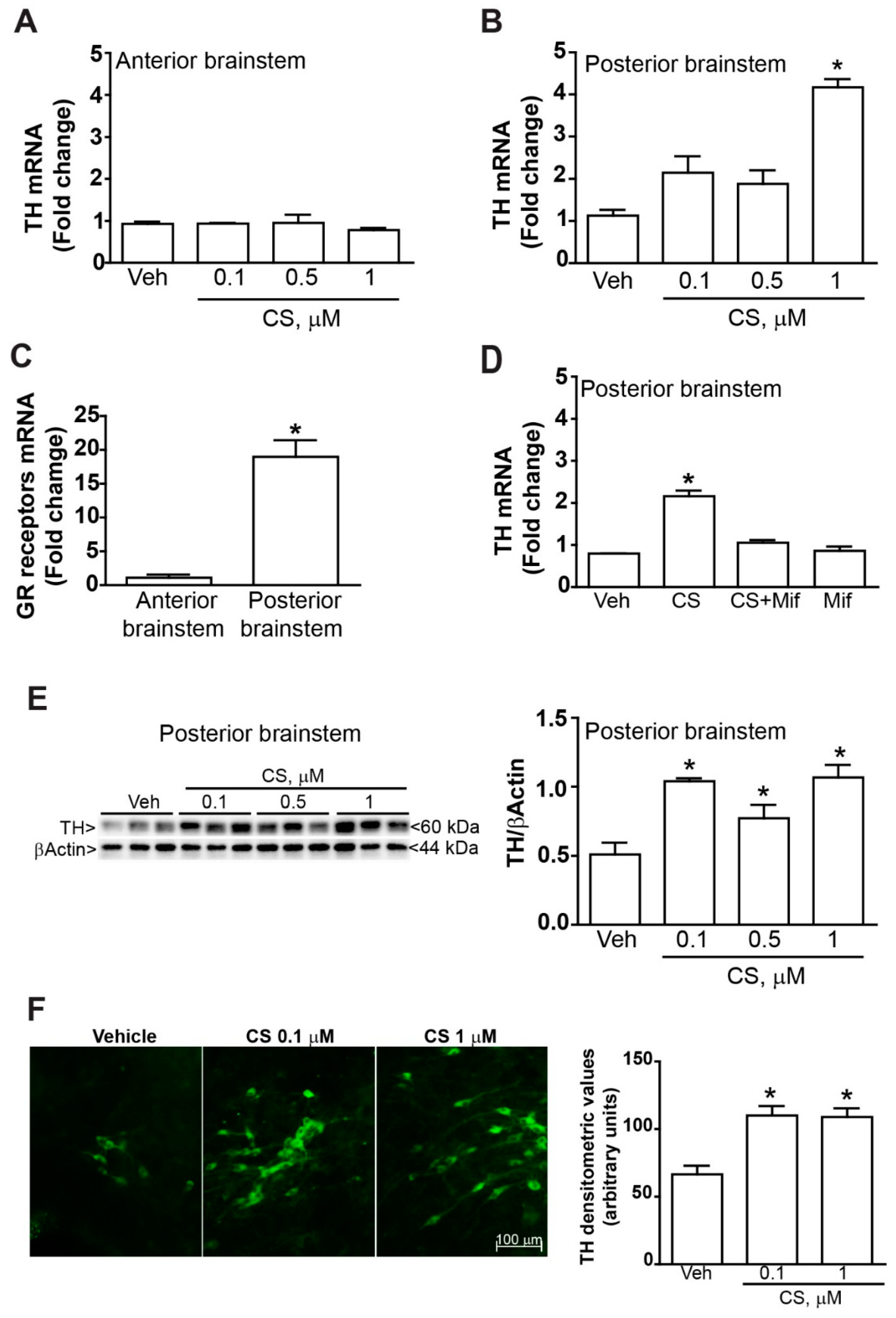
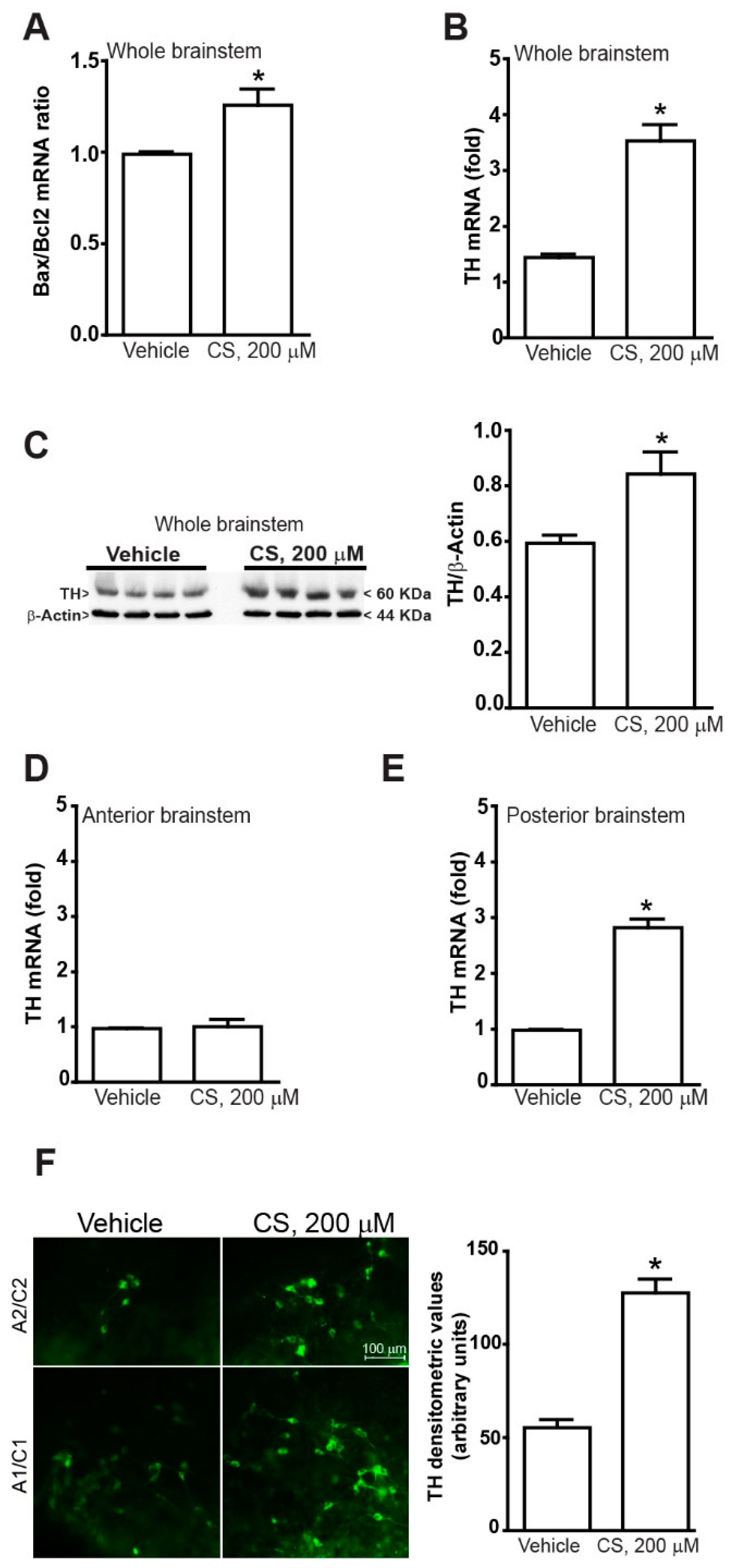
2.1. TH Increases within Organotypic Cultures Following Incubation with Corticosterone
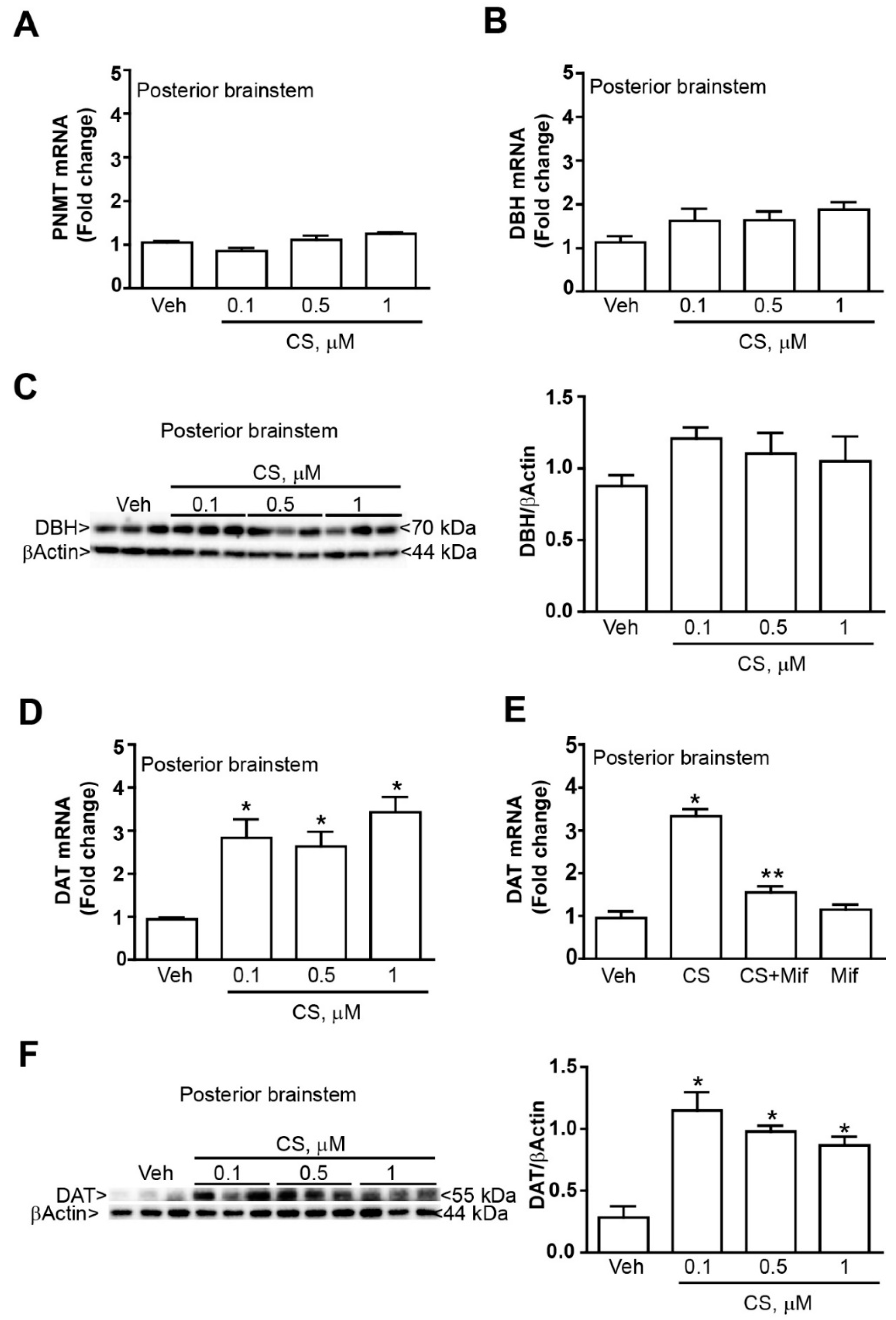
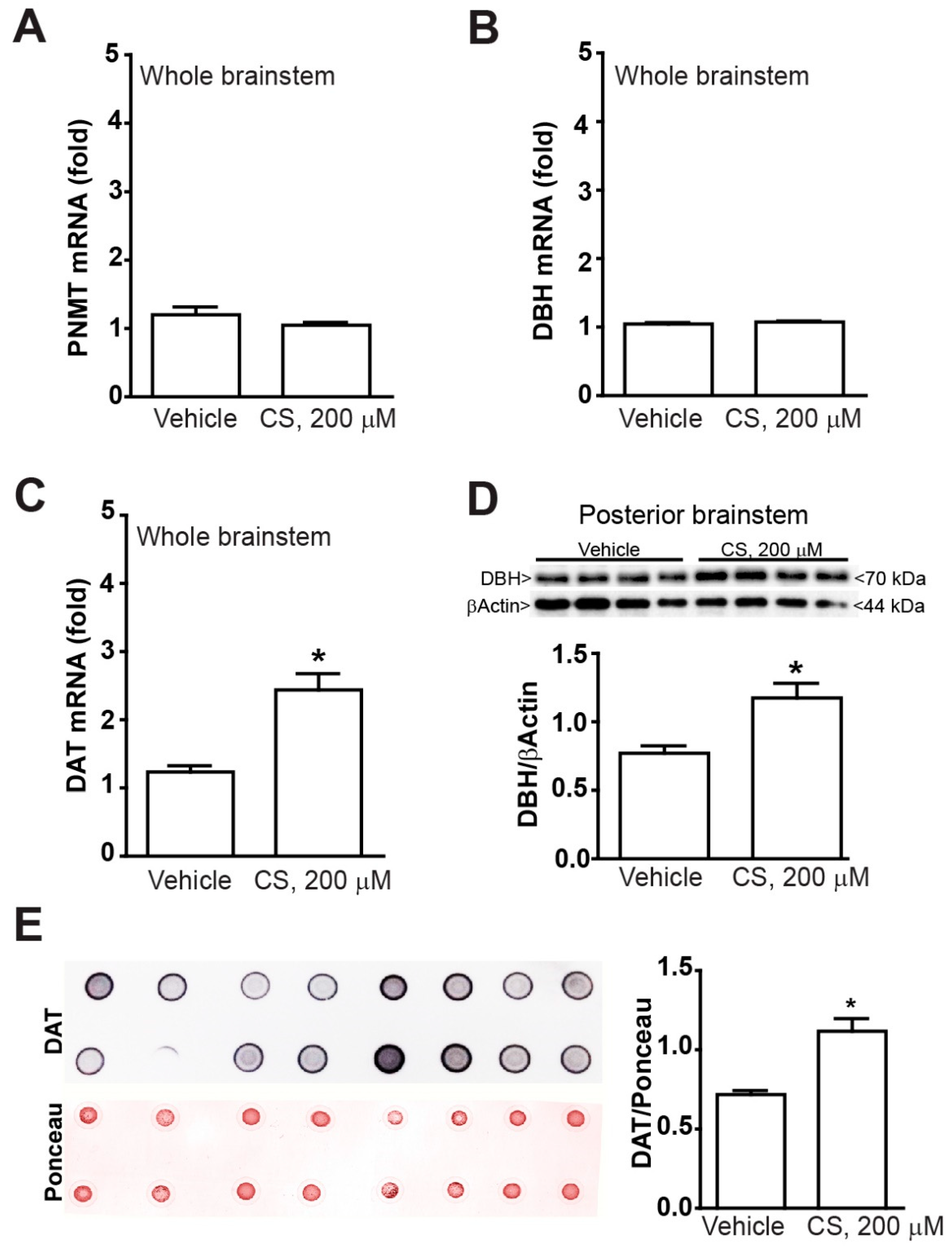
2.2. Low Doses of Corticosterone Increase DAT mRNA and Protein without Altering PNMT and DBH
3. Discussion
4. Materials and Methods
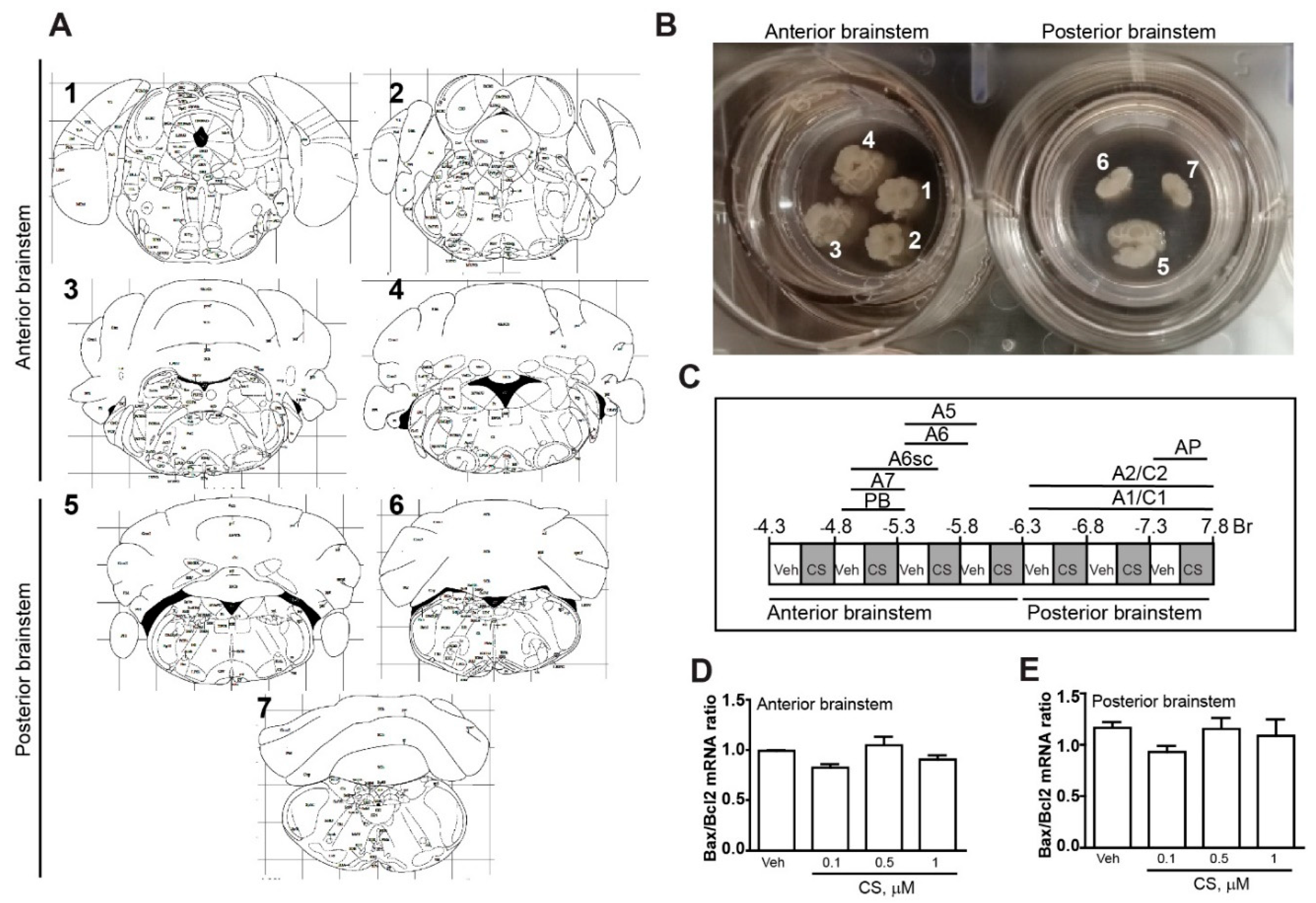
4.1. Organotypic Brainstem Cultures
4.2. Corticosterone ELISA
4.3. Experimental Planning
4.4. Western Blot
4.5. Dot Blot
4.6. Real Time PCR Analysis
4.7. Immunohistochemistry
4.8. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 6-OHDA | 6-hydroxydopamine |
| A | Adrenaline |
| CNS | Central nervous system |
| CS | Corticosterone |
| DA | Dopamine |
| DAT | Dopamine transporter |
| DBH | Beta-Hydroxylase |
| GR | Glucocorticoid receptors |
| LC | Locus coeruleus |
| NA | Noradrenaline |
| PNMT | Phenylethanolamine N-methyltransferase |
| TH | Tyrosine hydroxylase |
| VTA | Ventral tegmental area |
References
- Cain, D.W.; Cidlowski, J.A. Immune regulation by glucocorticoids. Nat. Rev. Immunol. 2017, 17, 233–247. [Google Scholar] [CrossRef] [PubMed]
- Straub, R.H.; Cutolo, M. Glucocorticoids and chronic inflammation. Rheumatology 2016, 55, ii6–ii14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dickmeis, T.; Foulkes, N.S. Glucocorticoids and circadian clock control of cell proliferation: At the interface between three dynamic systems. Mol. Cell. Endocrinol. 2011, 331, 11–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Lim, G.; Zeng, Q.; Sung, B.; Ai, Y.; Guo, G.; Yang, L.; Mao, J. Expression of central glucocorticoid receptors after peripheral nerve injury contributes to neuropathic pain behaviors in rats. J. Neurosci. 2004, 24, 8595–8605. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Lim, G.; Zeng, Q.; Sung, B.; Yang, L.; Mao, J. Central glucocorticoid receptors modulate the expression and function of spinal NMDA receptors after peripheral nerve injury. J. Neurosci. 2005, 25, 488–495. [Google Scholar] [CrossRef] [PubMed]
- Takasaki, I.; Kurihara, T.; Saegusa, H.; Zong, S.; Tanabe, T. Effects of glucocorticoid receptor antagonists on allodynia and hyperalgesia in mouse model of neuropathic pain. Eur. J. Pharmacol. 2005, 524, 80–83. [Google Scholar] [CrossRef] [PubMed]
- Alexander, J.K.; DeVries, A.C.; Kigerl, K.A.; Dahlman, J.M.; Popovich, P.G. Stress exacerbates neuropathic pain via glucocorticoid and NMDA receptor activation. Brain Behav. Immun. 2009, 23, 851–860. [Google Scholar] [CrossRef] [Green Version]
- De Guia, R.M.; Rose, A.J.; Herzig, S. Glucocorticoid hormones and energy homeostasis. Horm. Mol. Biol. Clin. Investig. 2014, 19, 117–128. [Google Scholar] [CrossRef]
- Rafacho, A.; Ortsäter, H.; Nadal, A.; Quesada, I. Glucocorticoid treatment and endocrine pancreas function: Implications for glucose homeostasis, insulin resistance and diabetes. J. Endocrinol. 2014, 223, R49–R62. [Google Scholar] [CrossRef]
- Wong, D.L.; Hayashi, R.J.; Ciaranello, R.D. Regulation of biogenic amine methyltransferases by glucocorticoids via S-adenosylmethionine and its metabolizing enzymes, methionine adenosyltransferase and S-adenosylhomocysteine hydrolase. Brain Res. 1985, 330, 209–216. [Google Scholar] [CrossRef]
- Wong, D.L.; Siddall, B.; Wang, W. Hormonal control of rat adrenal phenylethanolamine N-methyltransferase. Enzyme activity, the final critical pathway. Neuropsychopharmacology 1995, 13, 223–234. [Google Scholar] [CrossRef]
- Tai, T.C.; Wong, D.L. Phenylethanolamine N-methyltransferase gene regulation by cAMP-dependent protein kinase A and protein kinase C signaling pathways. Ann. N. Y. Acad. Sci. 2002, 971, 83–85. [Google Scholar] [CrossRef] [PubMed]
- Tai, T.C.; Claycomb, R.; Siddall, B.J.; Bell, R.A.; Kvetnansky, R.; Wong, D.L. Stress-induced changes in epinephrine expression in the adrenal medulla in vivo. J. Neurochem. 2007, 101, 1108–1118. [Google Scholar] [CrossRef] [PubMed]
- Sharara-Chami, R.I.; Joachim, M.; Pacak, K.; Majzoub, J.A. Glucocorticoid treatment-effect on adrenal medullary catecholamine production. Shock 2010, 33, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Hasan, K.M.; Rahman, M.S.; Arif, K.M.; Sobhani, M.E. Psychological stress and aging: Role of glucocorticoids (GCs). Age 2012, 34, 1421–1433. [Google Scholar] [CrossRef] [PubMed]
- Vyas, S.; Rodrigues, A.J.; Silva, J.M.; Tronche, F.; Almeida, O.F.; Sousa, N.; Sotiropoulos, I. Chronic Stress and Glucocorticoids: From Neuronal Plasticity to Neurodegeneration. Neural Plast. 2016, 2016. [Google Scholar] [CrossRef] [PubMed]
- De Quervain, D.; Schwabe, L.; Roozendaal, B. Stress, glucocorticoids and memory: Implications for treating fear-related disorders. Nat. Rev. Neurosci. 2017, 18, 7–19. [Google Scholar] [CrossRef] [PubMed]
- Stratakis, C.A. Cushing syndrome caused by adrenocortical tumors and hyperplasias (corticotropin- independent Cushing syndrome). Endocr. Dev. 2008, 13, 117–132. [Google Scholar] [CrossRef] [PubMed]
- Lacroix, A. ACTH-independent macronodular adrenal hyperplasia. Best Pract. Res. Clin. Endocrinol. Metab. 2009, 23, 245–259. [Google Scholar] [CrossRef]
- Arnaldi, G.; Angeli, A.; Atkinson, A.B.; Bertagna, X.; Cavagnini, F.; Chrousos, G.P.; Fava, G.A.; Findling, J.W.; Gaillard, R.C.; Grossman, A.B.; et al. Diagnosis and complications of Cushing’s syndrome: A consensus statement. J. Clin. Endocrinol. Metab. 2003, 88, 5593–5602. [Google Scholar] [CrossRef]
- Newell-Price, J.; Bertagna, X.; Grossman, A.B.; Nieman, L.K. Cushing’s syndrome. Lancet 2006, 367, 1605–1617. [Google Scholar] [CrossRef]
- Biller, B.M.; Grossman, A.B.; Stewart, P.M.; Melmed, S.; Bertagna, X.; Bertherat, J.; Buchfelder, M.; Colao, A.; Hermus, A.R.; Hofland, L.J.; et al. Treatment of adrenocorticotropin-dependent Cushing’s syndrome: A consensus statement. J. Clin. Endocrinol. Metab. 2008, 93, 2454–2462. [Google Scholar] [CrossRef] [PubMed]
- Nieman, L.K. Cushing’s syndrome: Update on signs, symptoms and biochemical screening. Eur. J. Endocrinol. 2015, 173, M33–M38. [Google Scholar] [CrossRef]
- Pivonello, R.; De Leo, M.; Vitale, P.; Cozzolino, A.; Simeoli, C.; De Martino, M.C.; Lombardi, G.; Colao, A. Pathophysiology of diabetes mellitus in Cushing’s syndrome. Neuroendocrinology 2010, 92, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Isidori, A.M.; Graziadio, C.; Paragliola, R.M.; Cozzolino, A.; Ambrogio, A.G.; Colao, A.; Corsello, S.M.; Pivonello, R. The hypertension of Cushing’s syndrome: Controversies in the pathophysiology and focus on cardiovascular complications. J. Hypertens. 2015, 33, 44–60. [Google Scholar] [CrossRef] [PubMed]
- Ferraù, F.; Korbonits, M. Metabolic comorbidities in Cushing’s syndrome. Eur. J. Endocrinol. 2015, 173, M133–M157. [Google Scholar] [CrossRef] [PubMed]
- Stratakis, C.A. Skin manifestations of Cushing’s syndrome. Rev. Endocr. Metab. Disord. 2016, 17, 283–286. [Google Scholar] [CrossRef]
- Cohen, S.; Nathan, J.A.; Goldberg, A.L. Muscle wasting in disease: Molecular mechanisms and promising therapies. Nat. Rev. Drug Discov. 2015, 14, 58–74. [Google Scholar] [CrossRef]
- Bratek, A.; Koźmin-Burzyńska, A.; Górniak, E.; Krysta, K. Psychiatric disorders associated with Cushing’s syndrome. Psychiatr. Danub. 2015, 27, S339–S343. [Google Scholar]
- Shipley, J.E.; Schteingart, D.E.; Tandon, R.; Starkman, M.N. Sleep architecture and sleep apnea in patients with Cushing’s disease. Sleep 1992, 15, 514–518. [Google Scholar] [CrossRef]
- Chandran, D.S.; Ali, N.; Jaryal, A.K.; Jyotsna, V.P.; Deepak, K.K. Decreased autonomic modulation of heart rate and altered cardiac sympathovagal balance in patients with Cushing’s syndrome: Role of endogenous hypercortisolism. Neuroendocrinology 2013, 97, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Pivonello, R.; Simeoli, C.; De Martino, M.C.; Cozzolino, A.; De Leo, M.; Iacuaniello, D.; Pivonello, C.; Negri, M.; Pellecchia, M.T.; Iasevoli, F.; et al. Neuropsychiatric disorders in Cushing’s syndrome. Front. Neurosci. 2015, 9, 129. [Google Scholar] [CrossRef] [PubMed]
- Brüne, M.; Schröder, S.G. Neuroleptic-induced akathisia and early onset tardive dyskinesia in affective disorder due to Cushing’s syndrome. Gen. Hosp. Psychiatry 1997, 19, 445–447. [Google Scholar] [CrossRef]
- Stenzel-Poore, M.P.; Cameron, V.A.; Vaughan, J.; Sawchenko, P.E.; Vale, W. Development of Cushing’s syndrome in corticotropin-releasing factor transgenic mice. Endocrinology 1992, 130, 3378–3386. [Google Scholar] [CrossRef] [PubMed]
- Westphal, C.H.; Muller, L.; Zhou, A.; Zhu, X.; Bonner-Weir, S.; Schambelan, M.; Steiner, D.F.; Lindberg, I.; Leder, P. The neuroendocrine protein 7B2 is required for peptide hormone processing in vivo and provides a novel mechanism for pituitary Cushing’s disease. Cell 1999, 96, 689–700. [Google Scholar] [CrossRef]
- Sun, Z.; Fan, Y.; Zha, Q.; Zhu, M.Y. Corticosterone Up-regulates Expression and Function of Norepinephrine Transporter in SK-N-BE(2)C Cells. J. Neurochem. 2010, 113, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Berry, J.N.; Meredith, A.; Saunders, M.A.; Sharrett-Field, L.J.; Reynolds, A.R.; Bardo, M.T.; James, R.; Pauly, J.R.; Prendergast, M.A. Corticosterone enhances N-methyl-d-aspartate receptor signaling to promote isolated ventral tegmental area activity in a reconstituted mesolimbic dopamine pathway. Brain Res. Bull. 2016, 120, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Zhou, H.; Li, X. Curcumin Protects PC12 Cells from Corticosterone-Induced Cytotoxicity: Possible Involvement of the ERK1/2 Pathway. Basic Clin. Pharmacol. Toxicol. 2009, 104, 236–240. [Google Scholar] [CrossRef] [PubMed]
- Mao, Q.Q.; Zhong, X.M.; Qiu, F.M.; Li, Z.Y.; Huang, Z. Protective effects of paeoniflorin against corticosterone-induced neurotoxicity in PC12 cells. Phytother. Res. 2012, 26, 969–973. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Li, H.; Zhao, L.; Li, X.; You, J.; Jiang, Q.; Li, S.; Jin, L.; Xu, Y. Protective effects of aqueous extract from Acanthopanax senticosus against corticosterone-induced neurotoxicity in PC12 cells. J. Ethnopharmacol. 2013, 148, 861–868. [Google Scholar] [CrossRef] [PubMed]
- Polman, J.A.; Welten, J.E.; Bosch, D.S.; de Jonge, R.T.; Balog, J.; van der Maarel, S.M.; de Kloet, E.R.; Datson, N.A. A genome-wide signature of glucocorticoid receptor binding in neuronal PC12 cells. BMC Neurosci. 2012, 13, 118. [Google Scholar] [CrossRef] [PubMed]
- Sheela Rani, C.S.; Soto-Pina, A.; Iacovitti, L.; Strong, R. Evolutionary conservation of an atypical glucocorticoid-responsive element in the human tyrosine hydroxylase gene. J. Neurochem. 2013, 126, 19–28. [Google Scholar] [CrossRef]
- Kalinina, T.S.; Shishkina, G.T.; Dygalo, N.N. Induction of tyrosine hydroxylase gene expression by glucocorticoids in the perinatal rat brain is age-dependent. Neurochem. Res. 2012, 37, 811–818. [Google Scholar] [CrossRef] [PubMed]
- Hensleigh, E.; Pritchard, L.M. Glucocorticoid receptor expression and sub-cellular localization in dopamine neurons of the rat midbrain. Neurosci. Lett. 2013, 556, 191–195. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, S.; Uchida, K.; Nakayama, H. Striatal TH-immunopositive fibers recover after an intrastriatal injection of 6-hydroxydopamine in golden hamsters treated with prednisolone: Roles of tumor necrosis factor-α and inducible nitric oxidesynthase in neurodegeneration. Neurosci. Res. 2013, 76, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Copeland, B.J.; Neff, N.H.; Hadjiconstantinou, M. Enhanced dopamine uptake in the striatum following repeated restraint stress. Synapse 2005, 57, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Czyrak, A.; Chocyk, A. Search for the presence of glucocorticoid receptors in dopaminergic neurons of rat ventral tegmental area and substantia nigra. Pol. J. Pharmacol. 2001, 53, 681–684. [Google Scholar]
- Harfstrand, A.; Fuxe, K.; Cintra, A.; Agnati, L.F.; Zini, I.; Wikstrom, A.C.; Okret, S.; Yu, Z.Y.; Goldstein, M.; Steinbusch, H.; et al. Glucocorticoid receptor immunoreactivity in monoaminergic neurons of rat brain. Proc. Natl. Acad. Sci. USA 1986, 83, 9779–9783. [Google Scholar] [CrossRef]
- Heydendael, W.; Jacobson, L. Glucocorticoid status affects antidepressant regulation of locus coeruleus tyrosine hydroxylase and dorsal raphe tryptophan hydroxylase gene expression. Brain Res. 2009, 1288, 69–78. [Google Scholar] [CrossRef]
- Li, M.; Han, F.; Shi, Y. Expression of locus coeruleus mineralocorticoid receptor and glucocorticoid receptor in rats under single-prolonged stress. Neurol. Sci. 2011, 32, 625–631. [Google Scholar] [CrossRef]
- Nakagaki, T.; Hirooka, Y.; Matsukawa, R.; Nishihara, M.; Nakano, M.; Ito, K.; Hoka, S.; Sunagawa, K. Activation of mineralocorticoid receptors in the rostral ventrolateral medulla is involved in hypertensive mechanisms in stroke-prone spontaneously hypertensive rats. Hypertens. Res. 2012, 35, 470–476. [Google Scholar] [CrossRef] [PubMed]
- Makino, S.; Smith, M.A.; Gold, P.W. Regulatory role of glucocorticoids and glucocorticoid receptor mRNA levels on tyrosine hydroxylase gene expression in the locus coeruleus during repeated immobilization stress. Brain Res. 2002, 943, 216–223. [Google Scholar] [CrossRef]
- Zhang, R.; Jankord, R.; Flak, J.N.; Solomon, M.B.; D’Alessio, D.A.; Herman, J.P. Role of glucocorticoids in tuning hindbrain stress integration. J. Neurosci. 2010, 30, 14907–14914. [Google Scholar] [CrossRef] [PubMed]
- Núñez, C.; Földes, A.; Pérez-Flores, D.; García-Borrón, J.C.; Laorden, M.L.; Kovács, K.J.; Milanés, M.V. Elevated glucocorticoid levels are responsible for induction of tyrosine hydroxylase mRNA expression, phosphorylation, and enzyme activity in the nucleus of the solitary tract during morphine withdrawal. Endocrinology 2009, 150, 3118–3127. [Google Scholar] [CrossRef] [PubMed]
- Virdee, K.; McArthur, S.; Brischoux, F.; Caprioli, D.; Ungless, M.A.; Robbins, T.W.; Dalley, J.W.; Gillies, G.E. Antenatal glucocorticoid treatment induces adaptations in adult midbrain dopamine neurons, which underpin sexually dimorphic behavioral resilience. Neuropsychopharmacology 2014, 39, 339–350. [Google Scholar] [CrossRef] [PubMed]
- McArthur, S.; McHale, E.; Dalley, J.W.; Buckingham, J.C.; Gillies, G.E. Altered mesencephalic dopaminergic populations in adulthood as a consequence of brief perinatal glucocorticoid exposure. J. Neuroendocrinol. 2005, 17, 475–482. [Google Scholar] [CrossRef]
- McArthur, S.; McHale, E.; Gillies, G.E. The size and distribution of midbrain dopaminergic populations are permanently altered by perinatal glucocorticoid exposure in a sex- region- and time-specific manner. Neuropsychopharmacology 2007, 32, 1462–1476. [Google Scholar] [CrossRef]
- Niwa, M.; Jaaro-Peled, H.; Tankou, S.; Seshadri, S.; Hikida, T.; Matsumoto, Y.; Cascella, N.G.; Kano, S.; Ozaki, N.; Nabeshima, T.; et al. Adolescent stress-induced epigenetic control of dopaminergic neurons via glucocorticoids. Science 2013, 339, 335–339. [Google Scholar] [CrossRef]
- Schefe, J.H.; Lehmann, K.E.; Buschmann, I.R.; Unger, T.; Funke-Kaiser, H. Quantitative real-time RT- PCR data analysis: Current concepts and the novel “gene expression’s CT difference” formula. J. Mol. Med. 2006, 84, 901–910. [Google Scholar] [CrossRef]
- Yuan, J.S.; Reed, A.; Chen, F.; Stewart, C.N., Jr. Statistical analysis of real-time PCR data. BMC Bioinform. 2006, 7, 85. [Google Scholar] [CrossRef]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Busceti, C.L.; Ferese, R.; Bucci, D.; Ryskalin, L.; Gambardella, S.; Madonna, M.; Nicoletti, F.; Fornai, F. Corticosterone Upregulates Gene and Protein Expression of Catecholamine Markers in Organotypic Brainstem Cultures. Int. J. Mol. Sci. 2019, 20, 2901. https://doi.org/10.3390/ijms20122901
Busceti CL, Ferese R, Bucci D, Ryskalin L, Gambardella S, Madonna M, Nicoletti F, Fornai F. Corticosterone Upregulates Gene and Protein Expression of Catecholamine Markers in Organotypic Brainstem Cultures. International Journal of Molecular Sciences. 2019; 20(12):2901. https://doi.org/10.3390/ijms20122901
Chicago/Turabian StyleBusceti, Carla L., Rosangela Ferese, Domenico Bucci, Larisa Ryskalin, Stefano Gambardella, Michele Madonna, Ferdinando Nicoletti, and Francesco Fornai. 2019. "Corticosterone Upregulates Gene and Protein Expression of Catecholamine Markers in Organotypic Brainstem Cultures" International Journal of Molecular Sciences 20, no. 12: 2901. https://doi.org/10.3390/ijms20122901
APA StyleBusceti, C. L., Ferese, R., Bucci, D., Ryskalin, L., Gambardella, S., Madonna, M., Nicoletti, F., & Fornai, F. (2019). Corticosterone Upregulates Gene and Protein Expression of Catecholamine Markers in Organotypic Brainstem Cultures. International Journal of Molecular Sciences, 20(12), 2901. https://doi.org/10.3390/ijms20122901