Aphid-Triggered Changes in Oxidative Damage Markers of Nucleic Acids, Proteins, and Lipids in Maize (Zea mays L.) Seedlings


Abstract
:1. Introduction
2. Results
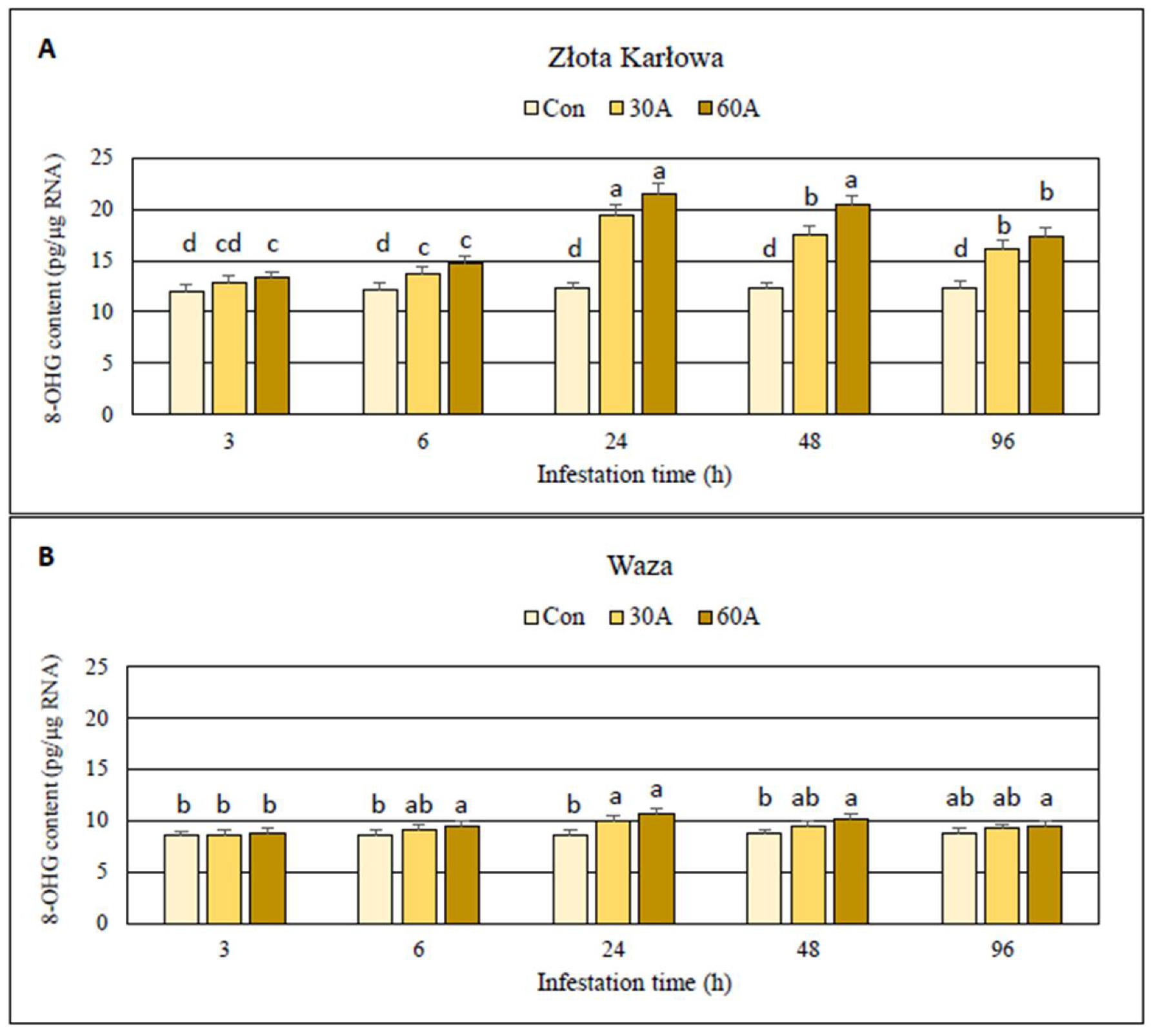
2.1. Impact of Aphid Herbivory on the Content of Oxidized Guanosine Residues in Genomic DNA and RNA Samples Derived from Maize Seedlings
2.2. Protein-Bound Carbonyls (PC) Content in Aphid-Challenged Maize Plants
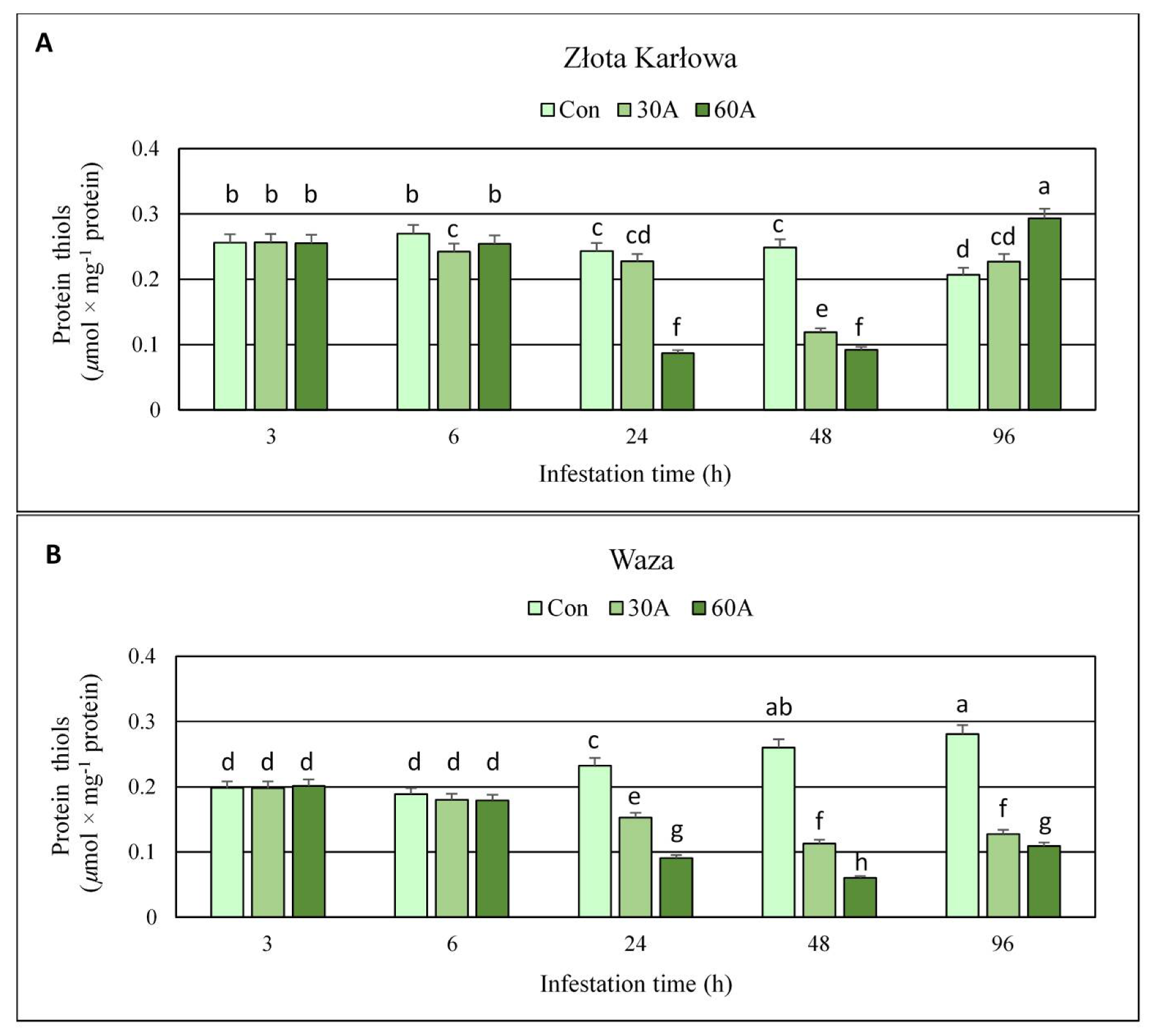
2.3. Effect of R. padi Herbivory on the Content of Protein Thiols (PT) in Maize Tissues
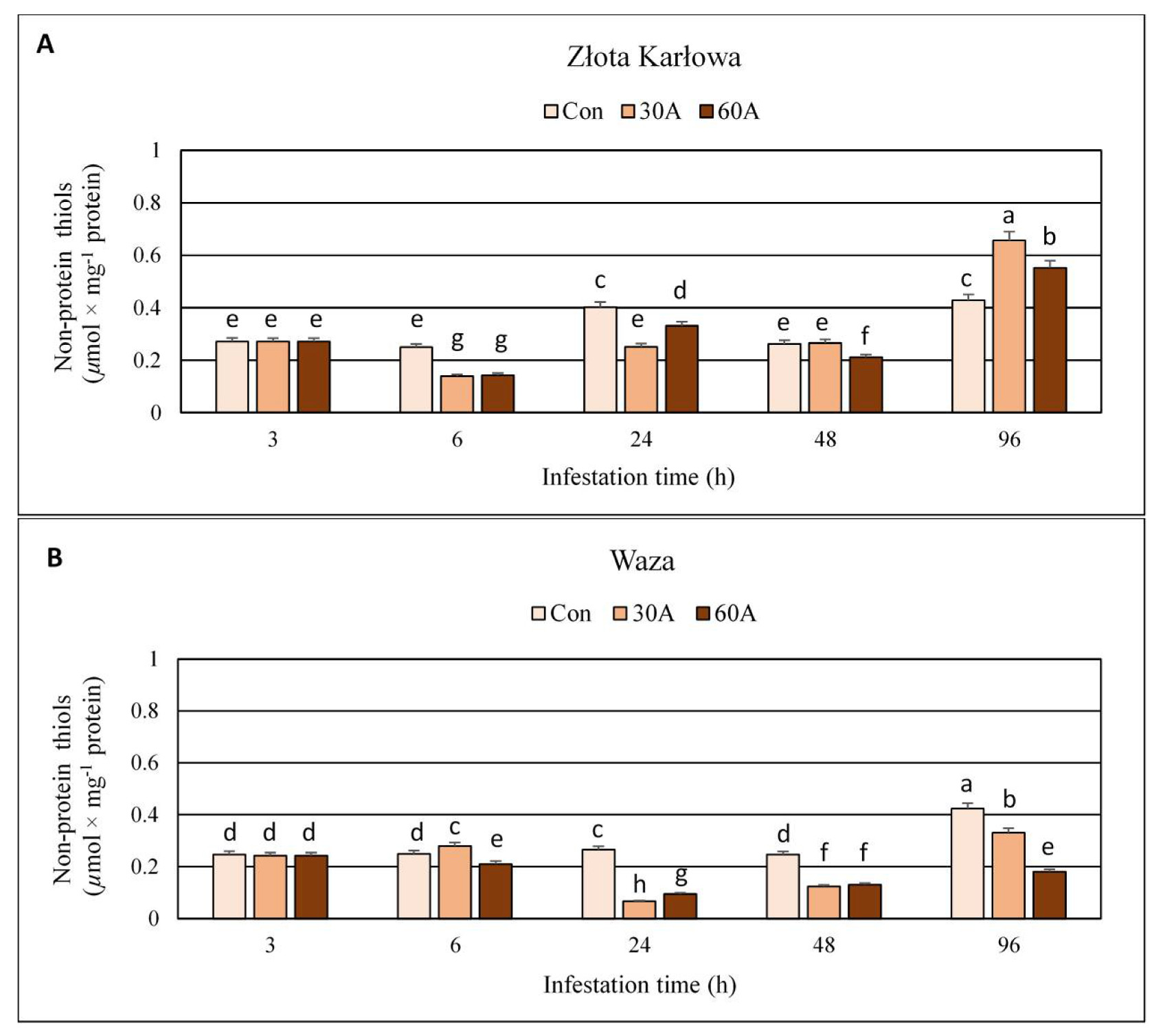
2.4. Influence of Aphid Feeding on the Content of Non-Protein Thiols (NPT) in Z. mays Seedlings
2.5. Aphid-Induced Changes in the Content of Total Thiols (TT) in Maize Seedlings
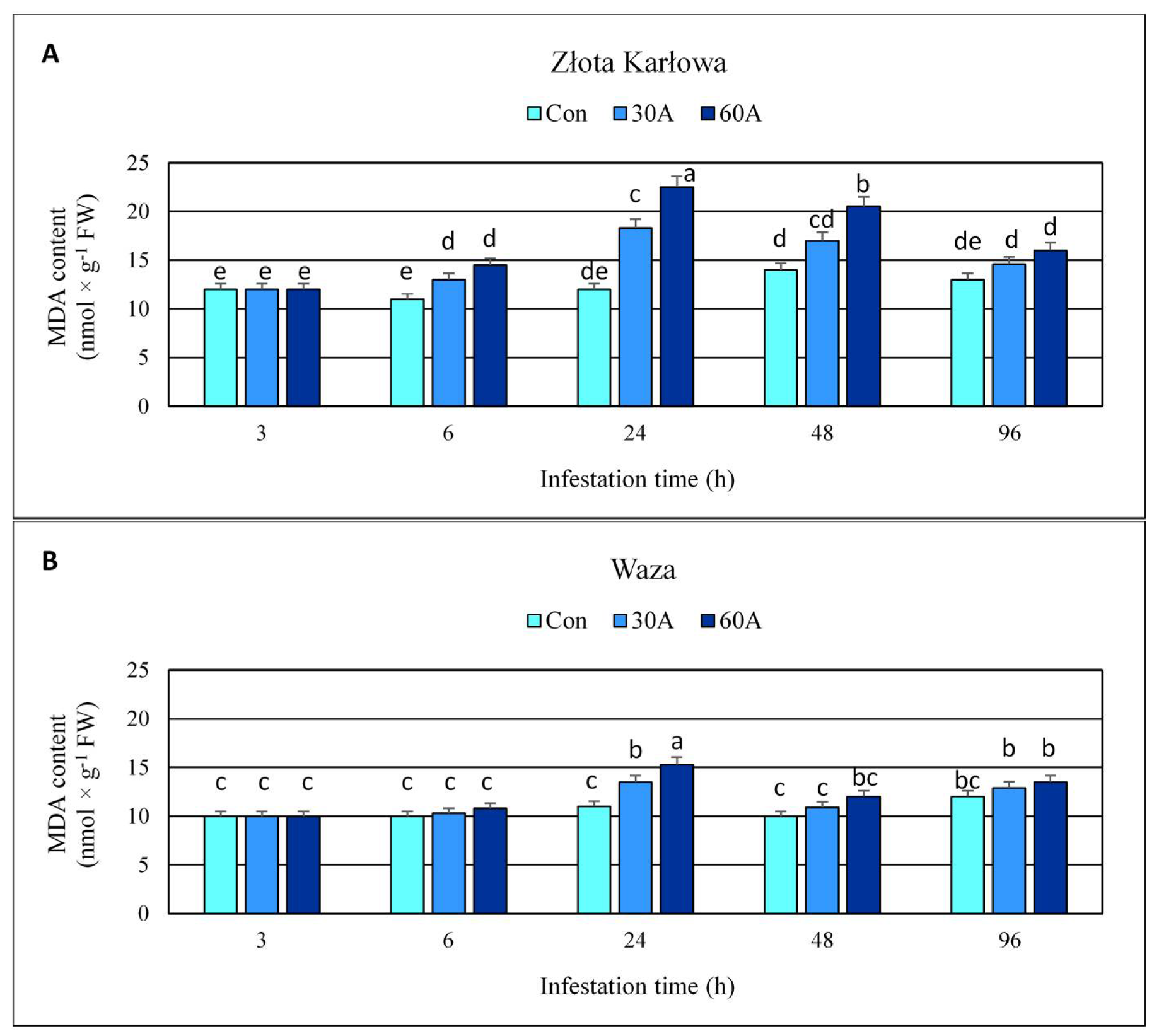
2.6. Effect of Insect Feeding on the Content of Malondialdehyde (MDA) and Plasma Membrane Stability in Maize Plants
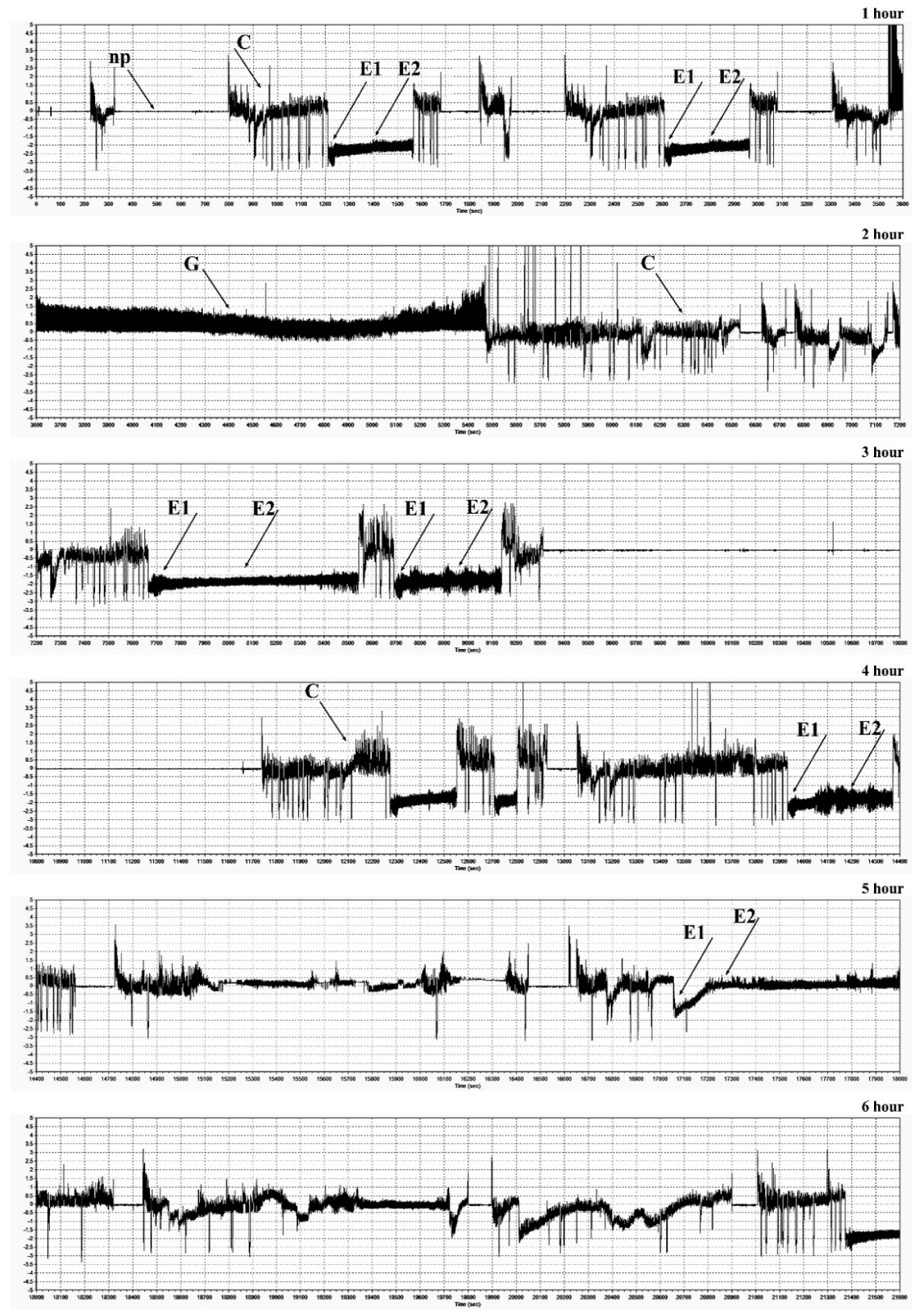
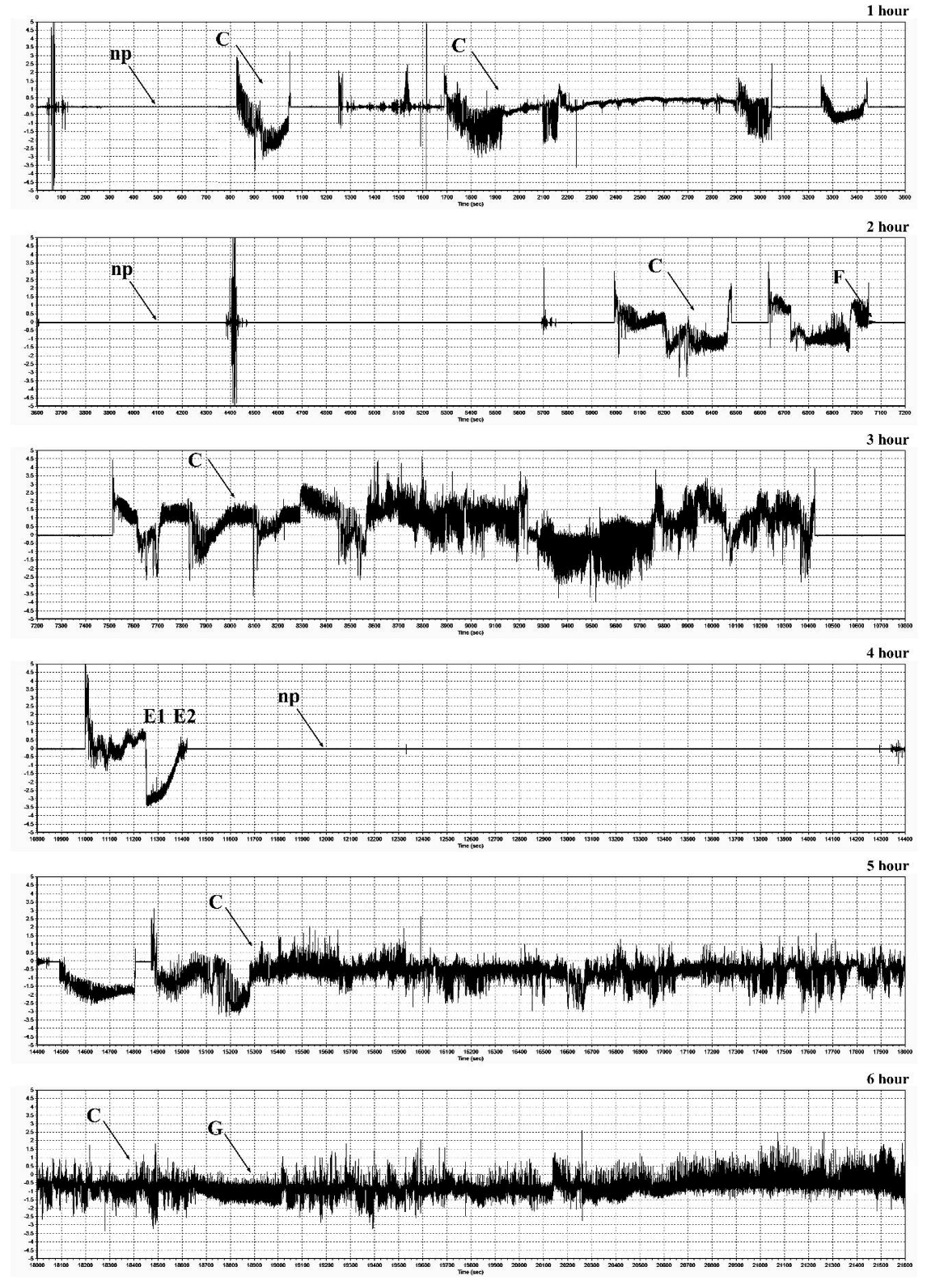
2.7. Electrical Penetration Graph (EPG) Recordings of Feeding Behavior of R. padi
3. Discussion
4. Materials and Methods
4.1. Aphids
4.2. Plant Material
4.3. Leaf Infestation Experiments
4.4. Extraction of Genomic DNA (gDNA)
4.5. Measurement of Oxidative Damages of Genomic DNA
4.6. Isolation of Total RNA and mRNA
4.7. Determination of Oxidative Damages of Total RNA and mRNA
4.8. Determination of Total Thiols (TT), Non-Protein Thiols (NPT) and Protein Thiols (PT)
4.9. Determination of Protein-Bound Carbonyls (CP)
4.10. Lipid Peroxidation Assay
4.11. Plasma Membrane Stability Assay
4.12. Electrical Penetration Graph (EPG) Recordings
4.13. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Singh, A.A.; Agrawal, S.B.; Shahi, J.P.; Agrawal, M. Yield and kernel nutritional quality in normal maize and quality protein maize cultivars exposed to ozone. J. Sci. Food. Agric. 2019, 99, 2205–2214. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, D.; Ponrajan, A.; Bonnet, J.P.; Rocheford, T.; Ferruzzi, M.G. Carotenoid stability during dry milling, storage, and extrusion processing of biofortified maize genotypes. J. Agric. Food Chem. 2018, 66, 4683–4691. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.; Ding, Z.; Li, P. Maize network analysis revealed gene modules involved in development, nutrients utilization, metabolism, and stress response. BMC Plant Biol. 2017, 17, 131. [Google Scholar] [CrossRef] [PubMed]
- Sytykiewicz, H. Transcriptional responses of catalase genes in maize seedlings exposed to cereal aphids’ herbivory. Biochem. Syst. Ecol. 2015, 60, 131–142. [Google Scholar] [CrossRef]
- Sytykiewicz, H. Differential expression of superoxide dismutase genes within aphid-stressed maize (Zea mays L.) seedlings. PLoS ONE 2014, 9, e94847. [Google Scholar] [CrossRef] [PubMed]
- Sytykiewicz, H.; Chrzanowski, G.; Czerniewicz, P.; Sprawka, I.; Łukasik, I.; Goławska, I.; Sempruch, C. Expression profiling of selected glutathione transferase genes in Zea mays (L.) seedlings infested with cereal aphids. PLoS ONE 2014, 9, e111863. [Google Scholar] [CrossRef] [PubMed]
- Sytykiewicz, H. Transcriptional reprogramming of genes related to ascorbate and glutathione biosynthesis, turnover and translocation in aphid-challenged maize seedlings. Biochem. Syst. Ecol. 2016, 69, 236–251. [Google Scholar] [CrossRef]
- Ruszkowska, M. Autumnal aphid fauna on maize as an indicator of environmental changes. Prog. Plant Prot. 2015, 55, 83–86. [Google Scholar] [CrossRef] [Green Version]
- Sytykiewicz, H.; Czerniewicz, P.; Sprawka, I.; Krzyżanowski, R. Chlorophyll content of aphid-infested seedling leaves of different maize genotypes. Acta Biol. Crac. Ser. Bot. 2013, 55, 51–60. [Google Scholar]
- Sytykiewicz, H. Expression patterns of genes involved in ascorbate-glutathione cycle in aphid-infested maize (Zea mays L.) seedlings. Int. J. Mol. Sci. 2016, 17, 268. [Google Scholar] [CrossRef]
- Sytykiewicz, H. Deciphering the role of NADPH oxidase in complex interactions between maize (Zea mays L.) genotypes and cereal aphids. Biochem. Biophys. Res. Commun. 2016, 476, 90–95. [Google Scholar] [CrossRef] [PubMed]
- Sytykiewicz, H.; Goławska, S.; Chrzanowski, G. Effect of the bird cherry-oat aphid Rhopalosiphum padi L. feeding on phytochemical responses within the bird cherry Prunus padus L. Pol. J. Ecol. 2011, 59, 329–338. [Google Scholar]
- Zehnder, G.W.; Nichols, A.J.; Edwards, O.R.; Ridsdill-Smith, T.J. Electronically monitored cowpea aphid feeding behavior on resistant and susceptible lupins. Entomol. Exp. Appl. 2001, 98, 259–269. [Google Scholar] [CrossRef]
- Tjallingii, W.F.; Hogen Esch, T. Fine structure of aphid stylet routes in plant tissues in correlation with EPG signals. Physiol. Entomol. 1993, 18, 317–328. [Google Scholar] [CrossRef]
- Song, J.; Bent, A.F. Microbial pathogens trigger host DNA double-strand breaks whose abundance is reduced by plant defense responses. PLoS Pathog. 2014, 10, e1004030. [Google Scholar] [CrossRef] [PubMed]
- Debiane, D.; Garçon, G.; Verdin, A.; Fontaine, J.; Durand, R.; Grandmougin-Ferjani, A.; Shirali, P.; Lounès-Hadj Sahraoui, A. In vitro evaluation of the oxidative stress and genotoxic potentials of anthracene on mycorrhizal chicory roots. Environ. Exp. Bot. 2008, 64, 120–127. [Google Scholar] [CrossRef]
- Bidar, G.; Pruvot, C.; Garçon, G.; Verdin, A.; Shirali, P.; Douay, F. Seasonal and annual variations of metal uptake, bioaccumulation, and toxicity in Trifolium repens and Lolium perenne growing in a heavy metal-contaminated field. Environ. Sci. Pollut. Res. 2009, 16, 42–53. [Google Scholar] [CrossRef] [PubMed]
- Bernard, F.; Dumez, S.; Lemière, S.; Platel, A.; Nesslany, F.; Deram, A.; Vandenbulcke, F.; Cuny, D. Impact of cadmium on forage kale (Brassica oleracea var. viridis cv “Prover”) after 3-,10- and 56-day exposure to a Cd-spiked field soil. Environ. Sci. Pollut. Res. Int. 2018, 1–9, (Epub ahead of print). [Google Scholar] [CrossRef]
- Manova, V.; Gruszka, D. DNA damage and repair in plants—From models to crops. Front. Plant Sci. 2015, 6, 885. [Google Scholar] [CrossRef]
- Chmielowska-Bąk, J.; Izbiańska, K.; Ekner-Grzyb, A.; Bayar, M.; Deckert, J. Cadmium stress leads to rapid increase in RNA oxidative modifications in soybean seedlings. Front. Plant Sci. 2018, 8, 2219. [Google Scholar] [CrossRef]
- Bazin, J.; Langlade, N.; Vincourt, P.; Arribat, S.; Balzergue, S.; El-Maarouf-Bouteau, H.; Bailly, C. Targeted mRNA oxidation regulates sunflower seed dormancy alleviation during dry after-ripening. Plant Cell 2011, 23, 2196–2208. [Google Scholar] [CrossRef]
- Gao, F.; Rampitsch, C.; Chitnis, V.R.; Humphreys, G.D.; Jordan, M.C.; Ayele, B.T. Integrated analysis of seed proteome and mRNA oxidation reveals distinct post-transcriptional features regulating dormancy in wheat (Triticum aestivum L.). Plant Biotechnol. J. 2013, 11, 921–932. [Google Scholar] [CrossRef] [PubMed]
- Chmielowska-Bąk, J.; Izbiańska, K.; Deckert, J. Products of lipid, protein and RNA oxidation as signals and regulators of gene expression in plants. Front. Plant Sci. 2015, 6, 405. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ye, X.Y.; Qiu, X.M.; Li, Z.G. Methylglyoxal triggers the heat tolerance in maize seedlings by driving AsA-GSH cycle and reactive oxygen species-/methylglyoxal-scavenging system. Plant Physiol. Biochem. 2019, 138, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Bi, W.; Liu, X.; Sun, L.; Li, J. Effects of waterlogging stress on the physiological response and grain-filling characteristics of spring maize (Zea mays L.) under field conditions. Acta Physiol. Plant. 2019, 41, 63. [Google Scholar] [CrossRef]
- Li, Z.G.; Xu, Y.; Bai, L.K.; Zhang, S.Y.; Wang, Y. Melatonin enhances thermotolerance of maize seedlings (Zea mays L.) by modulating antioxidant defense, methylglyoxal detoxification, and osmoregulation systems. Protoplasma 2019, 256, 471. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Cui, Q.; Yue, S.; Lu, Z.; Zhao, M. Enantioselective effect of glufosinate on the growth of maize seedlings. Environ. Sci. Pollut. Res. 2019, 26, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Janicka-Russak, M.; Kabała, K.; Burzynski, M. Different effect of cadmium and copper on H+-ATPase activity in plasma membrane vesicles from Cucumis sativus roots. J. Exp. Bot. 2012, 63, 4133–4142. [Google Scholar] [CrossRef]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell Longev. 2014, 360438. [Google Scholar] [CrossRef]
- Demidchik, V.; Straltsova, D.; Medvedev, S.S.; Pozhvanov, G.A.; Sokolik, A.; Yurin, V. Stress-induced electrolyte leakage: The role of K+-permeable channels and involvement in programmed cell death and metabolic adjustment. J. Exp. Bot. 2014, 65, 1259–1270. [Google Scholar] [CrossRef]
- Mendoza-Cózatl, D.G.; Moreno-Sánchez, R. Control of glutathione and phytochelatin synthesis under cadmium stress. Pathway modeling for plants. J. Theor. Biol. 2006, 238, 919–936. [Google Scholar] [CrossRef] [PubMed]
- Deneke, S.M. Thiol-based antioxidants. Curr. Top. Cell Regul. 2000, 36, 151–180. [Google Scholar] [PubMed]
- Sen, C.K. Cellular thiols and redox-regulated signal transduction. Curr. Top. Cell Regul. 2000, 36, 1–30. [Google Scholar] [PubMed]
- Bhoomika, K.; Pyngrope, S.; Dubey, R.S. Effect of aluminum on protein oxidation, non-protein thiols and protease activity in seedlings of rice cultivars differing in aluminum tolerance. J. Plant Physiol. 2014, 171, 497–508. [Google Scholar] [CrossRef] [PubMed]
- Nagalakshmi, N.; Prasad, M.N. Responses of glutathione cycle enzymes and glutathione metabolism to copper stress in Scendesmus bijugatus. Plant Sci. 2001, 160, 291–299. [Google Scholar] [CrossRef]
- Aly, A.A.; Mohamed, A.A. The impact of copper ion on growth, thiol compound and lipid peroxidation in two maize cultivars (Zea mays L.) grown in vitro. Aust. J. Crop Sci. 2012, 6, 541–549. [Google Scholar]
- Kaur, P.; Bali, S.; Sharma, A.; Vig, A.P.; Bhardwaj, R. Role of earthworms in phytoremediation of cadmium (Cd) by modulating the antioxidative potential of Brassica juncea L. Appl. Soil Ecol. 2018, 124, 306–316. [Google Scholar] [CrossRef]
- Nemat Alla, M.M.; Hassan, N.M. Changes of antioxidants and GSH-associated enzymes in isoproturon-treated maize. Acta Physiol. Plant. 2007, 29, 247–258. [Google Scholar] [CrossRef]
- Baisakhi, B.; Patra, J.; Panigrahy, R.K.; Panda, B.B. Differential antioxidant enzyme and thiol responses of tolerant and non-tolerant clones of Chloris barbata to cadmium-stress. Acta Physiol. Plant. 2003, 25, 357–363. [Google Scholar] [CrossRef]
- Rinalducci, S.; Murgiano, L.; Zolla, L. Redox proteomics: Basic principles and future perspectives for the detection of protein oxidation in plants. J. Exp. Bot. 2008, 59, 3781–3801. [Google Scholar] [CrossRef] [PubMed]
- Anjum, N.A.; Sofo, A.; Scopa, A.; Roychoudhury, A.; Gill, S.S.; Iqbal, M. Lipids and proteins-major targets of oxidative modifications in abiotic stressed plants. Environ. Sci. Pollut. Res. 2005, 22, 4099–4121. [Google Scholar] [CrossRef] [PubMed]
- Boguszewska-Mańkowska, D.; Nykiel, M.; Zagdańska, B. Protein oxidation and redox regulation of proteolysis. In Basic Principles and Clinical Significance of Oxidative Stress; InTech Open: London, UK, 2015; Volume 17, p. 35. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 2012, 26. [Google Scholar] [CrossRef]
- Ramakrishna, B.; Rao, S.S.R. 24-Epibrassinolide alleviated zinc-induced oxidative stress in radish (Raphanus sativus L.) seedlings by enhancing antioxidative system. Plant Growth Regul. 2012, 68, 249–259. [Google Scholar] [CrossRef]
- Zagorchev, L.; Terzieva, M.; Stoichkova, M.; Odjakova, M. Changes in protein thiols in response to salt stress in embryogenic suspension cultures of Dactylis glomerata L. Biotechnol. Biotechnol. Equip. 2014, 28, 616–621. [Google Scholar] [CrossRef] [PubMed]
- Gietler, M.; Nykiel, M.; Zagdańska, B. Changes in the reduction state of ascorbate and glutathione, protein oxidation and hydrolysis leading to the development of dehydration intolerance in Triticum aestivum L. seedlings. Plant Growth Regul. 2016, 79, 287–297. [Google Scholar] [CrossRef]
- Richter, T.; Munch, G.; Luth, H.J.; Arendt, T.; Kientsch-Engel, R.; Stahl, P.; Kuhla, B. Immunochemical crossreactivity of antibodies specific for “advanced glycation endproducts” with “advanced lipoxidation endproducts”. Neurobiol. Aging 2005, 26, 465–474. [Google Scholar] [CrossRef]
- Davies, M.J. The oxidative environment and protein damage. Biochim. Biophys. Acta. 2005, 1703, 93–109. [Google Scholar] [CrossRef]
- Møller, I.M.; Jensen, P.E.; Hansson, A. Oxidative modifications to cellular components in plants. Ann. Rev. Plant Biol. 2007, 58, 459–481. [Google Scholar] [CrossRef]
- Juszczuk, I.M.; Tybura, A.; Rychter, A.M. Protein oxidation in the leaves and roots of cucumber plants (Cucumis sativus L.) mutant MSC16 and wild type. J. Plant Physiol. 2008, 165, 355–365. [Google Scholar] [CrossRef]
- Pyngrope, S.; Kumari, B.; Dubey, R.S. Oxidative stress, protein carbonylation, proteolysis and antioxidative defense system as a model for depicting water deficit tolerance in Indica rice seedlings. Plant Growth Regul. 2013, 69, 149–165. [Google Scholar] [CrossRef]
- Roychoudhury, A.; Basu, S.; Sengupta, D.N. Antioxidants and stress-related metabolites in the seedlings of two indica rice varieties exposed to cadmium chloride toxicity. Acta Physiol. Plant. 2012, 34, 835–847. [Google Scholar] [CrossRef]
- Kaur, G.; Kaur, S.; Singh, H.P.; Batish, D.R.; Kohli, R.K.; Rishi, V. Biochemical adaptations in Zea mays roots to short-term Pb2+ exposure: ROS generation and metabolism. Bull. Environ. Contam. Toxicol. 2015, 9, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Maiti, S.; Ghosh, N.; Mandal, C.; Das, K.; Dey, N.; Adk, M.K. Responses of the maize plant to chromium stress with reference to antioxidation activity. Braz. J. Plant Physiol. 2012, 24, 203–212. [Google Scholar] [CrossRef] [Green Version]
- Boscolo, P.R.S.; Menossi, M.; Jorge, R.A. Aluminum-induced oxidative stress in maize. Phytochemistry 2003, 62, 181–189. [Google Scholar] [CrossRef]
- Dworak, A.; Nykiel, M.; Walczak, B.; Miazek, A.; Szworst-Łupina, D.; Zagdańska, B.; Kiełkiewicz, M. Maize proteomic responses to separate or overlapping soil drought and two-spotted spiker mite stresses. Planta 2016, 244, 939–960. [Google Scholar] [CrossRef] [PubMed]
- Ban, L.; Ahmed, E.; Ninkovic, V.; Dlp, G.; Glinwood, R. Infection with an insect virus affect olfactory behavior and interactions with host plant and natural enemies in an aphid. Entomol. Exp. Appl. 2008, 127, 108–117. [Google Scholar] [CrossRef]
- Goławska, S.; Leszczyński, B.; Oleszek, W. Effect of low and high-saponin lines of alfalfa on pea aphid. J. Insect Physiol. 2006, 52, 737–743. [Google Scholar] [CrossRef]
- Goławska, S.; Łukasik, I.; Leszczyński, B. Effect of alfalfa saponins and flavonoids on pea aphid. Entomol. Exp. Appl. 2008, 128, 147–153. [Google Scholar] [CrossRef]
- Gabryś, B.; Pawluk, M. Acceptability of different species of Brassiceae as hosts for the cabbage aphid. Entomol. Exp. Appl. 1999, 91, 105–109. [Google Scholar] [CrossRef]
- Kordan, B.; Gabryś, B.; Dancewicz, K.; Lahuta, L.B.; Piotrowicz-Cieślak, A.; Rowińska, E. European yellow lupine, Lupinus luteus, and narrow-leaf lupine, Lupinus angustifolius, as hosts for the pea aphid, Acyrthosiphon pisum. Entomol. Exp. Appl. 2008, 128, 139–146. [Google Scholar] [CrossRef]
- De Kok, L.J.; Kuiper, P.J.C. Effect of short-term dark incubation with sulfate, chloride and selenate on the glutathione content of spinach leaf discs. Physiol. Plant. 1986, 68, 477–482. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Levine, R.L.; Williams, J.A.; Stadtman, E.R.; Shacter, E. Carbonyl assays for determination of oxidatively modified proteins. Methods Enzymol. 1994, 233, 346–357. [Google Scholar] [PubMed]
- Jiang, M.; Zhang, J. Effect of abscisic acid on active oxygen species, antioxidative defence system and oxidative damage in leaves of maize seedlings. Plant Cell Physiol. 2001, 42, 1265–1273. [Google Scholar] [CrossRef] [PubMed]
- Tjallingii, W.F. Sieve element acceptance by aphids. Eur. J. Entomol. 1994, 91, 47–52. [Google Scholar]
- Sokal, R.R.; Rohlf, F.J. Biometry, 3rd ed.; Freeman, Co.: New York, NY, USA, 2001; p. 887. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| EPG Waveforms | EPG Activity | |||
|---|---|---|---|---|
| Number of Events | Total Duration (min) | Average Time (min) | Time to First Probing (min) | |
| Złota Karłowa cv. (susceptible) | ||||
| Np | 11.00 ± 4.71a | 53.42 ± 15.02b | 5.37 ± 2.13a | n/a |
| C | 19.93 ± 6.91a | 148.48 ± 62.01a | 7.90 ± 3.48a | 2.87 ± 1.77a |
| E1 | 9.33 ± 3.98a | 26.16 ± 20.69a | 2.80 ± 1.98a | 37.07 ± 10.71a |
| E2 | 6.73 ± 3.22a | 130.73 ± 75.24a | 23.88 ± 19.44a | 42.34 ± 11.22a |
| G | 1.20 ± 1.78a | 1.21 ± 128.62a | 0.46 ± 0.96b | 100.13 ± 142.93a |
| F | 0.00 ± 0.00b | 0.00 ± 0.00a | 0.00 ± 0.00a | 0.00 ± 0.00b |
| Waza cv. (relatively resistant) | ||||
| Np | 12.80 ± 4.54a | 143.31 ± 57.83a | 12.53 ± 6.74a | n/a |
| C | 19.27 ± 7.78a | 152.04 ± 41.09a | 8.52 ± 2.63a | 4.77 ± 2.96a |
| E1 | 6.00 ± 4.05a | 3.87 ± 3.39b | 0.58 ± 0.26a | 82.64 ± 17.28a |
| E2 | 5.00 ± 3.07a | 22.65 ± 15.78b | 4.21 ± 2.96b | 83.32 ± 17.38a |
| G | 2.93 ± 2.02a | 37.66 ± 28.66a | 11.92 ± 5.65a | 81.23 ± 48.53a |
| F | 1.07 ± 0.59a | 0.47 ± 0.39a | 0.38 ± 0.29a | 88.64 ± 66.42a |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sytykiewicz, H.; Łukasik, I.; Goławska, S.; Chrzanowski, G. Aphid-Triggered Changes in Oxidative Damage Markers of Nucleic Acids, Proteins, and Lipids in Maize (Zea mays L.) Seedlings. Int. J. Mol. Sci. 2019, 20, 3742. https://doi.org/10.3390/ijms20153742
Sytykiewicz H, Łukasik I, Goławska S, Chrzanowski G. Aphid-Triggered Changes in Oxidative Damage Markers of Nucleic Acids, Proteins, and Lipids in Maize (Zea mays L.) Seedlings. International Journal of Molecular Sciences. 2019; 20(15):3742. https://doi.org/10.3390/ijms20153742
Chicago/Turabian StyleSytykiewicz, Hubert, Iwona Łukasik, Sylwia Goławska, and Grzegorz Chrzanowski. 2019. "Aphid-Triggered Changes in Oxidative Damage Markers of Nucleic Acids, Proteins, and Lipids in Maize (Zea mays L.) Seedlings" International Journal of Molecular Sciences 20, no. 15: 3742. https://doi.org/10.3390/ijms20153742
APA StyleSytykiewicz, H., Łukasik, I., Goławska, S., & Chrzanowski, G. (2019). Aphid-Triggered Changes in Oxidative Damage Markers of Nucleic Acids, Proteins, and Lipids in Maize (Zea mays L.) Seedlings. International Journal of Molecular Sciences, 20(15), 3742. https://doi.org/10.3390/ijms20153742