Ovarian Expression of Adipokines in Polycystic Ovary Syndrome: A Role for Chemerin, Omentin, and Apelin in Follicular Growth Arrest and Ovulatory Dysfunction?


Abstract
:1. Introduction
2. Results
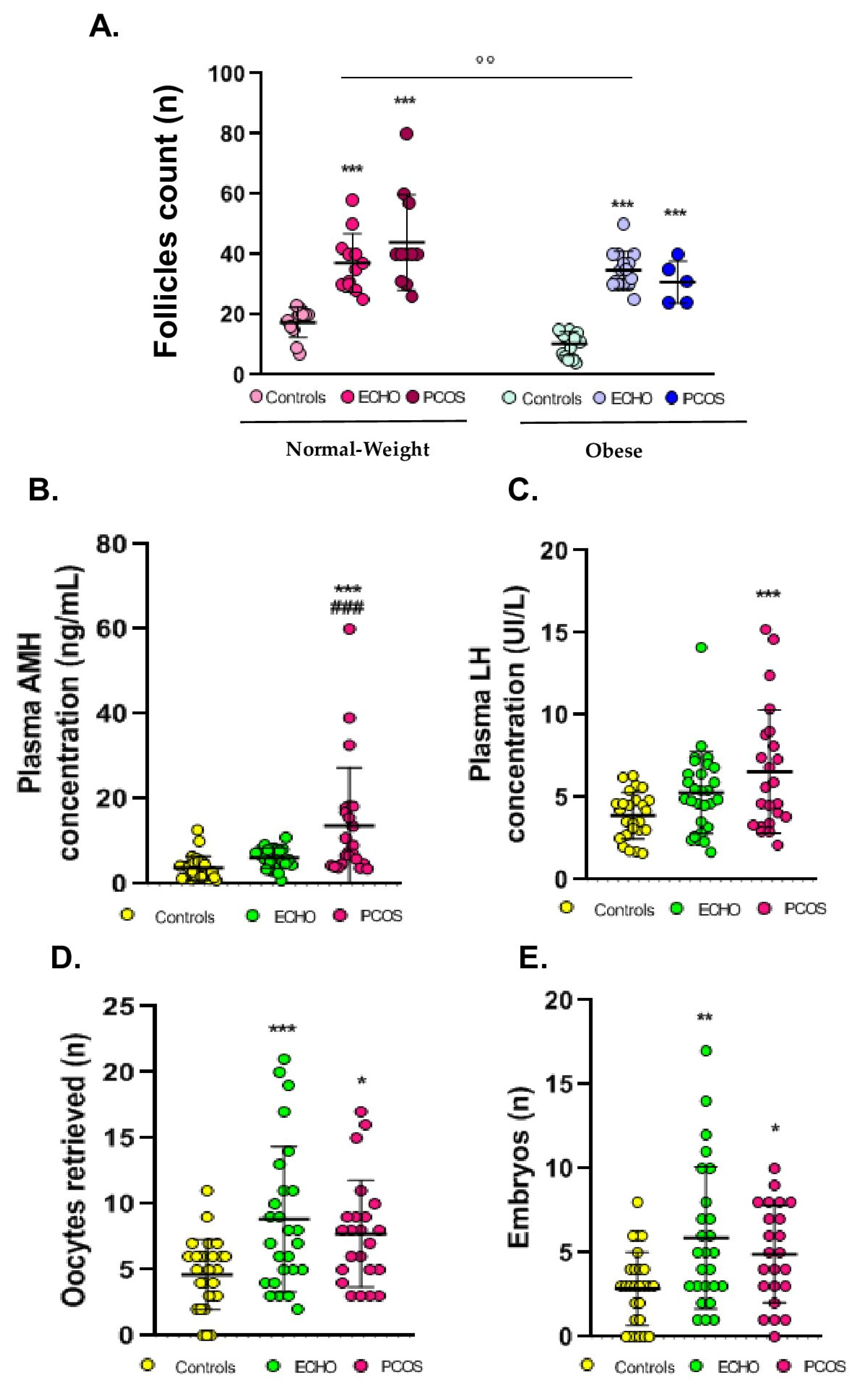
2.1. Anthropometric, Clinical, and Hormonal Data
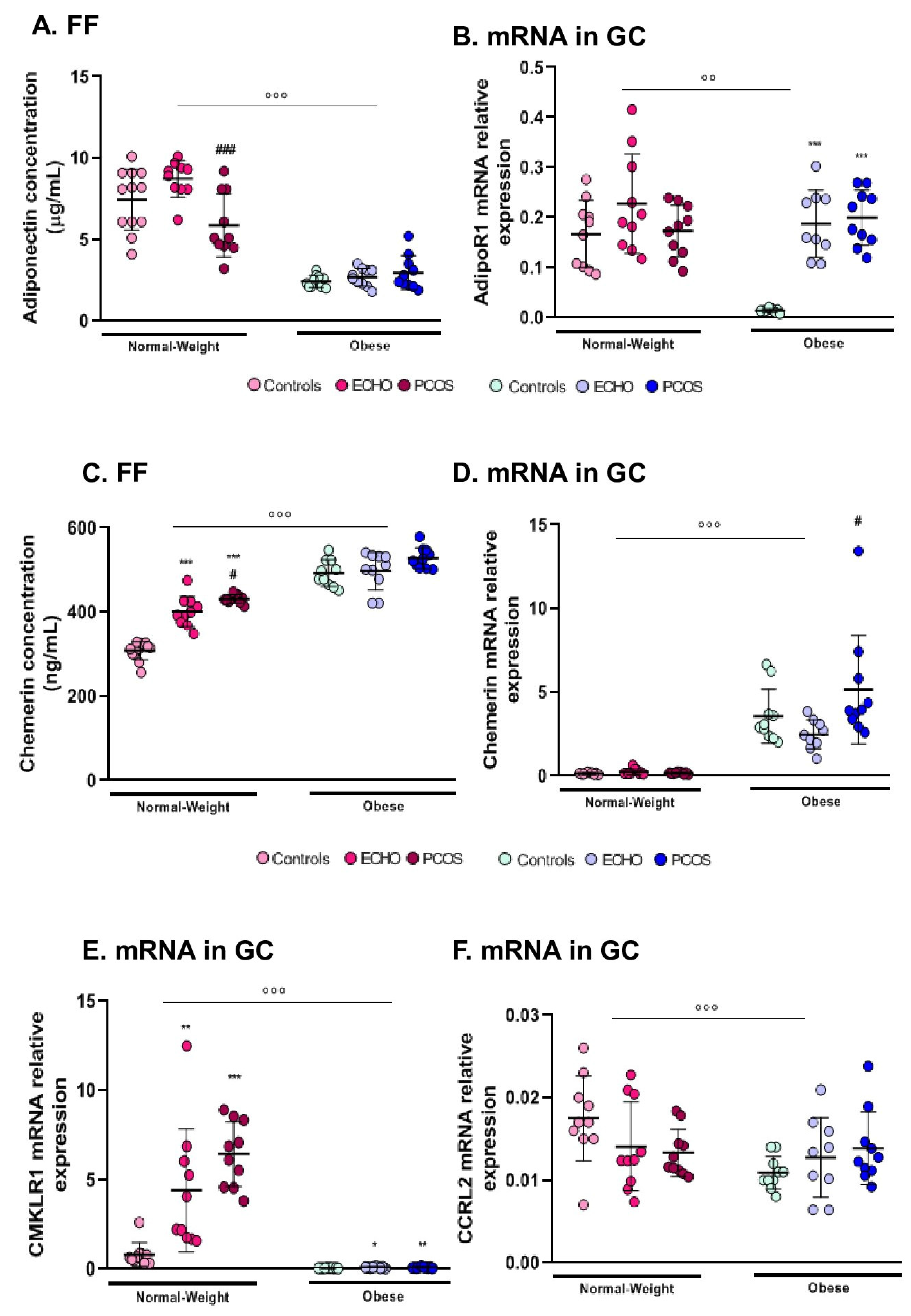
2.2. FF Adiponectin and AdipoR1 Expression in GC Varied Mainly According to BMI
2.3. Chemerin (FF and GC mRNA Expression) Was Higher in Obese Subjects and in Women with PCOS
2.4. CMKLR1 and CCRL2 mRNA Expression in GC was Markedly Reduced in Obese Women
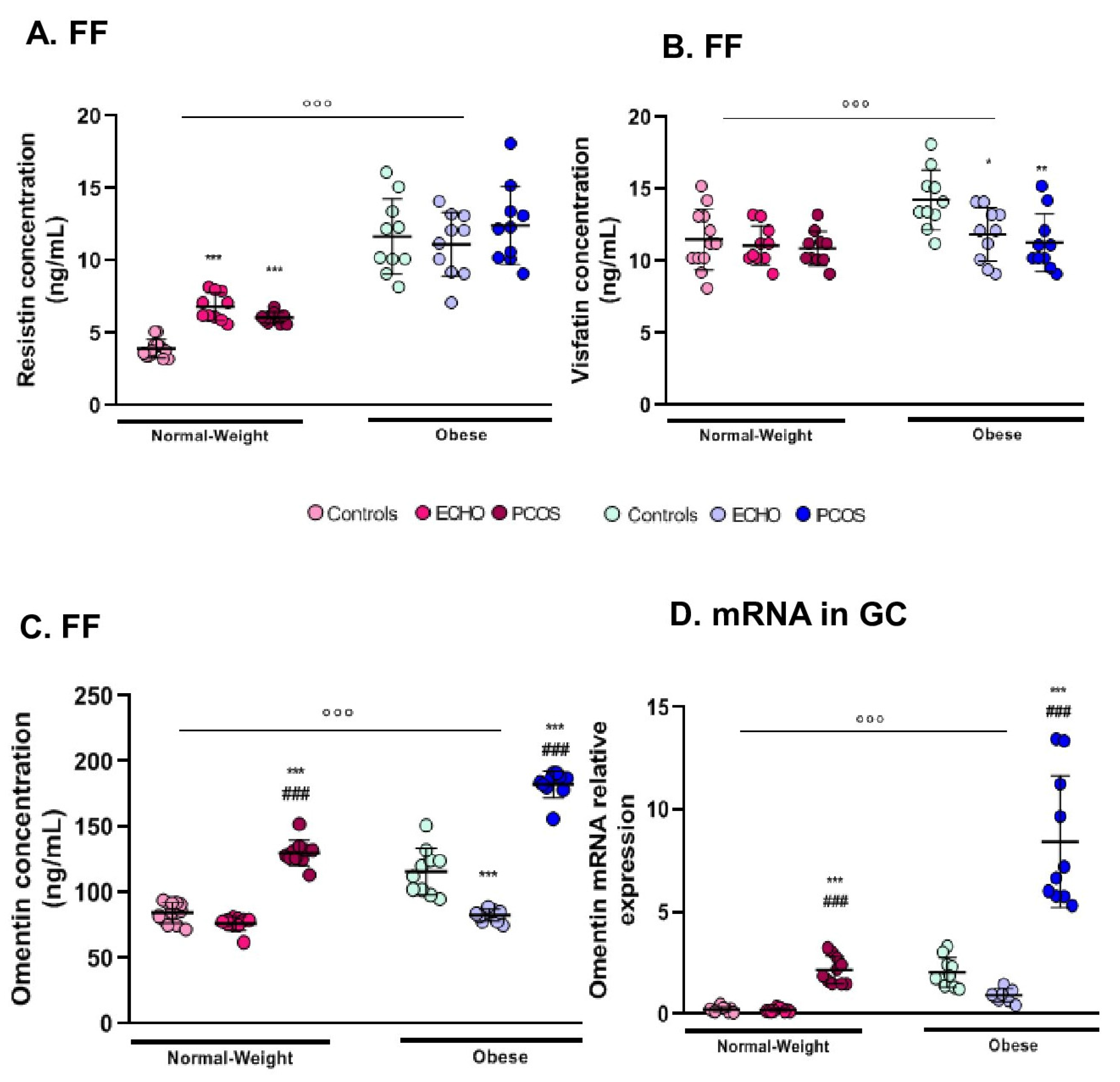
2.5. FF Resistin Was Higher in Obese Women
2.6. Visfatin Modifications Were Restrained to Its Concentration in FF
2.7. Omentin Expression (FF and GC mRNA) Was Markedly Predominant in Women with PCOS
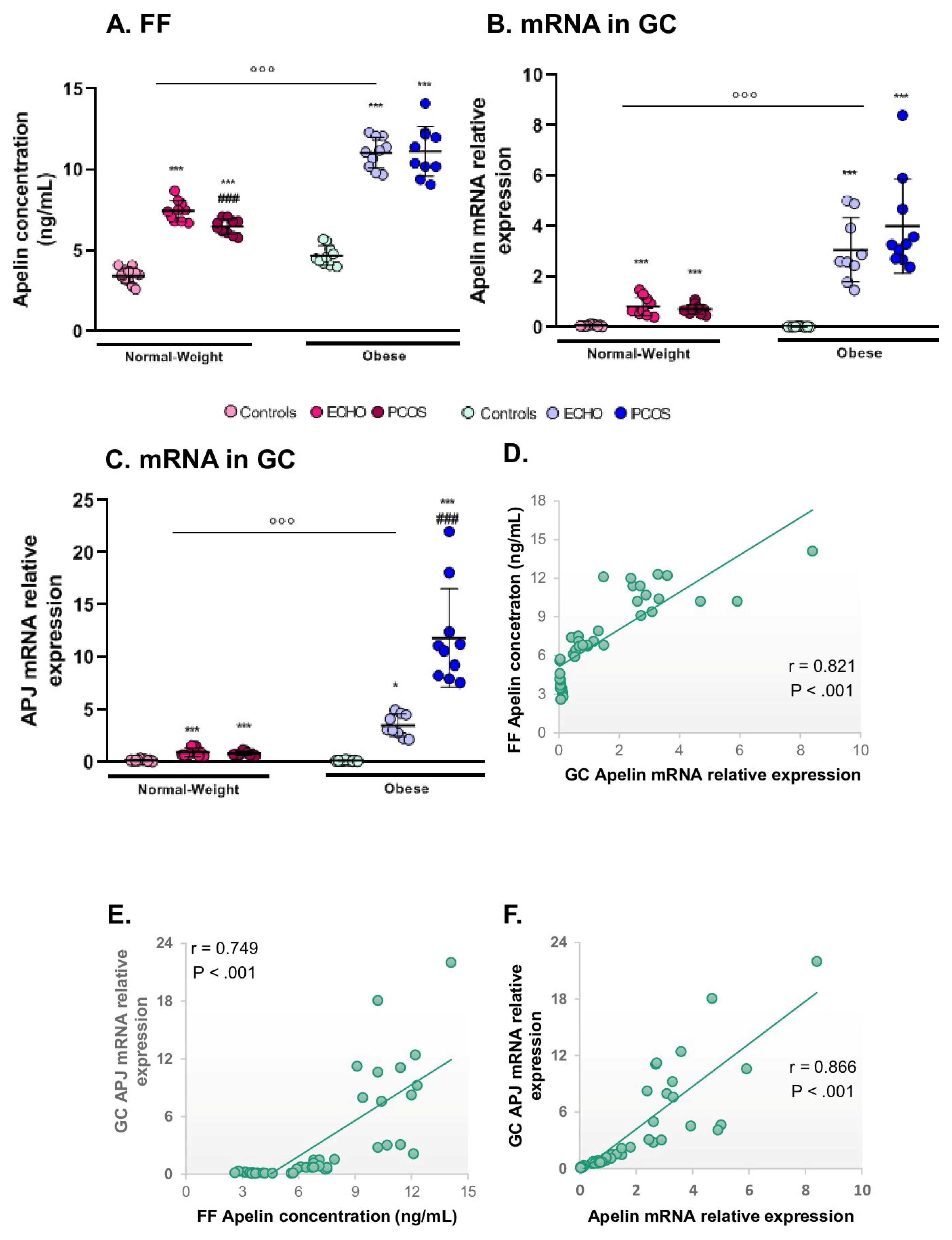
2.8. Apelin and Its Receptor APJ Were Mostly Expressed in Obese Subjects and in ECHO/PCOS Women
3. Discussion
3.1. Adiponectin
3.2. Resistin
3.3. Visfatin
3.4. Apelin
3.5. Omentin
3.6. Chemerin
3.7. ECHO Condition
3.8. Limitations and Perspectives
4. Materials and Methods
4.1. Ethics Approval
4.2. Study Population
4.3. Collection of FF Samples and Isolation of GCs
4.4. Adipokines Concentration in FF
4.5. RNA Extraction and Real-Time Quantitative PCR (qPCR)
4.6. Statistical Analyses
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Teede, H.; Deeks, A.; Moran, L. Polycystic ovary syndrome: A complex condition with psychological, reproductive and metabolic manifestations that impacts on health across the lifespan. BMC Med. 2010, 8, 41. [Google Scholar] [CrossRef] [PubMed]
- Rotterdam ESHRE/ASRM-Sponsored PCOS Consensus Workshop Group. Revised 2003 consensus on diagnostic criteria and long-term health risks related to polycystic ovary syndrome. Fertil. Steril. 2004, 81, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Teede, H.J.; Misso, M.L.; Costello, M.F.; Dokras, A.; Laven, J.; Moran, L.; Piltonen, T.; Norman, R.J.; International, P.N. Recommendations from the international evidence-based guideline for the assessment and management of polycystic ovary syndrome. Fertil. Steril. 2018, 110, 364–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toulis, K.A.; Goulis, D.G.; Farmakiotis, D.; Georgopoulos, N.A.; Katsikis, I.; Tarlatzis, B.C.; Papadimas, I.; Panidis, D. Adiponectin levels in women with polycystic ovary syndrome: A systematic review and a meta-analysis. Hum. Reprod. Update 2009, 15, 297–307. [Google Scholar] [CrossRef] [PubMed]
- Benrick, A.; Chanclon, B.; Micallef, P.; Wu, Y.; Hadi, L.; Shelton, J.M.; Stener-Victorin, E.; Wernstedt Asterholm, I. Adiponectin protects against development of metabolic disturbances in a PCOS mouse model. Proc. Natl. Acad. Sci. USA 2017, 114, E7187–E7196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasquali, R.; Pelusi, C.; Genghini, S.; Cacciari, M.; Gambineri, A. Obesity and reproductive disorders in women. Hum. Reprod. Update 2003, 9, 359–372. [Google Scholar] [CrossRef] [Green Version]
- Norman, J.E. The adverse effects of obesity on reproduction. Reproduction 2010, 140, 343–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasquali, R.; Gambineri, A.; Pagotto, U. The impact of obesity on reproduction in women with polycystic ovary syndrome. BJOG 2006, 113, 1148–1159. [Google Scholar] [CrossRef] [PubMed]
- Bluher, M. Adipose tissue dysfunction in obesity. Exp. Clin. Endocrinol. Diabetes 2009, 117, 241–250. [Google Scholar] [CrossRef]
- Ohashi, K.; Shibata, R.; Murohara, T.; Ouchi, N. Role of anti-inflammatory adipokines in obesity-related diseases. Trends. Endocrinol. Metab. 2014, 25, 348–355. [Google Scholar] [CrossRef]
- Vazquez, M.J.; Romero-Ruiz, A.; Tena-Sempere, M. Roles of leptin in reproduction, pregnancy and polycystic ovary syndrome: Consensus knowledge and recent developments. Metabolism 2015, 64, 79–91. [Google Scholar] [CrossRef] [PubMed]
- Landry, D.; Cloutier, F.; Martin, L.J. Implications of leptin in neuroendocrine regulation of male reproduction. Reprod. Biol. 2013, 13, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Dupont, J.; Pollet-Villard, X.; Reverchon, M.; Mellouk, N.; Levy, R. Adipokines in human reproduction. Horm. Mol. Biol. Clin. Investig. 2015, 24, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Chabrolle, C.; Tosca, L.; Rame, C.; Lecomte, P.; Royere, D.; Dupont, J. Adiponectin increases insulin-like growth factor I-induced progesterone and estradiol secretion in human granulosa cells. Fertil. Steril. 2009, 92, 1988–1996. [Google Scholar] [CrossRef] [PubMed]
- Reverchon, M.; Cornuau, M.; Rame, C.; Guerif, F.; Royere, D.; Dupont, J. Chemerin inhibits IGF-1-induced progesterone and estradiol secretion in human granulosa cells. Hum. Reprod. 2012, 27, 1790–1800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reverchon, M.; Cornuau, M.; Rame, C.; Guerif, F.; Royere, D.; Dupont, J. Resistin decreases insulin-like growth factor I-induced steroid production and insulin-like growth factor I receptor signaling in human granulosa cells. Fertil. Steril. 2013, 100, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Reverchon, M.; Cornuau, M.; Cloix, L.; Rame, C.; Guerif, F.; Royere, D.; Dupont, J. Visfatin is expressed in human granulosa cells: Regulation by metformin through AMPK/SIRT1 pathways and its role in steroidogenesis. Mol. Hum. Reprod. 2013, 19, 313–326. [Google Scholar] [CrossRef] [PubMed]
- Cloix, L.; Reverchon, M.; Cornuau, M.; Froment, P.; Rame, C.; Costa, C.; Froment, G.; Lecomte, P.; Chen, W.; Royere, D.; et al. Expression and regulation of INTELECTIN1 in human granulosa-lutein cells: Role in IGF-1-induced steroidogenesis through NAMPT. Biol. Reprod. 2014, 91, 50. [Google Scholar] [CrossRef]
- Roche, J.; Rame, C.; Reverchon, M.; Mellouk, N.; Cornuau, M.; Guerif, F.; Froment, P.; Dupont, J. Apelin (APLN) and Apelin Receptor (APLNR) in Human Ovary: Expression, Signaling, and Regulation of Steroidogenesis in Primary Human Luteinized Granulosa Cells. Biol. Reprod. 2016, 95, 104. [Google Scholar] [CrossRef]
- Tang, M.; Huang, C.; Wang, Y.F.; Ren, P.G.; Chen, L.; Xiao, T.X.; Wang, B.B.; Pan, Y.F.; Tsang, B.K.; Zabel, B.A.; et al. CMKLR1 deficiency maintains ovarian steroid production in mice treated chronically with dihydrotestosterone. Sci. Rep. 2016, 6, 21328. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Kim, J.Y.; Xue, K.; Liu, J.Y.; Leader, A.; Tsang, B.K. Chemerin, a novel regulator of follicular steroidogenesis and its potential involvement in polycystic ovarian syndrome. Endocrinology 2012, 153, 5600–5611. [Google Scholar] [CrossRef]
- Tan, B.K.; Chen, J.; Farhatullah, S.; Adya, R.; Kaur, J.; Heutling, D.; Lewandowski, K.C.; O’Hare, J.P.; Lehnert, H.; Randeva, H.S. Insulin and metformin regulate circulating and adipose tissue chemerin. Diabetes 2009, 58, 1971–1977. [Google Scholar] [CrossRef]
- Bozaoglu, K.; Segal, D.; Shields, K.A.; Cummings, N.; Curran, J.E.; Comuzzie, A.G.; Mahaney, M.C.; Rainwater, D.L.; VandeBerg, J.L.; MacCluer, J.W.; et al. Chemerin is associated with metabolic syndrome phenotypes in a Mexican-American population. J. Clin. Endocrinol. Metab. 2009, 94, 3085–3088. [Google Scholar] [CrossRef]
- Munir, I.; Yen, H.W.; Baruth, T.; Tarkowski, R.; Azziz, R.; Magoffin, D.A.; Jakimiuk, A.J. Resistin stimulation of 17alpha-hydroxylase activity in ovarian theca cells in vitro: Relevance to polycystic ovary syndrome. J. Clin. Endocrinol. Metab. 2005, 90, 4852–4857. [Google Scholar] [CrossRef]
- Baldani, D.P.; Skrgatic, L.; Kasum, M.; Zlopasa, G.; Kralik Oguic, S.; Herman, M. Altered leptin, adiponectin, resistin and ghrelin secretion may represent an intrinsic polycystic ovary syndrome abnormality. Gynecol. Endocrinol. 2019, 35, 401–405. [Google Scholar] [CrossRef] [Green Version]
- Chan, T.F.; Chen, Y.L.; Chen, H.H.; Lee, C.H.; Jong, S.B.; Tsai, E.M. Increased plasma visfatin concentrations in women with polycystic ovary syndrome. Fertil. Steril. 2007, 88, 401–405. [Google Scholar] [CrossRef]
- Panidis, D.; Farmakiotis, D.; Rousso, D.; Katsikis, I.; Delkos, D.; Piouka, A.; Gerou, S.; Diamanti-Kandarakis, E. Plasma visfatin levels in normal weight women with polycystic ovary syndrome. Eur. J. Intern. Med. 2008, 19, 406–412. [Google Scholar] [CrossRef]
- Tan, B.K.; Adya, R.; Farhatullah, S.; Chen, J.; Lehnert, H.; Randeva, H.S. Metformin treatment may increase omentin-1 levels in women with polycystic ovary syndrome. Diabetes 2010, 59, 3023–3031. [Google Scholar] [CrossRef]
- Orlik, B.; Madej, P.; Owczarek, A.; Skalba, P.; Chudek, J.; Olszanecka-Glinianowicz, M. Plasma omentin and adiponectin levels as markers of adipose tissue dysfunction in normal weight and obese women with polycystic ovary syndrome. Clin. Endocrinol. (Oxf) 2014, 81, 529–535. [Google Scholar] [CrossRef]
- Barbe, A.; Bongrani, A.; Mellouk, N.; Estienne, A.; Kurowska, P.; Grandhaye, J.; Elfassy, Y.; Levy, R.; Rak, A.; Froment, P.; et al. Mechanisms of Adiponectin Action in Fertility: An. Overview from Gametogenesis to Gestation in Humans and Animal Models in Normal and Pathological Conditions. Int. J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef]
- Guvenc, Y.; Var, A.; Goker, A.; Kuscu, N.K. Assessment of serum chemerin, vaspin and omentin-1 levels in patients with polycystic ovary syndrome. J. Int. Med. Res. 2016, 44, 796–805. [Google Scholar] [CrossRef] [Green Version]
- Farshchian, F.; Ramezani Tehrani, F.; Amirrasouli, H.; Rahimi Pour, H.; Hedayati, M.; Kazerouni, F.; Soltani, A. Visfatin and resistin serum levels in normal-weight and obese women with polycystic ovary syndrome. Int. J. Endocrinol. Metab. 2014, 12, e15503. [Google Scholar] [CrossRef]
- Akbarzadeh, S.; Ghasemi, S.; Kalantarhormozi, M.; Nabipour, I.; Abbasi, F.; Aminfar, A.; Jaffari, S.M.; Motamed, N.; Movahed, A.; Mirzaei, M.; et al. Relationship among plasma adipokines, insulin and androgens level as well as biochemical glycemic and lipidemic markers with incidence of PCOS in women with normal BMI. Gynecol. Endocrinol. 2012, 28, 521–524. [Google Scholar] [CrossRef]
- Kim, J.J.; Choi, Y.M.; Hong, M.A.; Kim, M.J.; Chae, S.J.; Kim, S.M.; Hwang, K.R.; Yoon, S.H.; Ku, S.Y.; Suh, C.S.; et al. Serum visfatin levels in non-obese women with polycystic ovary syndrome and matched controls. Obstet. Gynecol. Sci. 2018, 61, 253–260. [Google Scholar] [CrossRef]
- Tao, T.; Xu, B.; Liu, W. Ovarian HMW adiponectin is associated with folliculogenesis in women with polycystic ovary syndrome. Reprod. Biol. Endocrinol. 2013, 11, 99. [Google Scholar] [CrossRef]
- Tsouma, I.; Kouskouni, E.; Demeridou, S.; Boutsikou, M.; Hassiakos, D.; Chasiakou, A.; Hassiakou, S.; Baka, S. Correlation of visfatin levels and lipoprotein lipid profiles in women with polycystic ovary syndrome undergoing ovarian stimulation. Gynecol. Endocrinol. 2014, 30, 516–519. [Google Scholar] [CrossRef]
- Seow, K.M.; Juan, C.C.; Hsu, Y.P.; Ho, L.T.; Wang, Y.Y.; Hwang, J.L. Serum and follicular resistin levels in women with polycystic ovarian syndrome during IVF-stimulated cycles. Hum. Reprod. 2005, 20, 117–121. [Google Scholar] [CrossRef] [Green Version]
- Practice Committee of the American Society for Reproductive Medicine. Prevention and treatment of moderate and severe ovarian hyperstimulation syndrome: A guideline. Fertil. Steril. 2016, 106, 1634–1647. [Google Scholar] [CrossRef]
- Li, S.; Huang, X.; Zhong, H.; Peng, Q.; Chen, S.; Xie, Y.; Qin, X.; Qin, A. Low circulating adiponectin levels in women with polycystic ovary syndrome: An updated meta-analysis. Tumour. Biol. 2014, 35, 3961–3973. [Google Scholar] [CrossRef]
- Artimani, T.; Saidijam, M.; Aflatoonian, R.; Ashrafi, M.; Amiri, I.; Yavangi, M.; SoleimaniAsl, S.; Shabab, N.; Karimi, J.; Mehdizadeh, M. Downregulation of adiponectin system in granulosa cells and low levels of HMW adiponectin in PCOS. J. Assist. Reprod. Genet. 2016, 33, 101–110. [Google Scholar] [CrossRef]
- O’Connor, A.; Phelan, N.; Tun, T.K.; Boran, G.; Gibney, J.; Roche, H.M. High-molecular-weight adiponectin is selectively reduced in women with polycystic ovary syndrome independent of body mass index and severity of insulin resistance. J. Clin. Endocrinol. Metab. 2010, 95, 1378–1385. [Google Scholar] [CrossRef]
- Campos, D.B.; Palin, M.F.; Bordignon, V.; Murphy, B.D. The ‘beneficial’ adipokines in reproduction and fertility. Intj. Obes. (Lond.) 2008, 32, 223–231. [Google Scholar] [CrossRef]
- Patel, L.; Buckels, A.C.; Kinghorn, I.J.; Murdock, P.R.; Holbrook, J.D.; Plumpton, C.; Macphee, C.H.; Smith, S.A. Resistin is expressed in human macrophages and directly regulated by PPAR gamma activators. Biochem. Biophys. Res. Commun. 2003, 300, 472–476. [Google Scholar] [CrossRef]
- Fain, J.N.; Cheema, P.S.; Bahouth, S.W.; Lloyd Hiler, M. Resistin release by human adipose tissue explants in primary culture. Biochem. Biophys. Res. Commun. 2003, 300, 674–678. [Google Scholar] [CrossRef]
- Panidis, D.; Koliakos, G.; Kourtis, A.; Farmakiotis, D.; Mouslech, T.; Rousso, D. Serum resistin levels in women with polycystic ovary syndrome. Fertil. Steril. 2004, 81, 361–366. [Google Scholar] [CrossRef]
- Lee, J.H.; Chan, J.L.; Yiannakouris, N.; Kontogianni, M.; Estrada, E.; Seip, R.; Orlova, C.; Mantzoros, C.S. Circulating resistin levels are not associated with obesity or insulin resistance in humans and are not regulated by fasting or leptin administration: Cross-sectional and interventional studies in normal, insulin-resistant, and diabetic subjects. J. Clin. Endocrinol. Metab. 2003, 88, 4848–4856. [Google Scholar] [CrossRef]
- Seow, K.M.; Juan, C.C.; Hsu, Y.P.; Hwang, J.L.; Huang, L.W.; Ho, L.T. Amelioration of insulin resistance in women with PCOS via reduced insulin receptor substrate-1 Ser312 phosphorylation following laparoscopic ovarian electrocautery. Hum. Reprod. 2007, 22, 1003–1010. [Google Scholar] [CrossRef] [Green Version]
- Seow, K.M.; Juan, C.C.; Wu, L.Y.; Hsu, Y.P.; Yang, W.M.; Tsai, Y.L.; Hwang, J.L.; Ho, L.T. Serum and adipocyte resistin in polycystic ovary syndrome with insulin resistance. Hum. Reprod. 2004, 19, 48–53. [Google Scholar] [CrossRef] [Green Version]
- Samal, B.; Sun, Y.; Stearns, G.; Xie, C.; Suggs, S.; McNiece, I. Cloning and characterization of the cDNA encoding a novel human pre-B-cell colony-enhancing factor. Mol. Cell. Biol. 1994, 14, 1431–1437. [Google Scholar] [CrossRef]
- Fukuhara, A.; Matsuda, M.; Nishizawa, M.; Segawa, K.; Tanaka, M.; Kishimoto, K.; Matsuki, Y.; Murakami, M.; Ichisaka, T.; Murakami, H.; et al. Visfatin: A protein secreted by visceral fat that mimics the effects of insulin. Science 2005, 307, 426–430. [Google Scholar] [CrossRef]
- Berndt, J.; Kloting, N.; Kralisch, S.; Kovacs, P.; Fasshauer, M.; Schon, M.R.; Stumvoll, M.; Bluher, M. Plasma visfatin concentrations and fat depot-specific mRNA expression in humans. Diabetes 2005, 54, 2911–2916. [Google Scholar] [CrossRef]
- Chen, M.P.; Chung, F.M.; Chang, D.M.; Tsai, J.C.; Huang, H.F.; Shin, S.J.; Lee, Y.J. Elevated plasma level of visfatin/pre-B cell colony-enhancing factor in patients with type 2 diabetes mellitus. J. Clin. Endocrinol. Metab. 2006, 91, 295–299. [Google Scholar] [CrossRef]
- Shen, C.J.; Tsai, E.M.; Lee, J.N.; Chen, Y.L.; Lee, C.H.; Chan, T.F. The concentrations of visfatin in the follicular fluids of women undergoing controlled ovarian stimulation are correlated to the number of oocytes retrieved. Fertil. Steril. 2010, 93, 1844–1850. [Google Scholar] [CrossRef]
- Plati, E.; Kouskouni, E.; Malamitsi-Puchner, A.; Boutsikou, M.; Kaparos, G.; Baka, S. Visfatin and leptin levels in women with polycystic ovaries undergoing ovarian stimulation. Fertil. Steril. 2010, 94, 1451–1456. [Google Scholar] [CrossRef]
- Tan, B.K.; Chen, J.; Digby, J.E.; Keay, S.D.; Kennedy, C.R.; Randeva, H.S. Increased visfatin messenger ribonucleic acid and protein levels in adipose tissue and adipocytes in women with polycystic ovary syndrome: Parallel increase in plasma visfatin. J. Clin. Endocrinol. Metab. 2006, 91, 5022–5028. [Google Scholar] [CrossRef]
- Choi, K.H.; Joo, B.S.; Sun, S.T.; Park, M.J.; Son, J.B.; Joo, J.K.; Lee, K.S. Administration of visfatin during superovulation improves developmental competency of oocytes and fertility potential in aged female mice. Fertil. Steril. 2012, 97, 1234–1241. [Google Scholar] [CrossRef]
- Tatemoto, K.; Hosoya, M.; Habata, Y.; Fujii, R.; Kakegawa, T.; Zou, M.X.; Kawamata, Y.; Fukusumi, S.; Hinuma, S.; Kitada, C.; et al. Isolation and characterization of a novel endogenous peptide ligand for the human APJ receptor. Biochem. Biophys. Res. Commun. 1998, 251, 471–476. [Google Scholar] [CrossRef]
- Boucher, J.; Masri, B.; Daviaud, D.; Gesta, S.; Guigne, C.; Mazzucotelli, A.; Castan-Laurell, I.; Tack, I.; Knibiehler, B.; Carpene, C.; et al. Apelin, a newly identified adipokine up-regulated by insulin and obesity. Endocrinology 2005, 146, 1764–1771. [Google Scholar] [CrossRef]
- Chang, Y.H.; Chang, D.M.; Lin, K.C.; Shin, S.J.; Lee, Y.J. Visfatin in overweight/obesity, type 2 diabetes mellitus, insulin resistance, metabolic syndrome and cardiovascular diseases: A meta-analysis and systemic review. Diabetes Metab. Res. Rev. 2011, 27, 515–527. [Google Scholar] [CrossRef]
- Rak, A.; Drwal, E.; Rame, C.; Knapczyk-Stwora, K.; Slomczynska, M.; Dupont, J.; Gregoraszczuk, E.L. Expression of apelin and apelin receptor (APJ) in porcine ovarian follicles and in vitro effect of apelin on steroidogenesis and proliferation through APJ activation and different signaling pathways. Theriogenology 2017, 96, 126–135. [Google Scholar] [CrossRef]
- Shirasuna, K.; Shimizu, T.; Sayama, K.; Asahi, T.; Sasaki, M.; Berisha, B.; Schams, D.; Miyamoto, A. Expression and localization of apelin and its receptor APJ in the bovine corpus luteum during the estrous cycle and prostaglandin F2alpha-induced luteolysis. Reproduction 2008, 135, 519–525. [Google Scholar] [CrossRef]
- Roche, J.; Rame, C.; Reverchon, M.; Mellouk, N.; Rak, A.; Froment, P.; Dupont, J. Apelin (APLN) regulates progesterone secretion and oocyte maturation in bovine ovarian cells. Reproduction 2017, 153, 589–603. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, T.; Kosaka, N.; Murayama, C.; Tetsuka, M.; Miyamoto, A. Apelin and APJ receptor expression in granulosa and theca cells during different stages of follicular development in the bovine ovary: Involvement of apoptosis and hormonal regulation. Anim. Reprod. Sci. 2009, 116, 28–37. [Google Scholar] [CrossRef]
- Franks, S.; Stark, J.; Hardy, K. Follicle dynamics and anovulation in polycystic ovary syndrome. Hum. Reprod. Update 2008, 14, 367–378. [Google Scholar] [CrossRef]
- Taheri, S.; Murphy, K.; Cohen, M.; Sujkovic, E.; Kennedy, A.; Dhillo, W.; Dakin, C.; Sajedi, A.; Ghatei, M.; Bloom, S. The effects of centrally administered apelin-13 on food intake, water intake and pituitary hormone release in rats. Biochem. Biophys. Res. Commun. 2002, 291, 1208–1212. [Google Scholar] [CrossRef]
- Altinkaya, S.O.; Nergiz, S.; Kucuk, M.; Yuksel, H. Apelin levels in relation with hormonal and metabolic profile in patients with polycystic ovary syndrome. Eurj. Obstet. Gynecol. Reprod. Biol. 2014, 176, 168–172. [Google Scholar] [CrossRef]
- Olszanecka-Glinianowicz, M.; Madej, P.; Nylec, M.; Owczarek, A.; Szanecki, W.; Skalba, P.; Chudek, J. Circulating apelin level in relation to nutritional status in polycystic ovary syndrome and its association with metabolic and hormonal disturbances. Clin. Endocrinol. (Oxf) 2013, 79, 238–242. [Google Scholar] [CrossRef]
- Yang, R.Z.; Lee, M.J.; Hu, H.; Pray, J.; Wu, H.B.; Hansen, B.C.; Shuldiner, A.R.; Fried, S.K.; McLenithan, J.C.; Gong, D.W. Identification of omentin as a novel depot-specific adipokine in human adipose tissue: Possible role in modulating insulin action. Am. J. Physiol. Endocrinol. Metab. 2006, 290, E1253–E1261. [Google Scholar] [CrossRef]
- Mahde, A.; Shaker, M.; Al-Mashhadani, Z. Study of Omentin1 and Other Adipokines and Hormones in PCOS Patients. Oman. Med. J. 2009, 24, 108–118. [Google Scholar] [CrossRef]
- Choi, J.H.; Rhee, E.J.; Kim, K.H.; Woo, H.Y.; Lee, W.Y.; Sung, K.C. Plasma omentin-1 levels are reduced in non-obese women with normal glucose tolerance and polycystic ovary syndrome. Eur. J. Endocrinol. 2011, 165, 789–796. [Google Scholar] [CrossRef]
- Moreno-Navarrete, J.M.; Catalan, V.; Ortega, F.; Gomez-Ambrosi, J.; Ricart, W.; Fruhbeck, G.; Fernandez-Real, J.M. Circulating omentin concentration increases after weight loss. Nutr. Metab. (Lond) 2010, 7, 27. [Google Scholar] [CrossRef]
- Tan, B.K.; Pua, S.; Syed, F.; Lewandowski, K.C.; O’Hare, J.P.; Randeva, H.S. Decreased plasma omentin-1 levels in Type 1 diabetes mellitus. Diabet. Med. 2008, 25, 1254–1255. [Google Scholar] [CrossRef]
- de Souza Batista, C.M.; Yang, R.Z.; Lee, M.J.; Glynn, N.M.; Yu, D.Z.; Pray, J.; Ndubuizu, K.; Patil, S.; Schwartz, A.; Kligman, M.; et al. Omentin plasma levels and gene expression are decreased in obesity. Diabetes 2007, 56, 1655–1661. [Google Scholar] [CrossRef]
- Spritzer, P.M.; Lecke, S.B.; Satler, F.; Morsch, D.M. Adipose tissue dysfunction, adipokines, and low-grade chronic inflammation in polycystic ovary syndrome. Reproduction 2015, 149, R219–R227. [Google Scholar] [CrossRef] [Green Version]
- De Henau, O.; Degroot, G.N.; Imbault, V.; Robert, V.; De Poorter, C.; McHeik, S.; Gales, C.; Parmentier, M.; Springael, J.Y. Signaling Properties of Chemerin Receptors CMKLR1, GPR1 and CCRL2. PLoS ONE 2016, 11, e0164179. [Google Scholar] [CrossRef]
- Wittamer, V.; Franssen, J.D.; Vulcano, M.; Mirjolet, J.F.; Le Poul, E.; Migeotte, I.; Brezillon, S.; Tyldesley, R.; Blanpain, C.; Detheux, M.; et al. Specific recruitment of antigen-presenting cells by chemerin, a novel processed ligand from human inflammatory fluids. J. Exp. Med. 2003, 198, 977–985. [Google Scholar] [CrossRef]
- Bozaoglu, K.; Bolton, K.; McMillan, J.; Zimmet, P.; Jowett, J.; Collier, G.; Walder, K.; Segal, D. Chemerin is a novel adipokine associated with obesity and metabolic syndrome. Endocrinology 2007, 148, 4687–4694. [Google Scholar] [CrossRef]
- Goralski, K.B.; McCarthy, T.C.; Hanniman, E.A.; Zabel, B.A.; Butcher, E.C.; Parlee, S.D.; Muruganandan, S.; Sinal, C.J. Chemerin, a novel adipokine that regulates adipogenesis and adipocyte metabolism. J. Biol. Chem. 2007, 282, 28175–28188. [Google Scholar] [CrossRef]
- Kort, D.H.; Kostolias, A.; Sullivan, C.; Lobo, R.A. Chemerin as a marker of body fat and insulin resistance in women with polycystic ovary syndrome. Gynecol. Endocrinol. 2015, 31, 152–155. [Google Scholar] [CrossRef]
- Tsatsanis, C.; Dermitzaki, E.; Avgoustinaki, P.; Malliaraki, N.; Mytaras, V.; Margioris, A.N. The impact of adipose tissue-derived factors on the hypothalamic-pituitary-gonadal (HPG) axis. Horm. (Athens) 2015, 14, 549–562. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.Y.; Xue, K.; Cao, M.; Wang, Q.; Liu, J.Y.; Leader, A.; Han, J.Y.; Tsang, B.K. Chemerin suppresses ovarian follicular development and its potential involvement in follicular arrest in rats treated chronically with dihydrotestosterone. Endocrinology 2013, 154, 2912–2923. [Google Scholar] [CrossRef]
- Lima, P.D.A.; Nivet, A.L.; Wang, Q.; Chen, Y.A.; Leader, A.; Cheung, A.; Tzeng, C.R.; Tsang, B.K. Polycystic ovary syndrome: Possible involvement of androgen-induced, chemerin-mediated ovarian recruitment of monocytes/macrophages. Biol. Reprod. 2018, 99, 838–852. [Google Scholar] [CrossRef]
- Dunaif, A. Insulin resistance and the polycystic ovary syndrome: Mechanism and implications for pathogenesis. Endocr. Rev. 1997, 18, 774–800. [Google Scholar] [CrossRef]
- Panidis, D.; Tziomalos, K.; Misichronis, G.; Papadakis, E.; Betsas, G.; Katsikis, I.; Macut, D. Insulin resistance and endocrine characteristics of the different phenotypes of polycystic ovary syndrome: A prospective study. Hum. Reprod. 2012, 27, 541–549. [Google Scholar] [CrossRef]
- Dewailly, D.; Pigny, P.; Soudan, B.; Catteau-Jonard, S.; Decanter, C.; Poncelet, E.; Duhamel, A. Reconciling the definitions of polycystic ovary syndrome: The ovarian follicle number and serum anti-Mullerian hormone concentrations aggregate with the markers of hyperandrogenism. J. Clin. Endocrinol. Metab. 2010, 95, 4399–4405. [Google Scholar] [CrossRef]
- Dewailly, D.; Lujan, M.E.; Carmina, E.; Cedars, M.I.; Laven, J.; Norman, R.J.; Escobar-Morreale, H.F. Definition and significance of polycystic ovarian morphology: A task force report from the Androgen Excess and Polycystic Ovary Syndrome Society. Hum. Reprod. Update 2014, 20, 334–352. [Google Scholar] [CrossRef]
- Pigny, P.; Merlen, E.; Robert, Y.; Cortet-Rudelli, C.; Decanter, C.; Jonard, S.; Dewailly, D. Elevated serum level of anti-mullerian hormone in patients with polycystic ovary syndrome: Relationship to the ovarian follicle excess and to the follicular arrest. J. Clin. Endocrinol. Metab. 2003, 88, 5957–5962. [Google Scholar] [CrossRef]
- Teede, H.; Misso, M.; Tassone, E.C.; Dewailly, D.; Ng, E.H.; Azziz, R.; Norman, R.J.; Andersen, M.; Franks, S.; Hoeger, K.; et al. Anti-Mullerian Hormone in PCOS: A Review Informing International Guidelines. Trends Endocrinol. Metab. 2019, 30, 467–478. [Google Scholar] [CrossRef]
- Victoria, M.; Labrosse, J.; Krief, F.; Cedrin-Durnerin, I.; Comtet, M.; Grynberg, M. Anti Mullerian Hormone: More than a biomarker of female reproductive function. J. Gynecol. Obstet. Hum. Reprod. 2019, 48, 19–24. [Google Scholar] [CrossRef]
- Pierre, A.; Peigne, M.; Grynberg, M.; Arouche, N.; Taieb, J.; Hesters, L.; Gonzales, J.; Picard, J.Y.; Dewailly, D.; Fanchin, R.; et al. Loss of LH-induced down-regulation of anti-Mullerian hormone receptor expression may contribute to anovulation in women with polycystic ovary syndrome. Hum. Reprod. 2013, 28, 762–769. [Google Scholar] [CrossRef] [Green Version]
- Homburg, R.; Crawford, G. The role of AMH in anovulation associated with PCOS: A hypothesis. Hum. Reprod. 2014, 29, 1117–1121. [Google Scholar] [CrossRef]
- Cimino, I.; Casoni, F.; Liu, X.; Messina, A.; Parkash, J.; Jamin, S.P.; Catteau-Jonard, S.; Collier, F.; Baroncini, M.; Dewailly, D.; et al. Novel role for anti-Mullerian hormone in the regulation of GnRH neuron excitability and hormone secretion. Nat. Commun. 2016, 7, 10055. [Google Scholar] [CrossRef]
- Guerif, F.; Bidault, R.; Gasnier, O.; Couet, M.L.; Gervereau, O.; Lansac, J.; Royere, D. Efficacy of blastocyst transfer after implantation failure. Reprod. Biomed. Online 2004, 9, 630–636. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, RESEARCH0034. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Age (y) | BMI (kg/m2) | Cycle Duration (d) | FSH (UI/L) | Estradiol (ng/L) | Testosterone (µg/L) |
|---|---|---|---|---|---|---|
| NW Controls (n = 12) | 31.08 ± 3.82 | 21.77 ± 2.07 | 28.46 ± 2.15 | 6.78 ± 3.19 | 43.33 ± 14.39 | 0.33 ± 0.20 (n = 5) |
| NW ECHO (n = 13) | 31.69 ± 5.88 | 20.84 ± 1.86 | 30.25 ± 2.54 | 6.40 ± 2.15 | 42.85 ± 16.01 | 0.28 ± 0.15 (n = 6) |
| NW PCOS (n = 13) | 29.54 ± 3.36 | 20.68 ± 1.96 | 98.54 ± 67.77 * | 5.86 ± 1.66 | 41.55 ± 14.85 | 0.88 ± 0.93 (n = 6) |
| Obese Controls (n = 15) | 33.80 ± 4.62 | 33.13 ± 2.29 | 28.97 ± 2.21 | 6.71 ± 3.49 | 43.23 ± 17.24 | 0.41 ± 0.19 (n = 5) |
| Obese ECHO (n = 15) | 32.73 ± 4.50 | 31.53 ± 3.33 | 29.89 ± 1.30 | 5.68 ± 1.09 | 35.23 ± 13.43 | 0.43 ± 0.08 (n = 5) |
| Obese PCOS (n = 10) | 30.10 ± 3.25 | 33.38 ± 2.12 | 76.11 ± 61.79 # | 4.86 ± 1.64 | 34.13 ± 6.99 | 0.49 ± 0.24 (n = 7) |
| Condition Effect | p = 0.07 | p = 0.14 | p < 0.0001 | p = 0.13 | p = 0.38 | - |
| BMI Effect | p = 0.15 | p < 0.0001 | p = 0.37 | p = 0.28 | p = 0.13 | - |
| Interaction | p = 0.65 | p = 0.32 | p = 0.46 | p = 0.78 | p = 0.57 | - |
| Parameter | Adiponectin | Chemerin | Resistin | Visfatin | Omentin | Apelin |
|---|---|---|---|---|---|---|
| BMI (kg/m2) | r = −0.748 *** | r = 0.725 *** | r = 0.799 *** | r = 0.275 * | r = 0.446 *** | r = 0.441 *** |
| Cycle duration (d) | NS | NS | NS | NS | r = 0.421 *** | NS |
| Follicles Count (n) | NS | NS | NS | r = −0.352 ** | NS | r = 0.480* ** |
| AMH (ng/mL) | NS | NS | NS | r = −0.284 * | NS | NS |
| Estradiol (ng/L) | r = −0.300 * | NS | NS | NS | NS | r = −0.284 * |
| Oocytes Retrieved (n) | NS | NS | NS | r = −0.37** | NS | r = 0.300* |
| Embryos (n) | NS | NS | NS | r = −0.262* | NS | r = 0.268* |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bongrani, A.; Mellouk, N.; Rame, C.; Cornuau, M.; Guérif, F.; Froment, P.; Dupont, J. Ovarian Expression of Adipokines in Polycystic Ovary Syndrome: A Role for Chemerin, Omentin, and Apelin in Follicular Growth Arrest and Ovulatory Dysfunction? Int. J. Mol. Sci. 2019, 20, 3778. https://doi.org/10.3390/ijms20153778
Bongrani A, Mellouk N, Rame C, Cornuau M, Guérif F, Froment P, Dupont J. Ovarian Expression of Adipokines in Polycystic Ovary Syndrome: A Role for Chemerin, Omentin, and Apelin in Follicular Growth Arrest and Ovulatory Dysfunction? International Journal of Molecular Sciences. 2019; 20(15):3778. https://doi.org/10.3390/ijms20153778
Chicago/Turabian StyleBongrani, Alice, Namya Mellouk, Christelle Rame, Marion Cornuau, Fabrice Guérif, Pascal Froment, and Joëlle Dupont. 2019. "Ovarian Expression of Adipokines in Polycystic Ovary Syndrome: A Role for Chemerin, Omentin, and Apelin in Follicular Growth Arrest and Ovulatory Dysfunction?" International Journal of Molecular Sciences 20, no. 15: 3778. https://doi.org/10.3390/ijms20153778
APA StyleBongrani, A., Mellouk, N., Rame, C., Cornuau, M., Guérif, F., Froment, P., & Dupont, J. (2019). Ovarian Expression of Adipokines in Polycystic Ovary Syndrome: A Role for Chemerin, Omentin, and Apelin in Follicular Growth Arrest and Ovulatory Dysfunction? International Journal of Molecular Sciences, 20(15), 3778. https://doi.org/10.3390/ijms20153778