Genome-Wide Identification and Characterization of the Cyclophilin Gene Family in the Nematophagous Fungus Purpureocillium lilacinum


Abstract
:1. Introduction
2. Results
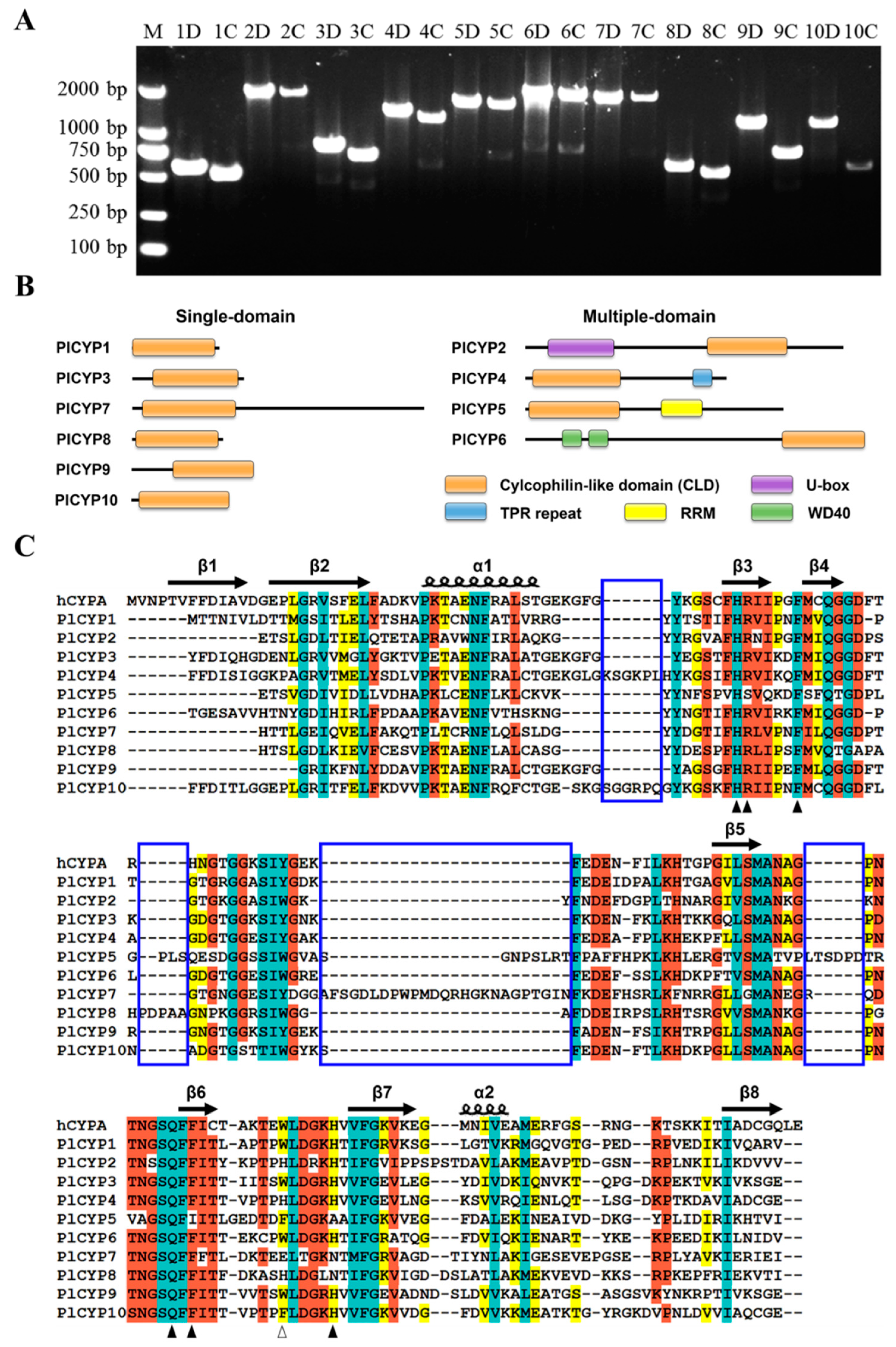
2.1. Ten CYP Genes Were Identified in P. lilacinum
2.2. The PlCYPs Contain the CLD Domain with Residue Variation
2.3. Phylogenetic Analysis Showed Evolutionary Divergence of CYPs between Ascomycotina and Saccharomycotina Fungi
2.4. The CYPs in Ascomycotina Fungi Display a More Complex Motif and Gene Structure Distribution than the CYPs in Saccharomycotina
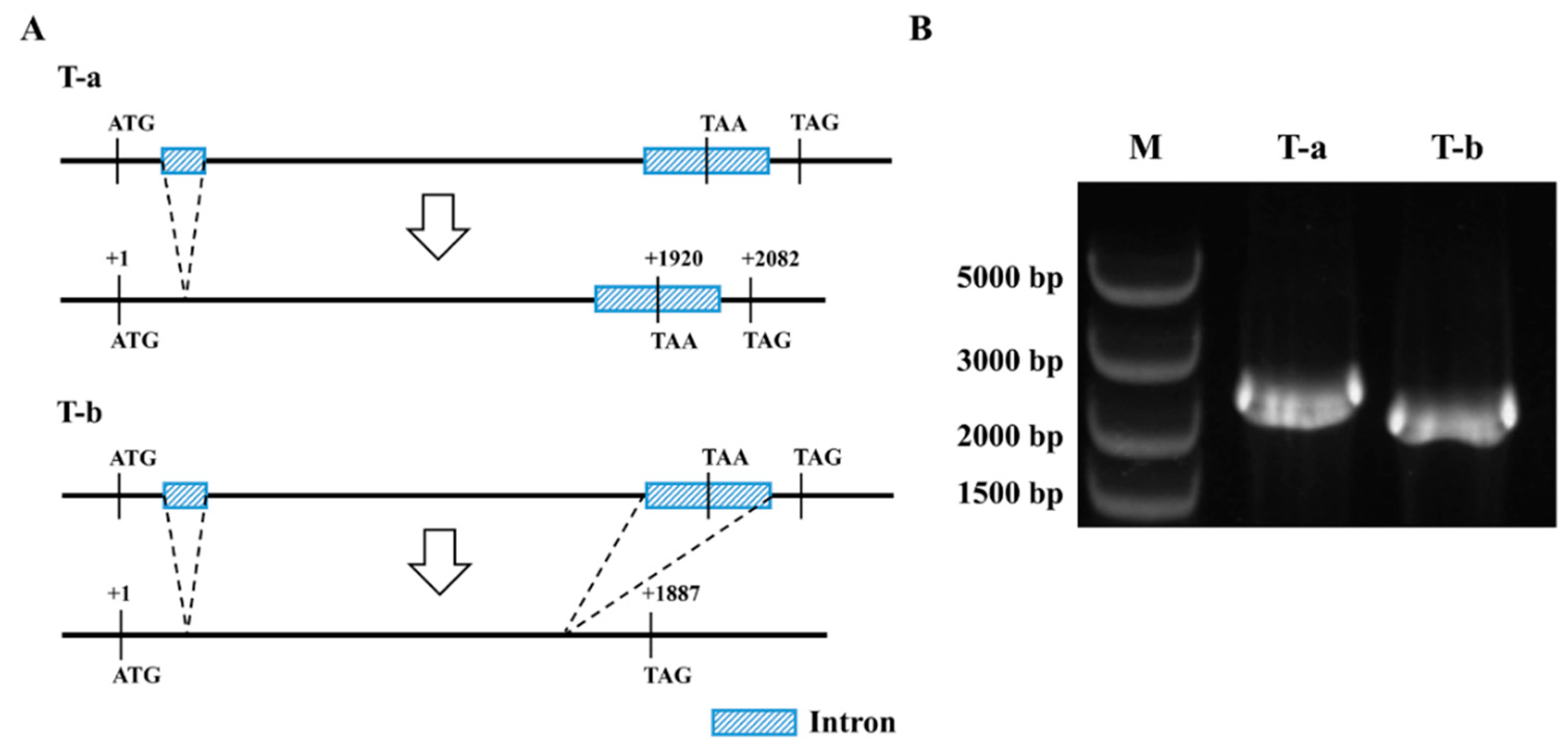
2.5. The PlCYP6 Gene Has Two Transcripts
2.6. PlCYPs Exhibit Different Expression Patterns in Different Fungal Morphologies and at the Egg Parasitic Stage
2.7. The Expression of PlCYP4 and PlCYP6 Were Induced under High Temperature and H2O2 Stresses
2.8. Heterologous Expressions of PlCYP4 and PlCYP6 in E. coli Enhance Tolerance towards Abiotic Stresses
3. Discussion
4. Materials and Methods
4.1. Fungal Strains and Growth Condition
4.2. Identification and Classification of CYP Genes in P. lilacinum Strain 36-1
4.3. Protein Alignment and Phylogenetic Analysis
4.4. Motifs and Gene Structure Analyses
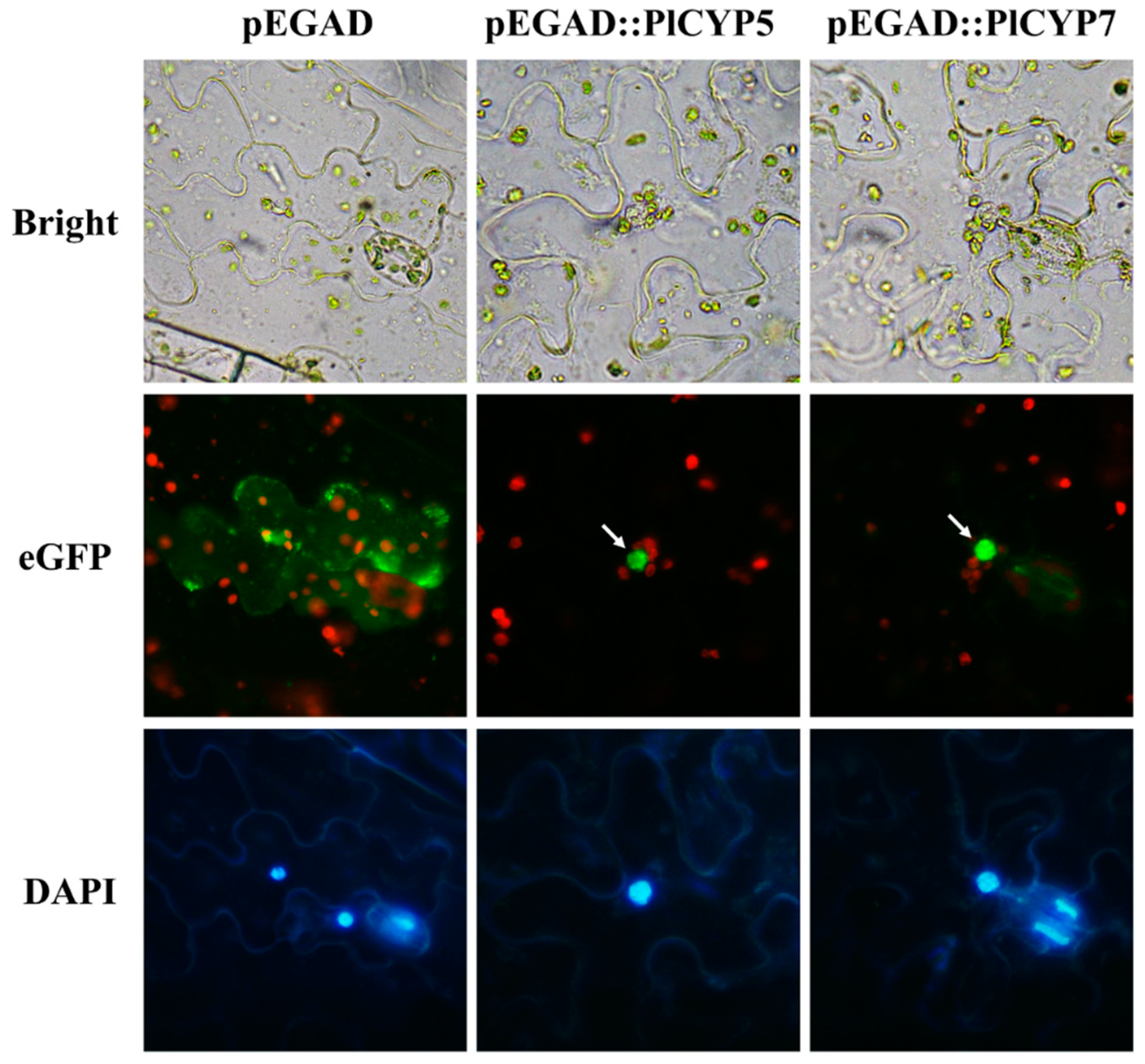
4.5. Transient Expression of Tobacco
4.6. Sample Treatments for qRT-PCR
4.7. RNA Extraction and qRT-PCR
4.8. Phenotypic Assay of PlCYP4 and PlCYP6 in E. coli
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Abad, P.; Gouzy, J.; Aury, J.; Castagnone-Sereno, P.; Danchin, E.G.J.; Deleury, E.; Caillaud, M.C. Genome sequence of the metazoan plant-parasitic nematode Meloidogyne incognita. Nat. Biotechnol. 2008, 26, 909–915. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.T.; Haegeman, A.; Danchin, E.G.J.; Gaur, H.S.; Helder, J.; Jones, M.G.K.; Perry, R.N. Top 10 plant-parasitic nematodes in molecular plant pathology. Mol. Plant Pathol. 2013, 14, 946–961. [Google Scholar] [CrossRef] [PubMed]
- Atkins, S.D.; Clark, I.M.; Pande, S.; Hirsch, P.R.; Kerry, B.R. The use of real-time PCR and species-specific primers for the identification and monitoring of Paecilomyces lilacinus. FEMS Microbiol. Ecol. 2005, 51, 257–264. [Google Scholar] [CrossRef]
- Anastasiadis, I.A.; Giannakou, I.O.; Prophetou-Athanasiadou, D.A.; Gowen, S.R. The combined effect of the application of a biocontrol agent Paecilomyces lilacinus, with various practices for the control of root-knot nematodes. Crop Prot. 2008, 27, 352–361. [Google Scholar] [CrossRef]
- Huang, W.K.; Cui, J.K.; Liu, S.M.; Kong, L.A.; Wu, Q.S.; Peng, H.; Peng, D.L. Testing various biocontrol agents against the root-knot nematode (Meloidogyne incognita) in cucumber plants identifies a combination of Syncephalastrum racemosum and Paecilomyces lilacinus as being most effective. Biol. Control 2016, 92, 31–37. [Google Scholar] [CrossRef]
- Hajji, L.; Hlaoua, W.; Regaieg, H.; Horrigue-Raouani, N. Biocontrol potential of Verticillium leptobactrum and Purpureocillium lilacinum Against Meloidogyne javanica and Globodera pallida on Potato (Solanum tuberosum). Am. J. Patato Res. 2017, 94, 178–183. [Google Scholar] [CrossRef]
- Kiewnick, S.; Neumann, S.; Sikora, R.A.; Frey, J.E. Effect of Meloidogyne incognita inoculum density and application rate of Paecilomyces lilacinus strain 251 on biocontrol efficacy and colonization of egg masses analyzed by real-time quantitative PCR. Phytopathology 2011, 101, 105–112. [Google Scholar] [CrossRef]
- Prasad, P.; Varshney, D.; Adholeya, A. Whole genome annotation and comparative genomic analyses of bio-control fungus Purpureocillium lilacinum. BMC Genom. 2015, 16, 1004. [Google Scholar] [CrossRef]
- Wang, G.; Liu, Z.G.; Lin, R.M.; Li, E.F.; Mao, Z.C.; Ling, J.; Xie, B. Biosynthesis of antibiotic leucinostatins in bio-control fungus purpureocillium lilacinum and their inhibition on Phytophthora revealed by genome mining. PloS Pathog. 2016, 12, e1005685. [Google Scholar] [CrossRef]
- Xie, J.L.; Li, S.J.; Mo, C.M.; Xiao, X.Q.; Peng, D.L.; Wang, G.F.; Xiao, Y. Genome and transcriptome sequences reveal the specific parasitism of the nematophagous Purpureocillium lilacinum 36-1. Front. Microbiol. 2016, 7, 1084. [Google Scholar] [CrossRef]
- Handschumacher, R.E.; Harding, M.W.; Rice, J.; Drugge, R.J.; Speicher, D.W. Cyclophilin: A specific cytosolic binding protein for cyclosporin A. Science 1984, 226, 544–547. [Google Scholar] [CrossRef] [PubMed]
- Gothel, S.F.; Marahiel, M.A. Peptidyl-prolyl cis-trans isomerases, a superfamily of ubiquitous folding catalysts. Cell. Mol. Life Sci. 1999, 55, 423–436. [Google Scholar] [CrossRef] [PubMed]
- Schiene-Fischer, C. Multidomain peptidyl prolyl cis/trans isomerases. Biochim. Biophys. Acta Gen. Subj. 2014, 1850, 2005–2016. [Google Scholar] [CrossRef] [PubMed]
- Hayano, T.; Takahashi, N.; Kato, S.; Maki, N.; Suzuki, M. Two distinct forms of peptidylprolyl-cis-trans-isomerase are expressed separately in periplasmic and cytoplasmic compartments of Escherichia coli cells. Biochemistry 1991, 30, 3041–3048. [Google Scholar] [CrossRef] [PubMed]
- Andrzej, G. Peptidylprolyl cis/trans isomerases (immunophilins): Biological diversity-targets-functions. Curr. Top. Med. Chem. 2003, 3, 1315–1347. [Google Scholar] [CrossRef]
- Liu, J.; Farmer, J.D.; Lane, W.S.; Friedman, J.; Weissman, I.; Schreiber, S.L. Calcineurin is a common target of cyclophilin-cyclosporin A and FKBP-FK506 complexes. Cell 1991, 66, 807–815. [Google Scholar] [CrossRef]
- Pemberton, T.J.; Kay, J.E. Identification and comparative analysis of the peptidyl-prolyl cis/trans isomerase repertoires of H. sapiens, D. melanogaster, C. elegans, S. cerevisiae and Sz. pombe. Comp. Funct. Genom. 2005, 6, 277–300. [Google Scholar] [CrossRef] [PubMed]
- Gan, P.H.P.; Shan, W.X.; Blackman, L.M.; Hardham, A.R. Characterization of cyclophilin-encoding genes in Phytophthora. Mol. Genet. Genom. 2009, 281, 565–578. [Google Scholar] [CrossRef]
- Arevalo-Rodriguez, M.; Wu, X.; Hanes, S.D.; Heitman, J. Prolyl isomerases in yeast. Front. Biosci. 2004, 9, 2420–2446. [Google Scholar] [CrossRef] [Green Version]
- Dolinski, K.; Muir, S.; Cardenas, M.; Heitman, J. All cyclophilins and FK506 binding proteins are, individually and collectively, dispensable for viability in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 1997, 94, 13093–13098. [Google Scholar] [CrossRef]
- Trivedi, D.K.; Yadav, S.; Vaid, N.; Tuteja, N. Genome wide analysis of Cyclophilin gene family from rice and Arabidopsis and its comparison with yeast. Plant Signal. Behav. 2012, 7, 1653–1666. [Google Scholar] [CrossRef] [PubMed]
- Mainali, H.R.; Chapman, P.; Dhaubhadel, S. Genome-wide analysis of Cyclophilin gene family in soybean (Glycine max). BMC Plant Biol. 2014, 14, 282. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.Q.; Wang, Y.; Chai, W.B.; Song, N.N.; Wang, J.; Cao, L.M.; Li, X. Systematic analysis of the maize cyclophilin gene family reveals ZmCYP15 involved in abiotic stress response. Plant Cell Tissue Organ Cult. 2017, 128, 543–561. [Google Scholar] [CrossRef]
- Anna, K.; Janine, B.; Anne-Marie, K.; Jean, C.; Maria, J.D.; Catherine, K. KIN241: A gene involved in cell morphogenesis in Paramecium tetraurelia reveals a novel protein family of cyclophilin-RNA interacting proteins (CRIPs) conserved from fission yeast to man. Mol. Microbiol. 2001, 42, 257–267. [Google Scholar] [CrossRef]
- Duina, A.A.; Chang, H.C.; Marsh, J.A.; Lindquist, S.; Gaber, R.F. A cyclophilin function in Hsp90-dependent signal transduction. Science 1996, 274, 1713–1715. [Google Scholar] [CrossRef] [PubMed]
- Horowitz, D.S.; Lee, E.J.; Mabon, S.A.; Misteli, T. A cyclophilin functions in pre-mRNA splicing. EMBO J. 2002, 21, 470–480. [Google Scholar] [CrossRef] [PubMed]
- Shaw, P.E. Peptidyl-prolyl isomerases: A new twist to transcription. EMBO Rep. 2002, 3, 521–526. [Google Scholar] [CrossRef] [PubMed]
- Kumari, S.; Singh, P.; Singla-Pareek, S.L.; Pareek, A. Heterologous expression of a salinity and developmentally regulated rice cyclophilin gene (OsCyp2) in E. coli and S. cerevisiae confers tolerance towards multiple abiotic stresses. Mol. Biotechnol. 2009, 43, 195–204. [Google Scholar] [CrossRef]
- Ruan, S.L.; Ma, H.S.; Wang, S.H.; Fu, Y.P.; Xin, Y.; Liu, W.Z.; Chen, H.Z. Proteomic identifcation of OsCYP2, a rice cyclophilin that confers salt tolerance in rice (Oryza sativa L.) seedlings when overexpressed. BMC Plant Biol. 2011, 11, 1117–1120. [Google Scholar] [CrossRef]
- Trivedi, D.K.; Ansari, M.W.; Tuteja, N. Multiple abiotic stress responsive rice cyclophilin: (OsCYP-25) mediates a wide range of cellular responses. Commun. Integr. Biol. 2013, 6, e25260. [Google Scholar] [CrossRef]
- Kim, S.K.; You, Y.N.; Park, J.C.; Joung, Y.; Kim, B.G.; Ahn, J.C.; Cho, H.S. The rice thylakoid luminal cyclophilin OsCYP20-2 confers enhanced environmental stress tolerance in tobacco and Arabidopsis. Plant Cell Rep. 2012, 31, 417–426. [Google Scholar] [CrossRef] [PubMed]
- Chu, Z.J.; Sun, H.H.; Ying, S.H.; Feng, M.G. Vital role for cyclophilin B (CypB) in asexual development, dimorphic transition and virulence of Beauveria bassiana. Fungal Genet. Biol. 2017, 105, 8–15. [Google Scholar] [CrossRef]
- Viaud, M.C.; Balhadere, P.V.; Talbot, N.J. A Magnaporthe grisea cyclophilin acts as a virulence determinant during plant infection. Plant Cell 2002, 14, 917–930. [Google Scholar] [CrossRef] [PubMed]
- Viaud, M.; Brunet-Simon, A.; Brygoo, Y.; Pradier, J.M.; Levis, C. Cyclophilin A and calcineurin functions investigated by gene inactivation, cyclosporin A inhibition and cDNA arrays approaches in the phytopathogenic fungus Botrytis cinerea. Mol. Microbiol. 2003, 50, 1451–1465. [Google Scholar] [CrossRef] [PubMed]
- Gullerova, M.; Barta, A.; Lorkovic, Z.J. AtCyp59 is a multidomain cyclophilin from Arabidopsis thaliana that interacts with SR proteins and the C-terminal domain of the RNA polymerase II. RNA 2006, 12, 631–643. [Google Scholar] [CrossRef] [PubMed]
- Gullerova, M.; Barta, A.; Lorkovic, Z.J. Rct1, a nuclear RNA recognition motif-containing cyclophilin, regulates phosphorylation of the RNA polymerase II C-terminal domain. Mol. Cell. Biol. 2007, 27, 3601–3611. [Google Scholar] [CrossRef]
- Zydowsky, L.D.; Etzkorn, F.A.; Chang, H.Y.; Ferguson, S.B.; Stolz, L.A.; Ho, S.; Walsh, C.T. Active-site mutants of human cyclophilin-a separate peptidyl-prolyl isomerase activity from cyclosporine-a binding and calcineurin inhibition. Protein Sci. 1992, 1, 1092–1099. [Google Scholar] [CrossRef] [PubMed]
- Pemberton, T.J. Identification and comparative analysis of sixteen fungal peptidyl-prolyl cis/trans isomerase repertoires. BMC Genom. 2006, 7, 244. [Google Scholar] [CrossRef]
- Han, Y.H.; Ding, T.; Su, B.; Jiang, H.Y. Genome-wide identification, characterization and expression analysis of the chalcone synthase family in maize. Int. J. Mol. Sci. 2016, 17, 161. [Google Scholar] [CrossRef]
- Wang, G.; Wang, T.; Jia, Z.H.; Xuan, Z.P.; Pan, D.L.; Guo, Z.R.; Zhang, J.Y. Genome-wide bioinformatics analysis of MAPK gene family in kiwifruit (Actinidia chinensis). Int. J. Mol. Sci. 2018, 19, 2510. [Google Scholar] [CrossRef]
- Li, J.B.; Zhang, J.; Jia, H.X.; Yue, Z.Q.; Lu, M.Z.; Xin, X.B.; Hu, J.J. Genome-wide characterization of the sHsp gene family in Salix suchowensis reveals its functions under different abiotic stresses. Int. J. Mol. Sci. 2018, 19, 3246. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Heitman, J. The cyclophilins. Genome Biol. 2005, 6, 226. [Google Scholar] [CrossRef] [PubMed]
- Cutler, S.R.; Ehrhardt, D.W.; Griffitts, J.S.; Somerville, C.R. Random GFP::cDNA fusions enable visualization of subcellular structures in cells of Arabidopsis at a high frequency. Proc. Natl. Acad. Sci. USA 2000, 97, 3718–3723. [Google Scholar] [CrossRef] [PubMed]
- Ying, S.H.; Feng, M.G. Novel blastospore-based transformation system for integration of phosphinothricin resistance and green fluorescence protein genes into Beauveria bassiana. Appl. Microbiol. Biotechnol. 2006, 72, 206–210. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Orthologs in Strain PLFJ-1 | ORF (bp) | Deduced Polypeptide | Predicted Subcellular Localization | ||
|---|---|---|---|---|---|---|
| Amino Acid (aa) | Molecular Weight (kDa) | Isoelectric Point | ||||
| PlCYP1 | XP_018182827.1 | 489 | 162 | 17.4 | 6.06 | Cytoplasm |
| PlCYP2 | XP_018178542.1 | 1764 | 587 | 64.0 | 8.49 | Cytoplasm |
| PlCYP3 | XP_018176208.1 | 627 | 208 | 22.8 | 7.89 | Endoplasmic reticulum |
| PlCYP4 | XP_018179002.1 | 1119 | 372 | 40.3 | 5.81 | Cytoplasm |
| PlCYP5 | XP_018178952.1 | 1437 | 478 | 54.8 | 5.93 | Nucleus |
| PlCYP6 | XP_018179645.1 | 1884 | 627 | 70.2 | 6.49 | Cytoplasm |
| PlCYP7 | XP_018177409.1 | 1623 | 540 | 60.2 | 5.98 | Nucleus |
| PlCYP8 | XP_018177405.1 | 507 | 168 | 18.1 | 7.01 | Cytoplasm |
| PlCYP9 | XP_018174420.1 | 684 | 227 | 24.6 | 9.49 | Mitochondria |
| PlCYP10 | XP_018175923.1 | 549 | 182 | 19.8 | 6.29 | Cytoplasm |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mo, C.; Xie, C.; Wang, G.; Liu, J.; Hao, Q.; Xiao, X.; Xiao, Y. Genome-Wide Identification and Characterization of the Cyclophilin Gene Family in the Nematophagous Fungus Purpureocillium lilacinum. Int. J. Mol. Sci. 2019, 20, 2978. https://doi.org/10.3390/ijms20122978
Mo C, Xie C, Wang G, Liu J, Hao Q, Xiao X, Xiao Y. Genome-Wide Identification and Characterization of the Cyclophilin Gene Family in the Nematophagous Fungus Purpureocillium lilacinum. International Journal of Molecular Sciences. 2019; 20(12):2978. https://doi.org/10.3390/ijms20122978
Chicago/Turabian StyleMo, Chenmi, Chong Xie, Gaofeng Wang, Juan Liu, Qiuyan Hao, Xueqiong Xiao, and Yannong Xiao. 2019. "Genome-Wide Identification and Characterization of the Cyclophilin Gene Family in the Nematophagous Fungus Purpureocillium lilacinum" International Journal of Molecular Sciences 20, no. 12: 2978. https://doi.org/10.3390/ijms20122978
APA StyleMo, C., Xie, C., Wang, G., Liu, J., Hao, Q., Xiao, X., & Xiao, Y. (2019). Genome-Wide Identification and Characterization of the Cyclophilin Gene Family in the Nematophagous Fungus Purpureocillium lilacinum. International Journal of Molecular Sciences, 20(12), 2978. https://doi.org/10.3390/ijms20122978