Selenium Modulates the Level of Auxin to Alleviate the Toxicity of Cadmium in Tobacco

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effects of Se on Tobacco Phenotype and Biomass under Cd Stress
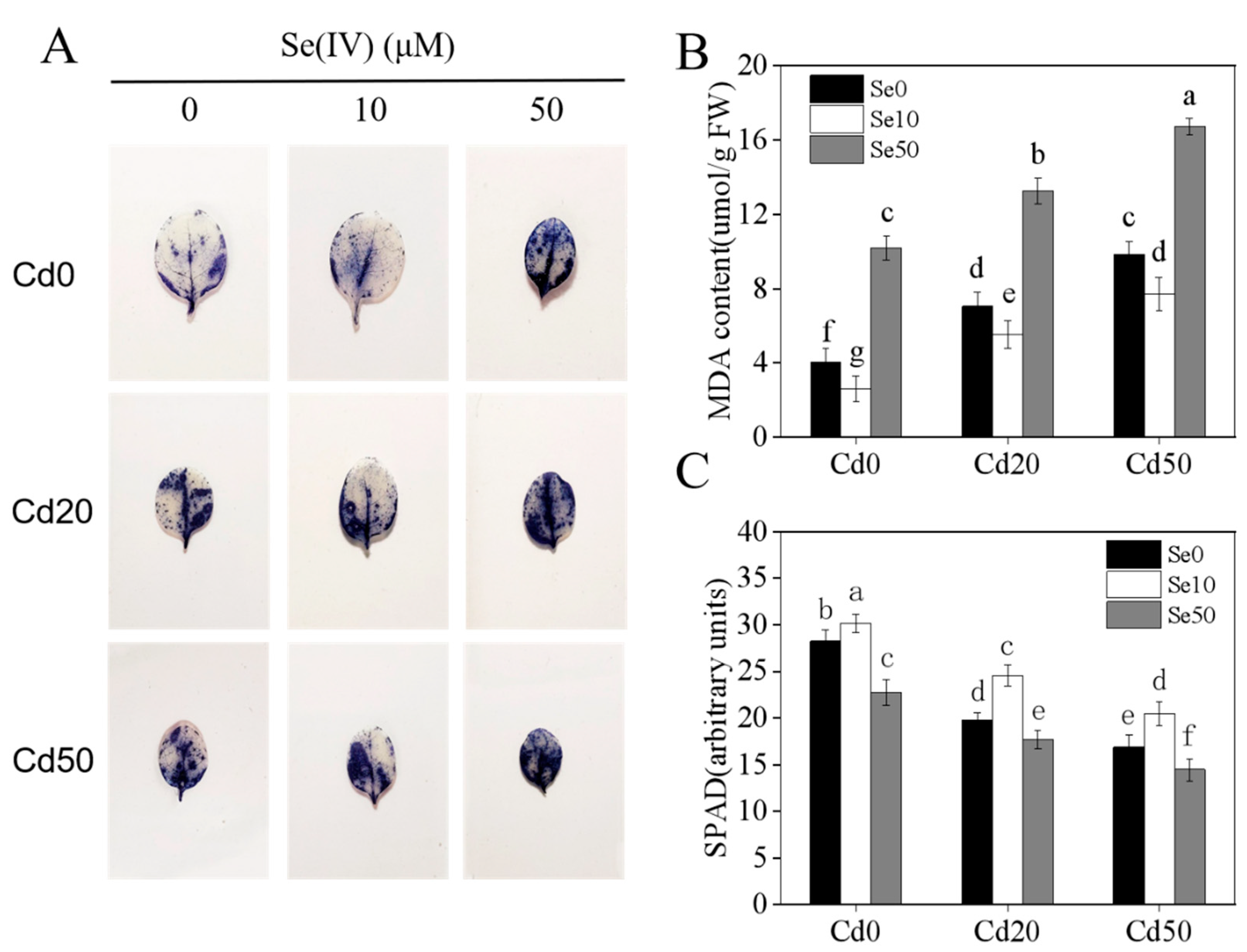
2.2. Effects of Se and Cd Interactions on Tobacco Antioxidant Capacity
2.3. Accumulation of Se and Cd in Tobacco
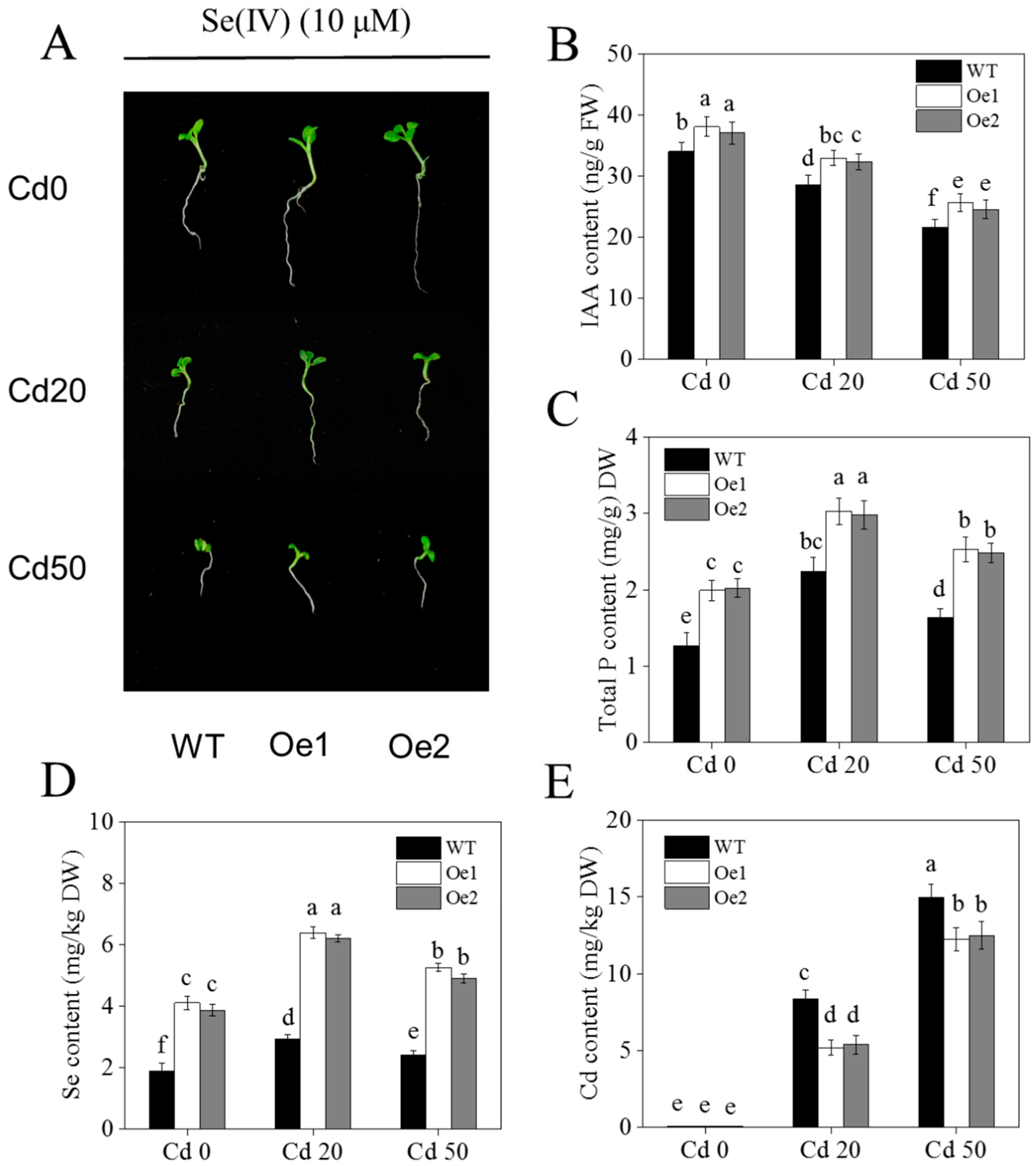
2.4. Effects of Se on Auxin and Expression of Auxin-Related Genes in Tobacco under Cd Stress
2.5. Overexpression of NtPT2 Could Enhance the Tolerance of Cd Stress under Low Se Conditions
3. Discussion
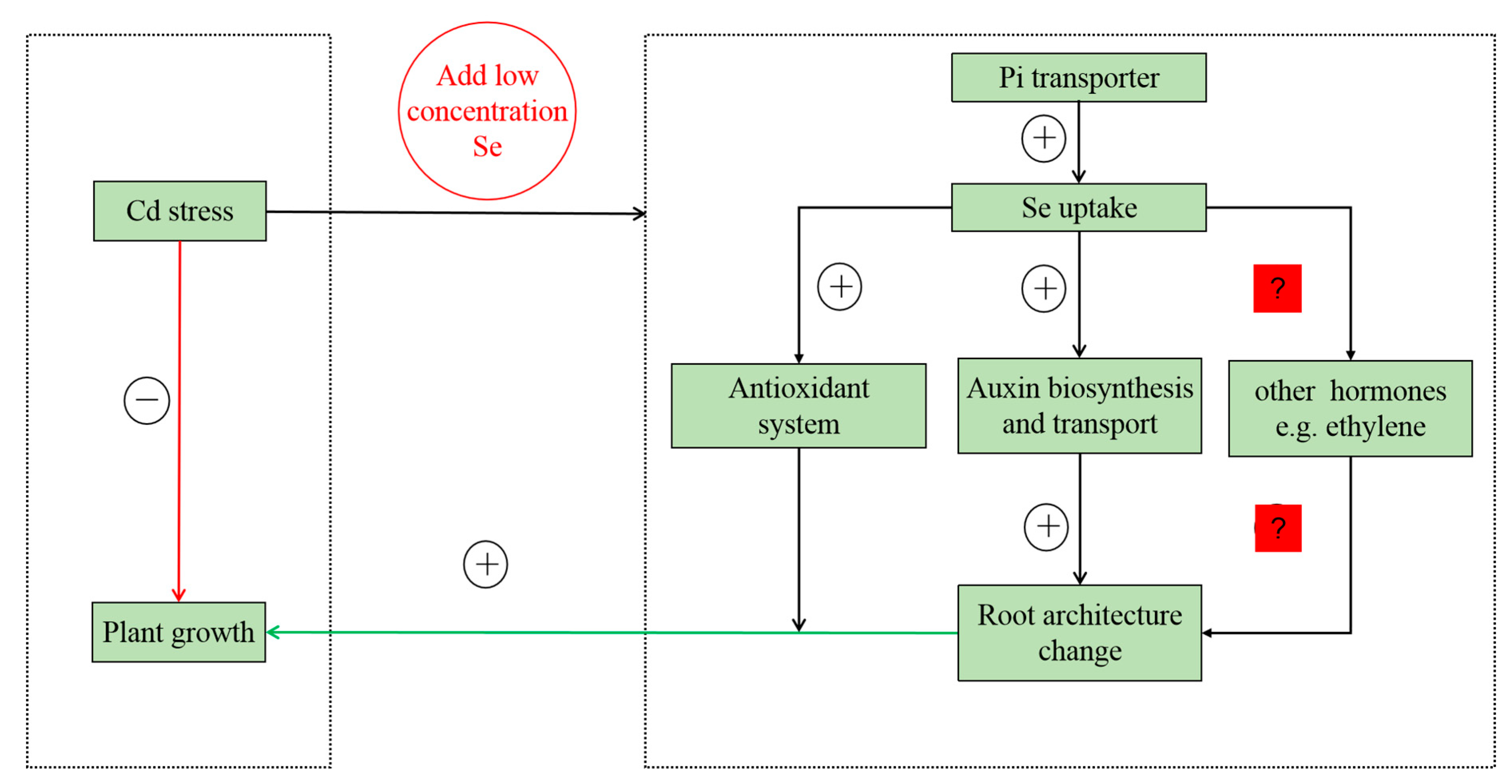
3.1. Se Affects the Growth of Tobacco Roots by Changing Auxin Concentration under Cd Stress
3.2. NtPT2 Might be a Potential Candidate Gene for Breeding Cd-Tolerant Plants
4. Materials and Methods
4.1. Plant Material and Experimental Conditions
4.2. GUS Staining and Nitroblue Tetrazolium (NBT) Staining of Plant Tissues
4.3. Chlorophyll and MDA Measurement
4.4. IAA Measurement
4.5. Reverse Transcription Polymerase Chain Reaction (qRT-PCR) Analysis
4.6. Determination of Total P in Plant
4.7. Measurement of Se and Cd Contents
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Raiesi, F.; Razmkhah, M.; Kiani, S. Salinity stress accelerates the effect of cadmium toxicity on soil N dynamics and cycling: Does joint effect of these stresses matter? Ecotox. Environ. Safe. 2018, 153, 160–167. [Google Scholar] [CrossRef]
- Chai, M.W.; Shi, F.C.; Li, R.L.; Liu, L.M.; Liu, Y.; Liu, F.C. Interactive effects of cadmium and carbon nanotubes on the growth and metal accumulation in a halophyte Spartina alterniflora (Poaceae). Plant Growth Regul. 2013, 71, 171–179. [Google Scholar] [CrossRef]
- Xue, Y.; Wang, Y.Y.; Yao, Q.H.; Song, K.; Zheng, X.Q.; Yang, J.J. Research progress of plants resistance to heavy metal Cd in soil. Ecol. Environ. Sci. 2014, 23, 528–534. [Google Scholar]
- He, X.; Richmond, M.E.; Williams, D.V.; Zheng, W.; Wu, F. Exogenous Glycinebetaine Reduces Cadmium Uptake and Mitigates Cadmium Toxicity in Two Tobacco Genotypes Differing in Cadmium Tolerance. Int. J. Mol. Sci. 2019, 20, 1612. [Google Scholar] [CrossRef]
- Zhang, S.S.; Zhang, H.M.; Qin, R.; Jiang, W.S.; Liu, D.H. Cadmium induction of lipid peroxidation and effects on root tip cells and antioxidant enzyme activities inVicia fabaL. Ecotoxicology 2009, 18, 814–823. [Google Scholar] [CrossRef]
- Dias, M.C.; Monteiro, C.; Moutinho-Pereira, J.; Correia, C.; Gonçalves, B.; Conceição, S. Cadmium toxicity affects photosynthesis and plant growth at different levels. Acta Physiol. Plant 2013, 35, 1281–1289. [Google Scholar] [CrossRef]
- Parmar, P.; Kumari, N.; Sharma, V. Structural and functional alterations in photosynthetic apparatus of plants under cadmium stress. Bot. Stud. 2013, 54, 45. [Google Scholar] [CrossRef]
- Fasahat, P.; Fasahat, P. Advances in Understanding of Cadmium Toxicity and Tolerance in Rice. Emir. J. Food Agr. 2014, 27, 94–105. [Google Scholar] [CrossRef]
- Pinto, F.R.; Mourato, M.P.; Sales, J.R.; Moreira, I.N.; Martins, L.L. Oxidative stress response in spinach plants induced by cadmium. J. Plant Nutr. 2017, 40, 268–276. [Google Scholar] [CrossRef]
- Kaur, N.; Sharma, S.; Kaur, S.; Nayyar, H. Selenium in agriculture: A nutrient or contaminant for crops? Arch. Agron Soil. Sci. 2014, 60, 1593–1624. [Google Scholar] [CrossRef]
- Çakir, Ö.; Turgut-Kara, N.; Ari, Ş. Selenium induced selenocysteine methyltransferase gene expression and antioxidant enzyme activities in Astragalus chrysochlorus. Acta Bot. Croat. 2016, 75, 11–16. [Google Scholar] [CrossRef] [Green Version]
- Hartikainen, H.; Xue, T.; Piironen, V. Selenium as an anti-oxidant and pro-oxidant in ryegrass. Plant Soil. 2000, 225, 193–200. [Google Scholar] [CrossRef]
- Sors, T.G.; Ellis, D.R.; Salt, D.E. Selenium uptake, translocation, assimilation and metabolic fate in plants. Photosynth. Res. 2005, 86, 373–389. [Google Scholar] [CrossRef]
- Zhang, L.H.; Hu, B.; Li, W.; Che, R.H.; Deng, K.; Li, H.; Yu, F.Y.; Ling, H.Q.; Li, Y.J.; Chu, C.C. OsPT2, a phosphate transporter, is involved in the active uptake of selenite in rice. New Phytol. 2014, 201, 1183–1191. [Google Scholar] [CrossRef]
- Li, H.F.; Steve, P.M.; Zhao, F.J. Selenium uptake, translocation and speciation in wheat supplied with selenate or selenite. New Phytol. 2008, 178, 92–102. [Google Scholar] [CrossRef]
- Jia, H.F.; Song, Z.P.; Wu, F.Y.; Ma, M.; Li, Y.T.; Han, D.; Yang, Y.X.; Zhang, S.T.; Cui, H. Low selenium increases the auxin concentration and enhances tolerance to low phosphorous stress in tobacco. Environ. Exp. Bot. 2018, 153, 127–134. [Google Scholar] [CrossRef]
- Cary, E.E. Effect of Selenium and Cadmium Additions to Soil on Their Concentrations in Lettuce and Wheat 1. Agron. J. 1981, 73, 703–706. [Google Scholar] [CrossRef]
- He, P.P.; Lv, X.Z.; Wang, G.Y. Effects of Se and Zn Supplementation on the Antagonism against Pb and Cd in Vegetables. Environ. Int. 2004, 30, 167–172. [Google Scholar] [CrossRef]
- Saidi, I.; Chtourou, Y.; Djebali, W. Selenium alleviates cadmium toxicity by preventing oxidative stress in sunflower (Helianthus annuus) seedlings. J. Plant Physiol. 2014, 171, 85–91. [Google Scholar] [CrossRef]
- Wan, Y.N.; Yu, Y.Y.; Wang, Q.; Qiao, Y.H.; Li, H.F. Cadmium uptake dynamics and translocation in rice seedling: Influence of different forms of selenium. Ecotox. Environ. Safe. 2016, 133, 127–134. [Google Scholar] [CrossRef]
- Qin, X.M.; Nie, Z.J.; Liu, H.G.; Zhao, P.; Qin, S.Y.; Shi, Z.W. Influence of selenium on root morphology and photosynthetic characteristics of winter wheat under cadmium stress. Environ. Exp. Bot. 2018, 150, 232–239. [Google Scholar] [CrossRef]
- Ulmasov, T.; Murfett, J.; Hagen, G.; Guilfoyle, T.J. Aux/IAA proteins repress expression of reporter genes containing natural and highly active synthetic auxin response elements. Plant Cell. 1997, 9, 1963–1971. [Google Scholar]
- Miura, K.; Lee, J.; Gong, Q.Q.; Ma, S.S.; Jin, J.B.; Yoo, C.Y.; Miura, T.; Sato, A.; Bohnert, H.J.; Hasegawa, P.M. SIZ1 Regulation of Phosphate Starvation-Induced Root Architecture Remodeling Involves the Control of Auxin Accumulation. Plant Physiol. 2011, 155, 1000–1012. [Google Scholar] [CrossRef]
- Sabatini, S.; Beis, D.; Wolkenfelt, H.; Murfett, J.; Guilfoyle, T.; Malamy, J.; Benfey, P.; Leyser, O.; Bechtold, N.; Weisbeek, P.; et al. An auxin-dependent distal organizer of pattern and polarity in the Arabidopsis root. Cell 1999, 99, 463–472. [Google Scholar] [CrossRef]
- Ding, Z.; Jirí, F. Auxin regulates distal stem cell differentiation in Arabidopsis roots. Proc. Natl. Acad. Sci. USA 2010, 107, 12046–12051. [Google Scholar] [CrossRef]
- Song, Z.P.; Shao, H.F.; Huang, H.G.; Shen, Y.; Wang, L.Z.; Wu, F.Y.; Han, D.; Song, J.Y.; Jia, H.F. Overexpression of the phosphate transporter gene, OsPT8, improves the Pi and selenium contents in, Nicotiana tabacum. Environ. Exp. Bot. 2017, 137, 158–165. [Google Scholar] [CrossRef]
- Nedjimi, B.; Daoud, Y. Cadmium accumulation in Atriplex halimus subsp. schweinfurthii and its influence on growth, proline, root hydraulic conductivity and nutrient uptake. Flora (Jena) 2009, 204, 316–324. [Google Scholar] [CrossRef]
- Li, H.; Li, X.; Xiang, L.; Zhao, H.M.; Li, Y.W.; Cai, Q.Y.; Zhu, L.; Mo, C.H.; Wong, M.H. Phytoremediation of soil co-contaminated with Cd and BDE-209 using hyperaccumulator enhanced by AM fungi and surfactant. Sci. Total Environ. 2017, 613–614, 447–455. [Google Scholar] [CrossRef]
- Li, L.; Zhou, W.H.; Dai, H.X.; Cao, F.B.; Zhang, G.P.; Wu, F.B. Selenium reduces cadmium uptake and mitigates cadmium toxicity in rice. J. Hazard. Mater. 2012, 235–236, 343–351. [Google Scholar]
- Han, D.; Xiong, S.L.; Tu, S.X.; Liu, J.C.; Chen, C. Interactive effects of selenium and arsenic on growth, antioxidant system, arsenic and selenium species of Nicotiana tabacum L. Environ. Exp. Bot. 2015, 117, 12–19. [Google Scholar] [CrossRef]
- Ding, Y.Z.; Feng, R.W.; Wang, R.G.; Guo, J.K.; Zheng, X.Q. A dual effect of Se on Cd toxicity: Evidence from plant growth, root morphology and responses of the antioxidative systems of paddy rice. Plant Soil. 2014, 375, 289–301. [Google Scholar] [CrossRef]
- Rafael, S.P.M.; Lucas, C.C.; Rodrigo, T.Á.; Thaline, M.P.; Lubia, S.T.; Fred, A.L.B.; Agustín, Z.; Wagner, L.A.; Ribeiro, D.M. Selenium downregulates auxin and ethylene biosynthesis in rice seedlings to modify primary metabolism and root architecture. Planta 2019, 250, 333–345. [Google Scholar]
- Ribeiro, D.M.; Silva, J.; Dalton, D.; Cardoso, F.B.; Martins, A.O.; Silva, W.A.; Nascimento, V.L.; Araújo, W.L. Growth inhibition by selenium is associated with changes in primary metabolism and nutrient levels inArabidopsis thaliana. Plant Cell Environ. 2016, 39, 2235–2246. [Google Scholar] [CrossRef]
- Lehotai, N.; Kolbert, Z.; Peto, A.; Feigl, G.; Ordog, A.; Kumar, D.; Tari, I.; Erdei, L. Selenite-induced hormonal and signalling mechanisms during root growth of Arabidopsis thaliana L. J. Exp. Bot. 2012, 63, 5677–5687. [Google Scholar] [CrossRef] [Green Version]
- Raghothama, K.G. Phosphate transport and signaling. Curr. Opin. Plant Biol. 2000, 3, 182–187. [Google Scholar] [CrossRef]
- Rausch, C.; Bucher, M. Molecular mechanisms of phosphate transport in plants. Planta (Berl.) 2002, 216, 23–37. [Google Scholar] [CrossRef]
- Yu, J.; Hu, S.; Wang, J.; Wong, G.K.; Li, S.; Liu, B.; Deng, Y.; Dai, L.; Zhou, Y.; Zhang, X.; et al. A Draft Sequence of the Rice Genome (Oryza sativa L. ssp. japonica). Science 2002, 296, 92–100. [Google Scholar] [CrossRef]
- Ai, P.H.; Sun, S.B.; Zhao, J.N.; Fan, X.R.; Xin, W.J.; Guo, Q.; Yu, L.; Shen, Q.R.; Wu, P.; Miller, A.J.; et al. Two rice phosphate transporters, OsPht1;2 and OsPht1;6, have different functions and kinetic properties in uptake and translocation. Plant J. 2009, 57, 798–809. [Google Scholar] [CrossRef]
- Wang, P.; Zhang, W.; Mao, C.; Xu, G.; Zhao, F.J. The role of OsPT8 in arsenate uptake and varietal difference in arsenate tolerance in rice. J. Exp. Bot. 2016, 67, 6051–6059. [Google Scholar] [CrossRef]
- Xie, L.H.; Xu, Z.R. The toxicity of heavy metal Cadmium to animals and humans. Acta Agric. Zhejianggensis 2003, 6, 52–57. (In Chinese) [Google Scholar]
- Prozialeck, W.C.; Edwards, J.R.; Nebert, D.W.; Woods, J.M.; Barchowsky, A.; Atchison, W.D. The Vascular System as a Target of Metal Toxicity. Toxicol. Sci. 2007, 102, 207–218. [Google Scholar] [CrossRef] [Green Version]
- Jia, H.F.; Zhang, S.T.; Wang, L.Z.; Yang, Y.X.; Zhang, H.Y.; Cui, H.; Shao, H.F.; Xu, G.H. OsPht1;8, a phosphate transporter, is involved in auxin and phosphate starvation response in rice. J. Exp. Bot. 2017, 68, 5057–5068. [Google Scholar] [CrossRef]
- Li, Z.; Tan, X.F.; Lu, K.; Zhang, L.; Long, H.X.; Lv, J.B.; Lin, Q. Influence of drought stress on the growth, leaf gas exchange, and chlorophyll fluorescence in two varieties of tung tree seedlings. Acta Ecol. Sin. 2017, 37, 1515–1524. [Google Scholar]
- Feng, R.W.; Wei, C.Y.; Tu, S.X.; Sun, X. Interactive effects of selenium and arsenic on their uptake by Pteris vittata L. under hydroponic conditions. Environ. Exp. Bot. 2009, 65, 363–368. [Google Scholar] [CrossRef]
- Han, S.; Fang, L.; Ren, X.J.; Wang, W.L.; Jiang, J. MPK6 controls H2O2-induced root elongation by mediating Ca2+ influx across the plasma membrane of root cells in Arabidopsis seedlings. New Phytol. 2015, 205, 695–706. [Google Scholar] [CrossRef]
- Sun, H.W.; Tao, J.Y.; Liu, S.J.; Huang, S.J.; Chen, S.; Yoneyama, Y.; Zhang, Y.L.; Xu, G.H. Strigolactones are involved in phosphate- and nitrate-deficiency-induced root development and auxin transport in rice. J. Exp. Bot. 2014, 65, 6735–6746. [Google Scholar] [CrossRef]
- Chen, A.Q.; Hu, J.; Sun, S.B.; Xu, G.H. Conservation and divergence of both phosphate- and mycorrhiza-regutelad physiological responses and expression patterns of phosphate transporters in solanaceous species. New Phytol. 2007, 173, 817–831. [Google Scholar] [CrossRef]
- Wei, C.Y.; Sun, X.; Wang, C.; Wang, W.Y. Factors influencing arsenic accumulation by Pteris vittata: A comparative field study at two sites. Environ. Pollut. 2006, 141, 488–493. [Google Scholar] [CrossRef]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, Y.; Wei, Y.; Sun, S.; Wang, J.; Wang, W.; Han, D.; Shao, H.; Jia, H.; Fu, Y. Selenium Modulates the Level of Auxin to Alleviate the Toxicity of Cadmium in Tobacco. Int. J. Mol. Sci. 2019, 20, 3772. https://doi.org/10.3390/ijms20153772
Luo Y, Wei Y, Sun S, Wang J, Wang W, Han D, Shao H, Jia H, Fu Y. Selenium Modulates the Level of Auxin to Alleviate the Toxicity of Cadmium in Tobacco. International Journal of Molecular Sciences. 2019; 20(15):3772. https://doi.org/10.3390/ijms20153772
Chicago/Turabian StyleLuo, Yong, Yuewei Wei, Shuguang Sun, Jian Wang, Weifeng Wang, Dan Han, Huifang Shao, Hongfang Jia, and Yunpeng Fu. 2019. "Selenium Modulates the Level of Auxin to Alleviate the Toxicity of Cadmium in Tobacco" International Journal of Molecular Sciences 20, no. 15: 3772. https://doi.org/10.3390/ijms20153772
APA StyleLuo, Y., Wei, Y., Sun, S., Wang, J., Wang, W., Han, D., Shao, H., Jia, H., & Fu, Y. (2019). Selenium Modulates the Level of Auxin to Alleviate the Toxicity of Cadmium in Tobacco. International Journal of Molecular Sciences, 20(15), 3772. https://doi.org/10.3390/ijms20153772