Seed Treatment with Trichoderma longibrachiatum T6 Promotes Wheat Seedling Growth under NaCl Stress Through Activating the Enzymatic and Nonenzymatic Antioxidant Defense Systems

Abstract
:1. Introduction
2. Results
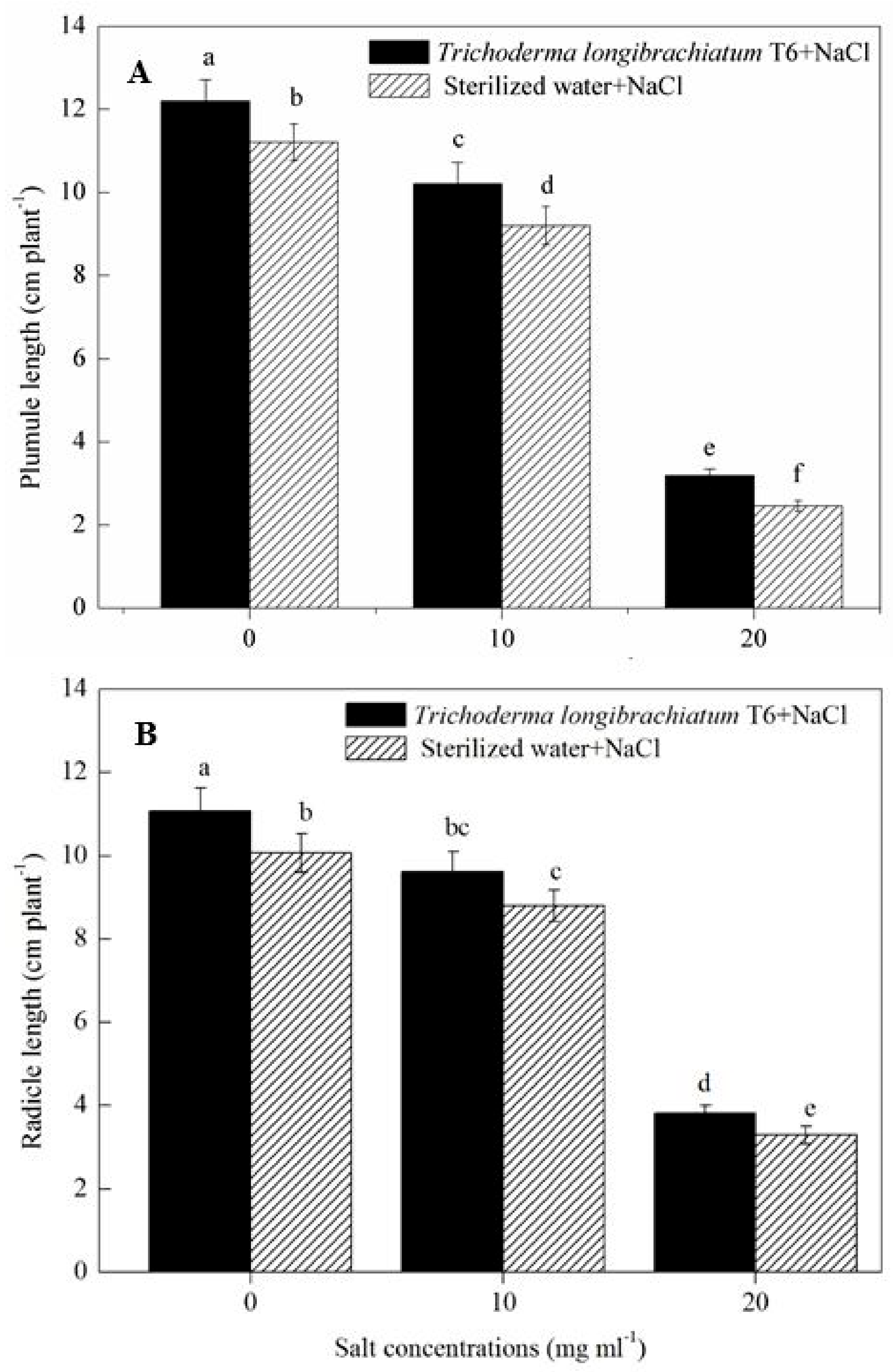
2.1. Effect of TL-6 on Wheat Seed Germination and Seedling Growth in Vitro
2.2. Effect of TL-6 on Wheat Seedling Growth in Greenhouse
2.3. Lipid Peroxides, H2O2 and Proline Content in Wheat Seedling
2.4. Ascorbate and Glutathione Contents in Wheat Seedling
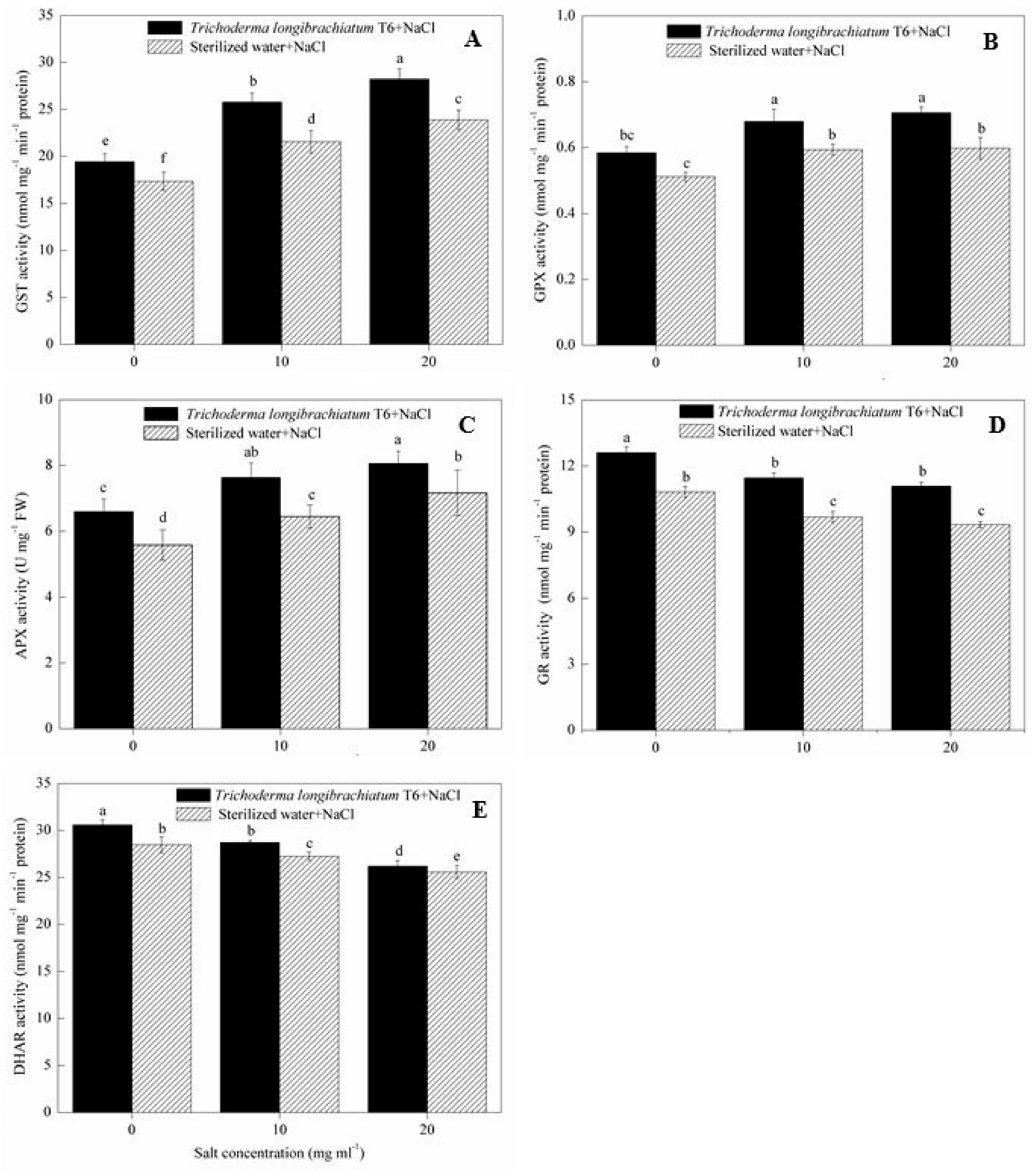
2.5. ROS Scavenging Enzymes Assay
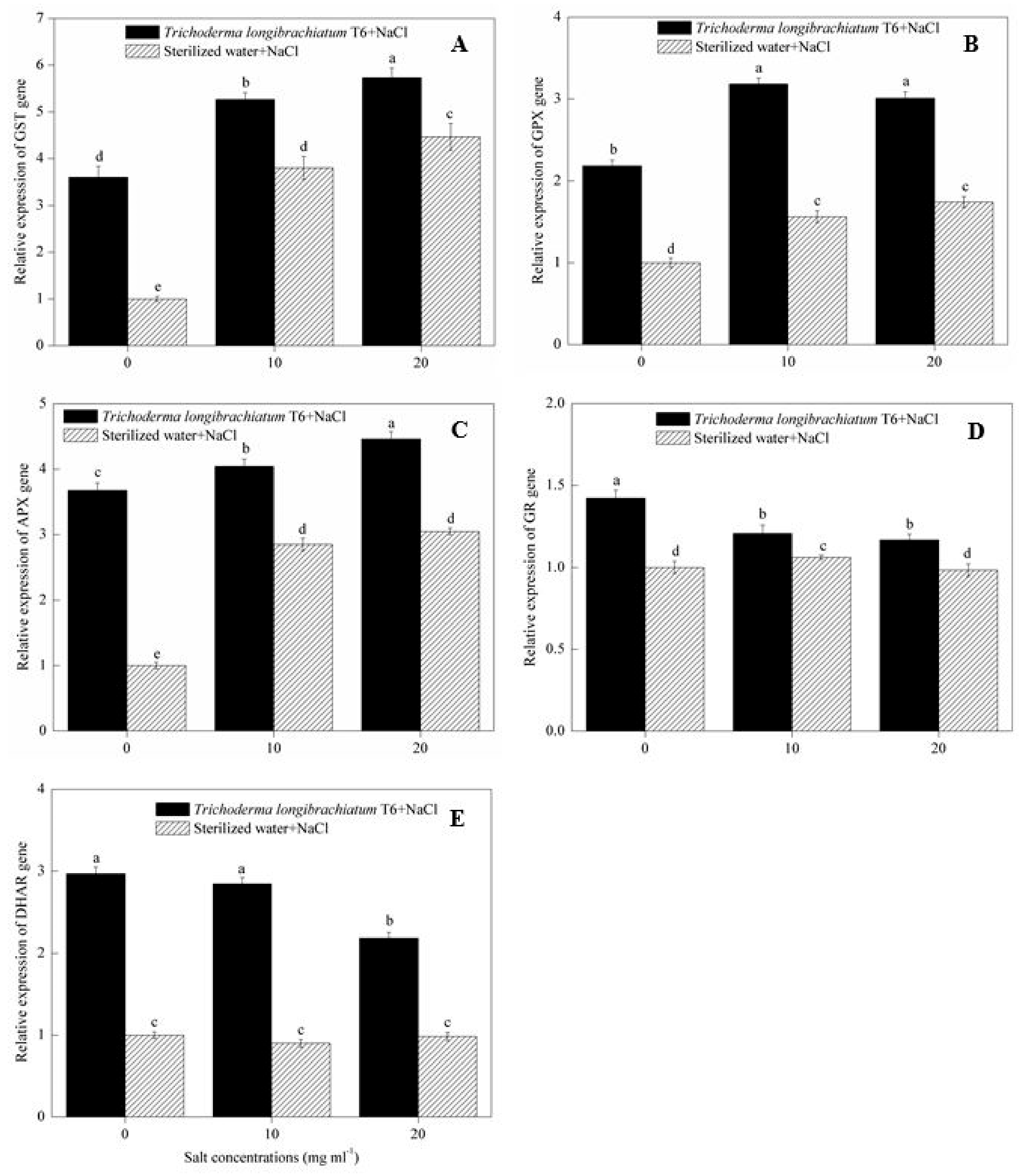
2.6. ROS Scavenging Enzyme Gene Expression
3. Discussion and Conclusions
4. Materials and Methods
4.1. Fungal Material
4.2. Seeds Treatment
4.3. Effect of TL-6 on Wheat Seed Germination under NaCl Stress
4.4. Effect of TL-6 on Wheat Seedling Growth under NaCl Stress in Greenhouse
4.5. Lipid Peroxides and Proline Contents Determination
4.6. H2O2 Assay in Wheat Seedling
4.7. Determination of Ascorbate and Glutathione
4.8. ROS Scavenging Enzymes Extraction and Determination
4.9. Total RNA Extraction and First Strand cDNA Synthesis
4.10. Quantitative Real-Time PCR (qRT-PCR) Analysis
4.11. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Mastouri, F.; Björkman, T.; Harman, G.E. Seed treatment with Trichoderma harzianum alleviates biotic, abiotic, and physiological stresses in germinating seeds and seedlings. Phytopathology 2010, 100, 1213–1221. [Google Scholar] [CrossRef] [PubMed]
- Contreras-Cornejo, H.A.; Macías-Rodríguez, L.; Alfaro-Cuevas, R.; López-Bucio, J. Trichoderma spp. improve growth of Arabidopsis seedlings under salt stress through enhanced root development, osmolite production, and Na+ elimination through root exudates. Mol. Plant Microbe Interact. 2014, 27, 503–514. [Google Scholar] [CrossRef] [PubMed]
- Hariadi, Y.; Marandon, K.; Tian, Y.; Jacobsen, S.E.; Shabala, S. Ionic and osmotic relations in quinoa (Chenopodium quinoa Willd.) plants grown at various salinity levels. J. Exp. Bot. 2011, 62, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Cabot, C.; Sibole, J.V.; Barcelo, J.; Poschenreider, C. Lessons from crop plants struggling with salinity. Plant Sci. 2014, 226, 2–13. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, P.M. Sodium (Na+) homeostasis and salt tolerance of plants. Environ. Exp. Bot. 2013, 92, 19–31. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Druzhinina, I.S.; Pan, X.Y.; Yuan, Z.L. Microbially mediated plant salt tolerance and microbiome-based solutions for saline agriculture. Biotechnol. Adv. 2016, 34, 1245–1259. [Google Scholar] [CrossRef] [PubMed]
- Harman, G.E. Overview of mechanisms and uses of Trichoderma spp. Phytopathology 2006, 96, 190–194. [Google Scholar] [CrossRef]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species: Opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2004, 2, 43–56. [Google Scholar] [CrossRef]
- Yedidia, I.; Srivastva, A.K.; Kapulnik, Y.; Chet, I. Effect of Trichoderma harzianum on microelement concentrations and increased growth of cucumber plants. Plant Soil 2001, 235, 235–242. [Google Scholar] [CrossRef]
- Gamalero, E.; Berta, G.; Massa, N.; Glick, B.R.; Lingua, G. Synergistic interactions between the ACC deaminase-producing bacterium Pseudomonas putida UW4 and the AM fungus Gigaspora rosea positively affect cucumber plant growth. FEMS Microbiol. Ecol. 2008, 64, 459–467. [Google Scholar] [CrossRef] [PubMed]
- Shabala, S.; Cuin, T.A. Potassium transport and plant salt tolerance. Physiol. Plant 2008, 133, 651–669. [Google Scholar] [CrossRef] [PubMed]
- De Palma, M.; D’Agostino, N.; Proietti, S.; Bertini, L.; Lorito, M.; Caruso, C.; Ruocco, M.; Chiusano, M.L.N.; Tucci, M. Suppression subtractive hybridization analysis provides new insights into the tomato (Solanum lycopersicum L.) response to the plant probiotic microorganism Trichoderma longibrachiatum MK1. J. Plant Physiol. 2016, 190, 79–94. [Google Scholar] [CrossRef] [PubMed]
- Brotman, Y.; Landau, U.; Cuadros-Inostroza, Á.; Tohge, T.; Fernie, A.R.; Chet, I.; Viterbo, A.; Willmitzer, L. Trichoderma-plant root colonization: Escaping early plant defense responses and activation of the antioxidant machinery for saline stress tolerance. PLoS Pathog. 2013, 9, e1003221. [Google Scholar] [CrossRef]
- Türkan, I.; Demiral, T. Recent developments in understanding salinity tolerance. Environ. Exp. Bot. 2009, 67, 2–9. [Google Scholar] [CrossRef]
- Zhang, S.W.; Gan, Y.T.; Xu, B.L. Efficacy of Trichoderma longibrachiatum in the control of Heterodera avenae. BioControl 2014, 59, 319–331. [Google Scholar] [CrossRef]
- Zhang, S.W.; Gan, Y.T.; Xu, B.L. Application of plant-growth-promoting fungi Trichoderma longibrachiatum T6 enhances tolerance of wheat to salt stress through improvement of antioxidative defense system and gene expression. Front. Plant Sci. 2016, 7, 1405. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.J.; Li, Y.Y.; Yu, C.M.; Wang, Y.; Li, X.M.; Li, N.; Chen, Q.; Bu, N. Alleviation of exogenous oligochitosan on wheat seedlings growth under salt stress. Protoplasma 2012, 249, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Mishra1, N.; Khan, S.S.; Sundari, S.K. Native isolate of Trichoderma: A biocontrol agent with unique stress tolerance properties. World J. Microbiol. Biotechnol. 2016, 32, 130. [Google Scholar] [CrossRef] [PubMed]
- Azooz, M.M.; Youssef, A.M.; Ahmad, P. Evaluation of salicylic acid (SA) application on growth, osmotic solutes and antioxidant enzyme activities on broad bean seedlings grown under diluted seawater. Int. J. Plant Physiol. Biochem. 2011, 3, 253–264. [Google Scholar]
- Rasool, S.; Ahmad, A.; Siddiqi, T.O.; Ahmad, P. Changes in growth, lipid peroxidation and some key antioxidant enzymes in chickpea genotypes under salt stress. Acta Physiol. Plant 2013, 35, 1039–1050. [Google Scholar] [CrossRef]
- Ahmad, P.; Ozturk, M.; Sharma, S.; Gucel, S. Effect of sodium carbonate-induced salinity-alkalinity on some key osmoprotectants, protein profile, antioxidant enzymes, and lipid peroxidation in two mulberry (Morus alba L.) cultivars. J. Plant Interact. 2014, 9, 460–467. [Google Scholar] [CrossRef]
- Contreras-Cornejo, H.A.; Macías-Rodríguez, L.; Cortés-Penagos, C.; López-Bucio, J. Trichoderma virens, a plant beneficial fungus, enhances biomass production and promotes lateral root growth through an auxin-dependent mechanism in Arabidopsis. Plant Physiol. 2009, 149, 1579–1592. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, P.; Jaleel, C.A.; Salem, M.A.; Nabi, G.; Sharma, S. Roles of enzymatic and non-enzymatic antioxidants in plants during abiotic stress. Crit. Rev. Biotechnol. 2010, 30, 161–175. [Google Scholar] [CrossRef] [PubMed]
- Shan, C.; Liang, Z. Jasmonic acid regulates ascorbate and glutathione metabolism in Agropyron cristatum leaves under water stress. Plant Sci. 2010, 178, 130–139. [Google Scholar] [CrossRef]
- Liu, Y.; Yuan, Y.; Liu, Y.; Fu, J.; Zheng, J.; Wang, G. Gene families of maize glutathione ascorbate redox cycle respond differently to abiotic stresses. J. Plant Physiol. 2012, 169, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Mastouri, F.; Bjorkman, T.; Harman, G.E. Trichoderma harzianum enhances antioxidant defense of tomato seedlings and resistance to water deficit. Mol. Plant Microbe Interact. 2012, 25, 1264–1271. [Google Scholar] [CrossRef]
- Aggarwal, A.; Kadian, N.; Neetu, K.; Tanwar, A.; Gupta, K.K. Arbuscular mycorrhizal symbiosis and alleviation of salinity stress. J. Appl. Nat. Sci. 2012, 4, 144–155. [Google Scholar] [CrossRef] [Green Version]
- Jogaiah, S.; Govind, S.R.; Tran, L.S. Systems biology-based approaches toward understanding drought tolerance in food crops. Crit. Rev. Biotechnol. 2013, 33, 23–39. [Google Scholar] [CrossRef]
- Li, Y. Physiological responses of tomato seedlings (Lycopersicon esculentum) to salt stress. Mod. Appl. Sci. 2009, 3, 171–176. [Google Scholar] [CrossRef]
- Saleem, M.; Ashraf, M.; Akram, N.A. Salt (NaCl)-induced modulation in some key physio-biochemical attributes in okra (Abelmoschus esculentus L.). J. Agron. Crop Sci. 2011, 197, 202–213. [Google Scholar] [CrossRef]
- Rawat, N.; Himabindu, K.; Neeraja, C.N.; Nair, S.; Bentur, J.S. Suppressive subtraction hybridization reveals that rice gall midge attack elicits plant-pathogen-like responses in rice. Plant Physiol. Biochem. 2013, 63, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Li, M.F.; Sun, B.G.; Xiao, Z.Z.; Sun, L. First characterization of a teleost Epstein-Barrvirus-induced gene 3 (EBI3) reveals a regulatory effect of EBI3 on the innate immune response of peripheral blood leukocytes. Dev. Comp. Immunol. 2013, 41, 514–522. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.H.; He, W.L.; Guo, J.K.; Chang, X.X.; Su, P.X.; Zhang, L.X. Increased sensitivity to salt stress in an ascorbate-deficient Arabidopsis mutant. J. Exp. Bot. 2005, 56, 3041–3049. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Fujita, M. Selenium pretreatment upregulates the antioxidant defense and methylglyoxal detoxification system and confers enhanced tolerance to drought stress in rapeseed seedlings. Biol. Trace Elem. Res. 2011, 143, 1758–1776. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.M.; Saltveit, M.E. Chilling tolerance of maize, cucumber and rice seedling leaves and roots are differentially affected by salicylic acid. Physiol. Plant 2002, 115, 571–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.C.; Liu, Z.M.; Cui, J.X.; Ding, J.G.; Xia, X.J.; Liu, D.L.; Yu, J.Q. Alleviation of chilling-induced oxidative damage by salicylic acid pretreatment and related gene expression in eggplant seedlings. Plant Growth Regul. 2011, 65, 101–108. [Google Scholar] [CrossRef]
- Duan, M.; Feng, H.L.; Wang, L.Y.; Li, D.; Meng, Q.W. Overexpression of thylakoidal ascorbate peroxidase shows enhanced resistance to chilling stress in tomato. J. Plant Physiol. 2012, 169, 867–877. [Google Scholar] [CrossRef] [PubMed]
- Sultana, S.; Khew, C.Y.; Morshed, M.M.; Namasivayam, P.; Napis, S.; Ho, C.L. Overexpression of monodehydroascorbate reductase from a mangrove plant (AeMDHAR) confers salt tolerance on rice. J. Plant Physiol. 2012, 169, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Hargrove, M.S. Nitric oxide in plants: The roles of ascorbate and hemoglobin. PLoS ONE 2013, 8, e82611. [Google Scholar] [CrossRef]
- Domingos, P.; Prado, A.M.; Wong, A.; Gehring, C.; Feijo, J.A. Nitric oxide: A multitasked signaling gas in plants. Mol. Plant 2015, 8, 506–520. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, P.; Latef, A.A.; Hashem, A.; Abd Allah, E.; Gucel, S.; Tran, L.S. Nitric oxide mitigates salt stress by regulating levels of osmolytes and antioxidant enzymes in chickpea. Front. Plant Sci. 2016, 7, 347. [Google Scholar] [CrossRef] [PubMed]
- Donia, M.; Maksimovic-Ivanic, D.; Mijatovic, S.; Mojic, M.; Miljkovic, D.; Timotijevic, G.; Fagone, P.; Caponnetto, S.; Al-Abed, Y.; McCubrey, J.; et al. In vitro and in vivo anticancer action of Saquinavir-NO, a novel nitric oxide-derivative of the protease inhibitor saquinavir, on hormone resistant prostate cancer cells. Cell Cycle 2011, 10, 492–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moloney, J.N.; Cotter, T.G. ROS signalling in the biology of cancer. Semin. Cell Dev. Biol. 2018, 80, 50–64. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.W.; Gan, Y.T.; Xue, Y.Y.; Xu, B.L. The parasitic and lethal effects of Trichoderma longibrachiatum against Heterodera avenae. Biol. Control 2014, 72, 1–8. [Google Scholar] [CrossRef]
- Niu, X.J.; Mi, L.N.; Li, Y.D.; Wei, A.S.; Yang, Z.Q.; Wu, J.D.; Zhang, D.; Song, X.F. Physiological and biochemical responses of rice seeds to phosphine exposure during germination. Chemosphere 2013, 93, 2239–2244. [Google Scholar] [CrossRef]
- Demiral, T.; Turkan, I. Comparative lipid peroxidation, antioxidant defense systems and proline content in roots of two rice cultivars differing in salt tolerance. Environ. Exp. Bot. 2005, 53, 247–257. [Google Scholar] [CrossRef]
- Tian, X.Y.; He, M.R.; Wang, Z.L.; Zhang, J.W.; Song, Y.L.; He, Z.L.; Dong, Y.J. Application of nitric oxide and calcium nitrate enhances tolerance of wheat seedlings to salt stress. Plant Growth Regul. 2015, 77, 343–356. [Google Scholar] [CrossRef]
- Bates, L.; Waldren, P.P.; Teare, J.D. Rapid determination of free proline of water stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Jena, S.; Choudhuri, M.A. Glycolate metabolism of three submerged aquatic angiosperms during aging. Aquat. Bot. 1981, 12, 345–354. [Google Scholar] [CrossRef]
- Foyer, C.H.; Rowell, J.; Walker, D. Measurements of the ascorbate content of spinach leaf protoplasts during illumination. Planta 1983, 157, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Fujita, M. Exogenous sodium nitroprusside alleviates arsenic-induced oxidative stress in wheat (Triticum aestivum L.) seedlings by enhancing antioxidant defense and glyoxalase system. Ecotoxicology 2013, 22, 584–596. [Google Scholar] [CrossRef] [PubMed]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach-chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- He, Z.Q.; Zou, Z.R.; He, C.X. Effect of AMF on GSH-Px activity and cell membrane osmosis of tomato. J. Northwest Sci. Tech. Univ. Agri. For. (Nat. Sci. Ed.) 2006, 34, 53–57. [Google Scholar]
- Elia, A.C.; Galarini, R.; Taticchi, M.I.; Dorr, A.J.M.; Mantilacci, L. Antioxidant responses and bioaccumulation in Ictalurus melas under mercury exposure. Ecotoxicol. Environ. Saf. 2003, 55, 162–167. [Google Scholar] [CrossRef]
- Foyer, C.H.; Halliwell, B. The presence of glutathione and glutathione reductase in chloroplasts: A proposed role in ascorbic acid metabolism. Planta 1976, 133, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Adriano, S.; Angelo, C.T.; Bartolomeo, D.; Cristos, X. Influence of water deficit and rewatering on the components of the ascorbate-glutathione cycle in four interspecific Prunus hybrids. Plant Sci. 2005, 169, 403–412. [Google Scholar]
- Qiu, Z.B.; Guo, J.L.; Zhu, A.J.; Zhang, L.; Zhang, M.M. Exogenous jasmonic acid can enhance tolerance of wheat seedlings to salt stress. Ecotoxicol. Environ. Saf. 2014, 104, 202–208. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Treatment | Salt Concentrations (mg mL−1) | Germination Rate (%) | Germination Potential (%) | Germination Index (%) |
|---|---|---|---|---|
| Trichoderma longibrachiatum T6 | 0 | 96.0 ± 1.2a | 89.0 ± 1.2a | 74.2 ± 0.7a |
| 10 | 83.3 ± 1.4b | 65.0 ± 1.2c | 52.2 ± 1.2c | |
| 20 | 68.3 ± 1.7d | 51.3 ± 1.4e | 40.2 ± 0.7e | |
| Sterile water | 0 | 93.3 ± 1.2a | 82.0 ± 1.2b | 68.1 ± 1.2b |
| 10 | 78.0 ± 2.3c | 61.3 ± 0.8d | 44.9 ± 1.2d | |
| 20 | 63.3 ± 1.2e | 47.0 ± 1.2f | 38.0 ± 1.7e |
| Treatment | Salt Concentration (mg mL−1) | Shoot Length (cm plant−1) | Root Length (cm plant−1) | Fresh Weight (g plant−1) | Dry Weight (g plant−1) |
|---|---|---|---|---|---|
| Trichoderma longibrachiatum T6 | 0 | 31.24 ± 0.7a | 24.79 ± 0.6a | 0.47 ± 0.01a | 0.14 ± 0.02a |
| 10 | 31.05 ± 0.7a | 22.55 ± 0.5b | 0.43 ± 0.01ab | 0.11 ± 0.01abc | |
| 20 | 28.67 ± 0.5b | 21.58 ± 0.4c | 0.34 ± 0.02c | 0.09 ± 0.01bc | |
| Sterile water | 0 | 28.55 ± 0.6b | 22.24 ± 0.3b | 0.41 ± 0.02b | 0.12 ± 0.01ab |
| 10 | 27.43 ± 0.6bc | 20.21 ± 0.5c | 0.35 ± 0.02c | 0.09 ± 0.02bc | |
| 20 | 24.22 ± 0.4c | 18.65 ± 0.4d | 0.29 ± 0.02d | 0.07 ± 0.01c |
| Treatment | Salt Concentration (mg mL−1) | MDA Content (µmol g−1 FW) | H2O2 Content (µmol g−1 FW) | Proline Content (µmol g−1 FW) |
|---|---|---|---|---|
| Trichoderma longibrachiatum T6 | 0 | 9.29 ± 0.6d | 5.84 ± 0.24d | 12.28 ± 1.2c |
| 10 | 11.46 ± 1.0c | 6.57 ± 0.26cd | 15.58 ± 0.9a | |
| 20 | 12.35 ± 0.8b | 6.75 ± 0.27bc | 15.46 ± 0.7a | |
| Sterile water | 0 | 10.88 ± 0.9c | 6.50 ± 0.27cd | 11.21 ± 1.1d |
| 10 | 14.78 ± 1.0ab | 7.43 ± 0.27ab | 13.93 ± 1.0c | |
| 20 | 15.33 ± 0.7a | 8.07 ± 0.12a | 14.01 ± 0.9b |
| Treatment | Salt Concentration (mg mL−1) | Glutathione (µmol g−1 FW) | Ascorbate (µmol g−1 FW) |
|---|---|---|---|
| Trichodermalongibrachiatum T6 | 0 | 0.99 ± 0.05cd | 0.88 ± 0.02a |
| 10 | 1.24 ± 0.04b | 0.75 ± 0.02b | |
| 20 | 1.46 ± 0.05a | 0.64 ± 0.02c | |
| Sterile water | 0 | 0.85 ± 0.01e | 0.83 ± 0.01a |
| 10 | 0.92 ± 0.04de | 0.65 ± 0.02c | |
| 20 | 1.11 ± 0.05c | 0.52 ± 0.03d |
| Genes | Premiers Sequence (5′-3′) |
|---|---|
| APX | F: AAAACCACCTACTGCCACCCTATC |
| R: AGCATTCGCTCCATGACTCAACT | |
| GST | F: AGCTCTTGGCGTCTTGGCT |
| R: AGGCTTCCCCTTGGAGCAC | |
| GPX | F: CCTAACTAACTCCAACTACACC |
| R: TCCTGCCCACCAAACTGAT | |
| GR | F: ATGAATACTCCCGTACATCAGT |
| R: TTTGTTACATCACCCACAGC | |
| DHAR | F: AGAAGTTTACGCCCTTCGGC |
| R: ACAAGTGATGGAGTTGGGT | |
| Actin | F: CCGTGGTGATGTTGTGCCAAAGGA |
| R: CGACGACACTGGTGGAGTTGGAGA |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, S.; Xu, B.; Gan, Y. Seed Treatment with Trichoderma longibrachiatum T6 Promotes Wheat Seedling Growth under NaCl Stress Through Activating the Enzymatic and Nonenzymatic Antioxidant Defense Systems. Int. J. Mol. Sci. 2019, 20, 3729. https://doi.org/10.3390/ijms20153729
Zhang S, Xu B, Gan Y. Seed Treatment with Trichoderma longibrachiatum T6 Promotes Wheat Seedling Growth under NaCl Stress Through Activating the Enzymatic and Nonenzymatic Antioxidant Defense Systems. International Journal of Molecular Sciences. 2019; 20(15):3729. https://doi.org/10.3390/ijms20153729
Chicago/Turabian StyleZhang, Shuwu, Bingliang Xu, and Yantai Gan. 2019. "Seed Treatment with Trichoderma longibrachiatum T6 Promotes Wheat Seedling Growth under NaCl Stress Through Activating the Enzymatic and Nonenzymatic Antioxidant Defense Systems" International Journal of Molecular Sciences 20, no. 15: 3729. https://doi.org/10.3390/ijms20153729
APA StyleZhang, S., Xu, B., & Gan, Y. (2019). Seed Treatment with Trichoderma longibrachiatum T6 Promotes Wheat Seedling Growth under NaCl Stress Through Activating the Enzymatic and Nonenzymatic Antioxidant Defense Systems. International Journal of Molecular Sciences, 20(15), 3729. https://doi.org/10.3390/ijms20153729