Galectin 13 (PP13) Facilitates Remodeling and Structural Stabilization of Maternal Vessels during Pregnancy

 ,
,  and
and
Abstract
:1. Galectins
2. The Placental Galectins
3. Galectin 13
3.1. The PP13 Protein and its mRNA
3.2. Insights on the Gene and Protein Structures
3.3. PP13 Secretion from the Placenta
3.4. PP13 and Preeclampsia
3.5. PP13 Polymorphism and Preeclampsia
3.6. PP13 and Immune Tolerance
3.7. PP13 Replenishing Studies in Animals
3.8. Modeling the Role of PP13 in Pregnancy
4. Multiple Galectins and Deep Placentation
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ABO blood groups | Blood group types A, B, AB and O |
| BP | Base pair |
| CRD | Carbohydrate-recognition domain |
| EC50 | Effective dose for reaching 50% effect |
| Enos | Endothelial nitric oxide synthase |
| Gal 1, Gal 3, etc. | Galectin 1, galectin 3 and other galectins according to their nomenclature |
| FGR | Fetal growth restriction |
| kDa | Kilo-Dalton |
| pMol | Pico molar quantity (10-12 M) |
| PP13 | Placental protein 13 also called gal-13 and LGALS13 (gene) |
| STBM | Syncytiotrophoblast microparticles |
| WBC | White blood cells |
References
- Barondes, S.H.; Castronovo, V.; Cooper, D.N.W.; Cummings, R.D.; Drickamer, K.; Felzi, T.; Gitt, M.A.; Hirabayashi, J.; Hughes, C.; Kasai, K.; et al. Galectins: A family of animal beta-galactoside-binding lectins. Cell 1994, 76, 597–598. [Google Scholar] [CrossRef]
- Johannes, L.; Jacob, R.; Leffler, H. Galectins at a glance. J. Cell Sci. 2018, 131, jcs208884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindstedt, R.; Apodaca, G.; Barondes, S.H.; Mostov, K.E.; Leffler, H. Apical secretion of a cytosolic protein by Madin-Darby canine kidney cells. Evidence for polarized release of an endogenous lectin by a nonclassical secretory pathway. J. Biol. Chem. 1993, 268, 11750–11757. [Google Scholar] [PubMed]
- Leffler, H.; Carlsson, S.; Hedlund, M.; Qian, Y.; Poirier, F. Introduction to galectins. Glycoconj. J. 2004, 19, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Visegrády, B.; Than, N.G.; Kilár, F.; Sümegi, B.; Than, G.N.; Bohn, H. Homology modelling and molecular dynamics studies of human placental tissue protein 13 (galectin-13). Protein Eng. 2001, 14, 875–880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Lella, S.; Sundblad, V.; Cerliani, J.P.; Guardia, C.M.; Estrin, D.A.; Vasta, G.R.; Rabinovich, G.A. When galectins recognize glycans: From biochemistry to physiology and back again. Biochemistry 2011, 50, 7842–7857. [Google Scholar] [CrossRef] [PubMed]
- Hirabayashi, J.; Hashidate, T.; Arata, Y.; Nishi, N.; Nakamura, T.; Hirashima, M.; Urashima, T.; Oka, T.; Futai, M.; Muller, W.E.; et al. Oligosaccharide specificity of galectins: A search by frontal affinity chromatography. Biochim. Biophys. Acta 2002, 1572, 232–254. [Google Scholar] [CrossRef]
- Kamili, N.A.; Arthur, C.M.; Gerner-Smidt, C.; Tafesse, E.; Blenda, A.; Dias-Baruffi, M.; Stowell, S.R. Key regulators of galectin-glycan interactions. Proteomics 2016, 16, 3111–3125. [Google Scholar] [CrossRef] [PubMed]
- Salomonsson, E.; Carlsson, M.C.; Osla, V.; Hendus-Altenburger, R.; Kahl-Knutson, B.; Öberg, C.T.; Sundin, A.; Nilsson, R.; Nordberg-Karlsson, E.; Nilsson, U.J.; et al. Mutational tuning of galectin-3 specificity and biological function. J. Biol. Chem. 2010, 285, 35079–35091. [Google Scholar] [CrossRef] [PubMed]
- Than, N.G.; Pick, E.; Bellyei, S.; Szigeti, A.; Burger, O.; Berente, Z.; Janaky, T.; Boronkai, A.; Kliman, H.; Meiri, H.; et al. Functional analyses of placental protein 13/galectin-13. Eur. J. Biochem. 2004, 71, 1065–1078. [Google Scholar] [CrossRef]
- Than, N.G.; Romero, R.; Meiri, H.; Erez, O.; Xu, Y.; Tarquini, F.; Barna, L.; Szilagyi, A.; Ackerman, R.; Sammar, M.; et al. PP13, maternal ABO blood groups and the risk assessment of pregnancy complications. PLoS ONE 2011, 6, e21564. [Google Scholar] [CrossRef] [PubMed]
- Seelenmeyer, C.; Wegehingel, S.; Tews, I.; Künzler, M.; Aebi, M.; Nickel, W. Cell surface counter receptors are essential components of the unconventional export machinery of galectin-1. J. Cell Biol. 2005, 171, 373–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, D.K.; Zuberi, R.I.; Liu, F.T. Biochemical and biophysical characterization of human recombinant IgE-binding protein, an S-type animal lectin. J. Biol. Chem. 1992, 267, 14167–14174. [Google Scholar] [PubMed]
- Massa, S.M.; Cooper, D.N.W.; Leffier, H.; Barondes, S.H. L-29, an endogenous lectin, binds to glycoconjugates ligands with positive cooperativity. Biochemistry 1993, 32, 260–267. [Google Scholar] [CrossRef] [PubMed]
- Kuklinski, S.; Probstmeier, R. Homophilic binding properties of galectin-3: Involvement of the carbohydrate recognition domain. J. Neurochem. 1998, 70, 814–823. [Google Scholar] [CrossRef] [PubMed]
- Banani, S.F.; Lee, H.O.; Hyman, A.A.; Rosen, M.K. Biomolecular condensates: Organizers of cellular biochemistry. Nat. Rev. Mol. Cell Biol. 2017, 18, 285–298. [Google Scholar] [CrossRef] [PubMed]
- Dennis, J.W. Many light touches convey the message. Trends Biochem. Sci. 2015, 40, 673–686. [Google Scholar] [CrossRef]
- Dagher, S.F.; Wang, J.L.; Patterson, R.J. Identification of galectin-3 as a factor in pre-mRNA splicing. Proc. Natl. Acad. Sci. USA 1995, 92, 1213–1217. [Google Scholar] [CrossRef] [PubMed]
- Vyakarnam, A.; Dagher, S.F.; Wang, J.L.; Patterson, R.J. Evidence for a role for galectin-1 in pre-mRNA splicing. Mol. Cell. Biol. 1997, 17, 4730–4737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michaud, S.; Reed, R. An ATP-independent complex commits pre-mRNA to the mammalian spliceosome assembly pathway. Genes Dev. 1991, 5, 2534–2546. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.G.; Kim, D.H.; Kim, S.J.; Cho, Y.; Jung, J.; Jang, W.; Chun, K.H. Galectin-3 supports stemness in ovarian cancer stem cells by activation of the Notch1 intracellular domain. Oncotarget 2016, 7, 68229–68241. [Google Scholar] [CrossRef] [PubMed]
- Blidner, A.G.; Méndez-Huergo, S.P.; Cagnoni, A.J.; Rabinovich, G.A. Re-wiring regulatory cell networks in immunity by galectin-glycan interactions. FEBS Lett. 2015, 589, 3407–3418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thiemann, S.; Baum, L.G. Galectins and immune responses-just how do they do those things they do? Annu. Rev. Immunol. 2016, 34, 243–264. [Google Scholar] [CrossRef] [PubMed]
- Glinsky, V.V.; Raz, A. Modified citrus pectin anti-metastatic properties: One bullet, multiple targets. Carbohydr. Res. 2009, 344, 1788–1791. [Google Scholar] [CrossRef] [PubMed]
- Glinsky, V.V.; Glinsky, G.V.; Rittenhouse-Olson, K.; Huflejt, M.E.; Glinskii, O.V.; Deutscher, S.L.; Quinn, T.P. The role of Thomsen-Friedenreich antigen in adhesion of human breast and prostate cancer cells to the endothelium. Cancer Res. 2001, 61, 4851–4857. [Google Scholar]
- Sindrewicz, P.; Lian, L.Y.; Yu, L.G. Interaction of the Oncofetal Thomsen-Friedenreich antigen with galectins in cancer progression and metastasis. Front. Oncol. 2016, 6, 79. [Google Scholar] [CrossRef]
- Li, S.; Wandel, M.P.; Li, F.; Liu, Z.; He, C.; Wu, J.; Shi, Y.; Randow, F. Sterical hindrance promotes selectivity of the autophagy cargo receptor NDP52 for the danger receptor galectin-8 in antibacterial autophagy. Sci. Signal. 2013, 6, 9. [Google Scholar] [CrossRef]
- Chen, W.S.; Cao, Z.; Leffler, H.; Nilsson, U.J.; Panjwani, N. Galectin-3 inhibition by a small-molecule inhibitor reduces both pathological corneal neovascularization and fibrosis. Investig. Ophthalmol. Vis. Sci. 2017, 58, 9–20. [Google Scholar] [CrossRef]
- Delaine, T.; Collins, P.; MacKinnon, A.; Sharma, G.; Stegmayr, J.; Rajput, V.K.; Mandal, S.; Cumpstey, I.; Larumbe, A.; Salameh, B.A.; et al. Galectin-3-binding glycomimetics that strongly reduce bleomycin-induced lung fibrosis and modulate intracellular glycan recognition. ChemBioChem 2016, 17, 1759–1770. [Google Scholar] [CrossRef]
- The Galectin Pharmacology List. Available online: https://galecto.com/ (accessed on 17 August 2018).
- Than, N.G.; Romero, R.; Goodman, M.; Weckle, A.; Xing, J.; Dong, Z.; Xu, Y.; Tarquini, F.; Szilagyi, A.; Gal, P.; et al. A primate subfamily of galectins expressed at the maternal-fetal interface that promote immune cell death. Proc. Natl. Acad. Sci. USA 2009, 106, 9731–9736. [Google Scholar] [CrossRef]
- Than, N.G.; Romero, R.; Xu, Y.; Erez, O.; Xu, Z.; Bhatti, G.; Leavitt, R.; Chung, T.H.; El-Azzamy, H.; LaJeunesse, C.; et al. Evolutionary origins of the placental expression of chromosome 19 cluster galectins, and their complex dysregulation in preeclampsia. Placenta 2014, 35, 855–865. [Google Scholar] [CrossRef]
- Su, E.U.; Bi, S.; Kane, L.P. Galectin-9 regulates T helper cell function independently of Tim-3. Glycobiology 2011, 21, 1258–1265. [Google Scholar] [CrossRef]
- Ackerman, S.J.; Liu, L.; Kwatia, M.A.; Savage, M.P.; Leonidas, D.D.; Swaminathan, G.J.; Acharya, K.R. Charcot-Leyden crystal protein (galectin-10) is not a dual function galectin with lysophospholipase activity but binds a lysophospholipase inhibitor in a novel structural fashion. J. Biol. Chem. 2002, 277, 14859–14868. [Google Scholar] [CrossRef] [PubMed]
- Su, J. A brief history of Charcot-Leyden Crystal Protein/Galectin-10 research. Molecules 2018, 23, 2931. [Google Scholar] [CrossRef] [PubMed]
- Than, N.G.; Romero, R.; Kim, C.J.; McGowen, M.R.; Papp, Z.; Wildman, D.E. Galectins: Guardians of eutherian pregnancy at the maternal-fetal interface. Trends Endocrinol. Metab. 2012, 23, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Bohn, H.; Kraus, W.; Winckler, W. Purification and characterization of two new soluble placental tissue proteins (PP13 and PP17). Oncodev. Biol. Med. 1983, 4, 343–350. [Google Scholar] [PubMed]
- Sammar, M.; Nisamblatt, S.; Gonen, R.; Huppertz, B.; Gizurarson, S.; Osol, G.; Meiri, H. The role of the carbohydrate recognition domain of placental protein 13 (PP13) in pregnancy evaluated with recombinant PP13 and the DelT221 PP13 variant. PLoS ONE 2014, 9, e102832. [Google Scholar] [CrossRef]
- Than, N.G.; Romero, R.; Balogh, A.; Karpati, E.; Mastrolia, S.A.; Staretz-Chacham, O.; Hahn, S.; Erez, O.; Papp, Z.; Kim, C.J. Galectins: Double-edged swords in the cross-roads of pregnancy complications and female reproductive tract inflammation and neoplasia. J. Pathol. Transl. Med. 2015, 49, 181–208. [Google Scholar] [CrossRef]
- Yang, R.Y.; Rabinovich, G.A.; Liu, F.T. Galectins: Structure, function and therapeutic potential. Expert Rev. Mol. Med. 2008, 10, e17. [Google Scholar] [CrossRef] [PubMed]
- Burgess, A.; Johnson, T.S.; Simanek, A.; Bell, T.; Founds, S. Maternal ABO blood type and factors associated with preeclampsia subtype. Biol. Res. Nurs. 2019, 21, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Huppertz, B.; Sammar, M.; Chefetz, I.; Neumaier-Wagner, P.; Bartz, C.; Meiri, H. Longitudinal determination of serum placental protein 13 during development of preeclampsia. Fetal Diagn. Ther. 2008, 24, 230–236. [Google Scholar] [CrossRef] [PubMed]
- Kliman, H.J.; Sammar, M.; Grimpel, Y.I.; Lynch, S.K.; Milano, K.M.; Pick, E.; Bejar, J.; Arad, A.; Lee, J.J.; Meiri, H.; et al. Placental protein 13 and decidual zones of necrosis: An immunologic diversion that may be linked to preeclampsia. Reprod. Sci. 2012, 19, 16–30. [Google Scholar] [CrossRef] [PubMed]
- Balogh, A.; Pozsgay, J.; Matkó, J.; Dong, Z.; Kim, C.J.; Várkonyi, T.; Sammar, M.; Rigó, J., Jr.; Meiri, H.; Romero, R.; et al. Placental protein 13 (PP13/galectin-13) undergoes lipid raft-associated subcellular redistribution in the syncytiotrophoblast in preterm preeclampsia and HELLP syndrome. Am. J. Obstet. Gynecol. 2011, 205, 156.e1–156.e14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Than, N.G.; Abdul Rahman, O.; Magenheim, R.; Nagy, B.; Fule, T.; Hargitai, B.; Sammar, M.; Hupuczi, P.; Tarca, A.L.; Szabo, G.; et al. Placental protein 13 (galectin-13) has decreased placental expression but increased shedding and maternal serum concentrations in patients presenting with preterm pre-eclampsia and HELLP syndrome. Virchows Arch. 2008, 453, 387–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Than, N.G.; Sumegi, B.; Than, G.N.; Berente, Z.; Bohn, H. Isolation and sequence analysis of a cDNA encoding human placental tissue protein 13 (PP13), a new lysophospholipase, homologue of human eosinophil Charcot-Leyden Crystal protein. Placenta 1999, 20, 703–710. [Google Scholar] [CrossRef] [PubMed]
- Orendi, K.; Gauster, M.; Moser, G.; Meiri, H.; Huppertz, B. The choriocarcinoma cell line BeWo: Syncytial fusion and expression of syncytium-specific proteins. Reproduction 2010, 140, 759–766. [Google Scholar] [CrossRef] [PubMed]
- Kliman, H.J.; Nestler, J.E.; Sermasi, E.; Sanger, J.M.; Strauss, J.F., III. Purification, characterization, and in vitro differentiation of cytotrophoblasts from human term placentae. Endocrinology 1986, 118, 1567–1582. [Google Scholar] [CrossRef] [PubMed]
- Orendi, K.; Gauster, M.; Moser, G.; Meiri, H.; Huppertz, B. Effects of vitamins C and E, acetylsalicylic acid and heparin on fusion, beta-hCG and PP13 expression in BeWo cells. Placenta 2010, 31, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Meiri, H.; PP13. PP13. ELISA KIT for IVD of pregnancy complications. IDABC―EUDAMED: European Database on Medical Devices. Available online: http://ec.europa.eu/idabc/en/document/2256/5637.html. (accessed on 31 August 2008).
- Madar-Shapiro, L.; Karady, I.; Trahtenherts, A.; Syngelaki, A.; Akolekar, R.; Poon, L.; Cohen, R.; Sharabi-Nov, A.; Huppertz, B.; Sammar, M.; et al. Predicting the risk to develop preeclampsia in the first trimester combining promoter variant-98A/C of LGALS13 (Placental Protein 13), black ethnicity, previous preeclampsia, obesity, and maternal age. Fetal Diagn. Ther. 2018, 43, 250–265. [Google Scholar] [CrossRef]
- Sammar, M.; Dragovic, R.; Meiri, H.; Vatish, M.; Sharabi-Nov, A.; Sargent, I.; Redman, C.; Tannetta, D. Reduced placental protein 13 (PP13) in placental derived syncytiotrophoblast extracellular vesicles in preeclampsia—A novel tool to study the impaired cargo transmission of the placenta to the maternal organs. Placenta 2018, 66, 17–25. [Google Scholar] [CrossRef]
- Hughes, R.C. Secretion of the galectin family of mammalian carbohydrate-binding proteins. Biochim. Biophys. Acta 1999, 1473, 172–185. [Google Scholar] [CrossRef]
- Ideo, H.; Hoshi, I.; Yamashita, K.; Sakamoto, M. Phosphorylation and externalization of galectin-4 is controlled by Src family kinases. Glycobiology 2013, 23, 1452–1462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burger, O.; Pick, E.; Zwickel, J.; Kliman, M.; Meiri, H.; Slotky, R.; Mandel, S.; Rabinovitch, L.; Paltieli, Y.; Admon, A.; et al. Placental protein 13 (PP-13): Effects on cultured trophoblasts, and its detection in human body fluids in normal and pathological pregnancies. Placenta 2004, 25, 608–622. [Google Scholar] [CrossRef] [PubMed]
- Furtak, V.; Hatcher, F.; Ochieng, J. Galectin-3 mediates the endocytosis of beta-1 integrins by breast carcinoma cells. Biochem. Biophys. Res. Commun. 2001, 289, 845–850. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.J. Pre-eclampsia. Lancet 2000, 356, 1260–1265. [Google Scholar] [CrossRef]
- Roberts, J.M.; Cooper, H. Pathogenesis and genetics of pre-eclampsia. Lancet 2001, 357, 53–56. [Google Scholar] [CrossRef]
- Redman, C.W.; Sargent, I. Latest advances in understanding preeclampsia. Science 2005, 308, 1592–1594. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Maternal Mortality: To Improve Maternal Health, Barriers that Limit Access to Quality Maternal Health Services Must be Identified and Addressed at All Levels of the Health System: Fact Sheet. World Health Organization, 2014. Available online: https://apps.who.int/iris/handle/10665/112318 (accessed on 9 June 2019).
- Meekins, J.W.; Pijnenborg, R.; Hanssens, M.; McFadyen, I.R.; van Asshe, A. A study of placental bed spiral arteries and trophoblast invasion in normal and severe pre-eclamptic pregnancies. Br. J. Obstet. Gynaecol. 1994, 101, 669–674. [Google Scholar] [CrossRef]
- Huppertz, B. Placental origins of preeclampsia: Challenging the current hypothesis. Hypertension 2008, 51, 970–975. [Google Scholar] [CrossRef]
- The Preeclampsia Foundation. Available online: https://www.preeclampsia.org/ (accessed on 9 June 2019).
- Cetin, I.; Huppertz, B.; Burton, G.; Cuckle, H.; Gonen, R.; Lapaire, O.; Mandia, L.; Nicolaides, K.; Redman, C.; Soothill, P.; et al. Pregenesys pre-eclampsia markers consensus meeting: What do we require from markers, risk assessment and model systems to tailor preventive strategies? Placenta 2011, 32, S4–S16. [Google Scholar] [CrossRef]
- Sammar, M.; Nisemblat, S.; Fleischfarb, Z.; Golan, A.; Sadan, O.; Meiri, H.; Huppertz, B.; Gonen, R. Placenta-bound and body fluid PP13 and its mRNA in normal pregnancy compared to preeclampsia, HELLP and preterm delivery. Placenta 2011, 32, S30–S36. [Google Scholar] [CrossRef] [PubMed]
- Huppertz, B.; Meiri, H.; Gizurarson, S.; Osol, G.; Sammar, M. Placental protein 13 (PP13): A new biological target shifting individualized risk assessment to personalized drug design combating pre-eclampsia. Hum. Reprod. Update 2013, 19, 391–405. [Google Scholar] [CrossRef]
- Meiri, H.; Sammar, M.; Herzog, A.; Grimpel, Y.I.; Fihaman, G.; Cohen, A.; Kivity, V.; Sharabi-Nov, A.; Gonen, R. Prediction of preeclampsia by placental protein 13 and background risk factors and its prevention by aspirin. J. Perinat. Med. 2014, 42, 591–601. [Google Scholar] [CrossRef] [PubMed]
- Maymon, R.; Trahtenherts, A.; Svirsky, R.; Melcer, Y.; Madar-Shapiro, L.; Klog, E.; Meiri, H.; Cuckle, H. Developing a new algorithm for first and second trimester preeclampsia screening in twin pregnancies. Hypertens Pregnancy 2017, 36, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Nicolaides, K.H.; Bindra, R.; Turan, O.M.; Chefetz, I.; Sammar, M.; Meiri, H.; Tal, J.; Cuckle, H.S. A novel approach to first-trimester screening for early pre-eclampsia combining serum PP-13 and Doppler ultrasound. Ultrasound Obstet. Gynecol. 2006, 27, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Spencer, K.; Cowans, N.J.; Chefetz, I.; Tal, J.; Meiri, H. First-trimester maternal serum PP-13, PAPP-A and second-trimester uterine artery Doppler pulsatility index as markers of pre-eclampsia. Ultrasound Obstet. Gynecol. 2007, 29, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Spencer, K.; Cowans, N.J.; Chefetz, I.; Tal, J.; Kuhnreich, I.; Meiri, H. Second-trimester uterine artery Doppler pulsatility index and maternal serum PP13 as markers of pre-eclampsia. Prenat. Diagn. 2007, 27, 258–263. [Google Scholar] [CrossRef]
- Chafetz, I.; Kuhnreich, I.; Sammar, M.; Tal, Y.; Gibor, Y.; Meiri, H.; Cuckle, H.; Wolf, M. First-trimester placental protein 13 screening for preeclampsia and intrauterine growth restriction. Am. J. Obstet. Gynecol. 2007, 197, e1–e7. [Google Scholar] [CrossRef] [PubMed]
- Cowans, N.J.; Spencer, K.; Meiri, H. First-trimester maternal placental protein 13 levels in pregnancies resulting in adverse outcomes. Prenat. Diagn. 2008, 28, 121–125. [Google Scholar] [CrossRef]
- Romero, R.; Kusanovic, J.P.; Than, N.G.; Erez, O.; Gotsch, F.; Espinoza, J.; Edwin, S.; Chefetz, I.; Gomez, R.; Nien, J.K.; et al. First-trimester maternal serum PP13 in the risk assessment for preeclampsia. Am. J. Obstet. Gynecol. 2008, 199, 122.e1–122.e11. [Google Scholar] [CrossRef] [Green Version]
- Gonen, R.; Shahar, R.; Grimpel, Y.I.; Chefetz, I.; Sammar, M.; Meiri, H.; Gibor, Y. Placental protein 13 as an early marker for pre-eclampsia: A prospective longitudinal study. Br. J. Obstet. Gynaecol. 2008, 115, 1465–1472. [Google Scholar] [CrossRef] [PubMed]
- Khalil, A.; Cowans, N.J.; Spencer, K.; Goichman, S.; Meiri, H.; Harrington, K. First trimester maternal serum placental protein 13 for the prediction of pre-eclampsia in women with a priori high risk. Prenat. Diagn. 2009, 29, 781–789. [Google Scholar] [CrossRef] [PubMed]
- Khalil, A.; Cowans, N.J.; Spencer, K.; Goichman, S.; Meiri, H.; Harrington, K. First-trimester markers for the prediction of pre-eclampsia in women with a-priori high risk. Ultrasound Obstet. Gynecol. 2010, 35, 671–679. [Google Scholar] [CrossRef] [PubMed]
- Meiri, H.; Huppertz, B.; Cetin, I. Development of early non-invasive markers and means for the diagnosis and progression monitoring of preeclampsia and tailoring putative therapies (project pregenesys 037244). Placenta 2011, 32, S1–S3. [Google Scholar] [CrossRef] [PubMed]
- Farina, A.; Bernabini, D.; Zucchini, C.; De Sanctis, P.; Quezada, M.S.; Mattioli, M.; Rizzo, N. Elevated maternal placental protein 13 serum levels at term of pregnancy in postpartum major hemorrhage (>1000 mLs). A prospective cohort study. Am. J. Reprod. Immunol. 2017, 78, e12702. [Google Scholar] [CrossRef] [PubMed]
- Shekizawa, A.; Purwosunu, Y.; Yoshimura, S.; Nakamura, M.; Shimizu, H. PP13 mRNA expression in trophoblasts from preeclamptic placentas. Reprod. Sci. 2009, 16, 408–413. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, H.; Sekizawa, A.; Purwosunu, Y.; Nakamura, M.; Farina, A.; Rizzo, N.; Okai, T. PP13 mRNA expression in the cellular component of maternal blood as a marker for preeclampsia. Prenat. Diagn. 2009, 29, 1231–1236. [Google Scholar] [CrossRef] [PubMed]
- Farina, A.; Zucchini, C.; Sekizawa, A.; Purwosunu, Y.; de Sanctis, P.; Santarsiero, G.; Rizzo, N.; Morano, D.; Okai, T. Performance of messenger RNAs circulating in maternal blood in the prediction of preeclampsia at 10–14 weeks. Am. J. Obstet. Gynecol. 2010, 203, 575.e1-7. [Google Scholar] [CrossRef]
- Grimpel, Y.I.; Kivity, V.; Cohen, A.; Meiri, H.; Sammar, M.; Gonen, R.; Huppertz, B. Effects of calcium, magnesium, low-dose aspirin and low-molecular-weight heparin on the release of PP13 from placental explants. Placenta 2011, 32, S55–S64. [Google Scholar] [CrossRef]
- Than, N.G.; Balogh, A.; Romero, R.; Kárpáti, E.; Erez, O.; Szilágyi, A.; Kovalszky, I.; Sammar, M.; Gizurarson, S.; Matkó, J.; et al. Placental Protein 13 (PP13)-A placental immunoregulatory galectin protecting pregnancy. Front. Immunol. 2014, 20, 348. [Google Scholar] [CrossRef]
- Gebhardt, S.; Bruiners, N.; Hillerman, R. A novel exonic variant (221delT) in the LGALS13 gene encoding placental protein 13 (PP13) is associated with preterm labour in a low risk population. J. Reprod. Immunol. 2009, 82, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Than, N.G.; Romero, R.; Hillermann, R.; Cozzi, V.; Nie, G.; Huppertz, B. Prediction of preeclampsia-a workshop report. Placenta 2008, 29, S83–S85. [Google Scholar] [CrossRef] [PubMed]
- Bruiners, N.; Bosman, M.; Postma, A.; Gebhardt, S.; Rebello, G.; Sammar, M.; Meiri, H.; Hillermann, R. Promoter variant-98A-C of the LGALS13 gene and pre-eclampsia. In Proceedings of the 8th World Congress of Prenatal Medicine and Fetal Development, Florence, Italy, 7 September 2007. [Google Scholar]
- Palmer, S.K.; Zamudio, S.; Coffin, C.; Parker, S.; Stamm, E.; Moore, L.G. Quantitative estimation of human uterine artery blood flow and pelvic blood flow redistribution in pregnancy. Obstet. Gynecol. 1992, 80, 1000–1006. [Google Scholar] [PubMed]
- Thornburg, K.L.; Jacobson, S.L.; Giraud, G.D.; Morton, M.J. Hemodynamic changes in pregnancy. Semin. Perinatol. 2000, 24, 11–14. [Google Scholar] [CrossRef]
- Moser, G.; Windsperger, K.; Pollheimer, J.; de Sousa Lopes, S.C.; Huppertz, B. Human trophoblast invasion: New and unexpected routes and functions. Histochem. Cell Biol. 2018, 150, 361–370. [Google Scholar] [CrossRef]
- Moser, G.; Drewlo, S.; Huppertz, B.; Armant, D.R. Trophoblast retrieval and isolation from the cervix: Origins of cervical trophoblasts and their potential value for risk assessment of ongoing pregnancies. Hum. Reprod. Update 2018, 24, 484–496. [Google Scholar] [CrossRef]
- Moser, G.; Huppertz, B. Implantation and extravillous trophoblast invasion: From rare archival specimens to modern biobanking. Placenta 2017, 56, 19–26. [Google Scholar] [CrossRef]
- Kaufmann, P.; Black, S.; Huppertz, B. Endovascular trophoblast invasion: Implications for the pathogenesis of intrauterine growth retardation and preeclampsia. Biol. Reprod. 2003, 69, 1–7. [Google Scholar] [CrossRef]
- Gokina, N.I.; Mandala, M.; Osol, G. Induction of localized differences in rat uterine radial artery behavior and structure during gestation. Am. J. Obstet. Gynecol. 2003, 189, 1489–1493. [Google Scholar] [CrossRef]
- Pijnenborg, R.; Vercruysse, L.; Hanssens, M. The uterine spiral arteries in human pregnancy: Facts and controversies. Placenta 2006, 27, 939–958. [Google Scholar] [CrossRef]
- Pijnenborg, R.; Vercruysse, L.; Brosens, I. Deep placentation. Best Pract. Res. Clin. Obstet. Gynaecol. 2011, 25, 273–285. [Google Scholar] [CrossRef] [PubMed]
- Moll, W. Structure adaptation and blood flow control in the uterine arterial system after hemochorial placentation. Eur. J. Obstet. Gynecol. Reprod. Biol. 2003, 110, S19–S27. [Google Scholar] [CrossRef]
- Burton, G.J.; Woods, A.W.; Jauniaux, E.; Kingdom, J.C. Rheological and physiological consequences of conversion of the maternal spiral arteries for uteroplacental blood flow during human pregnancy. Placenta 2009, 30, 473–482. [Google Scholar] [CrossRef] [PubMed]
- Browne, V.A.; Julian, C.G.; Toledo-Jaldin, L.; Cioffi-Ragan, D.; Vargas, E.; Moore, L.G. Uterine artery blood flow, fetal hypoxia and fetal growth. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20140068. [Google Scholar] [CrossRef] [PubMed]
- Konje, J.C.; Kaufmann, P.; Bell, S.C.; Taylor, D.J. A longitudinal study of quantitative uterine blood flow with the use of color power angiography in appropriate for gestational age pregnancies. Am. J. Obstet. Gynecol. 2001, 185, 608–613. [Google Scholar] [CrossRef] [PubMed]
- Mandala, M.; Osol, G. Physiological remodelling of the maternal uterine circulation during pregnancy. Basic Clin. Pharmacol. Toxicol. 2012, 110, 12–18. [Google Scholar] [CrossRef] [PubMed]
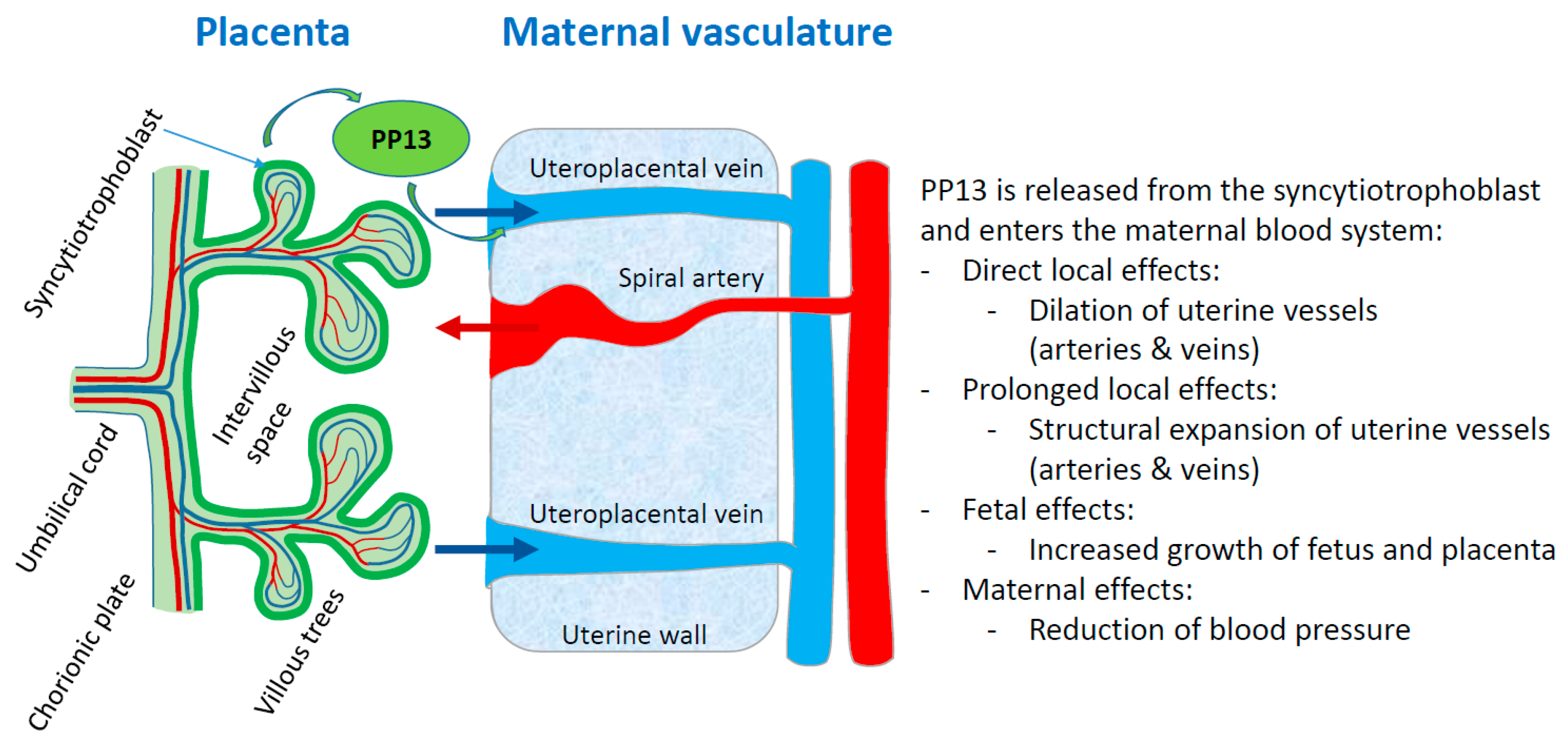
- Gizurarson, S.; Huppertz, B.; Osol, G.; Skarphedinsson, J.O.; Mandala, M.; Meiri, H. Effects of placental protein 13 on the cardiovascular system in gravid and non-gravid rodents. Fetal Diagn. Ther. 2013, 33, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Gizurarson, S.; Sigurdardottir, E.R.; Meiri, H.; Huppertz, B.; Sammar, M.; Sharabi-Nov, A.; Mandalá, M.; Osol, G. Placental protein 13 administration to pregnant rats lowers blood pressure and augments fetal growth and venous remodeling. Fetal Diagn. Ther. 2016, 39, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Drobnjak, T.; Gizurarson, S.; Gokina, N.I.; Meiri, H.; Mandalá, M.; Huppertz, B.; Osol, G. Placental protein 13 (PP13)-induced vasodilation of resistance arteries from pregnant and nonpregnant rats occurs via endothelial-signaling pathways. Hypertens. Pregnancy 2017, 36, 86–95. [Google Scholar] [CrossRef]
- Drobnjak, T.; Jónsdóttir, A.M.; Helgadóttir, H.; Runólfsdóttir, M.S.; Meiri, H.; Sammar, M.; Osol, G.; Mandalà, M.; Huppertz, B.; Gizurarson, S. Placental protein 13 (PP13) stimulates rat uterine vessels after slow subcutaneous administration. Int. J. Womens Health 2019, 11, 213–222. [Google Scholar] [CrossRef]
- Meiri, H.; Osol, G.; Cetin, I.; Gizurarson, S.; Huppertz, B. Personalized therapy against preeclampsia by replenishing placental protein 13 (PP13) targeted to patients with impaired PP13 molecule or function. Comput. Struct. Biotechnol. J. 2017, 15, 433–446. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; LaMarca, B.; Reckelhoff, J.F. A model of preeclampsia in rats: The reduced uterine perfusion pressure (RUPP) model. Am. J. Physiol. Heart Circ. Physiol. 2012, 303, H1–H8. [Google Scholar] [CrossRef] [PubMed]
- Collinot, H.; Marchiol, C.; Lagoutte, I.; Lager, F.; Siauve, N.; Autret, G.; Balvay, D.; Renault, G.; Salomon, L.J.; Vaiman, D. Preeclampsia induced by STOX1 overexpression in mice induces intrauterine growth restriction, abnormal ultrasonography and BOLD MRI signatures. J. Hypertens. 2018, 36, 1399–1406. [Google Scholar] [CrossRef] [PubMed]
- Makris, A.; Yeung, K.R.; Shirlene, M.; Lim, S.L.; Sunderland, N.; Heffernan, S.; Thompson, J.F.; Iliopoulos, J.; Killingsworth, M.C.; Yong, J.; et al. Placental growth factor reduces blood pressure in a uteroplacental ischemia model of preeclampsia in non-human primates. Hypertension 2016, 67, 1263–1272. [Google Scholar] [CrossRef] [PubMed]
- Grimes, P.S.; Bombay, K.; Lanes, A.; Walker, M.; Daniel, J.; Corsi, D.J. Potential biological therapies for severe preeclampsia: A systematic review and meta-analysis. BMC Pregnancy Childbirth 2019, 19, 163. [Google Scholar] [CrossRef]
- Gunnarsson, R.; Akerstorm, B.; Hansson, S.R.; Gram, M. Recombinant alpha-1-microglobulin: A potential treatment for preeclampsia. Drug Dis. Today 2017, 22, 736–743. [Google Scholar] [CrossRef] [PubMed]
- Chavatte-Palmer, P.; Tarrade, A. Placentation in different mammalian species. Ann. Endocrinol. 2016, 77, 67–74. [Google Scholar] [CrossRef]
- Enders, A.C.; Carter, A.M. The evolving placenta: Different developmental paths to a hemochorial relationship. Placenta 2012, 3, S92–S98. [Google Scholar] [CrossRef]
- Wildman, D.E.; Chen, C.; Erez, O.; Grossman, L.I.; Goodman, M.; Romero, R. Evolution of the mammalian placenta revealed by phylogenetic analysis. Proc. Natl. Acad. Sci. USA 2006, 103, 3203–3208. [Google Scholar] [CrossRef] [Green Version]
- Wildman, D.E.; Uddin, M.; Romero, R.; Gonzalez, J.M.; Than, N.G.; Murphy, J.; Hou, Z.C.; Fritz, J. Spontaneous abortion and preterm labor and delivery in nonhuman primates: Evidence from a captive colony of chimpanzees (Pan troglodytes). PLoS ONE 2011, 6, e24509. [Google Scholar] [CrossRef]
- Roberts, M.R.; Green, J.A.; Schulz, L.C. The evolution of the placenta. Reproduction 2016, 152, R179–R189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Searching for Orthologous Genes at NCBI. Available online: https://www.ncbi.nlm.nih.gov/kis/ortholog/29124/?scope=9526#genes-tab (accessed on 9 June 2019).
- Carter, A.M. Recent advances in understanding evolution of the placenta: Insights from transcriptomics. F1000Research 2018, 7. [Google Scholar] [CrossRef] [PubMed]
- Hou, Z.C.; Sterner, K.N.; Romero, R.; Than, N.G.; Gonzalez, J.M.; Weckle, A.; Xing, J.; Benirschke, K.; Goodman, M.; Wildman, D.E. Elephant transcriptome provides insights into the evolution of eutherian placentation. Genome Biol. Evol. 2012, 4, 713–725. [Google Scholar] [CrossRef] [PubMed]
- Pijnenborg, R.; Vercruysse, L.; Carter, A.M. Deep trophoblast invasion and spiral artery remodelling in the placental bed of the lowland gorilla. Placenta 2011, 32, 586–591. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Gal 10 Eosinophils | Gal 13 Placenta | Gal 14 Placenta | Gal 16 Placenta | Gal 17 Placenta | Gal 20 Placenta | Count | Invasion Level |
|---|---|---|---|---|---|---|---|---|
| Chimpanzee | 2 | 3+ | ||||||
| Orangutan | 10A | 17C | 4 | 2+ | ||||
| Baboon | 1 | 1+ | ||||||
| Human | 10A | 17A 17B | 5 (with 2 subtypes of Gal 17) | 4+ | ||||
| Gorilla | 1 | 3+ | ||||||
| Colobus | 1 | 1+ | ||||||
| Macaque | 17C | 4 | 2+ | |||||
| Marmoset | 10A 10B 10C | 4 (with 3 subtypes of Gal 10) | 2+ | |||||
| Sp. Monkeys | 10A 10B 10C | 4 (with 3 subtypes of Gal 10) | 2+ | |||||
| Total species # | 4 | 8 | 5 | 4 | 3 | 2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sammar, M.; Drobnjak, T.; Mandala, M.; Gizurarson, S.; Huppertz, B.; Meiri, H. Galectin 13 (PP13) Facilitates Remodeling and Structural Stabilization of Maternal Vessels during Pregnancy. Int. J. Mol. Sci. 2019, 20, 3192. https://doi.org/10.3390/ijms20133192
Sammar M, Drobnjak T, Mandala M, Gizurarson S, Huppertz B, Meiri H. Galectin 13 (PP13) Facilitates Remodeling and Structural Stabilization of Maternal Vessels during Pregnancy. International Journal of Molecular Sciences. 2019; 20(13):3192. https://doi.org/10.3390/ijms20133192
Chicago/Turabian StyleSammar, Marei, Tijana Drobnjak, Maurizio Mandala, Sveinbjörn Gizurarson, Berthold Huppertz, and Hamutal Meiri. 2019. "Galectin 13 (PP13) Facilitates Remodeling and Structural Stabilization of Maternal Vessels during Pregnancy" International Journal of Molecular Sciences 20, no. 13: 3192. https://doi.org/10.3390/ijms20133192
APA StyleSammar, M., Drobnjak, T., Mandala, M., Gizurarson, S., Huppertz, B., & Meiri, H. (2019). Galectin 13 (PP13) Facilitates Remodeling and Structural Stabilization of Maternal Vessels during Pregnancy. International Journal of Molecular Sciences, 20(13), 3192. https://doi.org/10.3390/ijms20133192