A Long Polymorphic GT Microsatellite within a Gene Promoter Mediates Non-Imprinted Allele-Specific DNA Methylation of a CpG Island in a Goldfish Inter-Strain Hybrid


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
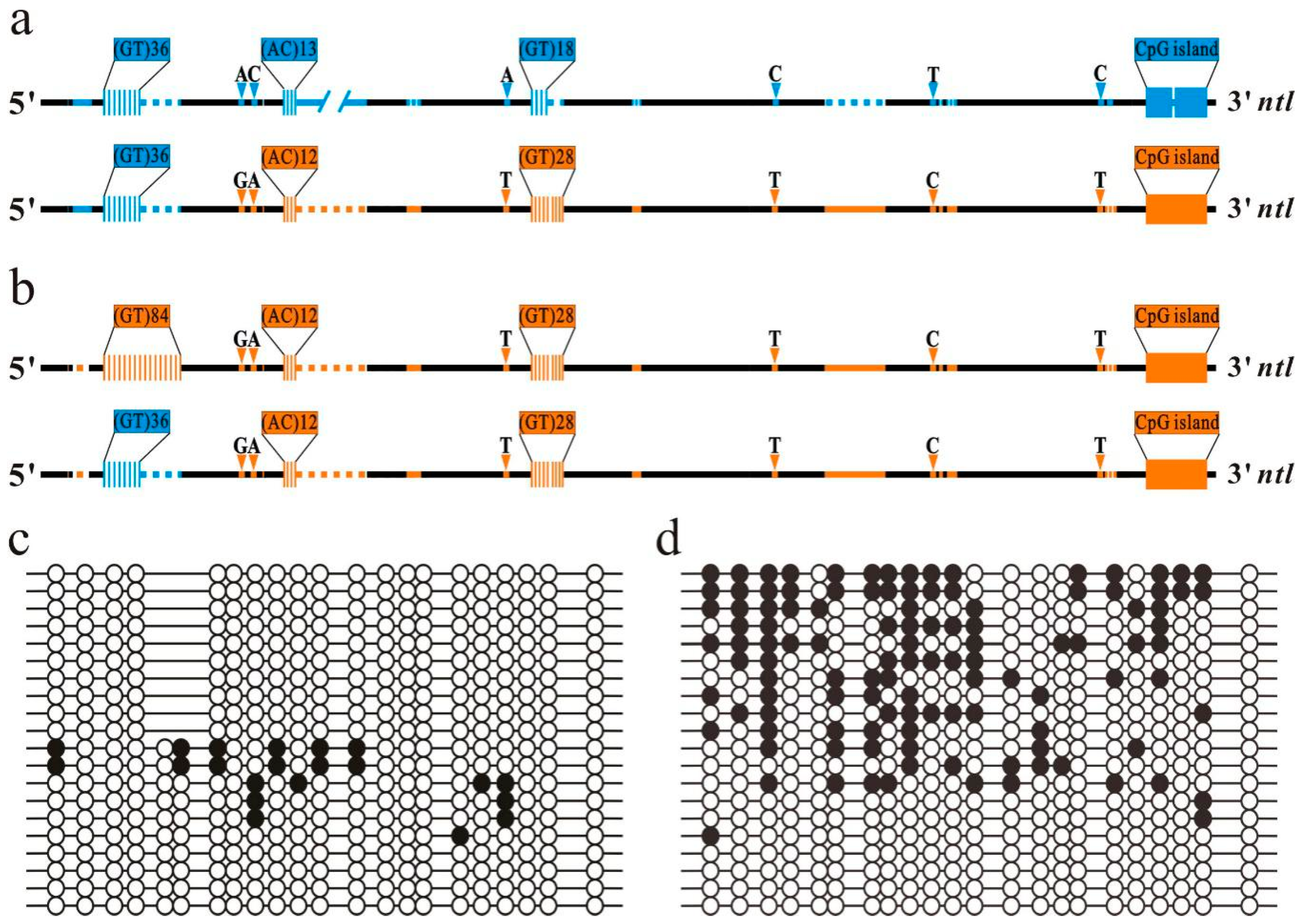
2. Results
2.1. Sequence Divergence within Upstream Region of the ntl Promoter Accompanying by Methylation Variation of Downstream CpG Island
2.2. Impact of Diversified Promoter Sequences on the Methylation of Downstream CpG Island in Hybrid
2.3. Homologous Recombination at ntl GT Microsatellite in the Eggs of the Hybrid
2.4. Length Effect of GTM 1 on Methylation of Downstream ntl CpG Island
2.5. ASM of the ntl CpG Island and ASE of ntl in Heterozygous Embryos
3. Discussion
4. Materials and Methods
4.1. Animals and Samples Preparation
4.2. Bisulfite Sequencing (BS) of the ntl CpG Island
4.3. Methylation-Specific PCR (MSP)
4.4. Measurements of Expression Differences between Chinese GF and Japanese GF ntl Alleles in the Heterozygous Embryos
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Weaver, J.R.; Susiarjo, M.; Bartolomei, M.S. Imprinting and epigenetic changes in the early embryo. Mamm. Genome 2009, 20, 532–543. [Google Scholar] [CrossRef]
- Reik, W.; Dean, W. DNA methylation and mammalian epigenetics. Electrophoresis 2001, 22, 2838–2843. [Google Scholar] [CrossRef]
- Kerkel, K.; Spadola, A.; Yuan, E.; Kosek, J.; Jiang, L.; Hod, E.; Li, K.; Murty, V.V.; Schupf, N.; Vilain, E.; et al. Genomic surveys by methylation-sensitive SNP analysis identify sequence-dependent allele-specific DNA methylation. Nat. Genet. 2008, 40, 904–908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Rohde, C.; Reinhardt, R.; Voelcker-Rehage, C.; Jeltsch, A. Non-imprinted allele-specific DNA methylation on human autosomes. Genome Biol. 2009, 10, R138. [Google Scholar] [CrossRef] [PubMed]
- Schalkwyk, L.C.; Meaburn, E.L.; Smith, R.; Dempster, E.L.; Jeffries, A.R.; Davies, M.N.; Plomin, R.; Mill, J. Allelic skewing of DNA methylation is widespread across the genome. Am. J. Hum. Genet. 2010, 86, 196–212. [Google Scholar] [CrossRef] [PubMed]
- Shoemaker, R.; Deng, J.; Wang, W.; Zhang, K. Allele-specific methylation is prevalent and is contributed by CpG-SNPs in the human genome. Genome Res. 2010, 20, 883–889. [Google Scholar] [CrossRef] [Green Version]
- Tycko, B. Allele-specific DNA methylation: Beyond imprinting. Hum. Mol. Genet. 2010, 19, R210–R220. [Google Scholar] [CrossRef]
- Schilling, E.; El, C.C.; Rehli, M. Allele-specific DNA methylation in mouse strains is mainly determined by cis-acting sequences. Genome Res. 2009, 19, 2028–2035. [Google Scholar] [CrossRef]
- Yang, H.H.; Hu, N.; Wang, C.; Ding, T.; Dunn, B.K.; Goldstein, A.M.; Taylor, P.R.; Lee, M.P. Influence of genetic background and tissue types on global DNA methylation patterns. PLoS ONE 2010, 5, e9355. [Google Scholar] [CrossRef]
- Hellman, A.; Chess, A. Extensive sequence-influenced DNA methylation polymorphism in the human genome. Epigenet. Chromatin 2010, 3, 1–11. [Google Scholar] [CrossRef]
- Qu, W.; Hashimoto, S.; Shimada, A.; Nakatani, Y.; Ichikawa, K.; Saito, T.L.; Ogoshi, K.; Matsushima, K.; Suzuki, Y.; Sugano, S.; et al. Genome-wide genetic variations are highly correlated with proximal DNA methylation patterns. Genome Res. 2012, 22, 1419–1425. [Google Scholar] [CrossRef]
- Lander, E.S.; Linton, L.M.; Birren, B.; Nusbaum, C.; Zody, M.C.; Baldwin, J.; Devon, K.; Dewar, K.; Doyle, M.; FitzHugh, W.; et al. Initial sequencing and analysis of the human genome. Nature 2001, 409, 860–921. [Google Scholar] [Green Version]
- Subramanian, S.; Mishra, R.K.; Singh, L. Genome-wide analysis of microsatellite repeats in humans: Their abundance and density in specific genomic regions. Genome Biol. 2003, 4, R13. [Google Scholar] [CrossRef]
- Ellegren, H. Microsatellites: Simple sequences with complex evolution. Nat. Rev. Genet. 2004, 5, 435–445. [Google Scholar] [CrossRef]
- Sun, J.X.; Helgason, A.; Masson, G.; Ebenesersd’ ottir, S.S.; Li, H.; Mallick, S.; Gnerre, S.; Patterson, N.; Kong, A.; Reich, D.; et al. A direct characterization of human mutation based on microsatellites. Nat. Genet. 2012, 44, 1161–1165. [Google Scholar] [CrossRef] [Green Version]
- Strand, M.; Prolla, T.A.; Liskay, R.M.; Petes, T.D. Destabilization of tracts of simple repetitive DNA in yeast by mutations affecting DNA mismatch repair. Nature 1993, 365, 274–276. [Google Scholar] [CrossRef]
- Paques, F.; Leung, W.Y.; Haber, J.E. Expansions and contractions in a tandem repeat induced by double-strand break repair. Mol. Cell. Biol. 1998, 18, 2045–2054. [Google Scholar] [CrossRef]
- Jankowski, C.; Nasar, F.; Nag, D.K. Meiotic instability of CAG repeat tracts occurs by double-strand break repair in yeast. Proc. Natl. Acad. Sci. USA 2000, 97, 2134–2139. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Bissler, J.J.; Sinden, R.R.; Leffak, M. Unstable spinocerebellar ataxia type 10 (ATTCT*(AGAAT) repeats are associated with aberrant replication at the ATX10 locus and replication origin-dependent expansion at an ectopic site in human cells. Mol. Cell. Biol. 2007, 27, 7828–7838. [Google Scholar] [CrossRef]
- Hardwick, R.J.; Tretyakov, M.V.; Dubrova, Y.E. Age-related accumulation of mutations supports a replication-dependent mechanism of spontaneous mutation at tandem repeat DNA loci in mice. Mol. Biol. Evol. 2009, 26, 2647–2654. [Google Scholar] [CrossRef]
- Gadgil, R.; Barthelemy, J.; Lewis, T.; Leffak, M. Replication stalling and DNA microsatellite instability. Biophys. Chem. 2017, 225, 38–48. [Google Scholar] [CrossRef]
- Warpeha, K.M.; Xu, W.; Liu, L.; Charles, I.G.; Patterson, C.C.; Ah-Fat, F.; Harding, S.; Hart, P.M.; Chakravarthy, U.; Hughes, A.E. Genotyping and functional analysis of a polymorphic (CCTTT)(n) repeat of NOS2A in diabetic retinopathy. FASEB J. 1999, 13, 1825–1832. [Google Scholar] [CrossRef]
- Contente, A.; Dittmer, A.; Koch, M.C.; Roth, J.; Dobbelstein, M. A polymorphic microsatellite that mediates induction of PIG3 by p53. Nat. Genet. 2002, 30, 315–320. [Google Scholar] [CrossRef]
- Quilez, J.; Guilmatre, A.; Garg, P.; Highnam, G.; Gymrek, M.; Erlich, Y.; Joshi, R.S.; Mittelman, D.; Sharp, A.J. Polymorphic tandem repeats within gene promoters act as modifiers of gene expression and DNA methylation in humans. Nucleic Acids Res. 2016, 44, 3750–3762. [Google Scholar] [CrossRef] [Green Version]
- Miesfeld, R.; Krystal, M.; Arnheim, N. A member of a new repeated sequence family which is conserved throughout eucaryotic evolution is found between the human delta and beta globin genes. Nucleic Acids Res. 1981, 9, 5931–5947. [Google Scholar] [CrossRef]
- Hamada, H.; Kakunaga, T. Potential Z-DNA forming sequences are highly dispersed in the human genome. Nature 1982, 298, 396–398. [Google Scholar] [CrossRef]
- Hamada, H.; Petrino, M.G.; Kakunaga, T. A novel repeated element with Z-DNA forming potential is widely found in evolutionarily diverse eukaryotic genomes. Proc. Natl. Acad. Sci. USA 1982, 79, 6465–6469. [Google Scholar] [CrossRef]
- Ma, S.; Huang, W.; Zhang, L.; Zhao, S.; Tong, Y.; Liu, Z.; Sun, L.; Chen, H.; Luo, C. Germ cell-specific DNA methylation and genome diploidization in primitive vertebrates. Epigenetics 2011, 6, 1471–1480. [Google Scholar] [CrossRef] [Green Version]
- Yamakoshi, K.; Shimoda, N. De novo DNA methylation at the CpG island of the zebrafish no tail gene. Genesis 2003, 37, 195–202. [Google Scholar] [CrossRef]
- Huang, W.; Zheng, J.; He, Y.; Luo, C. Tandem repeat modification during double strand break repair induced by an engineered TAL effector nuclease in zebrafish genome. PLoS ONE 2013, 8, e84176. [Google Scholar] [CrossRef]
- Majewski, J.; Ott, J. GT repeats are associated with recombinationon human chromosome 22. Genome Res. 2000, 10, 1108–1114. [Google Scholar] [CrossRef]
- Treco, D.; Arnheim, N. The evolutionarily conserved repetitive sequence d(TG.AC) promotes reciprocal exchange and generates unusual recombinant tetrads during yeast meiosis. Mol. Cell. Biol. 1986, 6, 3934–3947. [Google Scholar] [CrossRef]
- Grendrel, C.; Boulet, A.; Dutreix, M. (CA/GT)n microsatellites affect homologous recombination during yeast meiosis. Genes Dev. 2000, 14, 1261–11268. [Google Scholar]
- Panigrahi, G.B.; Lau, R.; Montgomery, S.E.; Leonard, M.R.; Pearson, C.E. Slipped (CTG)*(CAG) repeats can be correctly repaired, escape repair or undergo errorprone repair. Nat. Struct. Mol. Biol. 2005, 12, 654–662. [Google Scholar] [CrossRef]
- Axford, M.M.; Wang, Y.H.; Nakamori, M.; Zannis-Hadjopoulos, M.; Thornton, C.A.; Pearson, C.E. Detection of slipped-DNAs at the trinucleotide repeats of the myotonic dystrophy type I disease locus in patient tissues. PLoS Genet. 2013, 9, e1003866. [Google Scholar] [CrossRef]
- Slean, M.M.; Panigrahi, G.B.; Castel, A.L.; Pearson, A.B.; Tomkinson, A.E.; Pearson, C.E. Absence of MutSbeta leads to the formation of slipped-DNA for CTG/CAG contractions at primate replication forks. DNA Repair 2016, 42, 107–118. [Google Scholar] [CrossRef]
- Wahls, W.P.; Wallace, L.J.; Moore, P.D. The Z-DNA motif d(TG)30 promotes reception of information during gene conversion events while stimulating homologous recombination in human cells in culture. Mol. Cell. Biol. 1990, 10, 785–793. [Google Scholar] [CrossRef]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, J.; Xu, H.; Cao, H. A Long Polymorphic GT Microsatellite within a Gene Promoter Mediates Non-Imprinted Allele-Specific DNA Methylation of a CpG Island in a Goldfish Inter-Strain Hybrid. Int. J. Mol. Sci. 2019, 20, 3923. https://doi.org/10.3390/ijms20163923
Zheng J, Xu H, Cao H. A Long Polymorphic GT Microsatellite within a Gene Promoter Mediates Non-Imprinted Allele-Specific DNA Methylation of a CpG Island in a Goldfish Inter-Strain Hybrid. International Journal of Molecular Sciences. 2019; 20(16):3923. https://doi.org/10.3390/ijms20163923
Chicago/Turabian StyleZheng, Jianbo, Haomang Xu, and Huiwen Cao. 2019. "A Long Polymorphic GT Microsatellite within a Gene Promoter Mediates Non-Imprinted Allele-Specific DNA Methylation of a CpG Island in a Goldfish Inter-Strain Hybrid" International Journal of Molecular Sciences 20, no. 16: 3923. https://doi.org/10.3390/ijms20163923
APA StyleZheng, J., Xu, H., & Cao, H. (2019). A Long Polymorphic GT Microsatellite within a Gene Promoter Mediates Non-Imprinted Allele-Specific DNA Methylation of a CpG Island in a Goldfish Inter-Strain Hybrid. International Journal of Molecular Sciences, 20(16), 3923. https://doi.org/10.3390/ijms20163923