Cadmium and Plant Development: An Agony from Seed to Seed

 ,
,
Abstract
:1. Introduction
2. Seed Germination
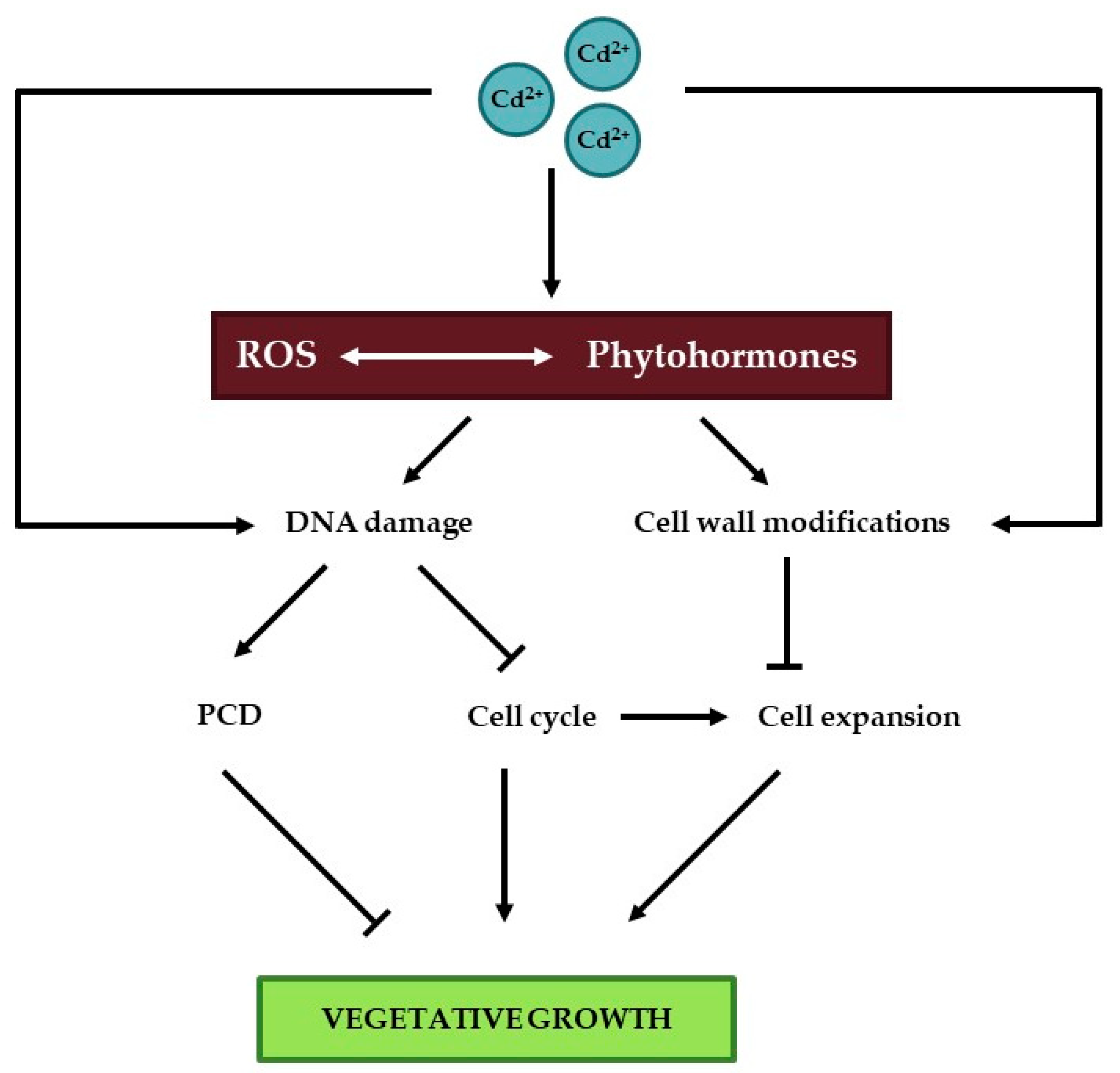
3. Vegetative Plant Growth
3.1. The DNA Damage Response
3.1.1. DNA Damage
3.1.2. The Cell Cycle
3.1.3. Cell Death
3.2. The Cell Wall
3.2.1. The Cell Wall as Major Storage Compartment for Cadmium
3.2.2. Cadmium-Induced Cell Wall Modifications
3.2.3. The Role of The Cell Wall in Cadmium Hyperaccumulators
4. Reproductive Growth
5. Conclusion and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ABA | Abscisic acid |
| ABI3 | Abscisic acid insensitive 3 |
| ACC | 1-aminocyclopropane-1-carboxylate |
| ACS | ACC synthase |
| AFLP | Amplified fragment length polymorphism |
| AsA | Ascorbate |
| AUX | Auxin |
| ATM | Ataxia telangiectasia mutated |
| ATR | RAD3-related |
| Ca | Calcium |
| CAT | Catalase |
| Cd | Cadmium |
| CDKs | Cyclin-dependent kinases |
| Cr | Chromium |
| CK | Cytokinin |
| DDR | DNA damage response |
| DSBs | Double-stranded breaks |
| EdU | 5-ethynyl-2′-deoxyuridine |
| ET | Ethylene |
| EXP | Expansin |
| FCM | Flow cytometry |
| Fe | Iron |
| FT-IR | Fourier transform infrared |
| GA | Gibberellic acid |
| GR | Glutathione reductase |
| Grx | Glutaredoxin |
| GSH | Glutathione |
| GTS | Genomic template stability |
| H2O2 | Hydrogen peroxide |
| HG | Homogalacturonan |
| IAA | Indole-3-acetic acid |
| ISSR | Inter-simple sequence repeat |
| JA | Jasmonic acid |
| Mn | Manganese |
| NAC | NO APICAL MERISTEM/ARABIDOPSIS TRANSCRIPTION ACTIVATION FACTOR/CUP-SHAPED COTYLEDON |
| NADPH | Nicotinamide adenine dinucleotide phosphate |
| •OH | Hydroxyl radical |
| O2• | Superoxide |
| Pb | Lead |
| PC | Phytochelatin |
| PCD | Programmed cell death |
| PME | Pectin methylesterase |
| POD | Class III peroxidase |
| Prx | Peroxiredoxin |
| qRT-PCR | Quantitative reverse transcription polymerase chain reaction |
| RAPD | Random amplified polymorphic DNA |
| ROS | Reactive oxygen species |
| RT-PCR | Reverse transcription polymerase chain reaction |
| SA | Salicylic acid |
| SAGs | Senescence-associated genes |
| SMR | SIAMESE-related |
| SOD | Superoxide dismutase |
| SOG1 | SUPPRESSOR OF GAMMA RESPONSE 1 |
| SRAP | Sequence-related amplified polymorphism |
| SSBs | Single-stranded breaks |
| SSR | Simple sequence repeat |
| SUSY | Sucrose synthase |
| TFs | Transcription factors |
| Trx | Thioredoxin |
| XTH | Endotransglucosylases/hydrolases |
| Zn | Zinc |
References
- Khan, M.A.; Khan, S.; Khan, A.; Alam, M. Soil contamination with cadmium, Consequences and remediation using organic amendments. Sci. Total Environ. 2017, 601, 1591–1605. [Google Scholar] [CrossRef] [PubMed]
- Verbruggen, N.; Hermans, C.; Schat, H. Mechanisms to cope with arsenic or cadmium excess in plants. Curr. Opin. Plant Biol. 2009, 12, 364–372. [Google Scholar] [CrossRef] [PubMed]
- DalCorso, G.; Farinati, S.; Maistri, S.; Furini, A. How plants cope with cadmium: Staking all on metabolism and gene expression. J. Integr. Plant Biol. 2008, 50, 1268–1280. [Google Scholar] [CrossRef] [PubMed]
- Cuypers, A.; Keunen, E.; Bohler, S.; Jozefczak, M.; Opdenakker, K.; Gielen, H.; Vercampt, H.; Bielen, A.; Schellingen, K.; Vangronsveld, J.; et al. Cadmium and copper stress induce a cellular oxidative challenge leading to damage versus signalling. In Metal Toxicity in Plants: Perception, Signaling and Remediation; Gupta, D.K., Sandalio, L.M., Eds.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 65–90. [Google Scholar] [CrossRef]
- Sharma, S.S.; Dietz, K.J. The relationship between metal toxicity and cellular redox imbalance. Trends Plant Sci. 2009, 14, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Cuypers, A.; Plusquin, M.; Remans, T.; Jozefczak, M.; Keunen, E.; Gielen, H.; Opdenakker, K.; Nair, A.R.; Munters, E.; Artois, T.J.; et al. Cadmium stress: An oxidative challenge. Biometals 2010, 23, 927–940. [Google Scholar] [CrossRef] [PubMed]
- Jozefczak, M.; Keunen, E.; Schat, H.; Bliek, M.; Hernandez, L.E.; Carleer, R.; Remans, T.; Bohler, S.; Vangronsveld, J.; Cuypers, A. Differential response of Arabidopsis leaves and roots to cadmium: Glutathione-related chelating capacity vs antioxidant capacity. Plant Physiol. Biochem. 2014, 83, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Schutzendubel, A.; Polle, A. Plant responses to abiotic stresses: Heavy metal-induced oxidative stress and protection by mycorrhization. J. Exp. Bot. 2002, 53, 1351–1365. [Google Scholar] [CrossRef] [PubMed]
- Cuypers, A.; Hendrix, S.; dos Reis, R.A.; De Smet, S.; Deckers, J.; Gielen, H.; Jozefczak, M.; Loix, C.; Vercampt, H.; Vangronsveld, J.; et al. Hydrogen peroxide, Signaling in disguise during metal phytotoxicity. Front. Plant Sci. 2016, 7, 25. [Google Scholar] [CrossRef] [PubMed]
- Moller, I.M.; Jensen, P.E.; Hansson, A. Oxidative modifications to cellular components in plants. Annu. Rev. Plant Biol. 2007, 58, 459–481. [Google Scholar] [CrossRef] [PubMed]
- Petrov, V.D.; Van Breusegem, F. Hydrogen peroxide-A central hub for information flow in plant cells. AoB Plants 2012. [Google Scholar] [CrossRef] [PubMed]
- Bucker-Neto, L.; Paiva, A.L.S.; Machado, R.D.; Arenhart, R.A.; Margis-Pinheiro, M. Interactions between plant hormones and heavy metals responses. Genet. Mol. Biol. 2017, 40, 373–386. [Google Scholar] [CrossRef] [PubMed]
- De Smet, S.; Cuypers, A.; Vangronsveld, J.; Remans, T. Gene networks involved in hormonal control of root development in Arabidopsis thaliana: A framework for studying its disturbance by metal stress. Int. J. Mol. Sci. 2015, 16, 19195–19224. [Google Scholar] [CrossRef] [PubMed]
- Schellingen, K.; Van Der Straeten, D.; Vandenbussche, F.; Prinsen, E.; Remans, T.; Vangronsveld, J.; Cuypers, A. Cadmium-induced ethylene production and responses in Arabidopsis thaliana rely on ACS2 and ACS6 gene expression. BMC Plant Biol. 2014, 14, 14. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Serrano, M.; Romero-Puertas, M.C.; Pazmino, D.M.; Testillano, P.S.; Risueno, M.C.; del Rio, L.A.; Sandalio, L.M. Cellular response of pea Plants to cadmium toxicity: Cross talk between reactive oxygen species, nitric oxide, and calcium. Plant Physiol. 2009, 150, 229–243. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Khan, A.L.; Kim, D.H.; Lee, S.Y.; Kim, K.M.; Waqas, M.; Jung, H.Y.; Shin, J.H.; Kim, J.G.; Lee, I.J. Silicon mitigates heavy metal stress by regulating P-type heavy metal ATPases, Oryza sativa low silicon genes, and endogenous phytohormones. BMC Plant Biol. 2014, 14, 13. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.F.; Zhou, G.Y.; Na, X.F.; Yang, L.J.; Nan, W.B.; Liu, X.; Zhang, Y.Q.; Li, J.L.; Bi, Y.R. Cadmium interferes with maintenance of auxin homeostasis in Arabidopsis seedlings. J. Plant Physiol. 2013, 170, 965–975. [Google Scholar] [CrossRef]
- Ronzan, M.; Piacentini, D.; Fattorini, L.; Della Rovere, F.; Eiche, E.; Rieman, M.; Altamura, M.M.; Falasca, G. Cadmium and arsenic affect root development in Oryza sativa L. negatively interacting with auxin. Environ. Exp. Bot. 2018, 151, 64–75. [Google Scholar] [CrossRef]
- Xia, X.J.; Zhou, Y.H.; Shi, K.; Zhou, J.; Foyer, C.H.; Yu, J.Q. Interplay between reactive oxygen species and hormones in the control of plant development and stress tolerance. J. Exp. Bot. 2015, 66, 2839–2856. [Google Scholar] [CrossRef] [Green Version]
- Vangronsveld, J.; Herzig, R.; Weyens, N.; Boulet, J.; Adriaensen, K.; Ruttens, A.; Thewys, T.; Vassilev, A.; Meers, E.; Nehnevajova, E.; et al. Phytoremediation of contaminated soils and groundwater: Lessons from the field. Environ. Sci. Pollut. Res. 2009, 16, 765–794. [Google Scholar] [CrossRef]
- Gubler, F.; Millar, A.A.; Jacobsen, J.V. Dormancy release, ABA and pre-harvest sprouting. Curr. Opin. Plant Biol. 2005, 8, 183–187. [Google Scholar] [CrossRef]
- Nee, G.; Xiang, Y.; Soppe, W.J.J. The release of dormancy, A wake-up call for seeds to germinate. Curr. Opin. Plant Biol. 2017, 35, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, M.V.; Barrero, J.M.; Corbineau, F.; Gubler, F.; Benech-Arnold, R.L. Dormancy in cereals (not too much, not so little): About the mechanisms behind this trait. Seed Sci. Res. 2015, 25, 99–119. [Google Scholar] [CrossRef]
- Finkelstein, R.; Reeves, W.; Ariizumi, T.; Steber, C. Molecular aspects of seed dormancy. Annu. Rev. Plant Biol. 2008, 59, 387–415. [Google Scholar] [CrossRef] [PubMed]
- Graeber, K.; Nakabayashi, K.; Miatton, E.; Leubner-Metzger, G.; Soppe, W.J.J. Molecular mechanisms of seed dormancy. Plant Cell Environ. 2012, 35, 1769–1786. [Google Scholar] [CrossRef] [PubMed]
- Bassel, G.W. To grow or not to grow? Trends Plant Sci. 2016, 21, 498–505. [Google Scholar] [CrossRef] [PubMed]
- Shu, K.; Liu, X.D.; Xie, Q.; He, Z.H. Two faces of one seed: Hormonal regulation of dormancy and germination. Mol. Plant. 2016, 9, 34–45. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.D.; Zhang, H.; Zhao, Y.; Feng, Z.Y.; Li, Q.; Yang, H.Q.; Luan, S.; Li, J.M.; He, Z.H. Auxin controls seed dormancy through stimulation of abscisic acid signaling by inducing ARF-mediated ABI3 activation in Arabidopsis. Proc. Natl. Acad. Sci. USA 2013, 110, 15485–15490. [Google Scholar] [CrossRef] [PubMed]
- Belin, C.; Megies, C.; Hauserova, E.; Lopez-Molina, L. Abscisic acid represses growth of the Arabidopsis embryonic axis after germination by enhancing auxin signaling. Plant Cell 2009, 21, 2253–2268. [Google Scholar] [CrossRef]
- Arc, E.; Sechet, J.; Corbineau, F.; Rajjou, L.; Marion-Poll, A. ABA crosstalk with ethylene and nitric oxide in seed dormancy and germination. Front. Plant Sci. 2013, 4, 19. [Google Scholar] [CrossRef]
- Linkies, A.; Muller, K.; Morris, K.; Tureckova, V.; Wenk, M.; Cadman, C.S.C.; Corbineau, F.; Strnad, M.; Lynn, J.R.; Finch-Savage, W.E.; et al. Ethylene interacts with abscisic acid to regulate endosperm rupture during germination: A comparative approach using Lepidium sativum and Arabidopsis thaliana. Plant Cell 2009, 21, 3803–3822. [Google Scholar] [CrossRef]
- Wojtyla, L.; Lechowska, K.; Kubala, S.; Garnczarska, M. Different modes of hydrogen peroxide action during seed germination. Front. Plant Sci. 2016, 7, 16. [Google Scholar] [CrossRef] [PubMed]
- El-Maarouf-Bouteau, H.; Sajjad, Y.; Bazin, J.; Langlade, N.; Cristescu, S.M.; Balzergue, S.; Baudouin, E.; Bailly, C. Reactive oxygen species, abscisic acid and ethylene interact to regulate sunflower seed germination. Plant Cell Environ. 2015, 38, 364–374. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.T.; Lin, C.; He, F.; Li, Z.; Guan, Y.J.; Hu, Q.J.; Hu, J. Exogenous spermidine improves seed germination of sweet corn via involvement in phytohormone interactions, H2O2 and relevant gene expression. BMC Plant Biol. 2017, 17, 1. [Google Scholar] [CrossRef]
- Bahin, E.; Bailly, C.; Sotta, B.; Kranner, I.; Corbineau, F.; Leymarie, J. Crosstalk between reactive oxygen species and hormonal signalling pathways regulates grain dormancy in barley. Plant Cell Environ. 2011, 34, 980–993. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Gao, Y.; Zhang, Y.C.; Lin, C.; Gong, D.T.; Guan, Y.J.; Hu, J. Reactive oxygen species and gibberellin acid mutual induction to regulate tobacco seed germination. Front. Plant Sci. 2018, 9, 1279. [Google Scholar] [CrossRef] [PubMed]
- Bailly, C.; El-Maarouf-Bouteau, H.; Corbineau, F. From intracellular signaling networks to cell death: The dual role of reactive oxygen species in seed physiology. C. R. Biol. 2008, 331, 806–814. [Google Scholar] [CrossRef] [PubMed]
- Bykova, N.V.; Hoehn, B.; Rampitsch, C.; Banks, T.; Stebbing, J.A.; Fan, T.; Knox, R. Redox-sensitive proteome and antioxidant strategies in wheat seed dormancy control. Proteomics 2011, 11, 865–882. [Google Scholar] [CrossRef] [PubMed]
- El-Maarouf-Bouteau, H.; Meimoun, P.; Job, C.; Job, D.; Bailly, C. Role of protein and mRNA oxidation in seed dormancy and germination. Front. Plant Sci. 2013, 4, 77. [Google Scholar] [CrossRef] [Green Version]
- Job, C.; Rajjou, L.; Lovigny, Y.; Belghazi, M.; Job, D. Patterns of protein oxidation in Arabidopsis seeds and during germination. Plant Physiol. 2005, 138, 790–802. [Google Scholar] [CrossRef]
- Oracz, K.; Bouteau, H.E.M.; Farrant, J.M.; Cooper, K.; Belghazi, M.; Job, C.; Job, D.; Corbineau, F.; Bailly, C. ROS production and protein oxidation as a novel mechanism for seed dormancy alleviation. Plant J. 2007, 50, 452–465. [Google Scholar] [CrossRef] [Green Version]
- Hou, L.Y.; Wang, M.Y.; Wang, H.; Zhang, W.H.; Mao, P.S. Physiological and proteomic analyses for seed dormancy and release in the perennial grass of Leymus chinensis. Environ. Exp. Bot. 2019, 162, 95–102. [Google Scholar] [CrossRef]
- Li, W.Q.; Khan, M.A.; Yamaguchi, S.; Kamiya, Y. Effects of heavy metals on seed germination and early seedling growth of Arabidopsis thaliana. Plant Growth Regul. 2005, 46, 45–50. [Google Scholar] [CrossRef]
- Raviv, B.; Aghajanyan, L.; Granot, G.; Makover, V.; Frenkel, O.; Gutterman, Y.; Grafi, G. The dead seed coat functions as a long-term storage for active hydrolytic enzymes. PLoS ONE 2017, 12, e0181102. [Google Scholar] [CrossRef] [PubMed]
- Kucera, B.; Cohn, M.A.; Leubner-Metzger, G. Plant hormone interactions during seed dormancy release and germination. Seed Sci. Res. 2005, 15, 281–307. [Google Scholar] [CrossRef]
- Bewley, J.D. Seed germination and dormancy. Plant Cell 1997, 9, 1055–1066. [Google Scholar] [CrossRef] [PubMed]
- Bentsink, L.; Koornneef, M. Seed dormancy and germination. Arabidopsis Book 2008, 6. [Google Scholar] [CrossRef] [PubMed]
- Sfaxi-Bousbih, A.; Chaoui, A.; El Ferjani, E. Cadmium impairs mineral and carbohydrate mobilization during the germination of bean seeds. Ecotoxicol. Environ. Safe. 2010, 73, 1123–1129. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.D.; Zhang, G.P.; Yao, H.G.; Zhang, H.M. Genotypic difference of germination and early seedling growth in response to Cd stress and its relation to Cd accumulation. J. Plant Nutr. 2008, 31, 702–715. [Google Scholar] [CrossRef]
- Kuriakose, S.V.; Prasad, M.N.V. Cadmium stress affects seed germination and seedling growth in Sorghum bicolor (L.) Moench by changing the activities of hydrolyzing enzymes. Plant Growth Regul. 2008, 54, 143–156. [Google Scholar] [CrossRef]
- Ahmad, I.; Akhtar, M.J.; Zahir, Z.A.; Jamil, A. Effects of cadmium on seed germination and seedling growth of four wheat (Triticum aestivum L. ) cultivars Pak. J. Bot. 2012, 44, 1569–1574. [Google Scholar]
- Farooqi, Z.R.; Iqbal, M.Z.; Kabir, M.; Shafiq, M. Toxic effects of lead and cadmium on germination and seedling growth of Albizia Lebbeck (L.) benth. Pak. J. Bot. 2009, 41, 27–33. [Google Scholar]
- Liu, S.J.; Yang, C.Y.; Xie, W.J.; Xia, C.H.; Fan, P. The effects of cadmium on germination and seedling growth of Suaeda salsa. In Proceedings of the Seventh International Conference on Waste Management and Technology (ICWMT 7), Beijing, China, 5–7 September 2012; Volume 16, pp. 293–298. [Google Scholar] [CrossRef]
- Alaraidh, I.A.; Alsahli, A.A.; Razik, E.S.A. Alteration of antioxidant gene expression in response to heavy metal stress in Trigonella foenum-graecum L. S. Afr. J. Bot. 2018, 115, 90–93. [Google Scholar] [CrossRef]
- Titov, A.F.; Talanova, V.V.; Boeva, N.P. Growth responses of barley and wheat seedlings to lead and cadmium. Biol. Plant. 1996, 38, 431–436. [Google Scholar] [CrossRef]
- Ahsan, N.; Lee, S.H.; Lee, D.G.; Lee, H.; Lee, S.W.; Bahk, J.D.; Lee, B.H. Physiological and protein profiles alternation of germinating rice seedlings exposed to acute cadmium toxicity. C. R. Biol. 2007, 330, 735–746. [Google Scholar] [CrossRef] [PubMed]
- Munzuroglu, O.; Zengin, F.K. Effect of cadmium on germination, coleoptile and root growth of barley seeds in the presence of gibberellic acid and kinetin. J. Environ. Biol. 2006, 27, 671–677. [Google Scholar] [PubMed]
- Lefevre, I.; Marchal, G.; Correal, E.; Zanuzzi, A.; Lutts, S. Variation in response to heavy metals during vegetative growth in Dorycnium pentaphyllum Scop. Plant Growth Regul. 2009, 59, 1–11. [Google Scholar] [CrossRef]
- Vijayaragavan, M.; Prabhahar, C.; Sureshkumar, J.; Natarajan, A.; Vijayarengan, P.; Sharavanan, S. Toxic effect of cadmium on seed germination, growth and biochemical contents of cowpea (Vigna unguiculata L.) plants. Int. Multidiscip. Res. J. 2011, 1, 1–6. [Google Scholar]
- De Lespinay, A.; Lequeux, H.; Lambillotte, B.; Lutts, S. Protein synthesis is differentially required for germination in Poa pratensis and Trifolium repens in the absence or in the presence of cadmium. Plant Growth Regul. 2010, 61, 205–214. [Google Scholar] [CrossRef]
- Kalai, T.; Bouthour, D.; Manai, J.; Ben Kaab, L.B.; Gouia, H. Salicylic acid alleviates the toxicity of cadmium on seedling growth, amylases and phosphatases activity in germinating barley seeds. Arch. Agron. Soil Sci. 2016, 62, 892–904. [Google Scholar] [CrossRef]
- He, J.Y.; Ren, Y.F.; Zhu, C.; Jiang, D. Effects of cadmium stress on seed germination, seedling growth and seed amylase activities in rice (Oryza sativa). Rice Sci. 2008, 15, 319–325. [Google Scholar] [CrossRef]
- Rivetta, A.; Negrini, N.; Cocucci, M. Involvement of Ca2+-calmodulin in Cd2+ toxicity during the early phases of radish (Raphanus sativus L) seed germination. Plant Cell Environ. 1997, 20, 600–608. [Google Scholar] [CrossRef]
- Cocucci, M.; Negrini, N. Change in the levels of calmodulin and of a calmodulin inhibitor in the early phases of radish (Raphanus sativus L) seed germination—Effects of ABA and fusicoccin. Plant Physiol. 1988, 88, 910–914. [Google Scholar] [CrossRef] [PubMed]
- Rahoui, S.; Chaoui, A.; El Ferjani, E. Reserve mobilization disorder in germinating seeds of Vicia faba L. exposed to cadmium. J. Plant Nutr. 2010, 33, 809–817. [Google Scholar] [CrossRef]
- Rahoui, S.; Chaoui, A.; El Ferjani, E. Membrane damage and solute leakage from germinating pea seed under cadmium stress. J. Hazard. Mater. 2010, 178, 1128–1131. [Google Scholar] [CrossRef] [PubMed]
- Jalmi, S.K.; Bhagat, P.K.; Verma, D.; Noryang, S.; Tayyeba, S.; Singh, K.; Sharma, D.; Sinha, A.K. Traversing the links between heavy metal stress and plant signaling. Front. Plant Sci. 2018, 9, 12. [Google Scholar] [CrossRef] [PubMed]
- Smiri, M.; Jelali, N.; El Ghoul, J. Cadmium affects the NADP-thioredoxin reductase/thioredoxin system in germinating pea seeds. J. Plant Interact. 2013, 8, 125–133. [Google Scholar] [CrossRef] [Green Version]
- Alkhalfioui, F.; Renard, M.; Vensel, W.H.; Wong, J.; Tanaka, C.K.; Hurkman, W.J.; Buchanan, B.B.; Montrichard, F. Thioredoxin-linked proteins are reduced during germination of Medicago truncatula seeds. Plant Physiol. 2007, 144, 1559–1579. [Google Scholar] [CrossRef]
- Smiri, M.; Chaoui, A.; Rouhier, N.; Gelhaye, E.; Jacquot, J.P.; El Ferjani, E. Cadmium affects the glutathione/glutaredoxin system in germinating pea seeds. Biol. Trace Elem. Res. 2011, 142, 93–105. [Google Scholar] [CrossRef]
- Smiri, M.; Jelali, N.; El Ghoul, J. Role for plant peroxiredoxin in cadmium chelation. J. Plant Interact. 2013, 8, 255–262. [Google Scholar] [CrossRef] [Green Version]
- Sneideris, L.C.; Gavassi, M.A.; Campos, M.L.; D’Amico-Damiao, V.; Carvalho, R.F. Effects of hormonal priming on seed germination of pigeon pea under cadmium stress. An. Acad. Bras. Cienc. 2015, 87, 1847–1852. [Google Scholar] [CrossRef] [Green Version]
- He, J.Y.; Ren, Y.F.; Pan, X.B.; Yan, Y.P.; Zhu, C.; Jiang, D. Salicylic acid alleviates the toxicity effect of cadmium on germination, seedling growth, and amylase activity of rice. J. Plant Nutr. Soil Sci. 2010, 173, 300–305. [Google Scholar] [CrossRef]
- Hu, Z.B.; Cools, T.; De Veylder, L. Mechanisms used by plants to cope with DNA Damage. Annu. Rev. Plant Biol. 2016, 67, 439–462. [Google Scholar] [CrossRef] [PubMed]
- Yoshiyama, K.O. SOG1: A master regulator of the DNA damage response in plants. Genes Genet. Syst. 2015, 90, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Bertin, G.; Averbeck, D. Cadmium: Cellular effects, modifications of biomolecules, modulation of DNA repair and genotoxic consequences (A review). Biochimie 2006, 88, 1549–1559. [Google Scholar] [CrossRef] [PubMed]
- Filipic, M. Mechanisms of cadmium induced genomic instability. Mutat. Res.-Fundam. Mol. Mech. Mutagen. 2012, 733, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Afanasieva, K.; Sivolob, A. Physical principles and new applications of comet assay. Biophys. Chem. 2018, 238, 1–7. [Google Scholar] [CrossRef]
- Ventura, L.; Giovannini, A.; Savio, M.; Dona, M.; Macovei, A.; Buttafava, A.; Carbonera, D.; Balestrazzi, A. Single Cell Gel Electrophoresis (Comet) assay with plants: Research on DNA repair and ecogenotoxicity testing. Chemosphere 2013, 92, 1–9. [Google Scholar] [CrossRef]
- Arya, S.K.; Mukherjee, A. Sensitivity of Allium cepa and Vicia faba towards cadmium toxicity. J. Soil Sci. Plant Nutr. 2014, 14, 447–458. [Google Scholar] [CrossRef]
- Koppen, G.; Verschaeve, L. The alkaline comet test on plant cells: A new genotoxicity test for DNA strand breaks in Vicia faba root cells. Mutat. Res.-Environ. Mutagen. Rel. Subj. 1996, 360, 193–200. [Google Scholar] [CrossRef]
- Lin, A.J.; Zhang, X.H.; Chen, M.M.; Cao, Q. Oxidative stress and DNA damages induced by cadmium accumulation. J. Environ. Sci. 2007, 19, 596–602. [Google Scholar] [CrossRef]
- Gichner, T.; Patkova, Z.; Szakova, J.; Demnerova, K. Cadmium induces DNA damage in tobacco roots, but no DNA damage, somatic mutations or homologous recombination in tobacco leaves. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2004, 559, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Tkalec, M.; Stefanic, P.P.; Cvjetko, P.; Sikic, S.; Pavlica, M.; Balen, B. The effects of cadmium-zinc interactions on biochemical responses in tobacco seedlings and adult plants. PLoS ONE 2014, 9, 13. [Google Scholar] [CrossRef] [PubMed]
- Gichner, T.; Patkova, Z.; Szakova, J.; Znidar, I.; Mukherjee, A. DNA damage in potato plants induced by cadmium, ethyl methanesulphonate and gamma-rays. Environ. Exp. Bot. 2008, 62, 113–119. [Google Scholar] [CrossRef]
- Silveira, G.L.; Lima, M.G.F.; dos Reis, G.B.; Palmieri, M.J.; Andrade-Vieria, L.F. Toxic effects of environmental pollutants: Comparative investigation using Allium cepa L. and Lactuca sativa L. Chemosphere 2017, 178, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Seth, C.S.; Misra, V.; Chauhan, L.K.S.; Singh, R.R. Genotoxicity of cadmium on root meristem cells of Allium cepa: Cytogenetic and Comet assay approach. Ecotox. Environ. Safe. 2008, 71, 711–716. [Google Scholar] [CrossRef] [PubMed]
- Balen, B.; Tkalec, M.; Sikic, S.; Tolic, S.; Cvjetko, P.; Pavlica, M.; Vidakovic-Cifrek, Z. Biochemical responses of Lemna minor experimentally exposed to cadmium and zinc. Ecotoxicology 2011, 20, 815–826. [Google Scholar] [CrossRef] [PubMed]
- Cvjetko, P.; Tolic, S.; Sikic, S.; Balen, B.; Tkalec, M.; Vidakovic-Cifrek, Z.; Pavlica, M. Effect of copper on the toxicity and genotoxicity of cadmium in duckweed (Lemna minor L.). Arh. Hig. Rada. Toksikol. 2010, 61, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, C.; Santos, C.; Pinho, S.; Oliveira, H.; Pedrosa, T.; Dias, M.C. Cadmium-induced cyto- and genotoxicity are organ-dependent in lettuce. Chem. Res. Toxicol. 2012, 25, 1423–1434. [Google Scholar] [CrossRef] [PubMed]
- Cao, F.B.; Chen, F.; Sun, H.Y.; Zhang, G.P.; Chen, Z.H.; Wu, F.B. Genome-wide transcriptome and functional analysis of two contrasting genotypes reveals key genes for cadmium tolerance in barley. BMC Genomics 2014, 15, 14. [Google Scholar] [CrossRef]
- Lanier, C.; Bernard, F.; Dumez, S.; Leclercq-Dransart, J.; Lemiere, S.; Vandenbulcke, F.; Nesslany, F.; Platel, A.; Devred, I.; Hayet, A.; et al. Combined toxic effects and DNA damage to two plant species exposed to binary metal mixtures (Cd/Pb). Ecotox. Environ. Safe. 2019, 167, 278–287. [Google Scholar] [CrossRef]
- Fenech, M.; Kirsch-Volders, M.; Natarajan, A.T.; Surralles, J.; Crott, J.W.; Parry, J.; Norppa, H.; Eastmond, D.A.; Tucker, J.D.; Thomas, P. Molecular mechanisms of micronucleus, nucleoplasmic bridge and nuclear bud formation in mammalian and human cells. Mutagenesis 2011, 26, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Vral, A.; Fenech, M.; Thierens, H. The micronucleus assay as a biological dosimeter of in vivo ionising radiation exposure. Mutagenesis 2011, 26, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Talukdar, D. Increasing nuclear ploidy enhances the capability of antioxidant defense and reduces chromotoxicity in Lathyrus sativus roots under cadmium stress. Turk. J. Bot. 2014, 38, 696–712. [Google Scholar] [CrossRef]
- Aslam, R.; Ansari, M.Y.K.; Choudhary, S.; Bhat, T.M.; Jahan, N. Genotoxic effects of heavy metal cadmium on growth, biochemical, cyto-physiological parameters and detection of DNA polymorphism by RAPD in Capsicum annuum L. - An important spice crop of India. Saudi J. Biol. Sci. 2014, 21, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Enan, M.R. Application of random amplified polymorphic DNA (RAPD) to detect the genotoxic effect of heavy metals. Biotechnol. Appl. Biochem. 2006, 43, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Cenkci, S.; Dogan, N. Random amplified polymorphic DNA as a method to screen metal-tolerant barley (Hordeum vulgare L.) genotypes. Turk. J. Bot. 2015, 39, 747–756. [Google Scholar] [CrossRef]
- Tanee, T.; Sudmoon, R.; Thamsenanupap, P.; Chaveerach, A. Effect of cadmium on DNA changes in Ipomoea aquatica Forssk. Pol. J. Environ. Stud. 2016, 25, 311–315. [Google Scholar] [CrossRef]
- Lima, M.G.F.; Rocha, L.C.; Silveira, G.L.; Alvarenga, I.F.S.; Andrade-Vieria, L.F. Nucleolar alterations are reliable parameters to determine the cytogenotoxicity of environmental pollutants. Ecotox. Environ. Safe. 2019, 174, 630–636. [Google Scholar] [CrossRef]
- Cui, W.N.; Wang, H.T.; Song, J.; Cao, X.; Rogers, H.J.; Francis, D.; Jia, C.Y.; Sun, L.Z.; Hou, M.F.; Yang, Y.S.; et al. Cell cycle arrest mediated by Cd-induced DNA damage in Arabidopsis root tips. Ecotox. Environ. Safe. 2017, 145, 569–574. [Google Scholar] [CrossRef]
- Cao, X.; Wang, H.T.; Zhuang, D.F.; Zhu, H.; Du, Y.L.; Cheng, Z.B.; Cui, W.N.; Rogers, H.J.; Zhang, Q.R.; Jia, C.J.; et al. Roles of MSH2 and MSH6 in cadmium-induced G2/M checkpoint arrest in Arabidopsis roots. Chemosphere 2018, 201, 586–594. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.L.; Liu, Z.H.; Chen, R.J.; Li, X.J.; Tai, P.D.; Gong, Z.Q.; Jia, C.Y.; Liu, W. DNA damage and genetic methylation changes caused by Cd in Arabidopsis thaliana seedlings. Environ. Toxicol. Chem. 2015, 34, 2095–2103. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.T.; He, L.; Song, J.; Cui, W.N.; Zhang, Y.Z.; Jia, C.Y.; Francis, D.; Rogers, H.J.; Sun, L.Z.; Tai, P.D.; et al. Cadmium-induced genomic instability in Arabidopsis: Molecular toxicological biomarkers for early diagnosis of cadmium stress. Chemosphere 2016, 150, 258–265. [Google Scholar] [CrossRef] [PubMed]
- Hendrix, S.; Keunen, E.; Mertens, A.I.G.; Beemster, G.T.S.; Vangronsveld, J.; Cuypers, A. Cell cycle regulation in different leaves of Arabidopsis thaliana plants grown under control and cadmium-exposed conditions. Environ. Exp. Bot. 2018, 155, 441–452. [Google Scholar] [CrossRef]
- Sudmoon, R.; Neeratanaphan, L.; Thamsenanupap, P.; Tanee, T. Hyperaccumulation of cadmium and DNA changes in popular vegetable, Brassica chinensis L. Int. J. Environ. Res. 2015, 9, 433–438. [Google Scholar]
- Venkatachalam, P.; Jayaraj, M.; Manikandan, R.; Geetha, N.; Rene, E.R.; Sharma, N.C.; Sahi, S.V. Zinc oxide nanoparticles (ZnONPs) alleviate heavy metal-induced toxicity in Leucaena leucocephala seedlings: A physiochemical analysis. Plant Physiol. Biochem. 2017, 110, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.Q.; Chen, H.N.; Jiang, H.; Lu, W.Y.; Pan, J.J.; Qian, Q.; Xue, D.W. Measuring the damage of heavy metal cadmium in rice seedlings by SRAP analysis combined with physiological and biochemical parameters. J. Sci. Food Agric. 2015, 95, 2292–2298. [Google Scholar] [CrossRef] [PubMed]
- Sorrentino, M.C.; Capozzi, F.; Giordano, S.; Spagnuolo, V. Genotoxic effect of Pb and Cd on in vitro cultures of Sphagnum palustre: An evaluation by ISSR markers. Chemosphere 2017, 181, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Ghiani, A.; Fumagalli, P.; Van, T.N.; Gentili, R.; Citterio, S. The Combined toxic and genotoxic effects of Cd and As to plant bioindicator Trifolium repens L. PLoS ONE 2014, 9, 11. [Google Scholar] [CrossRef] [PubMed]
- Lanier, C.; Bernard, F.; Dumez, S.; Leclercq, J.; Lemiere, S.; Vandenbulcke, F.; Nesslany, F.; Platel, A.; Devred, I.; Cuny, D.; et al. Combined effect of Cd and Pb spiked field soils on bioaccumulation, DNA damage, and peroxidase activities in Trifolium repens. Environ. Sci. Pollut. Res. 2016, 23, 1755–1767. [Google Scholar] [CrossRef] [PubMed]
- Dogan, I.; Ozyigit, I.I.; Tombuloglu, G.; Sakcali, M.S.; Tombuloglu, H. Assessment of Cd-induced genotoxic damage in Urtica pilulifera L. using RAPD-PCR analysis. Biotechnol. Biotechnol. Equip. 2016, 30, 284–291. [Google Scholar] [CrossRef]
- Monteiro, M.S.; Rodriguez, E.; Loureiro, J.; Mann, R.M.; Soares, A.; Santos, C. Flow cytometric assessment of Cd genotoxicity in three plants with different metal accumulation and detoxification capacities. Ecotox. Environ. Safe. 2010, 73, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Komaki, S.; Sugimoto, K. Control of the plant cell cycle by developmental and environmental cues. Plant Cell Physiol. 2012, 53, 953–964. [Google Scholar] [CrossRef]
- Dewitte, W.; Murray, J.A.H. The plant cell cycle. Annu. Rev. Plant Biol. 2003, 54, 235–264. [Google Scholar] [CrossRef] [PubMed]
- De Veylder, L.; Larkin, J.C.; Schnittger, A. Molecular control and function of endoreplication in development and physiology. Trends Plant Sci. 2011, 16, 624–634. [Google Scholar] [CrossRef] [PubMed]
- Barow, M.; Meister, A. Endopolyploidy in seed plants is differently correlated to systematics, organ, life strategy and genome size. Plant Cell Environ. 2003, 26, 571–584. [Google Scholar] [CrossRef]
- Bramsiepe, J.; Wester, K.; Weinl, C.; Roodbarkelari, F.; Kasili, R.; Larkin, J.C.; Hulskamp, M.; Schnittger, A. Endoreplication controls cell fate maintenance. PLoS Genet. 2010, 6, 14. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.O.; Davidson, J.M.; Duronio, R.J. Endoreplication: Polyploidy with purpose. Genes Dev. 2009, 23, 2461–2477. [Google Scholar] [CrossRef] [PubMed]
- Ager, F.J.; Ynsa, M.D.; Dominguez-Solis, J.R.; Gotor, C.; Respaldiza, M.A.; Romero, L.C. Cadmium localization and quantification in the plant Arabidopsis thaliana using micro-PIXE. Nucl. Instrum. Methods Phys. Res. Sect. B-Beam Interact. Mater. Atoms 2002, 189, 494–498. [Google Scholar] [CrossRef]
- Isaure, M.P.; Fayard, B.; Saffet, G.; Pairis, S.; Bourguignon, J. Localization and chemical forms of cadmium in plant samples by combining analytical electron microscopy and X-ray spectromicroscopy. Spectroc. Acta Pt. B-Atom. Spectr. 2006, 61, 1242–1252. [Google Scholar] [CrossRef] [Green Version]
- Scholes, D.R.; Paige, K.N. Plasticity in ploidy: A generalized response to stress. Trends Plant Sci. 2015, 20, 165–175. [Google Scholar] [CrossRef]
- Baryla, A.; Carrier, P.; Franck, F.; Coulomb, C.; Sahut, C.; Havaux, M. Leaf chlorosis in oilseed rape plants (Brassica napus) grown on cadmium-polluted soil: Causes and consequences for photosynthesis and growth. Planta 2001, 212, 696–709. [Google Scholar] [CrossRef] [PubMed]
- Dalla Vecchia, F.; La Rocca, N.; Moro, I.; De Faveri, S.; Andreoli, C.; Rascio, N. Morphogenetic, ultrastructural and physiological damages suffered by submerged leaves of Elodea canadensis exposed to cadmium. Plant Sci. 2005, 168, 329–338. [Google Scholar] [CrossRef]
- Fusconi, A.; Repetto, O.; Bona, E.; Massa, N.; Gallo, C.; Dumas-Gaudot, E.; Berta, G. Effects of cadmium on meristem activity and nucleus ploidy in roots of Pisum sativum L. cv. Frisson seedlings. Environ. Exp. Bot. 2006, 58, 253–260. [Google Scholar] [CrossRef]
- Repetto, O.; Massa, N.; Gianinazzi-Pearson, V.; Dumas-Gaudot, E.; Berta, G. Cadmium effects on populations of root nuclei in two pea genotypes inoculated or not with the arbuscular mycorrhizal fungus Glomus mosseae. Mycorrhiza 2007, 17, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Wojtylakuchta, B.; Gabara, B. Changes in the content of DNA and NYS-stained nuclear, nucleolar and cytoplasmic proteins in cortex cells of pea (Pisum sativum cv. De Grace) roots treated with cadmium. Biochem. Physiol. Pflanz. 1991, 187, 67–76. [Google Scholar] [CrossRef]
- Sobkowiak, R.; Deckert, J. Cadmium-induced changes in growth and cell cycle gene expression in suspension-culture cells of soybean. Plant Physiol. Biochem. 2003, 41, 767–772. [Google Scholar] [CrossRef]
- Sobkowiak, R.; Deckert, J. The effect of cadmium on cell cycle control in suspension culture cells of soybean. Acta Physiol. Plant. 2004, 26, 335–344. [Google Scholar] [CrossRef]
- Zhao, F.Y.; Hu, F.; Han, M.M.; Zhang, S.Y.; Liu, W. Superoxide radical and auxin are implicated in redistribution of root growth and the expression of auxin and cell-cycle genes in cadmium-stressed rice. Russ. J. Plant Physiol. 2011, 58, 851–863. [Google Scholar] [CrossRef]
- Zhao, F.Y.; Wang, K.; Zhang, S.Y.; Ren, J.; Liu, T.; Wang, X. Crosstalk between ABA, auxin, MAPK signaling, and the cell cycle in cadmium-stressed rice seedlings. Acta Physiol. Plant. 2014, 36, 1879–1892. [Google Scholar] [CrossRef]
- Tank, J.G.; Pandya, R.V.; Thaker, V.S. Phytohormones in regulation of the cell division and endoreduplication process in the plant cell cycle. RSC Adv. 2014, 4, 12605–12613. [Google Scholar] [CrossRef]
- Vivancos, P.D.; Dong, Y.P.; Ziegler, K.; Markovic, J.; Pallardo, F.V.; Pellny, T.K.; Verrier, P.J.; Foyer, C.H. Recruitment of glutathione into the nucleus during cell proliferation adjusts whole-cell redox homeostasis in Arabidopsis thaliana and lowers the oxidative defence shield. Plant J. 2010, 64, 825–838. [Google Scholar] [CrossRef] [PubMed]
- Vernoux, T.; Wilson, R.C.; Seeley, K.A.; Reichheld, J.P.; Muroy, S.; Brown, S.; Maughan, S.C.; Cobbett, C.S.; Van Montagu, M.; Inze, D.; et al. The ROOT MERISTEMLESS1/CADMIUM SENSITIVE2 gene defines a glutathione-dependent pathway involved in initiation and maintenance of cell division during postembryonic root development. Plant Cell 2000, 12, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Kosetsu, K.; Matsunaga, S.; Nakagami, H.; Colcombet, J.; Sasabe, M.; Soyano, T.; Takahashi, Y.; Hirt, H.; Machida, Y. The MAP Kinase MPK4 Is required for cytokinesis in Arabidopsis thaliana. Plant Cell 2010, 22, 3778–3790. [Google Scholar] [CrossRef] [PubMed]
- Yao, L.L.; Zhou, Q.; Pei, B.L.; Li, Y.Z. Hydrogen peroxide modulates the dynamic microtubule cytoskeleton during the defence responses to Verticillium dahliae toxins in Arabidopsis. Plant Cell Environ. 2011, 34, 1586–1598. [Google Scholar] [CrossRef] [PubMed]
- Mhamdi, A.; Van Breusegem, F. Reactive oxygen species in plant development. Development 2018, 145. [Google Scholar] [CrossRef] [PubMed]
- Yi, D.; Kamei, C.L.A.; Cools, T.; Vanderauwera, S.; Takahashi, N.; Okushima, Y.; Eekhout, T.; Yoshiyama, K.O.; Larkin, J.; Van den Daele, H.; et al. The Arabidopsis SIAMESE-RELATED cyclin-dependent Kinase inhibitors SMR5 and SMR7 regulate the DNA damage checkpoint in response to reactive oxygen species. Plant Cell 2014, 26, 296–309. [Google Scholar] [CrossRef]
- Zhan, Y.H.; Zhang, C.H.; Zheng, Q.X.; Huang, Z.A.; Yu, C.L. Cadmium stress inhibits the growth of primary roots by interfering auxin homeostasis in Sorghum bicolor seedlings. J. Plant Biol. 2017, 60, 593–603. [Google Scholar] [CrossRef]
- Locato, V.; Paradiso, A.; Sabetta, W.; De Gara, L.; de Pinto, M.C. Chapter nine - Nitric oxide and reactive oxygen species in PCD signaling. Adv. Bot. Res. 2016, 77, 165–192. [Google Scholar]
- Kuthanova, A.; Fischer, L.; Nick, P.; Opatrny, Z. Cell cycle phase-specific death response of tobacco BY-2 cell line to cadmium treatment. Plant Cell Environ. 2008, 31, 1634–1643. [Google Scholar] [CrossRef]
- Pena, L.B.; Barcia, R.A.; Azpilicueta, C.E.; Mendez, A.A.E.; Gallego, S.M. Oxidative post translational modifications of proteins related to cell cycle are involved in cadmium toxicity in wheat seedlings. Plant Sci. 2012, 196, 1–7. [Google Scholar] [CrossRef]
- Garnier, L.; Simon-Plas, F.; Thuleau, P.; Agnel, J.P.; Blein, J.P.; Ranjeva, R.; Montillet, J.L. Cadmium affects tobacco cells by a series of three waves of reactive oxygen species that contribute to cytotoxicity. Plant Cell Environ. 2006, 29, 1956–1969. [Google Scholar] [CrossRef] [PubMed]
- Tamas, L.; Mistrik, I.; Zelinova, V. Heavy metal-induced reactive oxygen species and cell death in barley root tip. Environ. Exp. Bot. 2017, 140, 34–40. [Google Scholar] [CrossRef]
- De Michele, R.; Vurro, E.; Rigo, C.; Costa, A.; Elviri, L.; Di Valentin, M.; Careri, M.; Zottini, M.; Sanita di Toppi, L.; Lo Schiavo, F. Nitric oxide is involved in cadmium-induced programmed cell death in Arabidopsis suspension cultures. Plant Physiol. 2009, 150, 217–228. [Google Scholar] [CrossRef] [PubMed]
- Elviri, L.; Speroni, F.; Careri, M.; Mangia, A.; Sanita di Toppi, L.; Zottini, M. Identification of in vivo nitrosylated phytochelatins in Arabidopsis thaliana cells by liquid chromatography-direct electrospray-linear ion trap-mass spectrometry. J. Chromatogr. A 2010, 1217, 4120–4126. [Google Scholar] [CrossRef] [PubMed]
- Arasimowicz-Jelonek, M.; Floryszak-Wieczorek, J.; Deckert, J.; Rucinska-Sobkowiak, R.; Gzyl, J.; Pawlak-Sprada, S.; Abramowski, D.; Jelonek, T.; Gwozdz, E.A. Nitric oxide implication in cadmium-induced programmed cell death in roots and signaling response of yellow lupine plants. Plant Physiol. Biochem. 2012, 58, 124–134. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.W.; Xu, W.Z.; Xu, H.; Chen, Y.S.; He, Z.Y.; Ma, M. Nitric oxide modulates cadmium influx during cadmium-induced programmed cell death in tobacco BY-2 cells. Planta 2010, 232, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Li, Z.; Xing, D. Nitric oxide promotes MPK6-mediated caspase-3-like activation in cadmium-induced Arabidopsis thaliana programmed cell death. Plant Cell Environ. 2013, 36, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Iakimova, E.T.; Woltering, E.J.; Kapchina-Toteva, V.M.; Harren, F.J.M.; Cristescu, S.M. Cadmium toxicity in cultured tomato cells - Role of ethylene, proteases and oxidative stress in cell death signaling. Cell Biol. Int. 2008, 32, 1521–1529. [Google Scholar] [CrossRef]
- Pormehr, M.; Ghanati, F.; Sharifi, M.; McCabe, P.F.; Hosseinkhani, S.; Zare-Maivan, H. The role of SIPK signaling pathway in antioxidant activity and programmed cell death of tobacco cells after exposure to cadmium. Plant Sci. 2019, 280, 416–423. [Google Scholar] [CrossRef]
- Zhang, W.N.; Chen, W.L. Role of salicylic acid in alleviating photochemical damage and autophagic cell death induction of cadmium stress in Arabidopsis thaliana. Photochem. Photobiol. Sci. 2011, 10, 947–955. [Google Scholar] [CrossRef]
- Gzyl, J.; Chmielowska-Bak, J.; Przymusinski, R. Gamma-tubulin distribution and ultrastructural changes in root cells of soybean (Glycine max L.) seedlings under cadmium stress. Environ. Exp. Bot. 2017, 143, 82–90. [Google Scholar] [CrossRef]
- Yue, J.Y.; Wei, X.J.; Wang, H.Z. Cadmium tolerant and sensitive wheat lines: Their differences in pollutant accumulation, cell damage, and autophagy. Biol. Plant. 2018, 62, 379–387. [Google Scholar] [CrossRef]
- De Araujo, R.P.; de Almeida, A.A.F.; Pereira, L.S.; Mangabeira, P.A.O.; Souza, J.O.; Pirovani, C.P.; Ahnert, D.; Baligar, V.C. Photosynthetic, antioxidative, molecular and ultrastructural responses of young cacao plants to Cd toxicity in the soil. Ecotox. Environ. Safe. 2017, 144, 148–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gou, W.; Li, X.; Guo, S.; Liu, Y.; Li, F.; Xie, Q. Autophagy in plant: A new orchestrator in the regulation of the phytohormones homeostasis. Int. J. Mol. Sci. 2019, 20, 2900. [Google Scholar] [CrossRef] [PubMed]
- Signorelli, S.; Tarkowski, L.P.; Van den Ende, W.; Bassham, D.C. Linking autophagy to abiotic and biotic stress responses. Trends Plant Sci. 2019, 24, 413–430. [Google Scholar] [CrossRef] [PubMed]
- Cosgrove, D.J. Growth of the plant cell wall. Nat. Rev. Mol. Cell Biol. 2005, 6, 850–861. [Google Scholar] [CrossRef] [PubMed]
- Mohnen, D. Pectin structure and biosynthesis. Curr. Opin. Plant Biol. 2008, 11, 266–277. [Google Scholar] [CrossRef] [PubMed]
- Scheller, H.V.; Ulvskov, P. Hemicelluloses. Annu. Rev. Plant Biol. 2010, 61, 263–289. [Google Scholar] [CrossRef]
- Krzeslowska, M. The cell wall in plant cell response to trace metals: Polysaccharide remodeling and its role in defense strategy. Acta Physiol. Plant. 2011, 33, 35–51. [Google Scholar] [CrossRef]
- Grant, G.T.; Morris, E.R.; Rees, D.A.; Smith, P.J.C.; Thom, D. Biological interactions between polysaccharides and divalent cations - Egg-box model. FEBS Lett. 1973, 32, 195–198. [Google Scholar] [CrossRef]
- Hall, J.L. Cellular mechanisms for heavy metal detoxification and tolerance. J. Exp. Bot. 2002, 53, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Tenhaken, R. Cell wall remodeling under abiotic stress. Front. Plant Sci. 2015, 5, 9. [Google Scholar] [CrossRef] [PubMed]
- Verslues, P.E.; Agarwal, M.; Katiyar-Agarwal, S.; Zhu, J.H.; Zhu, J.K. Methods and concepts in quantifying resistance to drought, salt and freezing, abiotic stresses that affect plant water status. Plant J. 2006, 45, 523–539. [Google Scholar] [CrossRef] [PubMed]
- Van Belleghem, F.; Cuypers, A.; Semane, B.; Smeets, K.; Vangronsveld, J.; d’Haen, J.; Valcke, R. Subcellular localization of cadmium in roots and leaves of Arabidopsis thaliana. New Phytol. 2007, 173, 495–508. [Google Scholar] [CrossRef] [PubMed]
- Vazquez, S.; Goldsbrough, P.; Carpena, R.O. Assessing the relative contributions of phytochelatins and the cell wall to cadmium resistance in white lupin. Physiol. Plant. 2006, 128, 487–495. [Google Scholar] [CrossRef]
- Huang, W.L.; Bai, Z.Q.; Jiao, J.; Yuan, H.L.; Bao, Z.A.; Chen, S.N.; Ding, M.H.; Liang, Z.S. Distribution and chemical forms of cadmium in Coptis chinensis Franch. determined by laser ablation ICP-MS, cell fractionation, and sequential extraction. Ecotox. Environ. Safe. 2019, 171, 894–903. [Google Scholar] [CrossRef] [PubMed]
- Kramer, U. Metal Hyperaccumulation in Plants. Annu. Rev. Plant Biol. 2010, 61, 517–534. [Google Scholar] [CrossRef]
- Zhao, H.; Jin, Q.J.; Wang, Y.J.; Chu, L.L.; Li, X.; Xu, Y.C. Effects of nitric oxide on alleviating cadmium stress in Typha angustifolia. Plant Growth Regul. 2016, 78, 243–251. [Google Scholar] [CrossRef]
- Dong, Y.J.; Chen, W.F.; Xu, L.L.; Kong, J.; Liu, S.; He, Z.L. Nitric oxide can induce tolerance to oxidative stress of peanut seedlings under cadmium toxicity. Plant Growth Regul. 2016, 79, 19–28. [Google Scholar] [CrossRef]
- Berni, R.; Luyckx, M.; Xu, X.; Legay, S.; Sergeant, K.; Hausman, J.F.; Lutts, S.; Cai, G.; Guerriero, G. Reactive oxygen species and heavy metal stress in plants: Impact on the cell wall and secondary metabolism. Environ. Exp. Bot. 2019, 161, 98–106. [Google Scholar] [CrossRef]
- Loix, C.; Huybrechts, M.; Vangronsveld, J.; Gielen, M.; Keunen, E.; Cuypers, A. Reciprocal interactions between cadmium-induced cell wall responses and oxidative stress in plants. Front. Plant Sci. 2017, 8, 19. [Google Scholar] [CrossRef] [PubMed]
- Parrotta, L.; Guerriero, G.; Sergeant, K.; Cal, G.; Hausman, J.F. Target or barrier? The cell wall of early- and later-diverging plants vs cadmium toxicity: Differences in the response mechanisms. Front. Plant Sci. 2015, 6, 16. [Google Scholar] [CrossRef] [PubMed]
- Douchiche, O.; Rihouey, C.; Schaumann, A.; Driouich, A.; Morvan, C. Cadmium-induced alterations of the structural features of pectins in flax hypocotyl. Planta 2007, 225, 1301–1312. [Google Scholar] [CrossRef] [PubMed]
- Paynel, F.; Schaumann, A.; Arkoun, M.; Douchiche, O.; Morvan, C. Temporal regulation of cell-wall pectin methylesterase and peroxidase isoforms in cadmium-treated flax hypocotyl. Ann. Bot. 2009, 104, 1363–1372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, J.; An, L.Y.; Lu, H.; Zhu, C. Exogenous nitric oxide enhances cadmium tolerance of rice by increasing pectin and hemicellulose contents in root cell wall. Planta 2009, 230, 755–765. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.Y.; He, C.X.; Wu, Z.C.; Liu, X.W.; Cai, M.M.; Jia, W.; Zhao, X.H. Selenium reduces cadmium accumulation in seed by increasing cadmium retention in root of oilseed rape (Brassica napus L.). Environ. Exp. Bot. 2019, 158, 161–170. [Google Scholar] [CrossRef]
- Li, H.B.; Zheng, X.W.; Tao, L.X.; Yang, Y.J.; Gao, L.; Xiong, J. Aeration increases cadmium (Cd) retention by enhancing iron plaque formation and regulating pectin synthesis in the roots of rice (Oryza sativa) seedlings. Rice 2019, 12, 14. [Google Scholar] [CrossRef]
- Finger-Teixeira, A.; Ferrarese, M.D.L.; Soares, A.R.; da Silva, D.; Ferrarese, O. Cadmium-induced lignification restricts soybean root growth. Ecotox. Environ. Safe. 2010, 73, 1959–1964. [Google Scholar] [CrossRef]
- Schutzendubel, A.; Schwanz, P.; Teichmann, T.; Gross, K.; Langenfeld-Heyser, R.; Godbold, D.L.; Polle, A. Cadmium-induced changes in antioxidative systems, hydrogen peroxide content, and differentiation in Scots pine roots. Plant Physiol. 2001, 127, 887–898. [Google Scholar] [CrossRef]
- Liu, Q.Q.; Luo, L.; Zheng, L.Q. Lignins: Biosynthesis and biological functions in plants. Int. J. Mol. Sci. 2018, 19, 335. [Google Scholar] [CrossRef]
- Rui, H.Y.; Chen, C.; Zhang, X.X.; Shen, Z.G.; Zhang, F.Q. Cd-induced oxidative stress and lignification in the roots of two Vicia sativa L. varieties with different Cd tolerances. J. Hazard. Mater. 2016, 301, 304–313. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, R.; Kunkowska, A.B.; Schippers, J.H.M. Role of reactive oxygen species during cell expansion in leaves. Plant Physiol. 2016, 172, 2098–2106. [Google Scholar] [CrossRef] [PubMed]
- Muller, K.; Linkies, A.; Vreeburg, R.A.M.; Fry, S.C.; Krieger-Liszkay, A.; Leubner-Metzger, G. In vivo cell wall loosening by hydroxyl radicals during cress seed germination and elongation growth. Plant Physiol. 2009, 150, 1855–1865. [Google Scholar] [CrossRef] [PubMed]
- Schopfer, P. Hydrogen peroxide-mediated cell-wall stiffening in vitro in maize coleoptiles. Planta 1996, 199, 43–49. [Google Scholar] [CrossRef]
- Vatehova, Z.; Malovikova, A.; Kollarova, K.; Kucerova, D.; Liskova, D. Impact of cadmium stress on two maize hybrids. Plant Physiol. Biochem. 2016, 108, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Yang, B.; Wan, H.B.; Fang, X.L.; Yang, C.Y. The differences of cell wall in roots between two contrasting soybean cultivars exposed to cadmium at young seedlings. Environ. Sci. Pollut. Res. 2018, 25, 29705–29714. [Google Scholar] [CrossRef]
- Somerville, C. Cellulose synthesis in higher plants. Annu. Rev. Cell Dev. Biol. 2006, 22, 53–78. [Google Scholar] [CrossRef] [PubMed]
- Haigler, C.H.; Ivanova-Datcheva, M.; Hogan, P.S.; Salnikov, V.V.; Hwang, S.; Martin, K.; Delmer, D.P. Carbon partitioning to cellulose synthesis. Plant Mol. Biol. 2001, 47, 29–51. [Google Scholar] [CrossRef] [PubMed]
- Gutsch, A.; Keunen, E.; Guerriero, G.; Renaut, J.; Cuypers, A.; Hausman, J.F.; Sergeant, K. Long-term cadmium exposure influences the abundance of proteins that impact the cell wall structure in Medicago sativa stems. Plant Biol. 2018, 20, 1023–1035. [Google Scholar] [CrossRef]
- Guo, H.P.; Hong, C.T.; Xiao, M.Z.; Chen, X.M.; Chen, H.M.; Zheng, B.S.; Jiang, D.A. Real-time kinetics of cadmium transport and transcriptomic analysis in low cadmium accumulator Miscanthus sacchariflorus. Planta 2016, 244, 1289–1302. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, D.C.; Zhang, H.Y.; Gao, H.B.; Guo, X.L.; Wang, D.M.; Zhang, X.Q.; Zhang, A.M. The alpha- and beta-expansin and xyloglucan endotransglucosylase/hydrolase gene families of wheat: Molecular cloning, gene expression, and EST data mining. Genomics 2007, 90, 516–529. [Google Scholar] [CrossRef] [PubMed]
- Cosgrove, D.J. Plant expansins: Diversity and interactions with plant cell walls. Curr. Opin. Plant Biol. 2015, 25, 162–172. [Google Scholar] [CrossRef] [PubMed]
- Kuluev, B.; Avalbaev, A.; Mikhaylova, E.; Nikonorov, Y.; Berezhneva, Z.; Chemeris, A. Expression profiles and hormonal regulation of tobacco expansin genes and their involvement in abiotic stress response. J. Plant Physiol. 2016, 206, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Zhang, Y.X.; Chai, T.Y. Cloning, characterization, and expression of the BjEXPA1 gene and its promoter region from Brassica juncea L. Plant Growth Regul. 2011, 64, 39–51. [Google Scholar] [CrossRef]
- Ren, Y.Q.; Chen, Y.H.; An, J.; Zhao, Z.X.; Zhang, G.Q.; Wang, Y.; Wang, W. Wheat expansin gene TaEXPA2 is involved in conferring plant tolerance to Cd toxicity. Plant Sci. 2018, 270, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Van Sandt, V.S.T.; Suslov, D.; Verbelen, J.P.; Vissenberg, K. Xyloglucan endotransglucosylase activity loosens a plant cell wall. Ann. Bot. 2007, 100, 1467–1473. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.P.; Li, C.L.; Zhang, F.; Yu, Q.Q.; Gao, S.; Zhang, M.L.; Tian, H.Y.; Zhang, J.; Yuan, X.Z.; Ding, Z.J. Ethylene promotes cadmium-induced root growth inhibition through EIN3 controlled XTH33 and LSU1 expression in Arabidopsis. Plant Cell Environ. 2018, 41, 2449–2462. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.S.; Sa, G.; Sun, J.; Shen, Z.D.; Zhao, R.; Ding, M.Q.; Deng, S.R.; Lu, Y.J.; Zhang, Y.H.; Shen, X.; et al. Overexpression of Populus euphratica xyloglucan endotransglucosylase/hydrolase gene confers enhanced cadmium tolerance by the restriction of root cadmium uptake in transgenic tobacco. Environ. Exp. Bot. 2014, 100, 74–83. [Google Scholar] [CrossRef]
- Yu, R.G.; Jiang, Q.; Xv, C.; Li, L.; Bu, S.J.; Shi, G.R. Comparative proteomics analysis of peanut roots reveals differential mechanisms of cadmium detoxification and translocation between two cultivars differing in cadmium accumulation. BMC Plant Biol. 2019, 19, 15. [Google Scholar] [CrossRef]
- Majda, M.; Robert, S. The role of auxin in cell wall expansion. Int. J. Mol. Sci. 2018, 19, 951. [Google Scholar] [CrossRef]
- Rascio, N.; Navari-Izzo, F. Heavy metal hyperaccumulating plants: How and why do they do it? And what makes them so interesting? Plant Sci. 2011, 180, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Benzarti, S.; Mohri, S.; Ono, Y. Plant response to heavy metal toxicity: Comparative study between the hyperaccumulator Thlaspi caerulescens (ecotype Ganges) and nonaccumulator plants: Lettuce, radish, and alfalfa. Environ. Toxicol. 2008, 23, 607–617. [Google Scholar] [CrossRef] [PubMed]
- Meyer, C.L.; Juraniec, M.; Huguet, S.; Chaves-Rodriguez, E.; Salis, P.; Isaure, M.P.; Goormaghtigh, E.; Verbruggen, N. Intraspecific variability of cadmium tolerance and accumulation, and cadmium-induced cell wall modifications in the metal hyperaccumulator Arabidopsis halleri. J. Exp. Bot. 2015, 66, 3215–3227. [Google Scholar] [CrossRef] [PubMed]
- Verbruggen, N.; Hermans, C.; Schat, H. Molecular mechanisms of metal hyperaccumulation in plants. New Phytol. 2009, 181, 759–776. [Google Scholar] [CrossRef] [PubMed]
- Li, T.Q.; Tao, Q.; Shohag, M.J.I.; Yang, X.E.; Sparks, D.L.; Liang, Y.C. Root cell wall polysaccharides are involved in cadmium hyperaccumulation in Sedum alfredii. Plant Soil 2015, 389, 387–399. [Google Scholar] [CrossRef]
- Baliardini, C.; Corso, M.; Verbruggen, N. Transcriptomic analysis supports the role of CATION EXCHANGER 1 in cellular homeostasis and oxidative stress limitation during cadmium stress. Plant Signal. Behav. 2016, 11, 5. [Google Scholar] [CrossRef]
- Peng, J.S.; Wang, Y.J.; Ding, G.; Ma, H.L.; Zhang, Y.J.; Gong, J.M. A pivotal role of cell wall in cadmium accumulation in the Crassulaceae hyperaccumulator Sedum plumbizincicola. Mol. Plant. 2017, 10, 771–774. [Google Scholar] [CrossRef]
- Anjum, N.A.; Umar, S.; Ahmad, A.; Iqbal, M.; Khan, N.A. Ontogenic variation in response of Brassica campestris L. to cadmium toxicity. J. Plant Interact. 2008, 3, 189–198. [Google Scholar] [CrossRef]
- Hancock, L.M.S.; Ernst, C.L.; Charneskie, R.; Ruane, L.G. Effects of cadmium and mycorrhizal fungi on growth, fitness and cadmium accumulation in flax (Linum unitatissimum; Linaceae). Am. J. Bot. 2012, 99, 1445–1452. [Google Scholar] [CrossRef]
- Keunen, E.; Truyens, S.; Bruckers, L.; Remans, T.; Vangronsveld, J.; Cuypers, A. Survival of Cd-exposed Arabidopsis thaliana: Are these plants reproductively challenged? Plant Physiol. Biochem. 2011, 49, 1084–1091. [Google Scholar] [CrossRef]
- Maistri, S.; DalCorso, G.; Vicentini, V.; Furini, A. Cadmium affects the expression of ELF4, a circadian clock gene in Arabidopsis. Environ. Exp. Bot. 2011, 72, 115–122. [Google Scholar] [CrossRef]
- Gusmão Lima, A.I.; Pereira, S.I.A.; de Almeida Figueira, E.M.; Caldeira, G.C.N.; Caldeira, H.D.Q. Cadmium uptake in pea plants under environmentally-relevant exposures: The risk of food-chain transfer. J. Plant Nutr. 2006, 29, 2165–2177. [Google Scholar] [CrossRef]
- Rodda, M.S.; Li, G.; Reid, R.J. The timing of grain Cd accumulation in rice plants: The relative importance of remobilisation within the plant and root Cd uptake post-flowering. Plant Soil 2011, 347, 105–114. [Google Scholar] [CrossRef]
- Hediji, H.; Djebali, W.; Belkadhi, A.; Cabasson, C.; Moing, A.; Rolin, D.; Brouquisse, R.; Gallusci, P.; Chaibi, W. Impact of long-term cadmium exposure on mineral content of Solanum lycopersicum plants: Consequences on fruit production. S. Afr. J. Bot. 2015, 97, 176–181. [Google Scholar] [CrossRef]
- Shekari, L.; Aroiee, H.; Mirshekari, A.; Nemati, H. Protective role of selenium on cucumber (Cucumis sativus L.) exposed to cadmium and lead stress during reproductive stage role of selenium on heavy metals stress. J. Plant Nutr. 2019, 42, 529–542. [Google Scholar] [CrossRef]
- Soldatova, N.A.; Khryanin, V.N. The effects of heavy metal salts on the phytohormonal status and sex expression in marijuana. Russ. J. Plant Physiol. 2010, 57, 96–100. [Google Scholar] [CrossRef]
- Pan, J.T.; Li, D.Q.; Shu, Z.F.; Jiang, X.; Ma, W.W.; Wang, W.D.; Zhu, J.J.; Wang, Y.H. CsPDC-E1 alpha, a novel pyruvate dehydrogenase complex E1 alpha subunit gene from Camellia sinensis, is induced during cadmium inhibiting pollen tube growth. Can. J. Plant Sci. 2018, 98, 62–70. [Google Scholar] [CrossRef]
- Sabrine, H.; Afif, H.; Mohamed, B.; Hamadi, B.; Maria, H. Effects of cadmium and copper on pollen germination and fruit set in pea (Pisum sativum L.). Sci. Hortic. 2010, 125, 551–555. [Google Scholar] [CrossRef]
- Xiong, Z.T.; Peng, Y.H. Response of pollen germination and tube growth to cadmium with special reference to low concentration exposure. Ecotox. Environ. Safe. 2001, 48, 51–55. [Google Scholar] [CrossRef]
- Sawidis, T. Effect of cadmium on pollen germination and tube growth in Lilium longiflorum and Nicotiana tabacum. Protoplasma 2008, 233, 95–106. [Google Scholar] [CrossRef]
- Sharafi, Y.; Talebi, S.F.; Talei, D. Effects of heavy metals on male gametes of sweet cherry. Caryologia 2017, 70, 166–173. [Google Scholar] [CrossRef]
- Wang, X.X.; Zhang, S.S.; Gao, Y.; Lu, W.G.; Sheng, X.Y. Different heavy metals have various effects on Picea wilsonii pollen germination and tube growth. Plant Signal. Behav. 2015, 10, 4. [Google Scholar] [CrossRef] [PubMed]
- Wudick, M.M.; Feijo, J.A. At the Intersection: Merging Ca2+ and ROS signaling pathways in pollen. Mol. Plant 2014, 7, 1595–1597. [Google Scholar] [CrossRef] [PubMed]
- Smirnova, A.V.; Matveyeva, N.P.; Yermakov, I.P. Reactive oxygen species are involved in regulation of pollen wall cytomechanics. Plant Biol. 2014, 16, 252–257. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, M.E.A.; Piotto, F.A.; Nogueira, M.L.; Gomes, F.G.; Chamma, H.; Pizzaia, D.; Azevedo, R.A. Cadmium exposure triggers genotype-dependent changes in seed vigor and germination of tomato offspring. Protoplasma 2018, 255, 989–999. [Google Scholar] [CrossRef] [PubMed]
- Truyens, S.; Beckers, B.; Thijs, S.; Weyens, N.; Cuypers, A.; Vangronsveld, J. Cadmium-induced and trans-generational changes in the cultivable and total seed endophytic community of Arabidopsis thaliana. Plant Biol. 2016, 18, 376–381. [Google Scholar] [CrossRef] [PubMed]
- Truyens, S.; Weyens, N.; Cuypers, A.; Vangronsveld, J. Changes in the population of seed bacteria of transgenerationally Cd-exposed Arabidopsis thaliana. Plant Biol. 2013, 15, 971–981. [Google Scholar] [CrossRef]
- Remans, T.; Thijs, S.; Truyens, S.; Weyens, N.; Schellingen, K.; Keunen, E.; Gielen, H.; Cuypers, A.; Vangronsveld, J. Understanding the development of roots exposed to contaminants and the potential of plant-associated bacteria for optimization of growth. Ann. Bot. 2012, 110, 239–252. [Google Scholar] [CrossRef]
- Bhat, J.A.; Shivaraj, S.M.; Singh, P.; Navadagi, D.B.; Tripathi, D.K.; Dash, P.K.; Solanke, A.U.; Sonah, H.; Deshmukh, R. Role of silicon in mitigation of heavy metal stresses in crop plants. Plants 2019, 8, 71. [Google Scholar] [CrossRef]
- Gutsch, A.; Vandionant, S.; Sergeant, K.; Jozefczak, M.; Vangronsveld, J.; Hausman, J.-F.; Cuypers, A. Systems Biology of Metal Tolerance in Plants: A Case Study on the Effects of Cd Exposure on Two Model Plants. In Plant Metallomics and Functional Omics: A System-Wide Perspective; Sablok, G., Ed.; Springer International Publishing: Cham, Switzerland, 2019; pp. 23–37. [Google Scholar]



{kind=link}
{kind=link}
| Species | Organ | Cd Concentration | Exposure Duration | Effect | Detection Method | Reference |
|---|---|---|---|---|---|---|
| Allium cepa | Root tip | 50–200 µM | 2 h + 24 h recovery | Micronucleus formation | Microscopic analysis | Arya and Mukherjee, 2014 [80] |
| Chromosomal aberrations | ||||||
| % tail DNA ↑ | Comet assay (alkaline) | |||||
| Root tip | 25 µM | 48 h | Chromosomal aberrations | Microscopic analysis | Silveira et al., 2017 [86] | |
| Micronucleus formation | ||||||
| % DNA damage ↑ | Comet assay (alkaline) | |||||
| Root tip | 25 µM | 48 h | Number of nucleoli ↑ | Microscopic analysis | Lima et al., 2019 [100] | |
| Arabidopsis thaliana | Root tip | 0.125–2.5 mg L−1 | 5 d | Altered expression DNA repair genes | qRT-PCR | Cui et al., 2017 [101] |
| Root | 1.25–4 mg L−1 | 5 d | Altered RAPD profile | RAPD | Cao et al., 2018 [102] | |
| Altered expression DNA repair genes | qRT-PCR | |||||
| Leaf | 0.5–5 mg L−1 | 16 d | Altered AFLP profile | AFLP | Li et al., 2015 [103] | |
| Leaf | 0.25–8 mg L−1 | 15 d | Microsatellite instability | SSR | Wang et al., 2016 [104] | |
| Altered RAPD profile | RAPD | |||||
| Leaf | 5 µM | 72 h | Altered expression DNA repair genes | qRT-PCR | Hendrix et al., 2018 [105] | |
| Brassica chinensis | Leaf | 15–120 mg kg−1 soil | 30 d | Altered RAPD profile | RAPD | Sudmoon et al., 2015 [106] |
| Brassica oleracea | Root | 2.5–20 mg kg−1 soil | 3–56 d | Altered % tail intensity | Comet assay (alkaline) | Lanier et al., 2019 [92] |
| Capsicum annuum | Root tip | 20–100 ppm | 24 h | Chromosomal aberrations | Microscopic analysis | Aslam et al., 2014 [96] |
| Leaf | 20–100 ppm | 24 h | Altered RAPD profile | RAPD | ||
| Hordeum vulgare | Root tip | 75–225 µM | 7 d | Altered RAPD profile (GTS ↓) | RAPD | Cenkci and Dogan, 2015 [98] |
| Leaf | 5 µM | 15 d | DNA damage ↑ | Comet assay (alkaline) | Cao et al., 2014 [91] | |
| Ipomoea aquatica | Entire seedling | 15–120 mg kg−1 soil | 21 d | Altered RAPD profile (GTS ↓) | RAPD | Tanee et al., 2016 [99] |
| Lactuca sativa | Root tip | 25 µM | 48 h | Chromosomal aberrations | Microscopic analysis | Silveira et al., 2017 [86] |
| Micronucleus formation | ||||||
| % DNA damage ↑ | Comet assay (alkaline) | |||||
| Lathyrus sativus | Root tip | 5–50 µM | 3–7 d | Chromosomal aberrations | Microscopic analysis | Talukdar, 2014 [95] |
| Micronucleus formation | ||||||
| Leucaena leucocephala | Leaf | 50 mg L−1 | 15 d | Altered RAPD profile | RAPD | Venkatachalam et al., 2017 [107] |
| Nicotiana tabacum | Root and leaf | 10–15 µM | 7 d | % tail DNA ↑ | Comet assay (alkaline) | Tkalec et al., 2014 [84] |
| Oryza sativa | Root tip | 50–200 µM | 48–96 h | Altered SRAP profil (GTS ↓) | SRAP | Zhang et al., 2015 [108] |
| Sphagnum palustre | Shoot | 0.1–10 µM | 24–48 h | Altered ISSR profile (GTS ↓) | ISSR | Sorrentino et al., 2017 [109] |
| Trifolium repens | Root | 2.5–20 mg kg−1 soil | 3–56 d | Altered % tail intensity | Comet assay (alkaline) | Lanier et al., 2019 [92] |
| Root and leaf | 20–60 mg kg−1 soil | 2 weeks | Altered RAPD profile | RAPD | Ghiani et al., 2014 [110] | |
| Leaf | 2.5–20 mg kg−1 soil | 3–56 d | Tail moment ↑ | Comet assay (alkaline) | Lanier et al., 2016 [111] | |
| Urtica pilulifera | Root tip | 100–200 µM | 2 months | Altered RAPD profile | RAPD | Dogan et al., 2016 [112] |
| Vicia faba | Root tip | 50–200 µM | 2 h + 24 h recovery | Micronucleus formation | Microscopic analysis | Arya and Mukherjee, 2014 [80] |
| Chromosomal aberrations | ||||||
| % tail DNA ↑ | Comet assay (alkaline) |
| Species | Organ | Cd Concentration | Exposure Duration | Effect | Detection Method | Reference |
|---|---|---|---|---|---|---|
| Allium cepa | Root tip | 50–200 µM | 2 h + 24 h recovery | Mitotic index ↓ | Microscopic analysis | Arya and Mukherjee, 2014 [80] |
| Root tip | 25 µM | 48 h | Mitotic index ↓ | Microscopic analysis | Silveira et al., 2017 [86] | |
| Arabidopsis thaliana | Root tip | 0.125–2.5 mg L−1 | 5 d | 2C ↓, 4C ↑, 8C ↑ | FCM | Cui et al., 2017 [101] |
| Altered cell cycle phase distribution | ||||||
| Altered expression cell cycle-related genes | qRT-PCR | |||||
| Root | 1.25–4 mg L−1 | 5 d | 2C ↓, 4C ↑ | FCM | Cao et al., 2018 [102] | |
| Altered expression of cell cycle-related genes | qRT-PCR | |||||
| Leaf | 5 µM | 3–12 d | Endoreduplication factor ↓ | FCM | Hendrix et al., 2018 [105] | |
| Epidermal cell number and cell surface area ↓ | Microscopic analysis | |||||
| Altered expression of cell-cycle related genes | qRT-PCR | |||||
| Capsicum annuum | Root tip | 20–100 ppm | 24 h | Mitotic index ↓ | Microscopic analysis | Aslam et al., 2014 [96] |
| Lactuca sativa | Root tip | 25 µM | 48 h | Mitotic index ↓ | Microscopic analysis | Silveira et al., 2017 [86] |
| Lathyrus sativus | Root tip | 5–50 µM | 3–7 d | Mitotic index ↓ | Microscopic analysis | Talukdar, 2014 [95] |
| Oryza sativa | Root | 200 µM | 7 d | Cortex cell length in elongation zone ↓ | Microscopic analysis | Zhao et al., 2014 [131] |
| Cortex cell number in elongation zone ↓ | ||||||
| 7–11 d | Altered expression of cell cycle-related genes | RT-PCR | ||||
| Sorghum bicolor | Root tip | 50–200 µM | 5 d | Inhibition of S phase progression | EdU assay | Zhan et al., 2017 [139] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huybrechts, M.; Cuypers, A.; Deckers, J.; Iven, V.; Vandionant, S.; Jozefczak, M.; Hendrix, S. Cadmium and Plant Development: An Agony from Seed to Seed. Int. J. Mol. Sci. 2019, 20, 3971. https://doi.org/10.3390/ijms20163971
Huybrechts M, Cuypers A, Deckers J, Iven V, Vandionant S, Jozefczak M, Hendrix S. Cadmium and Plant Development: An Agony from Seed to Seed. International Journal of Molecular Sciences. 2019; 20(16):3971. https://doi.org/10.3390/ijms20163971
Chicago/Turabian StyleHuybrechts, Michiel, Ann Cuypers, Jana Deckers, Verena Iven, Stéphanie Vandionant, Marijke Jozefczak, and Sophie Hendrix. 2019. "Cadmium and Plant Development: An Agony from Seed to Seed" International Journal of Molecular Sciences 20, no. 16: 3971. https://doi.org/10.3390/ijms20163971
APA StyleHuybrechts, M., Cuypers, A., Deckers, J., Iven, V., Vandionant, S., Jozefczak, M., & Hendrix, S. (2019). Cadmium and Plant Development: An Agony from Seed to Seed. International Journal of Molecular Sciences, 20(16), 3971. https://doi.org/10.3390/ijms20163971