JA-Induced Endocytosis of AtRGS1 Is Involved in G-Protein Mediated JA Responses



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
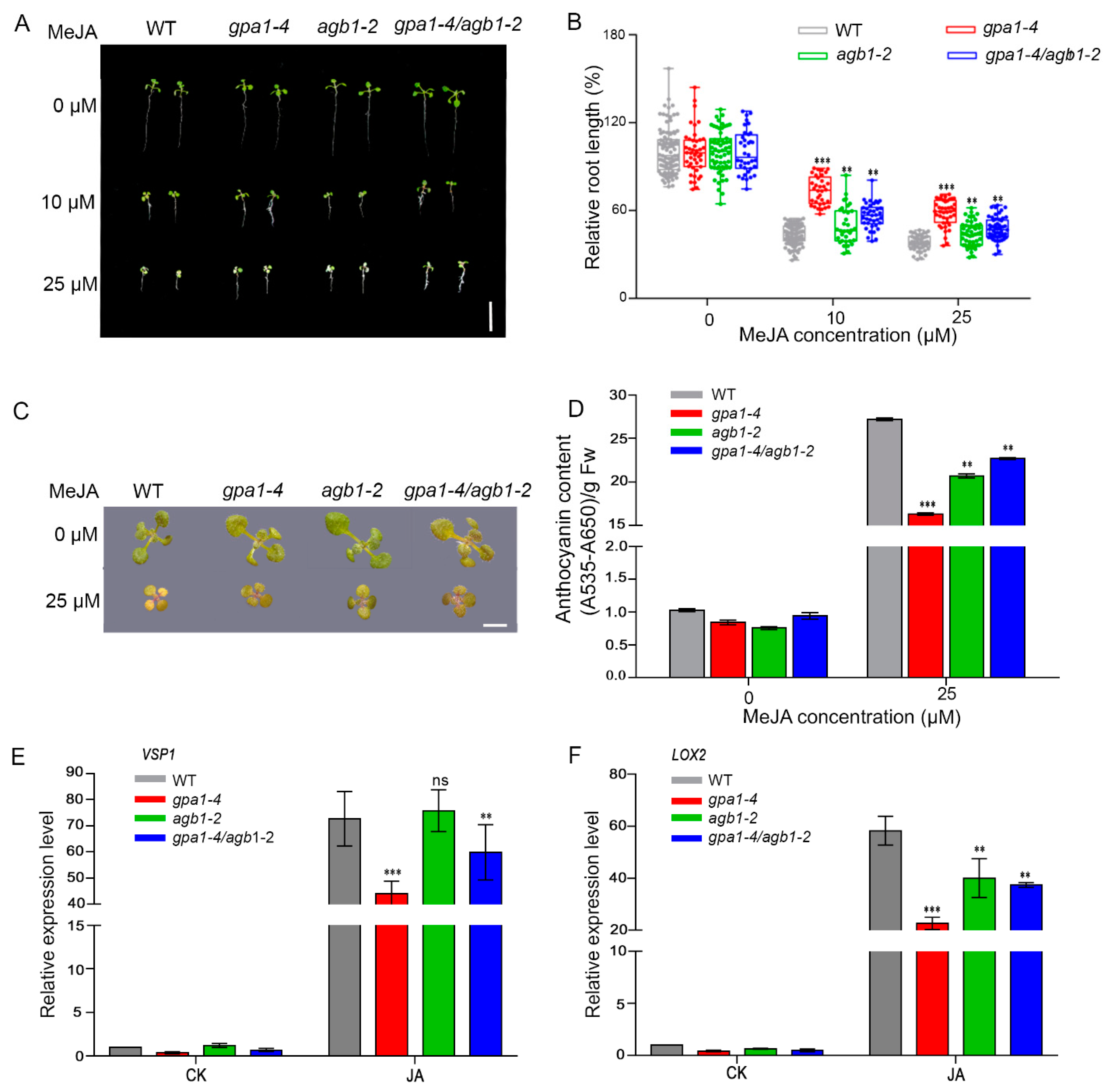
2.1. The Heterotrimeric G Protein Complex Is Involved in JA Signaling
2.2. JA Induces AtRGS1 Endocytosis and Dissociation from AtGPA1
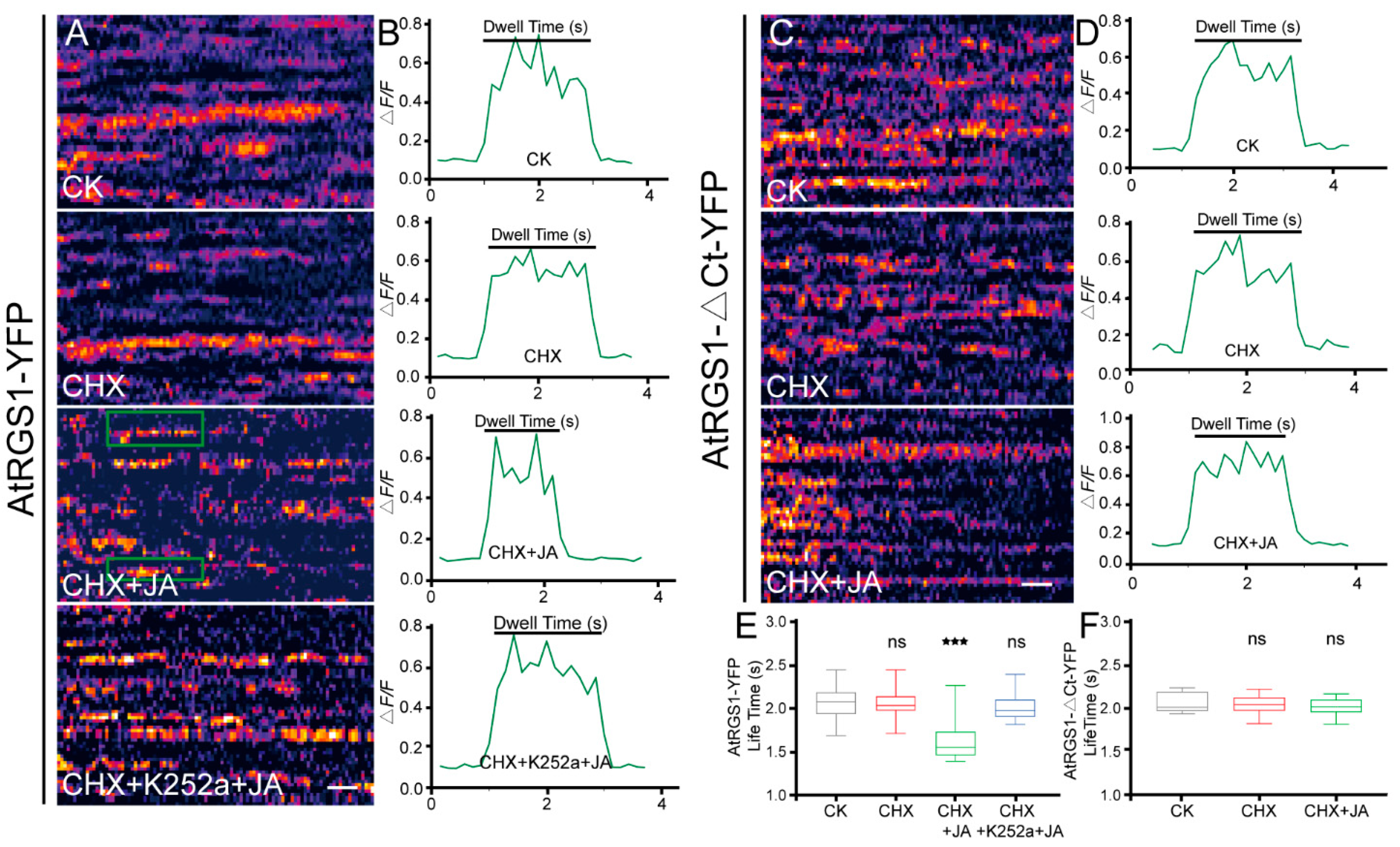
2.3. The Phosphorylation and C-Terminal Domain Are Required for JA-Induced AtRGS1 Endocytosis
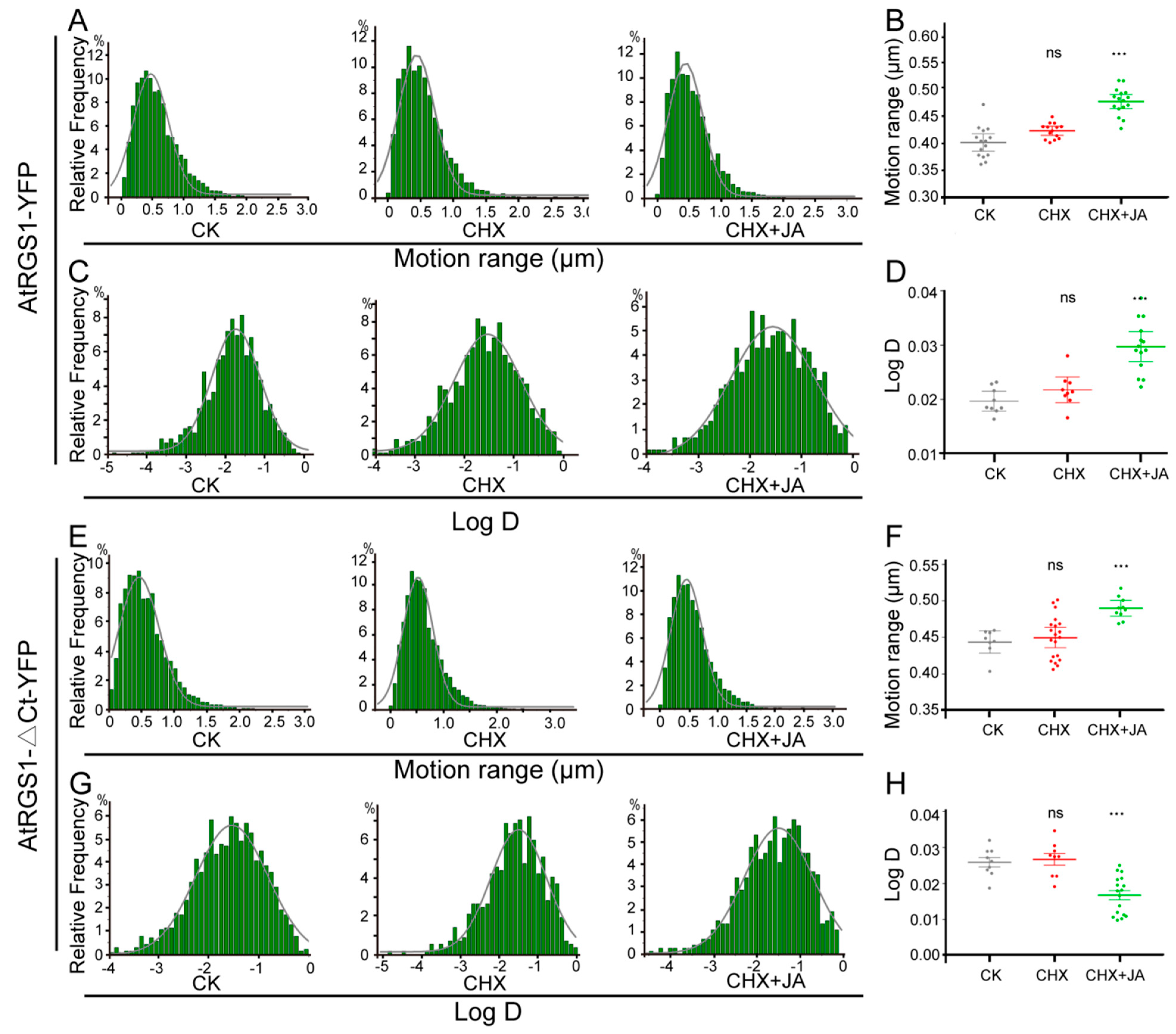
2.4. JA Treatment Affects PM Dynamics of AtRGS1 and AtRGS1-ΔCt
3. Materials and Methods
3.1. Plant Materials and Growth Conditions
3.2. Anthocyanin Measurements
3.3. Root Length Measurement
3.4. Quantitative Real-Time PCR Analysis
3.5. Transient Infiltration of N. benthamiana Leaves
3.6. Drug Treatment
3.7. Confocal Microscopy
3.8. VA-TIRFM and Fluorescence Image Analysis
3.9. FLIM-FRET Measurement
3.10. Western Blot Analysis
3.11. Data Analysis
3.12. Accession Numbers
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wasternack, C.; Song, S. Jasmonates: Biosynthesis, metabolism and signalling by proteins activating and repressing transcription. J. Exp. Bot. 2017, 6, 1303–1321. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Liu, B.; Liu, L.; Song, S. Jasmonate action in plant growth and development. J. Exp. Bot. 2017, 68, 1349–1359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shan, X.; Yan, J.; Xie, D. Comparison of phytohormone signalling mechanisms. Curr. Opin. Plant Biol. 2012, 15, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Gilman, A.G. G proteins: Transducers of receptor-generated signals. Annu. Rev. Biochem. 1987, 56, 615–649. [Google Scholar] [CrossRef] [PubMed]
- Wettschureck, N.; Offermanns, S. Mammalian G proteins and their cell type specific functions. Physiol. Rev. 2005, 85, 1159–1204. [Google Scholar] [CrossRef] [PubMed]
- Lefkowitz, R.J. Historical review: A brief history and personal retrospective of seven-transmembrane receptors. Trends Pharmacol. 2004, 28, 413–422. [Google Scholar] [CrossRef]
- Ma, H.; Yanofsky, M.F.; Meyerowitz, E.M. Molecular cloning and characterization of GPA1, a G protein alpha subunit gene from Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 1990, 87, 3821–3825. [Google Scholar] [CrossRef]
- Weiss, C.A.; Garnaat, C.W.; Mukai, K.; Hu, Y.; Ma, H. Isolation of cDNAs encoding guanine nucleotidebinding protein beta-subunit homologues from maize (ZGB1) and Arabidopsis (AGB1). Proc. Natl. Acad. Sci. USA 1994, 91, 9554–9558. [Google Scholar] [CrossRef]
- Mason, M.G.; Botella, J.R. Completing the heterotrimer: Isolation and characterization of an Arabidopsis thaliana G protein gamma-subunit cDNA. Proc. Natl. Acad. Sci. USA 2000, 97, 14784–14788. [Google Scholar] [CrossRef]
- Mason, M.G.; Botella, J.R. Isolation of a novel G-protein gamma-subunit from Arabidopsis thaliana and its interaction with Gβ. Biochim. Biophys. 2001, 1520, 147–153. [Google Scholar] [CrossRef]
- Chen, J.G.; Willard, F.S.; Huang, J.; Liang, J.; Chasse, S.A.; Jones, A.M.; Siderovski, D.P. A seventransmembrane RGS protein that modulates plant cell proliferation. Science 2003, 301, 1728–1731. [Google Scholar] [CrossRef] [PubMed]
- Johnston, C.A.; Taylor, J.P.; Gao, Y.; Kimple, A.J.; Grigston, J.C.; Chen, J.G.; Siderovski, D.P.; Jones, A.M.; Willard, F.S. GTPase acceleration as the rate-limiting step in Arabidopsis G protein-coupled sugar signalling. Proc. Natl. Acad. Sci. USA 2007, 104, 17317–17322. [Google Scholar] [CrossRef] [PubMed]
- Urano, D.; Chen, J.G.; Botella, J.R.; Jones, A.M. Heterotrimeric G protein signalling in the plant kingdom. Open Biol. 2013, 3, 120186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phan, N.; Urano, D.; Srba, M.; Fischer, L.; Jones, A.M. Sugar-induced endocytosis of plant 7TM-RGS proteins. Plant Signal. Behav. 2012, 8, e22814. [Google Scholar] [CrossRef] [PubMed]
- Urano, D.; Phan, N.; Jones, J.C.; Yang, J.; Huang, J.; Grigston, J.; Taylor, J.P.; Jones, A.M. Endocytosis of the seven-transmembrane RGS1 protein activates G-protein-coupled signalling in Arabidopsis. Nat. Cell Biol. 2012, 14, 1079–1088. [Google Scholar] [CrossRef] [PubMed]
- Tunc-Ozdemir, M.; Urano, D.; Jaiswal, D.K.; Clouse, S.D.; Jones, A.M. Direct modulation of heterotrimeric G protein-coupled signalling by a receptor kinase complex. J. Biol. Chem. 2016, 291, 13918–13925. [Google Scholar] [CrossRef]
- Tunc-Ozdemir, M.; Jones, A.M. Ligand-induced dynamics of heterotrimeric G protein-coupled receptor-like kinase complexes. PLoS ONE 2017, 12, e0171854. [Google Scholar] [CrossRef]
- Liang, X.; Ding, P.; Lian, K.; Wang, J.; Ma, M.; Li, L.; Li, L.; Li, M.; Zhang, X.; Chen, S.; et al. Arabidopsis heterotrimeric G proteins regulate immunity by directly coupling to the FLS2 receptor. eLife 2016, 5, e13568. [Google Scholar]
- Liang, X.; Ma, M.; Zhou, Z.; Wang, J.; Yang, X.; Rao, S.; Zhou, J.M. Ligand-triggered de-repression of Arabidopsis heterotrimeric G proteins coupled to immune receptor kinases. Cell Res. 2018, 28, 529–543. [Google Scholar] [CrossRef]
- Colaneri, A.C.; Tunc-Ozdemir, M.; Huang, J.; Jones, A.M. Growth attenuation under saline stress is mediated by the heterotrimeric G protein complex. BMC Plant Biol. 2014, 14, 129. [Google Scholar] [CrossRef]
- Wang, L.; Xue, Y.; Xing, J.; Song, K.; Lin, J. Exploring the spatiotemporal organization of membrane proteins in living plant cells. Annu. Rev. Plant Biol. 2018, 69, 525–551. [Google Scholar] [CrossRef] [PubMed]
- Shan, X.; Zhang, Y.; Peng, W.; Wang, Z.; Xie, D. Molecular mechanism for jasmonate-induction of anthocyanin accumulation in Arabidopsis. J. Exp. Bot. 2009, 60, 3849–3860. [Google Scholar] [CrossRef] [PubMed]
- Jaqaman, K.; Loerke, D.; Mettlen, M.; Kuwata, H.; Grinstein, S.; Schmid, S.L. Danuser G Robust single-particle tracking in live-cell time-lapse sequences. Nat. Methods 2008, 5, 695–702. [Google Scholar] [CrossRef] [PubMed]
- Eichel, K.; Jullié, D.; Zastrow, M.V. β-Arrestin drives MAP kinase signalling from clathrin-coated structures after GPCR dissociation. Nat. Cell Biol. 2016, 18, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Yu, M.; Yao, X.; Xing, J.; Lin, J.; Li, X. Single-particle tracking for the quantification of membrane protein dynamics in living plant cells. Mol. Plant 2018, 11, 1315–1321. [Google Scholar] [CrossRef] [PubMed]
- Long, Y.; Stahl, Y.; Weidtkamp-Peters, S.; Postma, M.; Zhou, W.; Goedhart, J.; Blilou, I. In vivo FRET–FLIM reveals cell-type-specific protein interactions in Arabidopsis roots. Nature 2017, 548, 97–102. [Google Scholar] [CrossRef] [PubMed]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, L.; Su, B.; Qi, X.; Zhang, X.; Song, S.; Shan, X. JA-Induced Endocytosis of AtRGS1 Is Involved in G-Protein Mediated JA Responses. Int. J. Mol. Sci. 2019, 20, 3779. https://doi.org/10.3390/ijms20153779
Li L, Su B, Qi X, Zhang X, Song S, Shan X. JA-Induced Endocytosis of AtRGS1 Is Involved in G-Protein Mediated JA Responses. International Journal of Molecular Sciences. 2019; 20(15):3779. https://doi.org/10.3390/ijms20153779
Chicago/Turabian StyleLi, Li, Bodan Su, Xueying Qi, Xi Zhang, Susheng Song, and Xiaoyi Shan. 2019. "JA-Induced Endocytosis of AtRGS1 Is Involved in G-Protein Mediated JA Responses" International Journal of Molecular Sciences 20, no. 15: 3779. https://doi.org/10.3390/ijms20153779
APA StyleLi, L., Su, B., Qi, X., Zhang, X., Song, S., & Shan, X. (2019). JA-Induced Endocytosis of AtRGS1 Is Involved in G-Protein Mediated JA Responses. International Journal of Molecular Sciences, 20(15), 3779. https://doi.org/10.3390/ijms20153779