Phenolic Compounds and Ginsenosides in Ginseng Shoots and Their Antioxidant and Anti-Inflammatory Capacities in LPS-Induced RAW264.7 Mouse Macrophages


Abstract
:
1. Introduction
2. Results and Discussion
2.1. Total Phenols, Total Flavonoids, and Antioxidant Activities in GSEs
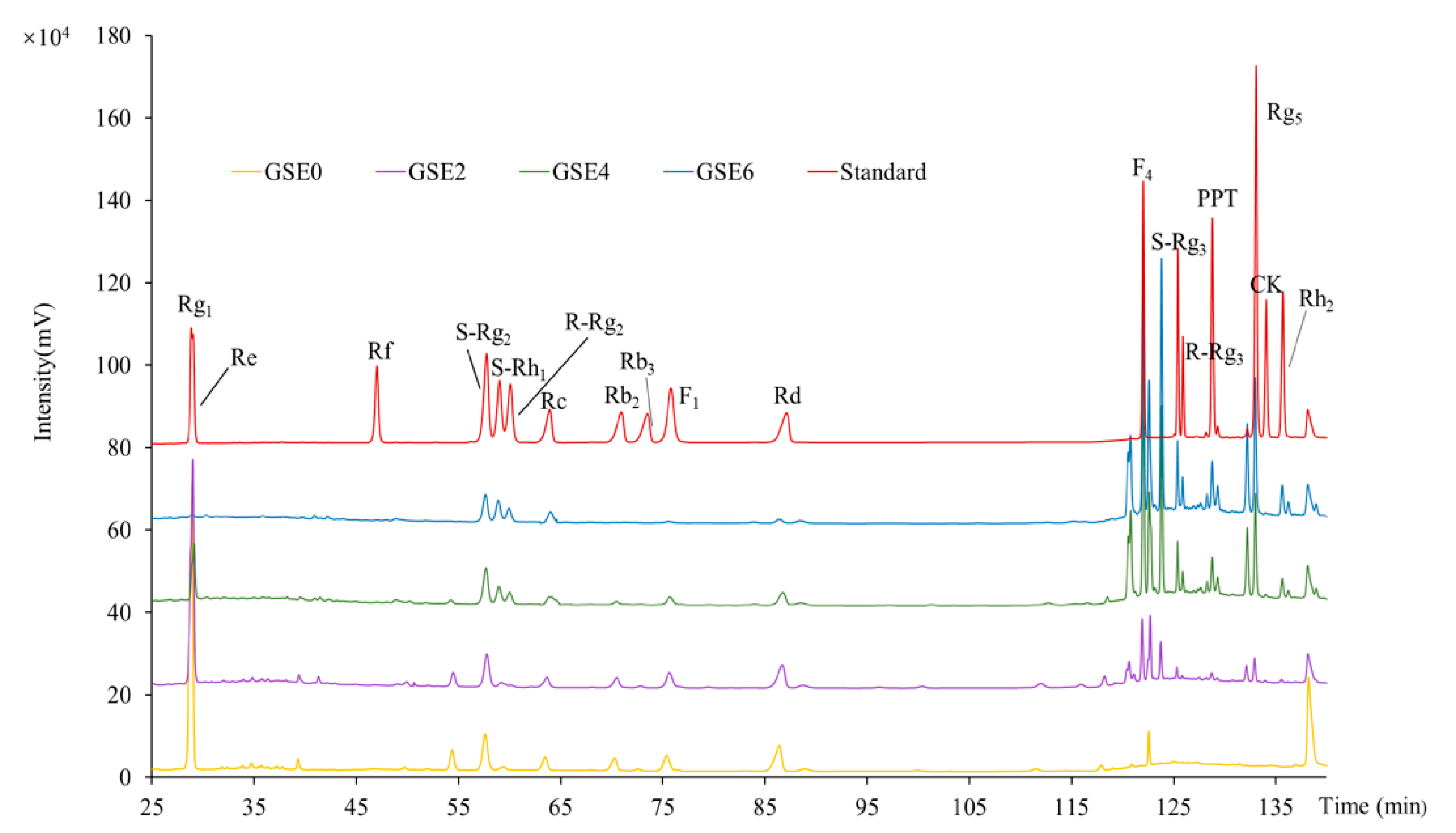
2.2. HPLC Profiles and Contents of Individual Ginsenosides in GSEs
2.3. Effects of GSEs on LPS-Induced RAW264.7 Cell Viability and Production of Three Inflammation Factors
2.4. Effects of GSEs on Relative mRNA Expression of Four Inflammation Factors in LPS-Induced RAW264.7 Cells
2.5. Correlation Analysis
3. Materials and Methods
3.1. Chemicals and Reagents, Preparation of Plant Extracts, and Culture of RAW264.7 Cells
3.2. Evaluation of Phenolic Content and Antioxidant Capacity and HPLC Analyses of Ginsenosides
3.3. Assays of Cell Viability, and Release of NO and the Cytokines IL-6 and TNF-α
3.4. qRT-PCR Assays of Inducible Nitric Oxide Synthase (iNOS), TNF-α, IL-6 and Cyclooxygenase-2 (COX-2) mRNAs
3.5. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chhetri, J.K.; de Souto Barreto, P.; Fougère, B.; Rolland, Y.; Vellas, B.; Cesari, M. Chronic inflammation and sarcopenia: A regenerative cell therapy perspective. Exp. Gerontol. 2018, 103, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Nandakumar, K.S. Recent advances in the development of vaccines for chronic inflammatory autoimmune diseases. Vaccine 2018, 36, 3208–3220. [Google Scholar] [CrossRef] [PubMed]
- Bartsch, H.; Nair, J. Chronic inflammation and oxidative stress in the genesis and perpetuation of cancer: Role of lipid peroxidation, DNA damage, and repair. Langenbecks Arch. Surg. 2006, 391, 499–510. [Google Scholar] [CrossRef] [PubMed]
- Wei, B.; Duan, Z.G.; Zhu, C.H.; Deng, J.J.; Fan, D.D. Anti-anemia effects of ginsenoside Rk3 and ginsenoside Rh4 on mice with ribavirin-induced anemia. Food Funct. 2018, 9, 2447–2455. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Min, D.S.; Lee, C.W.; Song, K.H.; Kim, Y.S.; Kim, H.P. Ginsenosides from Korean Red Ginseng ameliorate lung inflammatory responses: Inhibition of the MAPKs/NF-κB/c-Fos pathways. J. Ginseng Res. 2018, 42, 476–484. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Ren, D.Y.; Li, J.R.; Yuan, G.X.; Li, H.Y.; Xu, G.Y.; Han, X.; Du, P.G.; An, L.P. Effects of compound K on hyperglycemia and insulin resistance in rats with type 2 diabetes mellitus. Fitoterapia 2014, 95, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.K.; Kang, H.; Baek, C.W.; Jung, Y.H.; Woo, Y.C.; Choi, G.J.; Shin, H.Y.; Kim, K.S. Antinociceptive and anti-inflammatory effects of ginsenoside Rf in a rat model of incisional pain. J. Ginseng Res. 2018, 42, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.G.; Yoo, S.R.; Park, H.J.; Lee, N.H.; Shin, J.W.; Sathyanath, R.; Cho, J.H.; Son, C.G. Antioxidant effects of Panax ginseng C.A. Meyer in healthy subjects: A randomized, placebo-controlled clinical trial. Food Chem. Toxicol. 2011, 49, 2229–2235. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H. Pharmacological and medical applications of Panax ginseng and ginsenosides: A review for use in cardiovascular diseases. J. Ginseng Res. 2018, 42, 264–269. [Google Scholar] [CrossRef]
- Yi, Y.S. Ameliorative effects of ginseng and ginsenosides on rheumatic diseases. J. Ginseng Res. 2018, 1–7. [Google Scholar] [CrossRef]
- Choi, P.; Park, J.Y.; Kim, T.; Park, S.H.; Kim, H.; Kang, K.S.; Ham, J. Improved anticancer effect of ginseng extract by microwave-assisted processing through the generation of ginsenosides Rg3, Rg5 and Rk1. J. Funct. Foods 2015, 14, 613–622. [Google Scholar] [CrossRef]
- Li, B.H.; Zhao, J.; Wang, C.Z.; Searle, J.; He, T.C.; Yuan, C.S.; Du, W. Ginsenoside Rh2 induces apoptosis and paraptosis-like cell death in colorectal cancer cells through activation of p53. Cancer Lett. 2011, 301, 185–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Im, K.; Kim, J.; Min, H. Ginseng, the natural effectual antiviral: Protective effects of Korean Red Ginseng against viral infection. J. Ginseng Res. 2016, 40, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Kim, J.M.; Kim, D.H.; Park, S.J.; Liu, X.T.; Cai, M.D.; Hong, J.G.; Park, J.H.; Ryu, J.H. Effects of Sun ginseng on memory enhancement and hippocampal neurogenesis. Phytother. Res. 2013, 27, 1293–1299. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.M.; Yoon, H.; Park, H.M.; Song, B.C.; Yeum, K.J. Implications of red Panax ginseng in oxidative stress associated chronic diseases. J. Ginseng Res. 2017, 41, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Tsukioka, Y.; Nakao, K.; Moriyama, K.; Shintani, T.; Samukawa, K.; Itoh, K.; Takeshina, F.; Matsuda, H. Pharmacological study on Panax ginseng C. A. Meyer. XYI. Improving effects of Red Ginseng on peripheral circulation disorder. J. Trad. Med. 2008, 25, 43–51. [Google Scholar] [CrossRef]
- Ma, L.Y.; Yang, X.W. Six new dammarane-type triterpenes from acidic hydrolysate of the stems-leaves of Panax ginseng and their inhibitory–activities against three human cancer cell lines. Phytochem. Lett. 2015, 13, 406–412. [Google Scholar] [CrossRef]
- Li, X.; Yao, F.; Fan, H.; Li, K.; Sun, L.W.; Liu, Y.J. Intraconversion of Polar Ginsenosides, Their Transformation into Less-Polar Ginsenosides, and Ginsenoside Acetylation in Ginseng Flowers upon Baking and Steaming. Molecules 2018, 23, 759. [Google Scholar] [CrossRef]
- Kim, M.S.; Kim, S.H.; Park, S.J.; Sung, M.J.; Park, J.; Hwang, J.T.; Yang, H.J.; Kim, S.; Seo, D.; Shin, S.S.; et al. Ginseng berry improves hyperglycemia by downregulating hepatic gluconeogenesis and steatosis in mice with diet-induced type 2 diabetes. J. Funct. Foods 2017, 35, 295–302. [Google Scholar] [CrossRef]
- Lee, J.W.; Choi, B.R.; Kim, Y.C.; Choi, D.J.; Lee, Y.S.; Kim, G.S.; Baek, N.I.; Kim, S.Y.; Lee, D.Y. Comprehensive Profiling and Quantification of Ginsenosides in the Root, Stem, Leaf, and Berry of Panax ginseng by UPLC-QTOF/MS. Molecules 2017, 22, 2147. [Google Scholar] [CrossRef]
- Hwang, C.R.; Lee, S.H.; Jang, G.Y.; Hwang, I.G.; Kim, H.Y.; Woo, K.S.; Lee, J.; Jeong, H.S. Changes in ginsenoside compositions and antioxidant activities of hydroponic-cultured ginseng roots and leaves with heating temperature. J. Ginseng Res. 2014, 38, 180–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Saeedi, A.H.; Hossain, M.A. Total phenols, total flavonoids contents and free radical scavenging activity of seeds crude extracts of pigeon pea traditionally used in Oman for the treatment of several chronic diseases. Asian Pac. J. Trop. Dis. 2015, 5, 316–321. [Google Scholar] [CrossRef]
- Xu, G.H.; Ye, X.Q.; Chen, J.C.; Liu, D.H. Effect of heat treatment on the phenolic compounds and antioxidant capacity of citrus peel extract. J. Agric. Food Chem. 2007, 55, 330–335. [Google Scholar] [CrossRef] [PubMed]
- Budaraju, S.; Mallikarjunan, K.; Annor, G.; Schoenfuss, T.; Raun, R. Effect of pre-treatments on the antioxidant potential of phenolic extracts from barley malt rootlets. Food Chem. 2018, 266, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.S.; Yamabe, N.; Kim, H.Y.; Okamoto, T.; Sei, Y.; Yokozawa, T. Increase in the free radical scavenging activities of American ginseng by heat processing and its safety evaluation. J. Ethnopharmacol. 2007, 113, 225–232. [Google Scholar] [CrossRef]
- Hwang, I.G.; Kim, H.Y.; Joung, E.M.; Woo, K.S.; Jeong, J.H.; Yu, K.W.; Lee, J.; Jeong, H.S. Changes in ginsenosides and antioxidant activity of Korean ginseng (Panax ginseng C.A. Meyer) with Heating Temperature and Pressure. Food Sci. Biotechnol. 2010, 19, 941–949. [Google Scholar] [CrossRef]
- Sun, S.; Qi, L.W.; Du, G.J.; Mehendale, S.R.; Wang, C.Z.; Yuan, C.S. Red notoginseng: Higher ginsenoside content and stronger anticancer potential than Asian and American ginseng. Food Chem. 2011, 125, 1299–1305. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.F.; Gao, Y.; Xu, S.Y.; Liu, H.; Xue, X.; Zhang, Y.; Zhang, H.; Liu, M.N.; Xiong, H.; Lin, R.C. Remarkable impact of steam temperature on ginsenosides transformation from fresh ginseng to red ginseng. J. Ginseng Res. 2018, 42, 277–287. [Google Scholar] [CrossRef]
- Lee, H.J.; Jeong, H.S.; Kim, D.J.; Noh, Y.H.; Yuk, D.Y.; Hong, J.T. Inhibitory effect of citral on NO production by suppression of iNOS expression and NF-κB activation in RAW264.7 cells. Arch. Pharm. Res. 2008, 31, 342–349. [Google Scholar] [CrossRef]
- Guzik, T.J.; Korbut, R.; Adamek-Guzik, T. Nitric oxide and superoxide in inflammation and immune regulation. J. Physiol. Pharmacol. 2003, 54, 469–487. [Google Scholar]
- Conforti, F.; Marrelli, M.; Carmela, C.; Menichini, F.; Valentina, P.; Uzunov, D.; Statti, G.A.; Duez, P.; Menichini, F. Bioactive phytonutrients (omega fatty acids, tocopherols, polyphenols), in vitro inhibition of nitric oxide production and free radical scavenging activity of non-cultivated Mediterranean vegetables. Food Chem. 2011, 129, 1413–1419. [Google Scholar] [CrossRef]
- Shen, S.C.; Lee, W.R.; Lin, H.Y.; Huang, H.C.; Ko, C.H.; Yang, L.L.; Chen, Y.C. In vitro and in vivo inhibitory activities of rutin, wogonin, and quercetin on lipopolysaccharide-induced nitric oxide and prostaglandin E2 production. Eur. J. Pharmacol. 2002, 446, 187–194. [Google Scholar] [CrossRef]
- Choi, S.Y.; Ko, H.C.; Ko, S.Y.; Hwang, J.H.; Park, J.G.; Kang, S.H.; Han, S.H.; Yun, S.H.; Kim, S.J. Correlation between flavonoid content and the no production inhibitory activity of peel extracts from various citrus fruits. Biol. Pharm. Bull. 2007, 30, 772–778. [Google Scholar] [CrossRef] [PubMed]
- Popko, K.; Gorska, E.; Stelmaszczyk-Emmel, A.; Plywaczewski, R.; Stoklosa, A.; Pyrzak, B.; Demkow, U. Proinflammatory cytokines IL-6 and TNF-α and the development of inflammation in obese subjects. Eur. J. Med. Res. 2010, 15, 120–122. [Google Scholar] [PubMed]
- Crichton, M.B.; Nichols, J.E.; Zhao, Y.; Bulun, S.E.; Simpson, E.R. Expression of transcripts of interleukin-6 and related cytokines by human breast tumors, breast cancer cells, and adipose stromal cells. Mol. Cell. Endocrinol. 1996, 118, 215–220. [Google Scholar] [CrossRef]
- Kumari, N.; Dwarakanath, B.S.; Das, A.; Bhatt, A.N. Role of interleukin-6 in cancer progression and therapeutic resistance. Tumor Biol. 2016, 37, 11553–11572. [Google Scholar] [CrossRef] [PubMed]
- Unver, N.; McAllister, F. IL-6 family cytokines: Key inflammatory mediators as biomarkers and potential therapeutic targets. Cytokine Growth Factor Rev. 2018, 41, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Zeng, K.W.; Ma, X.L.; Song, F.J.; Jiang, Y.; Tu, P.F.; Wang, X.M. Resokaempferol-mediated anti-inflammatory effects on activated macrophages via the inhibition of JAK2/STAT3, NF-κB and JNK/p38 MAPK signaling pathways. Int. Immunopharmacol. 2016, 38, 104–114. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.W.; Seo, U.K.; Kim, J.H.; Leem, K.H.; Park, Y.K. Flower extract of Panax notoginseng attenuates lipopolysaccharide-induced inflammatory response via blocking of NF-κB signaling pathway in murine macrophages. J. Ethnopharmacol. 2009, 122, 313–319. [Google Scholar] [CrossRef]
- Hanada, T.; Yoshimura, A. Regulation of cytokine signaling and inflammation. Cytokine Growth Factor Rev. 2002, 13, 413–421. [Google Scholar] [CrossRef]
- Jung, K.H.; Ha, E.; Kim, M.J.; Won, H.J.; Zheng, L.T.; Kim, H.K.; Hong, S.J.; Chung, J.H.; Yim, S.V. Suppressive effects of nitric oxide (NO) production and inducible nitric oxide synthase (iNOS) expression by Citrus reticulata extract in RAW 264.7 macrophage cells. Food Chem. Toxicol. 2007, 45, 1545–1550. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.X.; Zhang, S.H.; Wang, X.L.; Ye, T.J.; Li, H.; Yan, X.F.; Wei, L.; Wu, Z.P.; Hu, J.; Zou, C.P.; et al. Panax Notoginseng flower saponins (PNFS) inhibit LPS-stimulated NO overproduction and iNOS gene overexpression via the suppression of TLR4-mediated MAPK/NF-kappa B signaling pathways in RAW264.7 macrophages. Chin. Med. 2015, 10, 15. [Google Scholar] [CrossRef] [PubMed]
- DuBois, M.E.; DuBois, R.N. CYCLOOXYGENASE-2: A Therapeutic Target. Annu. Rev. Med. 2002, 53, 55–57. [Google Scholar] [CrossRef]
- Rhule, A.; Navarro, S.; Smith, J.R.; Shepherd, D.M. Panax notoginseng attenuates LPS-induced pro-inflammatory mediators in RAW264.7 cells. J. Ethnopharmacol. 2006, 106, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Park, S.B.; Park, G.H.; Um, Y.; Kim, H.N.; Song, H.M.; Kim, N.; Kim, H.S.; Jeong, J.B. Wood-cultivated ginseng exerts anti-inflammatory effect in LPS-stimulated RAW264.7 cells. Int. J. Biol. Macromol. 2018, 116, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Baek, K.S.; Yi, Y.S.; Son, Y.J.; Yoo, S.; Sung, N.Y.; Kim, Y.; Hong, S.; Aravinthan, A.; Kim, J.H.; Cho, J.Y. In vitro and in vivo anti-inflammatory activities of Korean Red Ginseng-derived components. J. Ginseng Res. 2016, 40, 437–444. [Google Scholar] [CrossRef]
- Yang, L.G.; Yin, P.P.; Li, K.; Fan, H.; Xue, Q.; Li, X.; Sun, L.W.; Liu, Y.J. Seasonal dynamics of constitutive levels of phenolic components lead to alterations of antioxidant capacities in Acer truncatum leaves. Arab. J. Chem. 2018, 11, 14–25. [Google Scholar] [CrossRef]
- Li, K.; Fan, H.; Yin, P.P.; Yang, L.G.; Xue, Q.; Li, X.; Sun, L.W.; Liu, Y.J. Structure-activity relationship of eight high content flavonoids analyzed with a preliminary assign-score method and their contribution to antioxidant ability of flavonoids-rich extract from Scutellaria baicalensis shoots. Arab. J. Chem. 2018, 11, 159–170. [Google Scholar] [CrossRef]
- Fan, H.; Sun, L.W.; Yang, L.G.; Zhou, J.C.; Yin, P.P.; Li, K.; Xue, Q.; Li, X.; Liu, Y.J. Assessment of the bioactive phenolic composition of Acer truncatum seed coat as a byproduct of seed oil. Ind. Crops Prod. 2018, 118, 11–19. [Google Scholar] [CrossRef]
- Xue, Q.; Fan, H.; Li, K.; Yang, L.G.; Sun, L.W.; Liu, Y.J. Comparative evaluations on phenolic antioxidants of nine adulterants and anti-inflammation of four alternatives with their original herb Erycibe schmidtii. RSC Adv. 2017, 7, 51151–51161. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ginsenoside | GSE0 | GSE2 | GSE4 | GSE6 |
|---|---|---|---|---|
| Re | 0.591 ± 0.031 a | 0.422 ± 0.023 b | 0.096 ± 0.008 c | 0.002 ± 0.000 d |
| Rg1 | 0.241 ± 0.019 a | 0.218 ± 0.002 b | 0.059 ± 0.004 c | 0.003 ± 0.000 d |
| Rd | 0.154 ± 0.008 a | 0.146 ± 0.059 ab | 0.089 ± 0.012 b | 0.021 ± 0.002 c |
| S-Rg2 | 0.125 ± 0.012 a | 0.115 ± 0.006 a | 0.121 ± 0.007 a | 0.088 ± 0.004 b |
| Rb2 | 0.072 ± 0.004 a | 0.059 ± 0.000 b | 0.026 ± 0.000 c | - |
| F1 | 0.061 ± 0.004 a | 0.058 ± 0.001 a | 0.031 ± 0.002 b | - |
| Rc | 0.058 ± 0.003 a | 0.046 ± 0.003 b | 0.052 ± 0.006 ab | 0.049 ± 0.003 b |
| S-Rh1 | 0.022 ± 0.000 c | 0.021 ± 0.001 c | 0.062 ± 0.002 b | 0.075 ± 0.002 a |
| Rb3 | 0.013 ± 0.001 a | 0.012 ± 0.002 a | - | - |
| R-Rg2 | - * | 0.001 ± 0.000 c | 0.044 ± 0.007 b | 0.052 ± 0.002 a |
| Rf | - | - | - | - |
| Total | 1.338 ± 0.083 a | 1.099 ± 0.099 b | 0.578 ± 0.049 c | 0.290 ± 0.013 d |
| Ginsenoside | GSE0 | GSE2 | GSE4 | GSE6 |
|---|---|---|---|---|
| F4 | 0.006 ± 0.001 c | 0.046 ± 0.007 b | 0.167 ± 0.004 a | 0.170 ± 0.004 a |
| S-Rg3 | - * | 0.025 ± 0.001 c | 0.120 ± 0.013 b | 0.149 ± 0.004 a |
| Rg5 | - | 0.012 ± 0.001 b | 0.050 ± 0.002 a | 0.047 ± 0.004 a |
| Rh2 | - | 0.006 ± 0.001 c | 0.031 ± 0.002 b | 0.046 ± 0.006 a |
| PPT | - | 0.003 ± 0.000 c | 0.015 ± 0.001 b | 0.024 ± 0.183 a |
| R-Rg3 | - | 0.001 ± 0.000 c | 0.007 ± 0.000 b | 0.014 ± 0.002 a |
| CK | - | 0.004 ± 0.000 b | 0.010 ± 0.001 a | 0.004 ± 0.000 b |
| Total | 0.006 ± 0.000 b | 0.098 ± 0.011 b | 0.400 ± 0.024 a | 0.455 ± 0.203 a |
| Gene | Primer Sequence (5′ to 3′) | bp |
|---|---|---|
| iNOS Forward | AATGGCAACATCAGGTCGGCCATCACT | 27 |
| iNOS Reverse | GCTGTGTGTCACAGAAGTCTCGAACTC | 27 |
| COX-2 | TGAA GCCGTACACATCATTTGAA | 23 |
| COX-2 | TGGTCTCCCCAAAGATAGCATCT | 23 |
| IL-6 | GGGGATGTCTGTAGCTCATTCTGCTCTG | 28 |
| IL-6 | AAGGACTCTGGCTTTGTCTTTCTTGTTA | 28 |
| TNF-α | CTGTGAAGGGAATGGGTGTT | 20 |
| TNF-α | CAGGGAAGAATCTGGAAAGGTC | 22 |
| GraPDH | TGTTTCCTCGTCCCGTAG | 18 |
| GraPDH | CAATCTCCACTTTGCCACT | 19 |
| TPG | TLPG | TP | TF | DPPH | ABTS | NO-r | IL-6-r | TNF-α-r | iNOS-mR | IL-6-mR | TNF-α-mR | COX-2-mR | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| TPG | 1 | ||||||||||||
| TLPG | −0.988 * | 1 | |||||||||||
| TP | −0.923 | 0.918 | 1 | ||||||||||
| TF | −0.922 | 0.891 | 0.984 * | 1 | |||||||||
| DPPH | −0.924 | 0.965 * | 0.934 | 0.870 | 1 | ||||||||
| ABTS | −0.842 | 0.817 | 0.976 * | 0.982 * | 0.837 | 1 | |||||||
| GSE0, GSE2, GSE4, GSE6 at 25 µg/mL | |||||||||||||
| NO-r | 0.921 | −0.903 | −0.996 ** | −0.996 ** | −0.901 | −0.985 * | 1 | ||||||
| IL-6-r | 0.898 | −0.952 * | −0.895 | −0.817 | −0.995 ** | −0.782 | 0.855 | 1 | |||||
| TNF-α-r | 0.974 * | −0.994 ** | −0.941 | −0.902 | −0.987 * | −0.846 | 0.920 | 0.974 * | 1 | ||||
| iNOS-mR | 0.930 | −0.930 | −0.999 ** | −0.979 * | −0.946 | −0.967 * | 0.992 ** | 0.911 | 0.953 * | 1 | |||
| IL-6-mR | 0.993 ** | −0.991 ** | −0.880 | −0.870 | −0.922 | −0.777 | 0.873 | 0.906 | 0.971 * | 0.892 | 1 | ||
| TNF-α-mR | 0.986 * | −0.981 * | −0.974 * | −0.962 * | −0.955 * | −0.911 | 0.968 * | 0.925 | 0.984 * | 0.979 * | 0.965 * | 1 | |
| COX-2-mR | 0.846 | −0.824 | −0.575 | −0.599 | −0.663 | −0.440 | 0.580 | 0.659 | 0.760 | 0.592 | 0.887 | 0.745 | 1 |
| GSE0, GSE2, GSE4, GSE6 at 50 µg/mL | |||||||||||||
| NO-r | 0.625 | −0.725 | −0.736 | −0.605 | −0.876 | −0.683 | 1 | ||||||
| IL-6-r | 0.980 * | −0.989 * | −0.854 | −0.831 | −0.928 | −0.734 | 0.661 | 1 | |||||
| TNF-α-r | 0.978 * | −0.992 ** | −0.959 * | −0.927 | −0.982 * | −0.876 | 0.769 | 0.962 * | 1 | ||||
| iNOS-mR | 0.900 | −0.883 | −0.661 | −0.677 | −0.741 | −0.531 | 0.384 | 0.938 | 0.817 | 1 | |||
| IL-6-mR | 0.991 ** | −0.969 * | −0.868 | −0.881 | −0.873 | −0.780 | 0.532 | 0.978 * | 0.944 | 0.944 | 1 | ||
| TNF-α-mR | 0.920 | −0.930 | −0.995 ** | −0.963 * | −0.961 * | −0.956 * | 0.797 | 0.867 | 0.970 * | 0.662 | 0.860 | 1 | |
| COX-2-mR | 0.966 * | −0.932 | −0.968 * | −0.988 * | −0.881 | −0.941 | 0.575 | 0.894 | 0.949 | 0.781 | 0.942 | 0.947 | 1 |
| GSE0, GSE2, GSE4, GSE6 at 100 µg/mL | |||||||||||||
| NO-r | 0.995 ** | −0.987 * | −0.881 | −0.877 | −0.911 | −0.782 | 1 | ||||||
| IL-6-r | 0.971 * | −0.982 * | −0.974 * | −0.944 | −0.980 * | −0.903 | 0.951 * | 1 | |||||
| TNF-α-r | 0.991 ** | −0.959 * | −0.922 | −0.943 | −0.877 | −0.865 | 0.979 * | 0.949 | 1 | ||||
| iNOS-mR | 0.965 * | −0.977 * | −0.978 * | −0.946 | −0.981 * | −0.910 | 0.943 | 1.000 ** | 0.943 | 1 | |||
| IL-6-mR | 0.996 ** | −0.974 * | −0.893 | −0.905 | −0.887 | −0.813 | 0.995 ** | 0.945 | 0.994 ** | 0.937 | 1 | ||
| TNF-α-mR | 0.934 | −0.932 | −0.999 ** | −0.981 * | −0.944 | −0.968 * | 0.895 | 0.982 * | 0.929 | 0.985 * | 0.904 | 1 | |
| COX-2-mR | 0.895 | −0.881 | −0.996 ** | −0.990 ** | −0.898 | −0.992 ** | 0.845 | 0.950 * | 0.904 | 0.955 * | 0.865 | 0.992 ** | 1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yao, F.; Xue, Q.; Li, K.; Cao, X.; Sun, L.; Liu, Y. Phenolic Compounds and Ginsenosides in Ginseng Shoots and Their Antioxidant and Anti-Inflammatory Capacities in LPS-Induced RAW264.7 Mouse Macrophages. Int. J. Mol. Sci. 2019, 20, 2951. https://doi.org/10.3390/ijms20122951
Yao F, Xue Q, Li K, Cao X, Sun L, Liu Y. Phenolic Compounds and Ginsenosides in Ginseng Shoots and Their Antioxidant and Anti-Inflammatory Capacities in LPS-Induced RAW264.7 Mouse Macrophages. International Journal of Molecular Sciences. 2019; 20(12):2951. https://doi.org/10.3390/ijms20122951
Chicago/Turabian StyleYao, Fan, Qiang Xue, Ke Li, Xinxin Cao, Liwei Sun, and Yujun Liu. 2019. "Phenolic Compounds and Ginsenosides in Ginseng Shoots and Their Antioxidant and Anti-Inflammatory Capacities in LPS-Induced RAW264.7 Mouse Macrophages" International Journal of Molecular Sciences 20, no. 12: 2951. https://doi.org/10.3390/ijms20122951
APA StyleYao, F., Xue, Q., Li, K., Cao, X., Sun, L., & Liu, Y. (2019). Phenolic Compounds and Ginsenosides in Ginseng Shoots and Their Antioxidant and Anti-Inflammatory Capacities in LPS-Induced RAW264.7 Mouse Macrophages. International Journal of Molecular Sciences, 20(12), 2951. https://doi.org/10.3390/ijms20122951