Effects of AntagomiRs on Different Lung Diseases in Human, Cellular, and Animal Models

 ,
,  ,
, 
Abstract
:1. Introduction
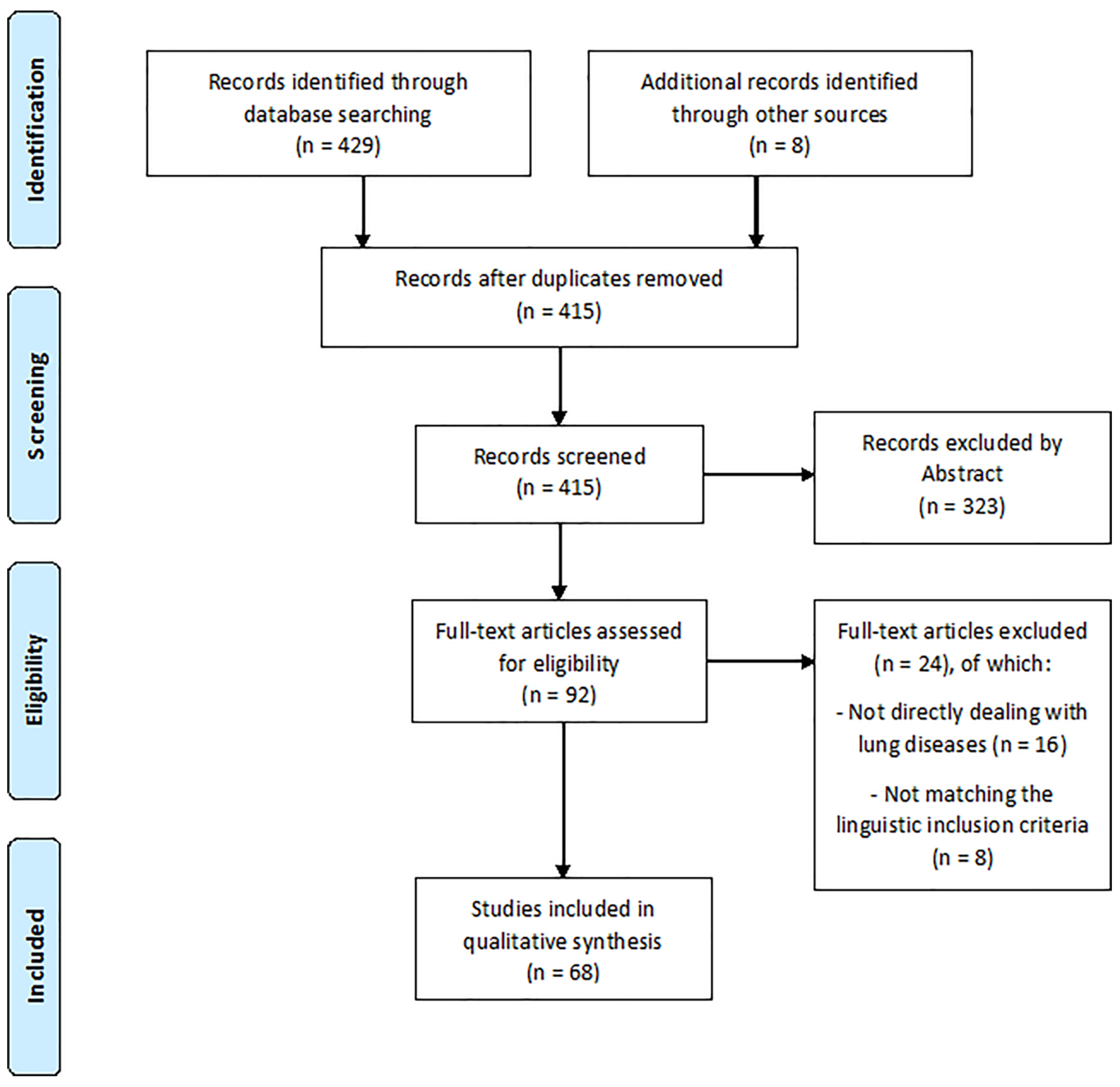
2. Materials and Methods
3. Results
3.1. Lung Cancer
3.1.1. Humans
3.1.2. Animals
3.1.3. Cell lines
3.2. Bronchial Hypersensitivity
3.2.1. Humans
3.2.2. Animals
3.2.3. Cell lines
3.3. Pulmonary Hypertension
3.3.1. Humans
3.3.2. Animals
3.3.3. Cell lines
3.4. Lung Injury
3.4.1. Animals
3.4.2. Cell lines
3.5. Other Conditions
3.5.1. Humans
3.5.2. Animals
3.5.3. Cell Lines
4. Discussion
5. Conclusions
- Strong evidence confirmed that miRNAs play a crucial role in several pathologic mechanisms, therefore they have been proposed as diagnostic or prognostic biomarkers and therapeutic targets for future treatments.
- AntagomiRs are chemically modified oligonucleotides able of silencing microRNAs and are now emerging as novel therapeutic agents in several conditions.
- It has been widely demonstrated that miRNAs have a fundamental role among several lung conditions, moreover, some of these molecules proved to have a lung-specific tropism, thus suggesting the idea of lung-specific miRNAs patterns.
- To our best knowledge, this is the first article describing the evidence on the involvement of miRNAs and the efficacy of respective antagomiRs in different lung diseases, including studies conducted on humans, animals, and cells.
- Coherence between these groups has been demonstrated, thus suggesting the importance of developing new studies on these agents as target therapies.
Funding
Conflicts of Interest
References
- Krol, J.; Loedige, I.; Filipowicz, W. The widespread regulation of microrna biogenesis, function and decay. Nat. Rev. Genet. 2010, 11, 597–610. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. Metazoan micrornas. Cell 2018, 173, 20–51. [Google Scholar] [CrossRef] [PubMed]
- Gambari, R.; Brognara, E.; Spandidos, D.A.; Fabbri, E. Targeting oncomirnas and mimicking tumor suppressor mirnas: Nuew trends in the development of mirna therapeutic strategies in oncology (review). Int. J. Oncol. 2016, 49, 5–32. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.D.; Chang, S. Development of novel therapeutic agents by inhibition of oncogenic micrornas. Int. J. Mol. Sci. 2017, 19. [Google Scholar]
- Jadideslam, G.; Ansarin, K.; Sakhinia, E.; Babaloo, Z.; Abhari, A.; Ghahremanzadeh, K.; Khalili, M.; Radmehr, R.; Kabbazi, A. Diagnostic biomarker and therapeutic target applications of mir-326 in cancers: A systematic review. J. Cell Physiol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Yoo, B.H.; Bochkareva, E.; Bochkarev, A.; Mou, T.C.; Gray, D.M. 2’-o-methyl-modified phosphorothioate antisense oligonucleotides have reduced non-specific effects in vitro. Nucleic Acids Res. 2004, 32, 2008–2016. [Google Scholar] [CrossRef] [PubMed]
- Krutzfeldt, J.; Rajewsky, N.; Braich, R.; Rajeev, K.G.; Tuschl, T.; Manoharan, M.; Stoffel, M. Silencing of micrornas in vivo with ’antagomirs’. Nature 2005, 438, 685–689. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Jung, Y.D.; Choi, Y.S.; Lee, Y.M. Targeting of runx3 by mir-130a and mir-495 cooperatively increases cell proliferation and tumor angiogenesis in gastric cancer cells. Oncotarget 2015, 6, 33269–33278. [Google Scholar] [CrossRef] [PubMed]
- Brognara, E.; Fabbri, E.; Montagner, G.; Gasparello, J.; Manicardi, A.; Corradini, R.; Bianchi, N.; Finotti, A.; Breveglieri, G.; Borgatti, M.; et al. High levels of apoptosis are induced in human glioma cell lines by co-administration of peptide nucleic acids targeting mir-221 and mir-222. Int. J. Oncol. 2016, 48, 1029–1038. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; Ahn, J.; Guo, D.; Votaw, J.R.; Shim, H. Microrna-302 replacement therapy sensitizes breast cancer cells to ionizing radiation. Pharm. Res. 2013, 30, 1008–1016. [Google Scholar] [CrossRef] [PubMed]
- Pace, E.; Di Vincenzo, S.; Di Salvo, E.; Genovese, S.; Dino, P.; Sangiorgi, C.; Ferraro, M.; Gangemi, S. Mir-21 upregulation increases il-8 expression and tumorigenesis program in airway epithelial cells exposed to cigarette smoke. J. Cell Physiol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Shrine, N.; Guyatt, A.L.; Erzurumluoglu, A.M.; Jackson, V.E.; Hobbs, B.D.; Melbourne, C.A.; Batini, C.; Fawcett, K.A.; Song, K.; Sakornsakolpat, P.; et al. New genetic signals for lung function highlight pathways and chronic obstructive pulmonary disease associations across multiple ancestries. Nat. Genet. 2019, 51, 481–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hobbs, B.D.; Tantisira, K.G. Micrornas in copd: Small molecules with big potential. Eur. Respir. J. 2019, 53. [Google Scholar] [CrossRef] [PubMed]
- Faiz, A.; Steiling, K.; Roffel, M.P.; Postma, D.S.; Spira, A.; Lenburg, M.E.; Borggrewe, M.; Eijgenraam, T.R.; Jonker, M.R.; Koppelman, G.H.; et al. Effect of long-term corticosteroid treatment on microrna and gene-expression profiles in copd. Eur. Respir. J. 2019, 53. [Google Scholar] [CrossRef] [PubMed]
- Heffler, E.; Allegra, A.; Pioggia, G.; Picardi, G.; Musolino, C.; Gangemi, S. Microrna profiling in asthma: Potential biomarkers and therapeutic targets. Am. J. Respir. Cell Mol. Biol. 2017, 57, 642–650. [Google Scholar] [CrossRef]
- Yu, B.; Yao, L.; Liu, C.; Tang, L.; Xing, T. Upregulation of microrna16 alters the response to inhaled betaagonists in patients with asthma though modulating expression of adrb2. Mol. Med. Rep. 2019, 19, 4027–4034. [Google Scholar] [PubMed]
- Galie, N.; Humbert, M.; Vachiery, J.L.; Gibbs, S.; Lang, I.; Torbicki, A.; Simonneau, G.; Peacock, A.; Vonk Noordegraaf, A.; Beghetti, M.; et al. 2015 esc/ers guidelines for the diagnosis and treatment of pulmonary hypertension: The joint task force for the diagnosis and treatment of pulmonary hypertension of the european society of cardiology (esc) and the european respiratory society (ers): Endorsed by: Association for european paediatric and congenital cardiology (aepc), international society for heart and lung transplantation (ishlt). Eur. Heart J. 2016, 37, 67–119. [Google Scholar]
- Wheeler, A.P.; Bernard, G.R. Acute lung injury and the acute respiratory distress syndrome: A clinical review. Lancet 2007, 369, 1553–1564. [Google Scholar] [CrossRef]
- Zhou, T.; Garcia, J.G.; Zhang, W. Integrating micrornas into a system biology approach to acute lung injury. Transl. Res. 2011, 157, 180–190. [Google Scholar] [CrossRef]
- Wang, Q.Z.; Xu, W.; Habib, N.; Xu, R. Potential uses of microrna in lung cancer diagnosis, prognosis, and therapy. Curr. Cancer Drug Targets 2009, 9, 572–594. [Google Scholar] [CrossRef]
- Yu, S.L.; Chen, H.Y.; Chang, G.C.; Chen, C.Y.; Chen, H.W.; Singh, S.; Cheng, C.L.; Yu, C.J.; Lee, Y.C.; Chen, H.S.; et al. Microrna signature predicts survival and relapse in lung cancer. Cancer Cell 2008, 13, 48–57. [Google Scholar] [CrossRef]
- Janssen, H.L.; Reesink, H.W.; Lawitz, E.J.; Zeuzem, S.; Rodriguez-Torres, M.; Patel, K.; van der Meer, A.J.; Patick, A.K.; Chen, A.; Zhou, Y.; et al. Treatment of hcv infection by targeting microrna. N. Engl. J. Med. 2013, 368, 1685–1694. [Google Scholar] [CrossRef]
- Bouchie, A. First microrna mimic enters clinic. Nat. Biotechnol. 2013, 31, 577. [Google Scholar] [CrossRef]
- Collison, A.; Mattes, J.; Plank, M.; Foster, P.S. Inhibition of house dust mite-induced allergic airways disease by antagonism of microrna-145 is comparable to glucocorticoid treatment. J. Allergy Clin. Immunol. 2011, 128, 160–167.e4. [Google Scholar] [CrossRef]
- Pullamsetti, S.S.; Doebele, C.; Fischer, A.; Savai, R.; Kojonazarov, B.; Dahal, B.K.; Ghofrani, H.A.; Weissmann, N.; Grimminger, F.; Bonauer, A.; et al. Inhibition of microrna-17 improves lung and heart function in experimental pulmonary hypertension. Am. J. Respir. Crit. Care Med. 2012, 185, 409–419. [Google Scholar] [CrossRef]
- Shentu, T.P.; Huang, T.S.; Cernelc-Kohan, M.; Chan, J.; Wong, S.S.; Espinoza, C.R.; Tan, C.; Gramaglia, I.; van der Heyde, H.; Chien, S.; et al. Thy-1 dependent uptake of mesenchymal stem cell-derived extracellular vesicles blocks myofibroblastic differentiation. Sci. Rep. 2017, 7, 18052. [Google Scholar] [CrossRef]
- Kim, R.Y.; Horvat, J.C.; Pinkerton, J.W.; Starkey, M.R.; Essilfie, A.T.; Mayall, J.R.; Nair, P.M.; Hansbro, N.G.; Jones, B.; Haw, T.J.; et al. Microrna-21 drives severe, steroid-insensitive experimental asthma by amplifying phosphoinositide 3-kinase-mediated suppression of histone deacetylase 2. J. Allergy Clin. Immunol. 2017, 139, 519–532. [Google Scholar] [CrossRef]
- Li, J.W.; Wei, L.; Han, Z.; Chen, Z. Mesenchymal stromal cells-derived exosomes alleviate ischemia/reperfusion injury in mouse lung by transporting anti-apoptotic mir-21-5p. Eur. J. Pharmacol. 2019, 852, 68–76. [Google Scholar] [CrossRef]
- Bhattacharyya, S.; Balakathiresan, N.S.; Dalgard, C.; Gutti, U.; Armistead, D.; Jozwik, C.; Srivastava, M.; Pollard, H.B.; Biswas, R. Elevated mir-155 promotes inflammation in cystic fibrosis by driving hyperexpression of interleukin-8. J. Biol. Chem. 2011, 286, 11604–11615. [Google Scholar] [CrossRef]
- Plank, M.W.; Maltby, S.; Tay, H.L.; Stewart, J.; Eyers, F.; Hansbro, P.M.; Foster, P.S. Microrna expression is altered in an ovalbumin-induced asthma model and targeting mir-155 with antagomirs reveals cellular specificity. PLoS ONE 2015, 10, e0144810. [Google Scholar] [CrossRef]
- Podsiad, A.; Standiford, T.J.; Ballinger, M.N.; Eakin, R.; Park, P.; Kunkel, S.L.; Moore, B.B.; Bhan, U. Microrna-155 regulates host immune response to postviral bacterial pneumonia via il-23/il-17 pathway. Am. J. Physiol. Lung Cell Mol. Physiol. 2016, 310, L465–L475. [Google Scholar] [CrossRef]
- Yuan, Z.; Syed, M.; Panchal, D.; Joo, M.; Bedi, C.; Lim, S.; Onyuksel, H.; Rubinstein, I.; Colonna, M.; Sadikot, R.T. Trem-1-accentuated lung injury via mir-155 is inhibited by lp17 nanomedicine. Am. J. Physiol. Lung Cell Mol. Physiol. 2016, 310, L426–L438. [Google Scholar] [CrossRef]
- He, S.; Li, Z.; Yu, Y.; Zeng, Q.; Cheng, Y.; Ji, W.; Xia, W.; Lu, S. Exosomal mir-499a-5p promotes cell proliferation, migration and emt via mtor signaling pathway in lung adenocarcinoma. Exp. Cell Res. 2019, 379, 203–213. [Google Scholar] [CrossRef]
- Liu, M.; Wang, Z.; Yang, S.; Zhang, W.; He, S.; Hu, C.; Zhu, H.; Quan, L.; Bai, J.; Xu, N. Tnf-alpha is a novel target of mir-19a. Int. J. Oncol. 2011, 38, 1013–1022. [Google Scholar]
- Lee, H.Y.; Lee, H.Y.; Choi, J.Y.; Hur, J.; Kim, I.K.; Kim, Y.K.; Kang, J.Y.; Lee, S.Y. Inhibition of microrna-21 by an antagomir ameliorates allergic inflammation in a mouse model of asthma. Exp. Lung Res. 2017, 43, 109–119. [Google Scholar] [CrossRef]
- Bartoszewska, S.; Kamysz, W.; Jakiela, B.; Sanak, M.; Kroliczewski, J.; Bebok, Z.; Bartoszewski, R.; Collawn, J.F. Mir-200b downregulates cftr during hypoxia in human lung epithelial cells. Cell Mol. Biol. Lett. 2017, 22, 23. [Google Scholar] [CrossRef]
- Baker, J.R.; Vuppusetty, C.; Colley, T.; Papaioannou, A.I.; Fenwick, P.; Donnelly, L.; Ito, K.; Barnes, P.J. Oxidative stress dependent microrna-34a activation via pi3kalpha reduces the expression of sirtuin-1 and sirtuin-6 in epithelial cells. Sci. Rep. 2016, 6, 35871. [Google Scholar] [CrossRef]
- Baker, J.R.; Vuppusetty, C.; Colley, T.; Hassibi, S.; Fenwick, P.S.; Donnelly, L.E.; Ito, K.; Barnes, P.J. Microrna-570 is a novel regulator of cellular senescence and inflammaging. FASEB J. 2019, 33, 1605–1616. [Google Scholar] [CrossRef]
- Brock, M.; Samillan, V.J.; Trenkmann, M.; Schwarzwald, C.; Ulrich, S.; Gay, R.E.; Gassmann, M.; Ostergaard, L.; Gay, S.; Speich, R.; et al. Antagomir directed against mir-20a restores functional bmpr2 signalling and prevents vascular remodelling in hypoxia-induced pulmonary hypertension. Eur. Heart J. 2014, 35, 3203–3211. [Google Scholar] [CrossRef]
- Cha, S.T.; Chen, P.S.; Johansson, G.; Chu, C.Y.; Wang, M.Y.; Jeng, Y.M.; Yu, S.L.; Chen, J.S.; Chang, K.J.; Jee, S.H.; et al. Microrna-519c suppresses hypoxia-inducible factor-1alpha expression and tumor angiogenesis. Cancer Res. 2010, 70, 2675–2685. [Google Scholar] [CrossRef]
- Chatterjee, V.; Beard, R.S., Jr.; Reynolds, J.J.; Haines, R.; Guo, M.; Rubin, M.; Guido, J.; Wu, M.H.; Yuan, S.Y. Microrna-147b regulates vascular endothelial barrier function by targeting adam15 expression. PLoS ONE 2014, 9, e110286. [Google Scholar] [CrossRef]
- Chiu, K.L.; Kuo, T.T.; Kuok, Q.Y.; Lin, Y.S.; Hua, C.H.; Lin, C.Y.; Su, P.Y.; Lai, L.C.; Sher, Y.P. Adam9 enhances cdcp1 protein expression by suppressing mir-218 for lung tumor metastasis. Sci. Rep. 2015, 5, 16426. [Google Scholar] [CrossRef]
- Fabbri, E.; Borgatti, M.; Montagner, G.; Bianchi, N.; Finotti, A.; Lampronti, I.; Bezzerri, V.; Dechecchi, M.C.; Cabrini, G.; Gambari, R. Expression of microrna-93 and interleukin-8 during pseudomonas aeruginosa-mediated induction of proinflammatory responses. Am. J. Respir. Cell Mol. Biol. 2014, 50, 1144–1155. [Google Scholar] [CrossRef]
- Fehl, J.; Pozarska, A.; Nardiello, C.; Rath, P.; Surate Solaligue, D.E.; Vadasz, I.; Mayer, K.; Herold, S.; Seeger, W.; Morty, R.E. Control interventions can impact alveolarization and the transcriptome in developing mouse lungs. Anat. Rec. (Hoboken) 2019, 302, 346–363. [Google Scholar] [CrossRef]
- Fu, L.; Zhu, P.; Qi, S.; Li, C.; Zhao, K. Microrna-92a antagonism attenuates lipopolysaccharide (lps)-induced pulmonary inflammation and injury in mice through suppressing the pten/akt/nf-kappab signaling pathway. Biomed. Pharmacother. 2018, 107, 703–711. [Google Scholar] [CrossRef]
- Guo, L.; Liu, Y.; Bai, Y.; Sun, Y.; Xiao, F.; Guo, Y. Gene expression profiling of drug-resistant small cell lung cancer cells by combining microrna and cdna expression analysis. Eur. J. Cancer 2010, 46, 1692–1702. [Google Scholar] [CrossRef]
- Huang, Y.; Huang, L.; Zhu, G.; Pei, Z.; Zhang, W. Downregulated microrna-27b attenuates lipopolysaccharide-induced acute lung injury via activation of nf-e2-related factor 2 and inhibition of nuclear factor kappab signaling pathway. J. Cell Physiol. 2019, 234, 6023–6032. [Google Scholar] [CrossRef]
- Incoronato, M.; Garofalo, M.; Urso, L.; Romano, G.; Quintavalle, C.; Zanca, C.; Iaboni, M.; Nuovo, G.; Croce, C.M.; Condorelli, G. Mir-212 increases tumor necrosis factor-related apoptosis-inducing ligand sensitivity in non-small cell lung cancer by targeting the antiapoptotic protein ped. Cancer Res. 2010, 70, 3638–3646. [Google Scholar] [CrossRef]
- Jiang, C.; Guo, Y.; Yu, H.; Lu, S.; Meng, L. Pleiotropic microrna-21 in pulmonary remodeling: Novel insights for molecular mechanism and present advancements. Allergy Asthma Clin. Immunol. 2019, 15, 33. [Google Scholar] [CrossRef]
- Li, J.J.; Tay, H.L.; Maltby, S.; Xiang, Y.; Eyers, F.; Hatchwell, L.; Zhou, H.; Toop, H.D.; Morris, J.C.; Nair, P.; et al. Microrna-9 regulates steroid-resistant airway hyperresponsiveness by reducing protein phosphatase 2a activity. J. Allergy Clin. Immunol. 2015, 136, 462–473. [Google Scholar] [CrossRef]
- Mondejar-Parreno, G.; Callejo, M.; Barreira, B.; Morales-Cano, D.; Esquivel-Ruiz, S.; Moreno, L.; Cogolludo, A.; Perez-Vizcaino, F. Mir-1 is increased in pulmonary hypertension and downregulates kv1.5 channels in rat pulmonary arteries. J. Physiol. 2019, 597, 1185–1197. [Google Scholar] [CrossRef]
- Zhang, Q.; Ran, R.; Zhang, L.; Liu, Y.; Mei, L.; Zhang, Z.; Gao, H.; He, Q. Simultaneous delivery of therapeutic antagomirs with paclitaxel for the management of metastatic tumors by a ph-responsive anti-microbial peptide-mediated liposomal delivery system. J. Control. Release 2015, 197, 208–218. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, M.; Hu, C. Exosomal transfer of mir-214 mediates gefitinib resistance in non-small cell lung cancer. Biochem. Biophys. Res. Commun. 2018, 507, 457–464. [Google Scholar] [CrossRef]
- Zhou, Z.; Li, X.; Liu, J.; Dong, L.; Chen, Q.; Liu, J.; Kong, H.; Zhang, Q.; Qi, X.; Hou, D.; et al. Honeysuckle-encoded atypical microrna2911 directly targets influenza a viruses. Cell Res. 2015, 25, 39–49. [Google Scholar] [CrossRef]
- Zhu, Q.; Zang, Q.; Jiang, Z.M. Enhanced expression of non coding mir 92a expression is implicated in the development of lung cancer. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 1028–1034. [Google Scholar]
- Rosenberger, C.M.; Podyminogin, R.L.; Navarro, G.; Zhao, G.W.; Askovich, P.S.; Weiss, M.J.; Aderem, A. Mir-451 regulates dendritic cell cytokine responses to influenza infection. J. Immunol. 2012, 189, 5965–5975. [Google Scholar] [CrossRef]
- Sharma, S.; Umar, S.; Centala, A.; Eghbali, M. Role of mir206 in genistein-induced rescue of pulmonary hypertension in monocrotaline model. J. Appl. Physiol. (1985) 2015, 119, 1374–1382. [Google Scholar] [CrossRef]
- Sharma, H.; Chinnappan, M.; Agarwal, S.; Dalvi, P.; Gunewardena, S.; O’Brien-Ladner, A.; Dhillon, N.K. Macrophage-derived extracellular vesicles mediate smooth muscle hyperplasia: Role of altered mirna cargo in response to hiv infection and substance abuse. FASEB J. 2018, 32, 5174–5185. [Google Scholar] [CrossRef]
- Shi, Y.; Liu, C.; Liu, X.; Tang, D.G.; Wang, J. The microrna mir-34a inhibits non-small cell lung cancer (nsclc) growth and the cd44hi stem-like nsclc cells. PLoS ONE 2014, 9, e90022. [Google Scholar] [CrossRef]
- Silveyra, P.; DiAngelo, S.L.; Floros, J. An 11-nt sequence polymorphism at the 3’utr of human sftpa1 and sftpa2 gene variants differentially affect gene expression levels and mirna regulation in cell culture. Am. J. Physiol. Lung Cell Mol. Physiol. 2014, 307, L106–L119. [Google Scholar] [CrossRef]
- Sun, B.; Yang, N.; Jiang, Y.; Zhang, H.; Hou, C.; Ji, C.; Liu, Y.; Zuo, P. Antagomir-1290 suppresses cd133(+) cells in non-small cell lung cancer by targeting fyn-related src family tyrosine kinase. Tumour Biol. 2015, 36, 6223–6230. [Google Scholar] [CrossRef]
- Sun, C.C.; Li, S.J.; Yuan, Z.P.; Li, D.J. Microrna-346 facilitates cell growth and metastasis, and suppresses cell apoptosis in human non-small cell lung cancer by regulation of xpc/erk/snail/e-cadherin pathway. Aging (Albany NY) 2016, 8, 2509–2524. [Google Scholar] [CrossRef]
- Vera, O.; Jimenez, J.; Pernia, O.; Rodriguez-Antolin, C.; Rodriguez, C.; Sanchez Cabo, F.; Soto, J.; Rosas, R.; Lopez-Magallon, S.; Esteban Rodriguez, I.; et al. DNA methylation of mir-7 is a mechanism involved in platinum response through mafg overexpression in cancer cells. Theranostics 2017, 7, 4118–4134. [Google Scholar] [CrossRef]
- Wu, T.; Chen, W.; Kong, D.; Li, X.; Lu, H.; Liu, S.; Wang, J.; Du, L.; Kong, Q.; Huang, X.; et al. Mir-25 targets the modulator of apoptosis 1 gene in lung cancer. Carcinogenesis 2015, 36, 925–935. [Google Scholar] [CrossRef]
- Wu, C.; Xu, B.; Zhou, Y.; Ji, M.; Zhang, D.; Jiang, J.; Wu, C. Correlation between serum il-1beta and mir-144-3p as well as their prognostic values in luad and lusc patients. Oncotarget 2016, 7, 85876–85887. [Google Scholar] [CrossRef]
- Wu, L.; Pu, X.; Wang, Q.; Cao, J.; Xu, F.; Xu, L.I.; Li, K. Mir-96 induces cisplatin chemoresistance in non-small cell lung cancer cells by downregulating samd9. Oncol. Lett. 2016, 11, 945–952. [Google Scholar] [CrossRef]
- Wu, Y.; He, H.; Ding, Y.; Liu, S.; Zhang, D.; Wang, J.; Jiang, H.; Zhang, D.; Sun, L.; Ye, R.D.; et al. Mk2 mediates macrophage activation and acute lung injury by regulating let-7e mirna. Am. J. Physiol. Lung Cell Mol. Physiol. 2018, 315, L371–L381. [Google Scholar] [CrossRef]
- Xu, Z.; Zhang, C.; Cheng, L.; Hu, M.; Tao, H.; Song, L. The microrna mir-17 regulates lung foxa1 expression during lipopolysaccharide-induced acute lung injury. Biochem. Biophys Res. Commun. 2014, 445, 48–53. [Google Scholar] [CrossRef]
- Yuan, Y.; Lin, D.; Feng, L.; Huang, M.; Yan, H.; Li, Y.; Chen, Y.; Lin, B.; Ma, Y.; Ye, Z.; et al. Upregulation of mir-196b-5p attenuates bcg uptake via targeting socs3 and activating stat3 in macrophages from patients with long-term cigarette smoking-related active pulmonary tuberculosis. J. Transl. Med. 2018, 16, 284. [Google Scholar] [CrossRef]
- Xie, Z.; Chen, W.; Chen, Y.; Wang, X.; Gao, W.; Liu, Y. Mir-768-3p is involved in the proliferation, invasion and migration of non-small cell lung carcinomas. Int. J. Oncol. 2017, 51, 1574–1582. [Google Scholar] [CrossRef]
- Ma, C.; Zhang, C.; Ma, M.; Zhang, L.; Zhang, L.; Zhang, F.; Chen, Y.; Cao, F.; Li, M.; Wang, G.; et al. Mir-125a regulates mitochondrial homeostasis through targeting mitofusin 1 to control hypoxic pulmonary vascular remodeling. J. Mol. Med. (Berl) 2017, 95, 977–993. [Google Scholar] [CrossRef]
- Zhou, M.; Hara, H.; Dai, Y.; Mou, L.; Cooper, D.K.; Wu, C.; Cai, Z. Circulating organ-specific micrornas serve as biomarkers in organ-specific diseases: Implications for organ allo- and xeno-transplantation. Int. J. Mol. Sci. 2016, 17. [Google Scholar] [CrossRef]
- Lin, C.W.; Chang, Y.L.; Chang, Y.C.; Lin, J.C.; Chen, C.C.; Pan, S.H.; Wu, C.T.; Chen, H.Y.; Yang, S.C.; Hong, T.M.; et al. Microrna-135b promotes lung cancer metastasis by regulating multiple targets in the hippo pathway and lzts1. Nat. Commun. 2013, 4, 1877. [Google Scholar] [CrossRef]
- Mao, G.; Liu, Y.; Fang, X.; Liu, Y.; Fang, L.; Lin, L.; Liu, X.; Wang, N. Tumor-derived microrna-494 promotes angiogenesis in non-small cell lung cancer. Angiogenesis 2015, 18, 373–382. [Google Scholar] [CrossRef]
- McCann, J.V.; Xiao, L.; Kim, D.J.; Khan, O.F.; Kowalski, P.S.; Anderson, D.G.; Pecot, C.V.; Azam, S.H.; Parker, J.S.; Tsai, Y.S.; et al. Endothelial mir-30c suppresses tumor growth via inhibition of tgf-beta-induced serpine1. J. Clin. Investig. 2019, 130, 1654–1670. [Google Scholar] [CrossRef]
- Hsu, A.C.; Parsons, K.; Moheimani, F.; Knight, D.A.; Hansbro, P.M.; Fujita, T.; Wark, P.A. Impaired antiviral stress granule and ifn-beta enhanceosome formation enhances susceptibility to influenza infection in chronic obstructive pulmonary disease epithelium. Am. J. Respir. Cell Mol. Biol. 2016, 55, 117–127. [Google Scholar] [CrossRef]
- Potus, F.; Malenfant, S.; Graydon, C.; Mainguy, V.; Tremblay, E.; Breuils-Bonnet, S.; Ribeiro, F.; Porlier, A.; Maltais, F.; Bonnet, S.; et al. Impaired angiogenesis and peripheral muscle microcirculation loss contribute to exercise intolerance in pulmonary arterial hypertension. Am. J. Respir. Crit Care Med. 2014, 190, 318–328. [Google Scholar] [CrossRef]
- Gubrij, I.B.; Pangle, A.K.; Pang, L.; Johnson, L.G. Reversal of microrna dysregulation in an animal model of pulmonary hypertension. PLoS ONE 2016, 11, e0147827. [Google Scholar] [CrossRef]
- Xie, W.; Lu, Q.; Wang, K.; Lu, J.; Gu, X.; Zhu, D.; Liu, F.; Guo, Z. Mir-34b-5p inhibition attenuates lung inflammation and apoptosis in an lps-induced acute lung injury mouse model by targeting progranulin. J. Cell Physiol. 2018, 233, 6615–6631. [Google Scholar] [CrossRef]
- Adyshev, D.M.; Moldobaeva, N.; Mapes, B.; Elangovan, V.; Garcia, J.G. Microrna regulation of nonmuscle myosin light chain kinase expression in human lung endothelium. Am. J. Respir. Cell Mol. Biol. 2013, 49, 58–66. [Google Scholar] [CrossRef]
- Adyshev, D.M.; Elangovan, V.R.; Moldobaeva, N.; Mapes, B.; Sun, X.; Garcia, J.G. Mechanical stress induces pre-b-cell colony-enhancing factor/nampt expression via epigenetic regulation by mir-374a and mir-568 in human lung endothelium. Am. J. Respir. Cell Mol. Biol. 2014, 50, 409–418. [Google Scholar]
- Ge, L.; Habiel, D.M.; Hansbro, P.M.; Kim, R.Y.; Gharib, S.A.; Edelman, J.D.; Konigshoff, M.; Parimon, T.; Brauer, R.; Huang, Y.; et al. Mir-323a-3p regulates lung fibrosis by targeting multiple profibrotic pathways. JCI Insight 2016, 1, e90301. [Google Scholar] [CrossRef]
- Chiba, Y.; Tanabe, M.; Goto, K.; Sakai, H.; Misawa, M. Down-regulation of mir-133a contributes to up-regulation of rhoa in bronchial smooth muscle cells. Am. J. Respir. Crit Care Med. 2009, 180, 713–719. [Google Scholar] [CrossRef]
- Pandit, K.V.; Corcoran, D.; Yousef, H.; Yarlagadda, M.; Tzouvelekis, A.; Gibson, K.F.; Konishi, K.; Yousem, S.A.; Singh, M.; Handley, D.; et al. Inhibition and role of let-7d in idiopathic pulmonary fibrosis. Am. J. Respir. Crit. Care Med. 2010, 182, 220–229. [Google Scholar] [CrossRef]
- Asquith, M.; Pasala, S.; Engelmann, F.; Haberthur, K.; Meyer, C.; Park, B.; Grant, K.A.; Messaoudi, I. Chronic ethanol consumption modulates growth factor release, mucosal cytokine production, and microrna expression in nonhuman primates. Alcohol. Clin. Exp. Res. 2014, 38, 980–993. [Google Scholar] [CrossRef]
- Zhou, S.; Wang, Y.; Meng, Y.; Xiao, C.; Liu, Z.; Brohawn, P.; Higgs, B.W.; Jallal, B.; Jia, Q.; Qu, B.; et al. In vivo therapeutic success of microrna-155 antagomir in a mouse model of lupus alveolar hemorrhage. Arthritis Rheumatol. 2016, 68, 953–964. [Google Scholar] [CrossRef]
- Morales, L.; Oliveros, J.C.; Fernandez-Delgado, R.; tenOever, B.R.; Enjuanes, L.; Sola, I. Sars-cov-encoded small rnas contribute to infection-associated lung pathology. Cell Host Microbe 2017, 21, 344–355. [Google Scholar] [CrossRef]
- Zhou, P.; Tu, L.; Lin, X.; Hao, X.; Zheng, Q.; Zeng, W.; Zhang, X.; Zheng, Y.; Wang, L.; Li, S. Cfa-mir-143 promotes apoptosis via the p53 pathway in canine influenza virus h3n2-infected cells. Viruses 2017, 9. [Google Scholar] [CrossRef]
- Tamgue, O.; Gcanga, L.; Ozturk, M.; Whitehead, L.; Pillay, S.; Jacobs, R.; Roy, S.; Schmeier, S.; Davids, M.; Medvedeva, Y.A.; et al. Differential targeting of c-maf, bach-1, and elmo-1 by microrna-143 and microrna-365 promotes the intracellular growth of mycobacterium tuberculosis in alternatively il-4/il-13 activated macrophages. Front. Immunol. 2019, 10, 421. [Google Scholar] [CrossRef]
- Zhang, F.; Sun, X.; Zhu, Y.; Qin, W. Downregulation of mir-146a inhibits influenza a virus replication by enhancing the type i interferon response in vitro and in vivo. Biomed. Pharmacother 2019, 111, 740–750. [Google Scholar] [CrossRef]
- Liao, W.; Dong, J.; Peh, H.Y.; Tan, L.H.; Lim, K.S.; Li, L.; Wong, W.F. Oligonucleotide therapy for obstructive and restrictive respiratory diseases. Molecules 2017, 22. [Google Scholar] [CrossRef]
- Mei, D.; Tan, W.S.D.; Wong, W.S.F. Pharmacological strategies to regain steroid sensitivity in severe asthma and copd. Curr. Opin. Pharmacol. 2019, 46, 73–81. [Google Scholar] [CrossRef]
- Testa, U.; Pelosi, E.; Castelli, G.; Labbaye, C. Mir-146 and mir-155: Two key modulators of immune response and tumor development. Noncoding RNA 2017, 3. [Google Scholar] [CrossRef]
- Sessa, R.; Hata, A. Role of micrornas in lung development and pulmonary diseases. Pulm. Circ. 2013, 3, 315–328. [Google Scholar] [CrossRef]
- Panwar, B.; Omenn, G.S.; Guan, Y. Mirmine: A database of human mirna expression profiles. Bioinformatics 2017, 33, 1554–1560. [Google Scholar] [CrossRef]
- Zhang, J.G.; Wang, J.J.; Zhao, F.; Liu, Q.; Jiang, K.; Yang, G.H. Microrna-21 (mir-21) represses tumor suppressor pten and promotes growth and invasion in non-small cell lung cancer (nsclc). Clin. Chim. Acta 2010, 411, 846–852. [Google Scholar] [CrossRef]
- Porteous, M.K.; Lee, J.C. Primary graft dysfunction after lung transplantation. Clin. Chest Med. 2017, 38, 641–654. [Google Scholar] [CrossRef]
- Liu, G.; Friggeri, A.; Yang, Y.; Milosevic, J.; Ding, Q.; Thannickal, V.J.; Kaminski, N.; Abraham, E. Mir-21 mediates fibrogenic activation of pulmonary fibroblasts and lung fibrosis. J. Exp. Med. 2010, 207, 1589–1597. [Google Scholar] [CrossRef]
- O’Connell, R.M.; Taganov, K.D.; Boldin, M.P.; Cheng, G.; Baltimore, D. Microrna-155 is induced during the macrophage inflammatory response. Proc. Natl. Acad. Sci. USA 2007, 104, 1604–1609. [Google Scholar] [CrossRef]
- Comer, B.S.; Camoretti-Mercado, B.; Kogut, P.C.; Halayko, A.J.; Solway, J.; Gerthoffer, W.T. Cyclooxygenase-2 and microrna-155 expression are elevated in asthmatic airway smooth muscle cells. Am. J. Respir. Cell Mol. Biol. 2015, 52, 438–447. [Google Scholar] [CrossRef]
- Suojalehto, H.; Toskala, E.; Kilpelainen, M.; Majuri, M.L.; Mitts, C.; Lindstrom, I.; Puustinen, A.; Plosila, T.; Sipila, J.; Wolff, H.; et al. Microrna profiles in nasal mucosa of patients with allergic and nonallergic rhinitis and asthma. Int. Forum Allergy Rhinol. 2013, 3, 612–620. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Lung Cancer | ||||
|---|---|---|---|---|
| Humans | ||||
| Study | Disease | Sample | miRNAs | Findings |
| Incoronato et al. (2010) [48] | Lung cancer | Cells: Calu-1 NSCLC cells; H460 cells Human Tissues: 18 snap-frozen normal and malignant lung tissues | miR-212 | AntagomiR 212 increases PED protein expression and resistance to TRAIL treatment (p < 0.05) |
| Liu et al. (2011) [34] | Lung cancer | Cells: esophageal cancer cell lines KYSE150, KYSE410, KYSE70, EC109, and EC9706. Clinical samples: 28 ESCC and 28 normal tissue samples from surgically resected esophageal carcinoma specimens. Animals: five nude mice, injected with EC9706/miR-17-92 and EC9706/Control cells | miR-19a | Antagomir-19a treatment impairs tumor growth in vivo (p < 0.05) |
| Liang et al. (2015) [10] | Lung cancer | 20 NSCLC patients, 20 controls | miR-223 | miR-223 antagomir decreases tumor cell invasion and increases EPB41L3 in A549 cells (p < 0.001) |
| Wu et al. (2015) [64] | Lung cancer | Humans: 81 NSCLC patients (12 Stage I, 14 Stage II, 26 Stage III, 29 Stage IV), 41 controls. Animals: 4-week-old BALB/c nude mice. Cells: NSCLC cell lines (A549, H1299, H1975, Hcc827), human embryonic kidney (HEK) 293T cells | miR-25 | miR-25 antagomir inhibited lung cancer growth via upregulation of MOAP1 in a mouse xenograft model (p < 0.01) |
| Sun et al. (2016) [62] | Lung cancer | Humans: NSCLC tumor tissue samples. Cells: human NSCLC cell lines A549, H1299, SPC-A-1, 95D, SK-MES-1, NCI-H520, NCI-H460, human normal lung epithelial cell line 16HBE | miR-346 | AntagomiR-346 inhibited NSCLC cell growth and metastasis |
| Vera et al. (2017) [63] | Lung cancer | Fifteen human cancer cell lines, ovarian cancer samples (n = 138), high-grade serous carcinoma (n = 22); normal ovarian samples (n = 10), peripheral blood mononuclear cells (n = 10) | miR-7, miR-132, miR-335, and miR-148a | Relative miR-7 and MAFG expression levels decreased when treated with antagomir |
| Wu et al. (2016a) [65] | Lung cancer | Humans: patients with lung adenocarcinoma (n = 129) and lung squamous cell carcinoma (n = 54). Cells: human NSCLC cell line A549 | miR-144-3p | miR-144-3p antagomir could enhance proliferation of IL-1β (p < 0.001) |
| Wu et al. (2016b) [66] | Lung cancer | Humans: human NSCLC tumor, adjacent normal lung tissues (n = 5). Cells: human NSCLC cell lines H358 and H23 | miR-96 | Antagomir-96 increased SAMD9 expression and the cisplatin-induced apoptosis and decreased cisplatin IC50 (p < 0.05) |
| Xie et al. (2017) [70] | Lung cancer | Humans: n = 83 patients with NSCLC. Animals: NSCLC xenograft nude mouse model. Cells: bronchoepithelial cell line BEAS-2B, A549, and HCC4006 human NSCLC cell line | miR-768-3p | miR-768-3p antagomir induced apoptosis and Fas/FasL expressional alteration of NSCLC cells; antimiR-768-3p transduction decreased viability, migration, invasion, MMP-2, MMP-9 activities in A549 and HCC4006 cells; antimiR-768-3p transfection inhibited growth and proliferation of NSCLC xenografts in nude mice (p < 0.05) |
| Zhu et al. (2018) [55] | Lung cancer | Humans: whole blood samples from patients with NSCLC and controls; tumor and nontumorous tissues obtained from NSCLC patients (n = 20). Cells: NSCLC cell lines SPCA1, A549, H2170 | miR-92a | Proliferation of SPCA1, A549, H2170 inhibited by antimiR-92a (p < 0.05) |
| He et al. (2019) [33] | Lung adenocarcinoma | Human lung cancer cells A549, human bronchial epithelial cells BEAS-2B, human lung cancer cells SPCA1, SPC-A-1-BM human lung adenocarcinoma cell line | miR-499a-5p | Inhibition of miR-499a-5p by antagomirs restrained tumor growth in vivo (p < 0.01) |
| Animals | ||||
| Study | Disease | Sample | miRNAs | Findings |
| Cha et al. (2010) [40] | Lung cancer | Animals: n = 20 5-week-old male BALB/c nude mice. Cell lines: CL1-0, CL1-5 lung adenocarcinoma cell lines. H1299, PC14, H928, A549 lung cancer, MCF-7, MDA-MB231, MDA-MD431, T47D, SKBR3 breast cancer cell lines | miR-519c | Antagomir inhibition of miR-519c increased HIF-1α protein and enhanced angiogenic activity (p < 0.05) |
| Liu et al. (2011) [34] | Lung cancer | Cells: esophageal cancer cell lines KYSE150, KYSE410, KYSE70, EC109, EC9706. Clinical samples: 28 ESCC, 28 normal tissue samples from surgically resected esophageal carcinoma specimens. Animals: five nude mice injected with EC9706/miR-17-92 and EC9706/Control cells | miR-19a | Antagomir-19a treatment impairs tumor growth in vivo (p < 0.05) |
| Lin et al. (2013) [73] | Lung cancer | Animals: n = 10 mice. Cells: lung cancer cell lines CL1-0, CL1-1, CL1-5 and CL1-5-F4. A549, HOP-62, H441, CL141 cells, melanoma cell line UACC-257. H1299 and HEK-293 cells | miR-135b | miR-135b antagomirs suppress cancer cell invasion, orthotopic lung tumor growth and metastasis in mouse model (p < 0.05) |
| Shi et al. (2014) [59] | Lung cancer | Animals: immune-deficient NOD-SCID mice. Cells: human NSCLC cell lines A549, H460, and H1299 | miR-34a | Expression of antimiR-34a in the CD44lo H460 cells promoted tumor development (p < 0.05) |
| Chiu et al. (2015) [42] | Lung cancer | Human cells: human lung cancer cell lines CL1-0, F4, Bm7, Bm7brmx2, A549, H1299. Animals: Lung cancer cells injected into SCID mice | miR-218 | CDCP1 protein levels increased in cells treated with miR-218 antagomirs (p < 0.05) |
| Mao et al. (2015) [74] | Lung cancer | Human cells: HUVECs, tumor cell lines A549, H1299, HCC827. Animals: male BALB/c nude mice | miR-494 | MiR-494 antagomiR inhibited angiogenesis and attenuated the growth of tumor xenografts in nude mice (p < 0.05) |
| Wu et al. (2015) [64] | Lung cancer | Humans: 81 NSCLC patients (12 Stage I, 14 Stage II, 26 Stage III, 29 Stage IV), 41 controls. Animals: BALB/c nude mice. Cells: NSCLC cell lines A549, H1299, H1975, Hcc827, HEK 293T cells | miR-25 | miR-25 antagomir inhibited lung cancer growth via upregulation of MOAP1 in a mouse xenograft model (p < 0.01) |
| Xie et al. (2017) [70] | Lung cancer | Humans: n = 83 NSCLC patients. Animals: NSCLC xenograft nude mouse model. Cells: bronchoepithelial cell line BEAS-2B, A549 and HCC4006 human NSCLC cell line | miR-768-3p | miR-768-3p antagomir induced distinctly apoptosis and Fas/FasL expressional alteration of NSCLC cells; miR-768-3p antagomir transduction decreased viability, migration, invasion, MMP-2, and MMP-9 activities in A549 and HCC4006 cells; miR-768-3p antagomir transfection inhibited the growth and proliferation of NSCLC xenografts in nude mice (p < 0.05) |
| Zhang et al. (2018) [53] | Lung cancer | Animals: 20 male BALB/c nu/nu mice. Cells: EGFR mutant non-small cell lung cancer cell line PC-9 | miR-214 | AntagomiR-214 reversed gefitinib resistance conferred by PC-9GR-derived exosomes in vitro and reversed gefitinib resistance in vivo (p < 0.01 in both cases) |
| McCann et al. (2019) [75] | Lung cancer | Animals: mice models. Cells: primary endothelial cells isolated from normal or tumor tissue from mice | miR-30c | miR-30c antagomiRs promoted PAI-1–dependent tumor growth and increased fibrin abundance (p < 0.05) |
| Cell Lines | ||||
| Study | Disease | Sample | miRNAs | Findings |
| Cha et al. (2010) [40] | Lung cancer | Animals: n = 20 5-week-old male BALB/c nude mice. Cell lines: CL1-0, CL1-5 lung adenocarcinoma cell lines. H1299, PC14, H928, A549 lung cancer, MCF-7, MDA-MB231, MDA-MD431, T47D, SKBR3 breast cancer cell lines | miR-519c | Antagomir inhibition of miR-519c increased the level of HIF-1α protein and enhanced angiogenic activity (p < 0.05) |
| Guo et al. (2010) [46] | Lung cancer | Human small lung cancer cell line NCI-H69 and drug-resistant subline H69AR | miR-134, miR-379, miR-495 | Sensitivity to anti-cancer drugs Cisplatin, Etoposide, and Doxorubicin reduced after transfection of drug-resistant H69AR cells with the antagomirs of miR-134, miR-379 and miR-495 (p < 0.05) |
| Incoronato et al. (2010) [48] | Lung cancer | Cells: Calu-1 NSCLC cells; H460 cells Human Tissues: 18 snap-frozen normal and malignant lung tissues | miR-212 | AntagomiR 212 increases PED protein expression and resistance to TRAIL treatment (p < 0.05) |
| Liu et al. (2011) [34] | Lung cancer | Cells: esophageal cancer cell lines KYSE150, KYSE410, KYSE70, EC109, and EC9706. Clinical samples: 28 ESCC and 28 normal tissue samples from surgically resected esophageal carcinoma specimens. Animals: five nude mice, injected with EC9706/miR-17-92 and EC9706/Control cells | miR-19a | Antagomir-19a treatment impairs tumor growth in vivo (p < 0.05) |
| Lin et al. (2013) [73] | Lung cancer | Animals: n = 10 mice. Cells: lung cancer cell lines CL1-0, CL1-1, CL1-5, and CL1-5-F4. A549, HOP-62, H441, CL141 cells, melanoma cell line UACC-257. H1299 and HEK-293 cells | miR-135b | miR-135b antagomirs suppress cancer cell invasion, orthotopic lung tumor growth, and metastasis in mouse model (p < 0.05) |
| Shi et al. (2014) [59] | Lung cancer | Animals: Immune-deficient NOD-SCID mice. Cells: human NSCLC cell lines A549, H460, and H1299 | miR-34a | Expression of miR-34a antagomirs in the CD44lo H460 cells promoted tumor development (p < 0.05) |
| Silveyra et al. (2014) [60] | Lung cancer | Cells: lung adenocarcinoma cell line NCI- H441, three Chinese hamster ovary (CHOK1) cell lines expressing the human SP-A variants 1A0, 6A2, and 6A4 | miR-183, miR-4507 | antagomir-183 reversed the effects of mir-183 on SP-A mRNA levels (p < 0.05) |
| Chiu et al. (2015) [42] | Lung cancer | Human cells: human lung cancer cell lines CL1-0, F4, Bm7, Bm7brmx2, A549, and H1299. Animals: lung cancer cells injected intracardially into SCID mice | miR-218 | CDCP1 levels increased in cells treated with antimiR-218 (p < 0.05) |
| Mao et al. (2015) [74] | Lung cancer | Human cells: HUVECs, tumor cell lines A549, H1299, HCC827. Animals: male BALB/c nude mice | miR-494 | AntimiR-494 inhibited angiogenesis and attenuated the growth of tumor xenografts in nude mice (p < 0.05) |
| Sun et al. (2015) [61] | Lung cancer | NSCLC cell line A549 | miR-1290 | AntimiR-1290 suppressed tumor volume and weight initiated by CD133+ cells in vivo; Anti-miR-1290 inhibited proliferation, clonogenicity, invasion, and migration of CD133+ (p < 0.05) |
| Wu et al. (2015) [64] | Lung cancer | Humans: 81 NSCLC patients (12 Stage I, 14 Stage II, 26 Stage III, 29 Stage IV), 41 controls. Animals: BALB/c nude mice. Cells: NSCLC cell lines A549, H1299, H1975, Hcc827, HEK 293T cells | miR-25 | miR-25 antagomir inhibited lung cancer growth via upregulation of MOAP1 in mice (p < 0.01) |
| Sun et al. (2016) [62] | Lung cancer | Humans: NSCLC tumor tissues. Cells: human NSCLC cell lines A549, H1299, SPC-A-1, 95D, SK-MES-1, NCI-H520, NCI-H460, human normal lung epithelial cell line 16HBE | miR-346 | AntimiR-346 inhibited NSCLC cell growth and metastasis |
| Wu et al. (2016a) [65] | Lung cancer | Humans: patients with lung adenocarcinoma (n = 129) and lung squamous cell carcinoma (n = 54). Cells: human NSCLC cell line A549 | miR-144-3p | miR-144-3p antagomir could enhance IL-1β proliferation (p < 0.001) |
| Wu et al. (2016b) [66] | Lung cancer | Humans: human NSCLC tumor, adjacent normal lung tissues (n = 5). Cells: human NSCLC cell lines H358 and H23 | miR-96 | Antagomir-96 increased SAMD9 expression and the cisplatin-induced apoptosis, it decreased cisplatin IC50 (p < 0.05) |
| Vera et al. (2017) [63] | Lung cancer | 15 human cancer cell lines, ovarian cancer samples (n = 138), high-grade serous carcinoma (n = 22); normal ovarian samples (n = 10), peripheral blood mononuclear cells (n = 10) | miR-7, miR-132, miR-335, and miR-148a | Relative miR-7 and MAFG expression levels decreased when treated with antagomir |
| Xie et al. (2017) [70] | Lung cancer | Humans: n = 83 NSCLC patients. Animals: NSCLC xenograft nude mouse model. Cells: bronchoepithelial cell line BEAS-2B, A549 and HCC4006 human NSCLC cell line | miR-768-3p | miR-768-3p antagomir induced distinctly apoptosis and Fas/FasL expressional alteration of NSCLC cells; miR-768-3p antagomir transduction decreased viability, migration, invasion, MMP-2, and MMP-9 activities in A549 and HCC4006 cells; miR-768-3p antagomir transfection inhibited the growth and proliferation of NSCLC xenografts in nude mice (p < 0.05) |
| Zhang et al. (2018) [53] | Lung cancer | Animals: 20 male BALB/c nu/nu mice. Cells: EGFR mutant non-small cell lung cancer cell line PC-9 | miR-214 | AntagomiR-214 reversed gefitinib resistance in vitro and in vivo (p < 0.01) |
| Zhu et al. (2018) [55] | Lung cancer | Humans: whole blood samples collected from healthy individuals and NSCLC patients; tumor and adjacent nontumorous tissues from NSCLC patients (n = 20). Cells: NSCLC cell lines SPCA1, A549, H2170 | miR-92a | Proliferation of SPCA1, A549, H2170 inhibited by antimiR-92a (p < 0.05) |
| He et al. (2019) [33] | Lung adenocarcinoma | Human lung cancer cells A549, human bronchial epithelial cells BEAS-2B, human lung cancer cells SPCA1, SPC-A-1-BM | miR-499a-5p | Inhibition of miR-499a-5p by antagomirs restrained tumor growth in vivo (p < 0.01) |
| McCann et al. (2019) [75] | Lung cancer | Animals: mice models. Cells: primary endothelial cells isolated from normal or tumor tissue from mice | miR-30c | miR-30c antagomiRs promoted PAI-1–dependent tumor growth and increased fibrin abundance (p < 0.05) |
| Bronchial Hypersensitivity | ||||
|---|---|---|---|---|
| Humans | ||||
| Study | Disease | Sample | miRNAs | Findings |
| Baker et al. (2016) [37] | COPD | Peripheral lung samples from COPD patients and controls; airway epithelial cells | miR-34a | miR-34a antagomirs increased SIRT1 (p < 0.01)/-6 (p < 0.05) mRNA levels, decreasing cellular senescence markers in COPD (p < 0.05) |
| Hsu et al. (2016) [76] | COPD | Five COPD, five smokers, five controls | miR-132 | Ectopic expression of PKR or miR-132 antagomiR alone failed to restore IFN-β induction (p > 0.05), co-treatment increased avSG formation, induction of p300, and IFN-β in COPD pBECs (p < 0.05) |
| Jiang et al. (2018) [49] | COPD | Humans: 73 patients with PH, 32 controls. Animals: hypoxia-induced PH mice | miR-190a-5p | Antagomir-190a-5p reduced right ventricular systolic pressure (p < 0.01) and enhanced KLF15 expression (p < 0.0001) in lung tissue |
| Baker et al. (2019) [38] | COPD | 30 COPD/18 controls: lung tissue from tissue bank; 14 COPD, 10 non-smoking controls: human primary SAECs cultured; 13 COPD, five controls: sputum samples collected | miR-570-3p | Inhibition of elevated miR-570-3p in COPD small airway epithelial cells, using an antagomir, restores sirtuin-1, and suppresses markers of cellular senescence, restoring cellular growth (p < 0.05) |
| Animals | ||||
| Study | Disease | Sample | miRNAs | Findings |
| Collison et al. (2011) [24] | Allergic airway disease | BALB/c mice sensitized with house dust mite | miR-145, miR-21, let-7b | Inhibition of miR-145 (p < 0.05), but not miR-21 or let-7b (both p > 0.05), inhibited eosinophilic inflammation, mucus hypersecretion, TH2 cytokine production, and airway hyper-responsiveness |
| Li et al. (2015) [50] | Airway hyper-responsiveness | Wild-type specific pathogen-free BALB/c mice | miR-9 | AntagomiR-9 increased PP2A activity and GR nuclear translocation in macrophages (p < 0.05), restored steroid sensitivity in steroid-resistant airway hyper-responsiveness |
| Plank et al. (2015) [30] | Asthma | Specific pathogen-free BALB/c mice | miR-155-5p | Antagomir administration reduced miR-155-5p expression (p < 0.01), but failed to alter the disease phenotype (p > 0.05). It exhibits poor uptake in lymphocytes |
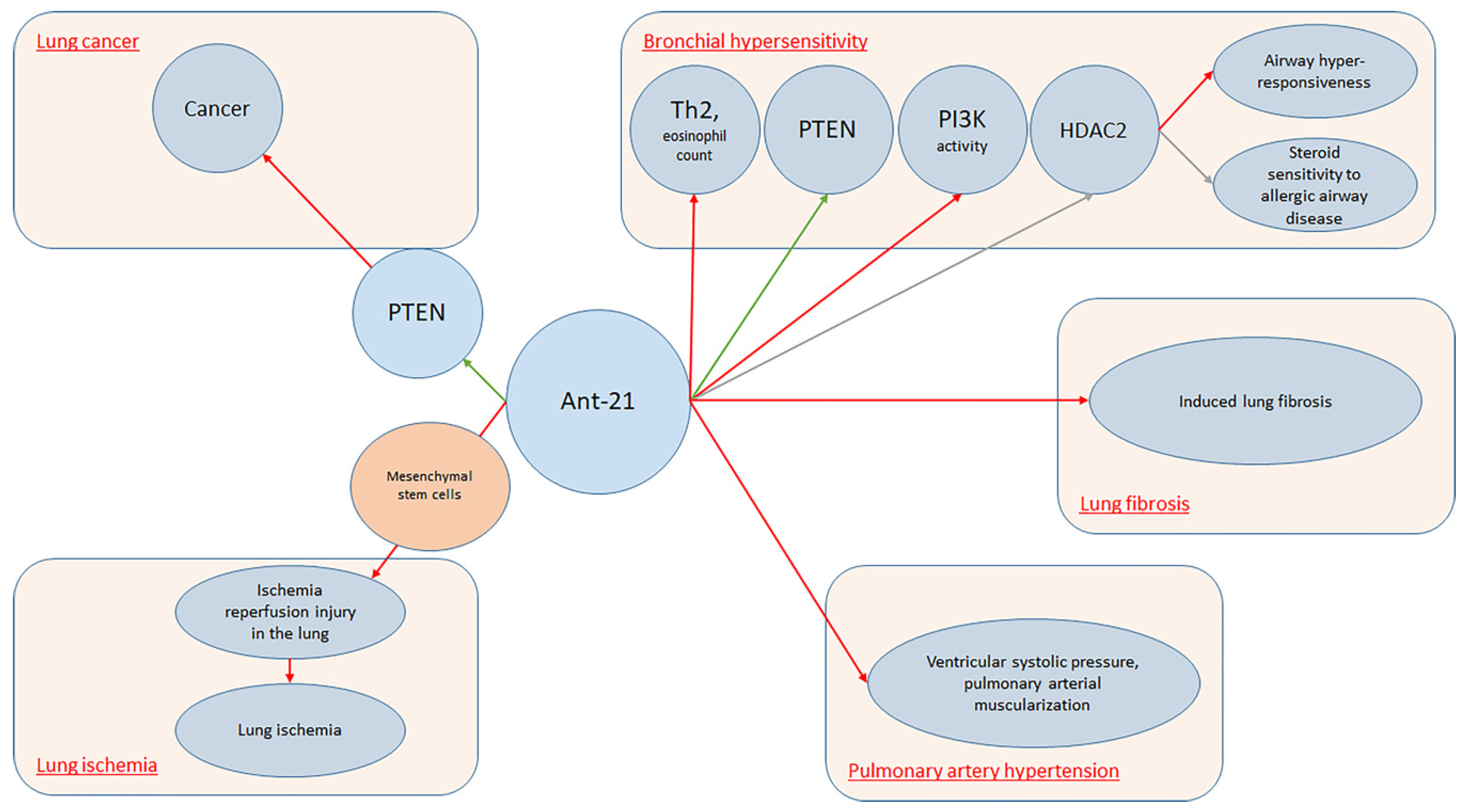
| Kim et al. (2017) [27] | Asthma | BALB/c mice | miR-21 | Antagomir-21 increased PTEN levels (p < 0.05). Treatment with Ant-21 reduced PI3K activity and restored HDAC2 levels (p < 0.05), suppressing airway hyper-responsiveness and restoring steroid sensitivity to allergic airway disease |
| Lee et al. (2017) [35] | Acute bronchial asthma | BALB/c mice sensitized and challenged with ovalbumin | miR-21 | MiR-21 expression down-regulated in mice lungs treated with anti-miR-21. It reduced total cell (p < 0.001) and eosinophil counts (p < 0.01) in BAL fluid and the levels of IL-5 and IL-13 (p < 0.05) |
| Jiang et al. (2018) [49] | COPD | Humans: 73 patients with PH, 32 controls. Animals: hypoxia-induced PH mice | miR-190a-5p | Antagomir-190a-5p reduced right ventricular systolic pressure (p < 0.01) and enhanced the KLF15 expression levels (p < 0.0001) in lung tissue |
| Cell Lines | ||||
| Study | Disease | Sample | miRNAs | Findings |
| Baker et al. (2016) [37] | COPD | Peripheral lung samples from COPD patients and controls; airway epithelial cells | miR-34a | miR-34a antagomirs increased SIRT1 (p < 0.01)/-6 (p < 0.05) mRNA levels, decreasing markers of cellular senescence in airway epithelial cells from COPD (p < 0.05) |
| Hsu et al. (2016) [76] | COPD | Five COPD, five smokers, five controls | miR-132 | Ectopic expression of PKR or miR-132 antagomiR alone failed to restore IFN-β induction (p > 0.05), co-treatment increased avSG formation, induction of p300 and IFN-β in COPD pBECs (p < 0.05) |
| Baker et al. (2019) [38] | COPD | 30 COPD/18 controls: lung tissue from a tissue bank; 14 COPD, 10 non-smoking controls: human primary SAECs cultured; 13 COPD, five controls: sputum samples collected | miR-570-3p | Inhibition of elevated miR-570-3p in COPD small airway epithelial cells, using an antagomir, restores sirtuin-1 and suppresses markers of cellular senescence, restoring cellular growth (p < 0.05) |
| Pulmonary Hypertension | ||||
|---|---|---|---|---|
| Humans | ||||
| Study | Disease | Sample | miRNAs | Findings |
| Potus et al. (2014) [77] | Pulmonary hypertension | Humans: percutaneous biopsy of vastus lateralis (n = 11 patients, n = 9 controls). Animals: male Sprague-Dawley rats. Cells: CD311 cells isolated from two idiopathic PAH, two heritable PAH, and three control quadriceps biopsies | miR-126 | AntagomiR-126 in healthy CD31+ cells mimicked the PAH phenotype. In skeletal muscle of healthy rats, it decreased muscle capillarity (p < 0.05) and exercise tolerance in treadmill tests (p < 0.05) |
| Animals | ||||
| Study | Disease | Sample | miRNAs | Findings |
| Pullamsetti et al. (2012) [25] | Pulmonary hypertension | Animals: mice and rat models. Cells: pooled human umbilical vein ECs and human pulmonary artery SMCs | miR-17, miR-21, miR-92a | Ant-17 and Ant-21 reduced right ventricular systolic pressure, all antagomirs decreased pulmonary arterial muscularization. Ant-17 reduced hypoxia-induced right ventricular hypertrophy, improved pulmonary artery acceleration time. In rats, Ant-17 decreased right ventricular systolic pressure and total pulmonary vascular resistance index, increased pulmonary artery acceleration time, normalized cardiac output, and decreased pulmonary vascular remodeling. In human pulmonary artery smooth muscle cells, Ant-17 increased p21 |
| Brock et al. (2014) [39] | Pulmonary hypertension | Animals: four mice samples (three in hypoxic condition, one control). In vitro: HPASMCs | miR-20a | Animals: AntagomiR-20a enhanced BMPR2 expression levels in lung tissues by 59.3% (p < 0.001), reduced wall thickness (p < 0.01), luminal occlusion of small pulmonary arteries (p < 0.001) and right ventricular hypertrophy (p < 0.01). In vitro: Transfection of HPASMCs with antimiR-20a activates downstream targets of BMPR2 increasing activation of Id-1 and Id-2 (p < 0.05). HPASMCs proliferation reduced upon transfection with antagomiR-20a (p < 0.05) |
| Potus et al. (2014) [77] | Pulmonary hypertension | Humans: percutaneous biopsy of vastus lateralis (n = 11 patients, n = 9 controls). Animals: male Sprague-Dawley rats. Cells: CD311 cells isolated from two idiopathic PAH, two heritable PAH, and three control quadriceps biopsies | miR-126 | AntagomiR-126 in healthy CD31+ cells mimicked the PAH phenotype. In skeletal muscle of healthy rats, it decreased muscle capillarity (p < 0.05) and exercise tolerance in treadmill tests (p < 0.05) |
| Sharma et al. (2015) [57] | Pulmonary hypertension | Male Sprague-Dawley rats | miR-206 | Knockdown of miR-206 reduced right ventricular pressure and right ventricular hypertrophy index |
| Gubrij et al. (2016) [78] | Pulmonary hypertension | MCT-PAH rats | miR-223 | A223 reduced levels of miR-223 in pulmonary artery and lungs of MCT PAH rats as compared to controls (p < 0.05), but did not attenuate MCT PAH (p > 0.05) |
| Mondejar-Parreño et al. (2019) [51] | Pulmonary hypertension | Pathogen-free male Wistar rats | miR-1 | AntagomiR-1 prevented (p < 0.05) hypoxia-induced decline in voltage-dependent potassium channel Kv1.5 currents |
| Cell Lines | ||||
| Study | Disease | Sample | miRNAs | Findings |
| Pullamsetti et al. (2012) [25] | Pulmonary hypertension | Animals: mice and rat models. Cells: pooled human umbilical vein ECs and human pulmonary artery SMCs | miR-17, miR-21, miR-92a | Ant-17 and Ant-21 reduced right ventricular systolic pressure, all antagomirs decreased pulmonary arterial muscularization. Ant-17 reduced hypoxia-induced right ventricular hypertrophy, improved pulmonary artery acceleration time. In rats, Ant-17 decreased right ventricular systolic pressure and total pulmonary vascular resistance index, increased pulmonary artery acceleration time, normalized cardiac output, and decreased pulmonary vascular remodeling. In human pulmonary artery smooth muscle cells, Ant-17 increased p21 |
| Brock et al. (2014) [39] | Pulmonary hypertension | Animals: four mice samples (three in hypoxic condition, one control). In vitro: HPASMCs | miR-20a | Animals: AntagomiR-20a enhanced BMPR2 expression levels in lung tissues by 59.3% (p < 0.001), reduced wall thickness (p < 0.01), luminal occlusion of small pulmonary arteries (p < 0.001) and right ventricular hypertrophy (p < 0.01). In vitro: Transfection of HPASMCs with antimiR-20a activates downstream targets of BMPR2 increasing activation of Id-1 and Id-2 (p < 0.05). HPASMCs proliferation reduced upon transfection with antagomiR-20a (p < 0.05) |
| Potus et al. (2014) [77] | Pulmonary hypertension | Humans: percutaneous biopsy of vastus lateralis (n = 11 patients, n = 9 controls). Animals: male Sprague-Dawley rats. Cells: CD311 cells isolated from two idiopathic PAH, two heritable PAH, and three control quadriceps biopsies | miR-126 | AntagomiR-126 in healthy CD31+ cells mimicked the PAH phenotype. In skeletal muscle of healthy rats, it decreased muscle capillarity (p < 0.05) and exercise tolerance in treadmill tests (p < 0.05) |
| Lung Injury | ||||
|---|---|---|---|---|
| Animals | ||||
| Study | Disease | Sample | miRNAs | Findings |
| Xu et al. (2014) [68] | Lung injury | Animals: healthy male C57BL/6 mice. Cells: Epithelial cells | miR-17 | miR-17 antagomir increased the expression of FoxA1 in Acute Lung Injury mice (p < 0.05) |
| Yuan et al. (2015) [32] | Lung inflammation | Animals: male wild-type C57BL/6J mice. Cells: bone marrow-derived macrophages | miR-155 | Increased expression of miR155 by mTREM-1 suppressed by antimiR-155 (p < 0.05) |
| Fu et al. (2018) [45] | Pulmonary inflammation | Animals: male BALB/c mice. Cells: murine macrophage RAW264.7 cells | miR-92a | Antagomir-92a reduced pathological changes associated with lung inflammation, reduces lung wet/dry ratio (p < 0.01), and Evans blue dye extravasation (p < 0.01). Inhibition of miR-92a reduced the repression of TNF-α, IL-1β, IL-6 (p < 0.01) in lung tissues |
| Wu et al. (2018) [67] | Acute lung injury | MK2 deficient mice (C57BL/6) (B6.129P2-Mapkapk2tm1Dgen/J, and MK2flox/flox mice | Let-7e | Transfection of anti-let-7e into MK2-/- BMDM rescued LPS-induced expression of TNF-α, IL-6, and MIP-2 (p < 0.05) |
| Xie et al. (2018) [79] | Lung inflammation, lung injury | Animals: male C57BL/6 mice. Cells: RAW264.7 cells | miR-34b-5p | miR-34b-5p antagomir in vivo inhibited miR-34b-5p up-regulation, reduced inflammatory cytokine release, decreased alveolar epithelial cell apoptosis, attenuated lung inflammation, improved survival by targeting PGRN during acute lung injury (p < 0.05) |
| Huang et al. (2019) [47] | Acute Lung Injury | Sixty healthy male-specific pathogen free C57BL/6 mice | miR-27b | Downregulation of miR-27b decreased the levels of IL-1β, IL-6, and TNF-α in BALF of Acute Lung Injury mice (p < 0.05) |
| Cell Lines | ||||
| Study | Disease | Sample | miRNAs | Findings |
| Adyshev et al. (2013) [80] | Lung injury | Human pulmonary artery endothelial cells | hsa-miR-374a, hsa-miR-374b, hsa-miR-520c-3p, hsa-miR-1290 | Antagomirs for each MYLK miRNA increased 3′UTR luciferase activity (1.2–2.3 FI) and rescued the decreased MLCK-3′UTR reporter activity produced by miRNA mimics (70%–110% increases for each miRNA; p < 0.05) |
| Adyshev et al. (2014) [81] | Lung inflammation | Human pulmonary artery endothelial cells | hsa-miR-374a, hsa-miR-568 | Antagomirs for each PBEF/NAMPT miRNA increased the endogenous PBEF/NAMPTmRNA and protein levels and 3′-UTR luciferase activity compared with controls (p < 0.05) |
| Xu et al. (2014) [68] | Lung injury | Animals: healthy male C57BL/6 mice. Cells: epithelial cells | miR-17 | miR-17 antagomir increased the expression of FoxA1 in Acute Lung Injury mice (p < 0.05) |
| Yuan et al. (2015) [32] | Lung inflammation | Animals: Male wild-type C57BL/6J mice. Cells: bone marrow-derived macrophages | miR-155 | Increased expression of miR155 by mTREM-1 suppressed by antagomir against miR-155 (p < 0.05) |
| Fu et al. (2018) [45] | Pulmonary inflammation | Animals: male BALB/c mice. Cells: murine macrophage RAW264.7 cells | miR-92a | Antagomir-92a reduced pathological changes associated with lung inflammation, reduces lung wet/dry ratio (p < 0.01), and Evans blue dye extravasation (p < 0.01). Inhibition of miR-92a ameliorated the inflammatory response by reducing the repression of TNF-α, IL-1β, IL-6 (p < 0.01) in lung tissues |
| Xie et al. (2018) [79] | Lung inflammation, lung injury | Animals: male C57BL/6 mice. Cells: RAW264.7 cells | miR-34b-5p | miR-34b-5p antagomir in vivo inhibited miR-34b-5p up-regulation, reduced inflammatory cytokine release, decreased alveolar epithelial cell apoptosis, attenuated lung inflammation, improved survival by targeting PGRN during acute lung injury (p < 0.05) |
| Other Conditions | ||||
|---|---|---|---|---|
| Humans | ||||
| Study | Disease | Sample | miRNAs | Findings |
| Chatterjee et al. (2014) [41] | Lung cell dysfunction | HLMECs, HUVECs | miR-147b | MiR-147b antagomir increased total and cell surface expression of ADAM15 in endothelial cells (p < 0.05) |
| Ge et al. (2016) [82] | Lung fibrosis | Humans: bronchial epithelia from lung transplant patients. Cells: primary fibroblasts isolated from human lungs | miR-323a-3p | Antagomirs for miR-323a-3p augment murine lung fibrosis after bleomycin injury (p < 0.05) |
| Sharma et al. (2018) [58] | HIV infection and substance abuse | Human monocyte derived macrophages, HPASMCs | miR-130a | Transfection of HPASMCs with antagomir-130a–ameliorated the extracellular vesicles-induced effect (p < 0.001) |
| Yuan et al. (2018) [69] | Tuberculosis | Fifty patients, 20 controls. Monocytes isolated from peripheral blood mononuclear cells | miR-196b-5p | antagomir-196b-5p promoted Bacillus Calmette–Guérin uptake in MDMs or differentiated U937 cells (p < 0.05) |
| Animals | ||||
| Study | Disease | Sample | miRNAs | Findings |
| Krützfeldt et al. (2005) [7] | Various conditions | Mice models | miR-16, miR-122, miR-192, miR-194 | Intravenous administration of antagomirs reduced miRNA levels in liver, lung, kidney, heart, intestine, fat, skin, bone marrow, muscle, ovaries, and adrenals |
| Chiba et al. (2009) [83] | Abnormal BSM contraction | BSM cells, bronchial tissues of BALB/c mice | miR-133a | Up-regulation of RhoA when endogenous miR-133a function inhibited by its antagomir in hBSMCs (p < 0.05). No effect (p > 0.05) of miR-133b and let-7a antagomirs |
| Pandit et al. (2010) [84] | Pulmonary Fibrosis | Animals: mice models. Cells: 10 Idiopathic Pulmonary Fibrosis, 10 control tissues | Let-7d | Let-7d antagomir decreased expression of CDH1 and TJP1 and increased COL1A1 and HMGA2 expression in the lungs (p < 0.05) |
| Rosenberger et al. (2012) [56] | Influenza | C57Bl/6, MyD88null mice | miR-451 | Three types of primary dendritic cells treated with antagomirs against miR-451 secreted elevated levels of IL-6 (p< 0.01), TNF (p < 0.05), CCL5/RANTES (p < 0.05), and CCL3/MIP1α (p < 0.01) |
| Asquith et al. (2014) [85] | Chronic ethanol consumption | From non-human primates: PBMC, mesenteric and tracheobronchial lymph nodes, jejunum, duodenum, ileum, and descending colon | miR-181a, miR-221 (in PBMC), miR-155 (in colon) | Transfection of miRNA antagomirs upregulated both STAT-3 (p < 0.05)/ARNT (p < 0.001), VEGF (p < 0.05)/HGF (p < 0.01)/G-CSF (p < 0.05) |
| Zhang et al. (2015) [52] | Various disorders | Animals: BALB/c mice. Cells: 4T1 murine breast cancer cells | miR-10b | Antagomir-10b and PTX delivered by D-Lip delays the growth of 4T1 tumors and reduce lung metastases; Hoxd10 expression in tumors up-regulated (p < 0.01) |
| Zhou et al. (2015) [54] | Influenza | Animals: six groups of mice (five mice per group), including control group. Cells: MDCK cells | miR-2911 | Inhibitory effect of HS decoction on viral replication abolished by anti-miR2911 (p < 0.05) |
| Podsiad et al. (2015) [31] | Pneumonia | Animals: wild-type C57BL/6 mice. Cells: Human lung macrophages | miR-155 | miR-155 antagomir improved lung bacterial clearance by 4.2-fold |
| Zhou et al. (2016) [86] | Systemic Lupus Erythematosus | Animals: C57BL/6J (B6) and B6.Cg-Mir155tm1.1Rsky/J mice. Cells: Hepa 1-6 cells | miR-155 | Disease progression reduced by 20% by in vivo using of antimiR-155 |
| Ma et al. (2017) [71] | Hypoxia | Animals: adult male Wistar rats. Cells: PASMC cultured | miR-125a | miR-125a antagomir mimicked the hypoxic damage effects to mitochondrial homeostasis (p < 0.05) |
| Morales et al. (2017) [87] | SARS-CoV | Animals: female mice. Cells: mouse delayed brain tumor cells expressing the murine SARS-CoV receptor ACE2 | svRNA-nsp3.1, svRNA-nsp3.2, svRNA-N, miR-877 | Antagomirs reduced partially (svRNA-nsp3.1), or totally (svRNA-nsp3.2, svRNA-N, miR-877), the luciferase activity |
| Zhou et al. (2017) [88] | Influenza | Animals: beagles. Cells: MDCK cells | cfa-miR-143 | Anti-cfa-miR-143 caused upregulation of Igfbp5 in CIV-infected MDCK cells |
| Fehl et al. (2019) [44] | Bronchopulmonary dysplasia | Newborn C57BL/6J mice | N/A | AntagomiRs impacted lung volume (p < 0.05), septal thickness (p < 0.01), and the transcriptome (p < 0.05) of developing mouse lungs |
| Li et al. (2019) [28] | Lung ischemia | Mail C57/BL6 mice | miR-21-5p | Pre-treatment of MSCs with miR-21-5p antagomir decreased miR-21-5p expression level in exosomes secreted |
| Tamgue et al. (2019) [89] | Tuberculosis | Bone marrow-derived macrophages generated from male BALB/c mice | miR-143, miR-365 | Antagomirs for miR-143 and miR-365 decreased the intracellular growth of Mtb HN878, reduced the production of IL-6 (p < 0.001) and CCL5 (p < 0.01 for miR-143, p < 0.05 for miR-365) and promoted the apoptotic death of Mtb HN878-infected BMDMs (p < 0.01 for miR-143, p < 0.05 for miR-365) |
| Zhang et al. (2019) [90] | Influenza | Animals: C57BL/6 mice. Cells: human pulmonary epithelial cell line A549 | miR-146a | Downregulation of miR-146a inhibits Influenza A Virus replication by enhancing type I IFN response through TRAF6 in vitro and in vivo (p < 0.01) |
| Cell Lines | ||||
| Study | Disease | Sample | miRNAs | Findings |
| Chiba et al. (2009) [83] | Abnormal BSM contraction | BSM cells, bronchial tissues of BALB/c mice | miR-133a | Up-regulation of RhoA when endogenous miR-133a function inhibited by its antagomir in hBSMCs (p < 0.05). No effect (p > 0.05) of miR-133b and let-7a antagomirs |
| Pandit et al. (2010) [84] | Pulmonary Fibrosis | Animals: mice models. Cells: 10 Idiopathic Pulmonary Fibrosis and 10 control tissues | Let-7d | Let-7d antagomir decreased expression of CDH1 and TJP1, and increased COL1A1 and HMGA2 expression in the lungs (p < 0.05) |
| Bhattacharyya et al. (2011) [29] | Cystic Fibrosis | Lung epithelial cells | miR-155 | Antagomir-155 in CF cells down-regulates miR-155 expression by 85%; IL-8 mRNA levels decreased of 70% and IL-8 protein levels by 11-fold |
| Chatterjee et al. (2014) [41] | Lung cell dysfunction | HLMECs, HUVECs | miR-147b | MiR-147b antagomir increased total and cell surface expression of ADAM15 in endothelial cells (p < 0.05) |
| Fabbri et al. (2014) [43] | Cystic Fibrosis | CF bronchial epithelial IB3-1 cells infected by Pseudomonas aeruginosa | miR-93 | IL-8 up-regulation in uninfected cells treated with antagomiR-93 (p < 0.01) |
| Zhang et al. (2015) [52] | Various disorders | Animals: BALB/c mice. Cells: 4T1 murine breast cancer cells | miR-10b | Antagomir-10b and PTX delivered by D-Lip delays the growth of 4T1 tumors and reduce the lung metastases; up-regulated Hoxd10 in tumors (p < 0.01) |
| Zhou et al. (2015) [54] | Influenza | Animals: six groups of mice (five mice per group), including control group. Cells: MDCK cells | miR-2911 | Inhibitory effect of HS decoction on viral replication abolished by anti-miR2911 (p < 0.05) |
| Ge et al. (2016) [82] | Lung fibrosis | Humans: bronchial epithelia from lung transplant patients. Cells: primary fibroblasts from human lung explants | miR-323a-3p | Antagomirs for miR-323a-3p augment murine lung fibrosis after bleomycin injury (p < 0.05) |
| Podsiad et al. (2015) [31] | Pneumonia | Animals: wild-type C57BL/6 mice. Cells: human lung macrophages | miR-155 | AntimiR-155 improved lung bacterial clearance by 4.2-fold compared with controls |
| Zhou et al. (2016) [86] | Systemic Lupus Erythematosus | Animals: C57BL/6J (B6) and B6.Cg-Mir155tm1.1Rsky/J mice. Cells: Hepa 1-6 cells | miR-155 | Disease progression, reduced by 20% by in vivo silencing of miR-155 using antimiR-155 |
| Bartoszewska et al. (2017) [36] | Hypoxia | Hypoxia-induced human airway epithelial cell lines Calu-3 and 16HBE14o-; normal primary bronchial epithelial cells | miR-200b | Manipulation of miRNA levels during normoxia and hypoxia by antagomirs increased CFTR mRNA levels (p < 0.05) |
| Ma et al. (2017) [71] | Hypoxia | Animals: adult male Wistar rats. Cells: PASMC cultured | miR-125a | miR-125a antagomir mimicked the hypoxic damage effects to mitochondrial homeostasis (p < 0.05) |
| Morales et al. (2017) [87] | SARS-CoV | Animals: Female mice. Cells: mouse delayed brain tumor cells expressing the murine SARS-CoV receptor ACE2 | svRNA-nsp3.1, svRNA-nsp3.2, svRNA-N, miR-877 | Antagomirs reduced partially (svRNA-nsp3.1), or totally (svRNA-nsp3.2, svRNA-N, miR-877), the luciferase activity |
| Shentu et al. (2017) [26] | Lung fibrosis | Human bone marrow-derived Mesenchymal Stem Cells | miR-199a/b-3p, 21-5p, 630, 22-3p, 196a-5p, 199b-5p, 34a-5p, and 148a-3p | AntagomiR-630 abrogated the effect of extracellular vesicles on CDH2 expression (p < 0.05) |
| Zhou et al. (2017) [88] | Influenza | Animals: beagles. Cells: MDCK cells | cfa-miR-143 | Anti-cfa-miR-143 caused upregulation of Igfbp5 in CIV-infected MDCK cells |
| Sharma et al. (2018) [58] | HIV infection and substance abuse | Human monocyte derived macrophages, HPASMCs | miR-130a | Transfection of HPASMCs with antagomir-130a ameliorated the extracellular vesicles-induced effect (p < 0.001) |
| Yuan et al. (2018) [69] | Tuberculosis | Fifty patients, 20 controls. Monocytes isolated from peripheral blood mononuclear cells | miR-196b-5p | Antagomir-196b-5p promoted Bacillus Calmette–Guérin uptake in MDMs or differentiated U937 cells (p < 0.05) |
| Tamgue et al. (2019) [89] | Tuberculosis | Bone marrow-derived macrophages generated from male BALB/c mice | miR-143, miR-365 | Antagomirs for miR-143 and miR-365 decreased the intracellular growth of Mtb HN878, reduced the production of IL-6 (p < 0.001) and CCL5 (p < 0.01 for miR-143, p < 0.05 for miR-365), and promoted the apoptotic death of Mtb HN878-infected BMDMs (p < 0.01 for miR-143, p < 0.05 for miR-365) |
| Zhang et al. (2019) [90] | Influenza | Animals: C57BL/6 mice. Cells: human pulmonary epithelial cell line A549 | miR-146a | Downregulation of miR-146a inhibits Influenza A Virus replication by enhancing type I IFN response through TRAF6 in vitro and in vivo (p < 0.01) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Murdaca, G.; Tonacci, A.; Negrini, S.; Greco, M.; Borro, M.; Puppo, F.; Gangemi, S. Effects of AntagomiRs on Different Lung Diseases in Human, Cellular, and Animal Models. Int. J. Mol. Sci. 2019, 20, 3938. https://doi.org/10.3390/ijms20163938
Murdaca G, Tonacci A, Negrini S, Greco M, Borro M, Puppo F, Gangemi S. Effects of AntagomiRs on Different Lung Diseases in Human, Cellular, and Animal Models. International Journal of Molecular Sciences. 2019; 20(16):3938. https://doi.org/10.3390/ijms20163938
Chicago/Turabian StyleMurdaca, Giuseppe, Alessandro Tonacci, Simone Negrini, Monica Greco, Matteo Borro, Francesco Puppo, and Sebastiano Gangemi. 2019. "Effects of AntagomiRs on Different Lung Diseases in Human, Cellular, and Animal Models" International Journal of Molecular Sciences 20, no. 16: 3938. https://doi.org/10.3390/ijms20163938
APA StyleMurdaca, G., Tonacci, A., Negrini, S., Greco, M., Borro, M., Puppo, F., & Gangemi, S. (2019). Effects of AntagomiRs on Different Lung Diseases in Human, Cellular, and Animal Models. International Journal of Molecular Sciences, 20(16), 3938. https://doi.org/10.3390/ijms20163938