The Interaction of Helicobacter pylori with TFF1 and Its Role in Mediating the Tropism of the Bacteria Within the Stomach

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. Site-Specific Tropism of Helicobacter Pylori in the Stomach
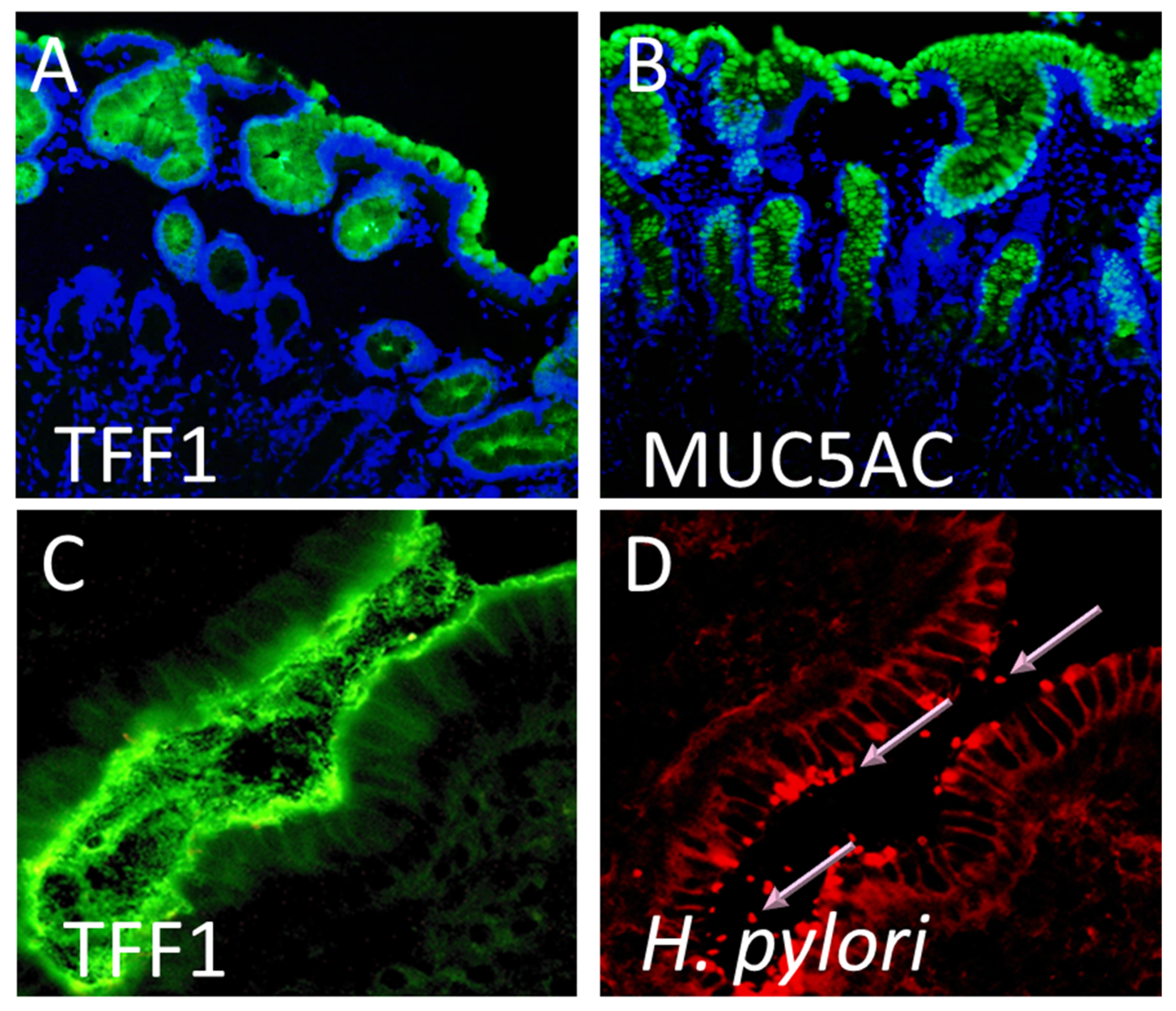
1.2. Site-Specific Localisation of Trefoil Proteins and Co-Expression with Mucins in the Gastrointestinal Tract
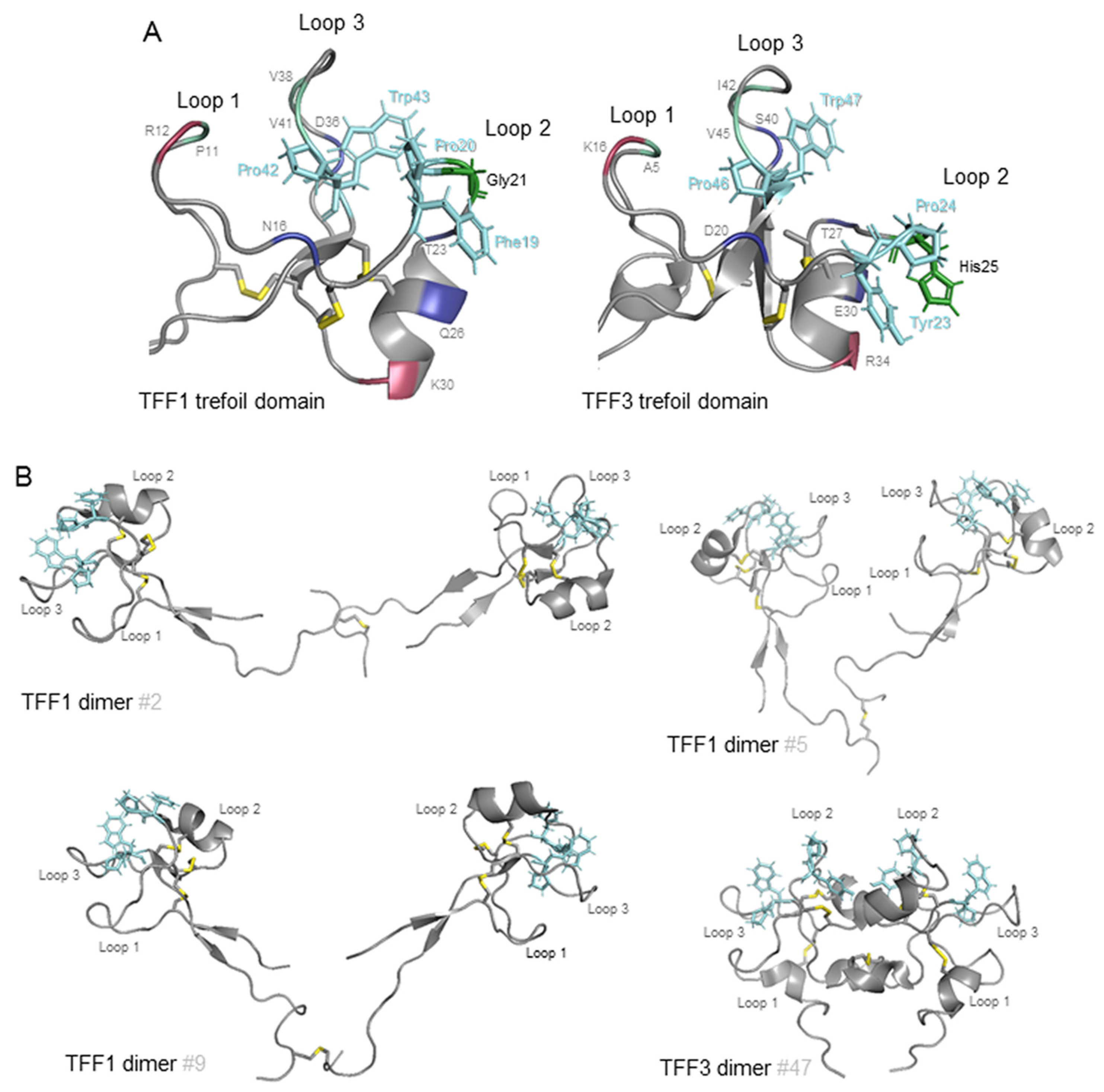
2. Helicobacter pylori Interacts with the Dimeric Form of TFF1
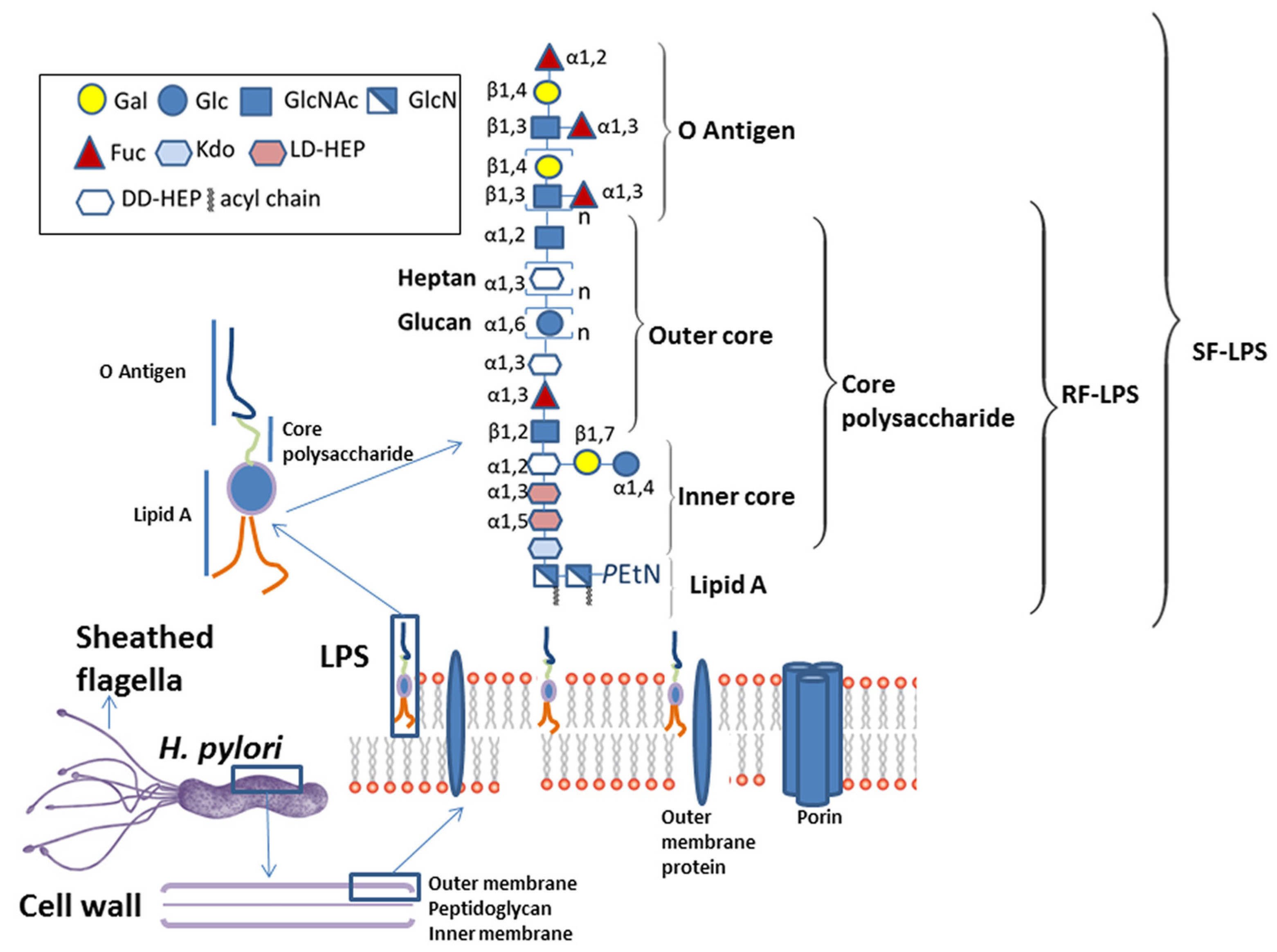
3. The Interaction of H. pylori with TFF1 is Mediated via Glycans Present on the Core Oligosaccharide Region of Lipopolysaccharide
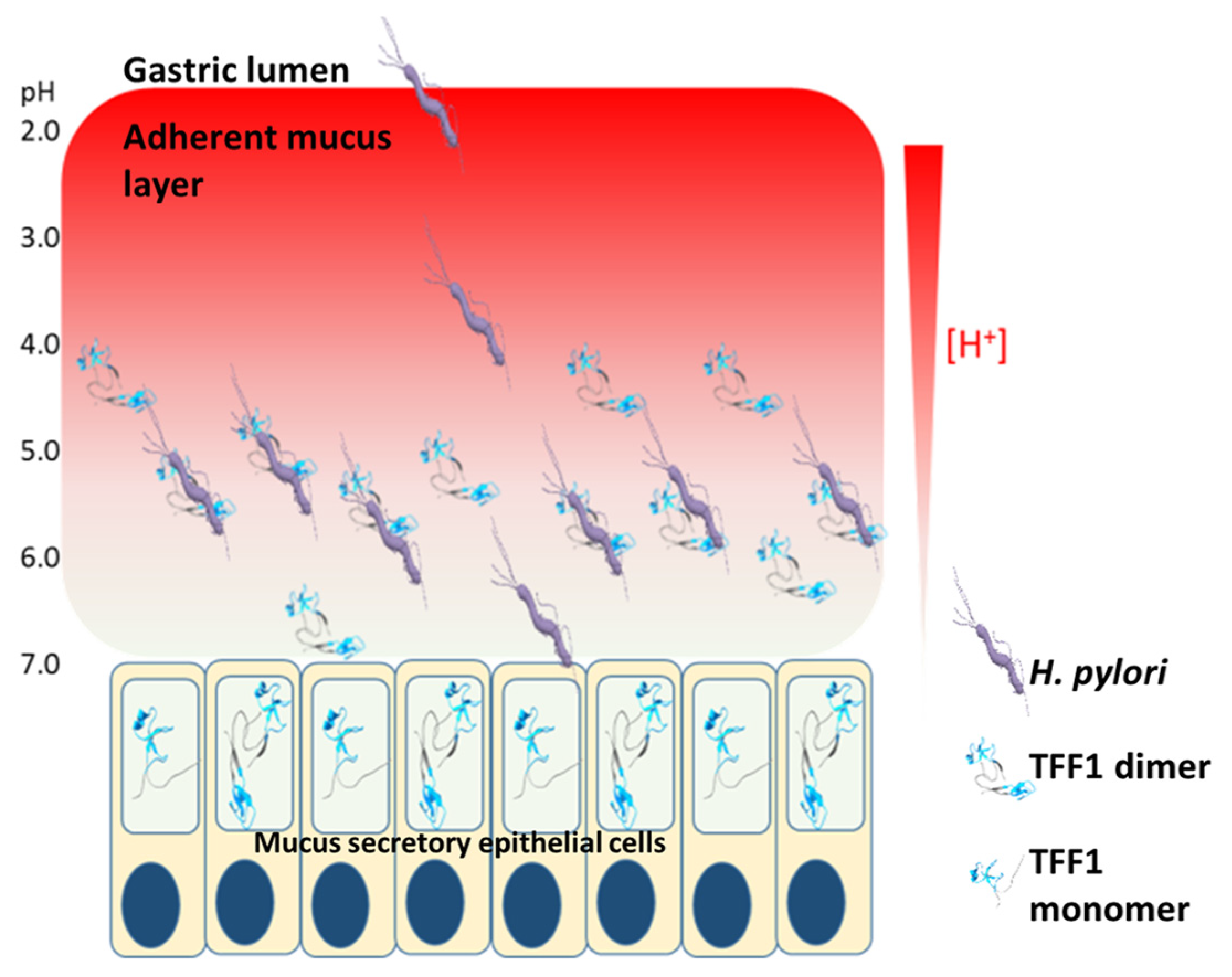
4. The Interaction of H. pylori with TFF1 Explains Site-Specific Localisation of the Bacteria Within the Stomach
Funding
Conflicts of Interest
References
- Hooi, J.K.Y.; Lai, W.Y.; Ng, W.K.; Suen, M.M.Y.; Underwood, F.E.; Tanyingoh, D.; Malfertheiner, P.; Graham, D.Y.; Wong, V.W.S.; Wu, J.C.Y.; et al. Global prevalence of Helicobacter pylori infection: Systematic review and meta-analysis. Gastroenterology 2017, 153, 420–429. [Google Scholar] [CrossRef] [PubMed]
- Drumm, B.; Perez-Perez, G.I.; Blaser, M.J.; Sherman, P.M. Intrafamilial clustering of Helicobacter pylori infection. N. Engl. J. Med. 1990, 322, 359–363. [Google Scholar] [CrossRef] [PubMed]
- Rowland, M.; Daly, L.; Vaughan, M.; Higgins, A.; Bourke, B.; Drumm, B. Age-specific incidence of Helicobacter pylori. Gastroenterology 2006, 130, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Blanchard, T.G.; Czinn, S.J. Identification of Helicobacter pylori and the evolution of an efficacious childhood vaccine to protect against gastritis and peptic ulcer disease. Pediatr. Res. 2017, 81, 170–176. [Google Scholar] [CrossRef] [PubMed]
- McColl, K.E. Clinical practice. Helicobacter pylori infection. N. Engl. J. Med. 2010, 362, 1597–1604. [Google Scholar] [PubMed]
- Leow, A.H.; Lim, Y.Y.; Liew, W.C.; Goh, K.L. Time trends in upper gastrointestinal diseases and Helicobacter pylori infection in a multiracial asian population—a 20-year experience over three time periods. Aliment. Pharmacol. Ther. 2016, 43, 831–837. [Google Scholar] [CrossRef]
- Sonnenberg, A. Review article: Historic changes of Helicobacter pylori-associated diseases. Aliment. Pharmacol. Ther. 2013, 38, 329–342. [Google Scholar] [CrossRef]
- Plummer, M.; Franceschi, S.; Vignat, J.; Forman, D.; de Martel, C. Global burden of gastric cancer attributable to Helicobacter pylori. Int. J. Cancer 2015, 136, 487–490. [Google Scholar] [CrossRef]
- Howitt, M.R.; Lee, J.Y.; Lertsethtakarn, P.; Vogelmann, R.; Joubert, L.M.; Ottemann, K.M.; Amieva, M.R. ChePep controls Helicobacter pylori infection of the gastric glands and chemotaxis in the Epsilonproteobacteria. MBio 2011, 2. [Google Scholar] [CrossRef]
- Keilberg, D.; Zavros, Y.; Shepherd, B.; Salama, N.R.; Ottemann, K.M. Spatial and temporal shifts in bacterial biogeography and gland occupation during the development of a chronic infection. MBio 2016, 7. [Google Scholar] [CrossRef]
- Fung, C.; Tan, S.; Nakajima, M.; Skoog, E.C.; Camarillo-Guerrero, L.F.; Klein, J.A.; Lawley, T.D.; Solnick, J.V.; Fukami, T.; Amieva, M.R. High-resolution mapping reveals that microniches in the gastric glands control Helicobacter pylori colonization of the stomach. PLoS Biol. 2019, 17, e3000231. [Google Scholar] [CrossRef] [PubMed]
- Hessey, S.J.; Spencer, J.; Wyatt, J.I.; Sobala, G.; Rathbone, B.J.; Axon, A.T.; Dixon, M.F. Bacterial adhesion and disease activity in Helicobacter associated chronic gastritis. Gut 1990, 31, 134–138. [Google Scholar] [CrossRef] [PubMed]
- Hidaka, E.; Ota, H.; Hidaka, H.; Hayama, M.; Matsuzawa, K.; Akamatsu, T.; Nakayama, J.; Katsuyama, T. Helicobacter pylori and two ultrastructurally distinct layers of gastric mucous cell mucins in the surface mucous gel layer. Gut 2001, 49, 474–480. [Google Scholar] [CrossRef] [PubMed]
- Wyatt, J.I.; Rathbone, B.J.; Sobala, G.M.; Shallcross, T.; Heatley, R.V.; Axon, A.T.; Dixon, M.F. Gastric epithelium in the duodenum: Its association with Helicobacter pylori and inflammation. J. Clin. Pathol 1990, 43, 981–986. [Google Scholar] [CrossRef] [PubMed]
- Van de Bovenkamp, J.H.; Mahdavi, J.; Korteland-Van Male, A.M.; Buller, H.A.; Einerhand, A.W.; Boren, T.; Dekker, J. The muc5ac glycoprotein is the primary receptor for Helicobacter pylori in the human stomach. Helicobacter 2003, 8, 521–532. [Google Scholar] [CrossRef] [PubMed]
- Van den Brink, G.R.; Tytgat, K.M.; Van der Hulst, R.W.; Van der Loos, C.M.; Einerhand, A.W.; Buller, H.A.; Dekker, J.H. pylori colocalises with MUC5AC in the human stomach. Gut 2000, 46, 601–607. [Google Scholar] [CrossRef]
- Genta, R.M.; Gurer, I.E.; Graham, D.Y.; Krishnan, B.; Segura, A.M.; Gutierrez, O.; Kim, J.G.; Burchette, J.L., Jr. Adherence of Helicobacter pylori to areas of incomplete intestinal metaplasia in the gastric mucosa. Gastroenterology 1996, 111, 1206–1211. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, A.; David, L.; Reis, C.A.; Costa, J.; Sobrinho-Simoes, M. Expression of mucins (MUC1, MUC2, MUC5AC, and MUC6) and type 1 Lewis antigens in cases with and without Helicobacter pylori colonization in metaplastic glands of the human stomach. J. Pathol. 2002, 197, 37–43. [Google Scholar] [CrossRef]
- Johansson, M.E.; Sjovall, H.; Hansson, G.C. The gastrointestinal mucus system in health and disease. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 352–361. [Google Scholar] [CrossRef] [Green Version]
- Lewis, O.L.; Keener, J.P.; Fogelson, A.L. A physics-based model for maintenance of the ph gradient in the gastric mucus layer. Am. J. Physiol. Gastrointest. Liver Physiol. 2017, 313, G599–G612. [Google Scholar] [CrossRef]
- Clyne, M.; Labigne, A.; Drumm, B. Helicobacter pylori requires an acidic environment to survive in the presence of urea. Infect. Immun. 1995, 63, 1669–1673. [Google Scholar] [PubMed]
- Schreiber, S.; Konradt, M.; Groll, C.; Scheid, P.; Hanauer, G.; Werling, H.O.; Josenhans, C.; Suerbaum, S. The spatial orientation of Helicobacter pylori in the gastric mucus. Proc. Natl. Acad. Sci. USA 2004, 101, 5024–5029. [Google Scholar] [CrossRef] [PubMed]
- Ilver, D.; Arnqvist, A.; Ogren, J.; Frick, I.M.; Kersulyte, D.; Incecik, E.T.; Berg, D.E.; Covacci, A.; Engstrand, L.; Boren, T. Helicobacter pylori adhesin binding fucosylated histo-blood group antigens revealed by retagging. Science 1998, 279, 373–377. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.; Barone, A.; Boren, T.; Teneberg, S. Helicobacter pylori-binding nonacid glycosphingolipids in the human stomach. J. Biol. Chem. 2018, 293, 17248–17266. [Google Scholar] [CrossRef] [PubMed]
- Mahdavi, J.; Sonden, B.; Hurtig, M.; Olfat, F.O.; Forsberg, L.; Roche, N.; Angstrom, J.; Larsson, T.; Teneberg, S.; Karlsson, K.A.; et al. Helicobacter pylori SabA adhesin in persistent infection and chronic inflammation. Science 2002, 297, 573–578. [Google Scholar] [CrossRef]
- Rossez, Y.; Gosset, P.; Boneca, I.G.; Magalhaes, A.; Ecobichon, C.; Reis, C.A.; Cieniewski-Bernard, C.; Joncquel Chevalier Curt, M.; Leonard, R.; Maes, E.; et al. The lacdinac-specific adhesin LabA mediates adhesion of Helicobacter pylori to human gastric mucosa. J. Infect. Dis. 2014, 210, 1286–1295. [Google Scholar] [CrossRef]
- May, F.E.; Westley, B.R. Trefoil proteins: Their role in normal and malignant cells. J. Pathol 1997, 183, 4–7. [Google Scholar] [CrossRef]
- Thim, L.; Madsen, F.; Poulsen, S.S. Effect of trefoil factors on the viscoelastic properties of mucus gels. Eur. J. Clin. Investig. 2002, 32, 519–527. [Google Scholar] [CrossRef]
- Thim, L.; May, F.E. Structure of mammalian trefoil factors and functional insights. Cell Mol. Life Sci. 2005, 62, 2956–2973. [Google Scholar] [CrossRef]
- Chadwick, M.P.; Westley, B.R.; May, F.E. Homodimerization and hetero-oligomerization of the single-domain trefoil protein pNR-2/pS2 through cysteine 58. Biochem. J. 1997, 327(Pt. 1), 117–123. [Google Scholar] [CrossRef]
- May, F.E.; Church, S.T.; Major, S.; Westley, B.R. The closely related estrogen-regulated trefoil proteins TFF1 and TFF3 have markedly different hydrodynamic properties, overall charge, and distribution of surface charge. Biochemistry 2003, 42, 8250–8259. [Google Scholar] [CrossRef] [PubMed]
- Masiakowski, P.; Breathnach, R.; Bloch, J.; Gannon, F.; Krust, A.; Chambon, P. Cloning of cDNA sequences of hormone-regulated genes from the MCF-7 human breast cancer cell line. Nucleic Acids Res. 1982, 10, 7895–7903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- May, F.E.; Westley, B.R. Cloning of estrogen-regulated messenger RNA sequences from human breast cancer cells. Cancer Res. 1986, 46, 6034–6040. [Google Scholar] [PubMed]
- Jorgensen, K.H.; Thim, L.; Jacobsen, H.E. Pancreatic spasmolytic polypeptide (pSP): I. Preparation and initial chemical characterization of a new polypeptide from porcine pancreas. Regul. Pept. 1982, 3, 207–219. [Google Scholar] [CrossRef]
- Suemori, S.; Lynch-Devaney, K.; Podolsky, D.K. Identification and characterization of rat intestinal trefoil factor: Tissue- and cell-specific member of the trefoil protein family. Proc. Natl. Acad. Sci. USA 1991, 88, 11017–11021. [Google Scholar] [CrossRef] [PubMed]
- Madsen, J.; Nielsen, O.; Tornoe, I.; Thim, L.; Holmskov, U. Tissue localization of human trefoil factors 1, 2, and 3. J. Histochem. Cytochem. 2007, 55, 505–513. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos Silva, E.; Ulrich, M.; Doring, G.; Botzenhart, K.; Gott, P. Trefoil factor family domain peptides in the human respiratory tract. J. Pathol. 2000, 190, 133–142. [Google Scholar] [CrossRef]
- Ahmed, A.R.H.; Griffiths, A.B.; Tilby, M.T.; Westley, B.R.; May, F.E.B. TFF3 is a normal breast epithelial protein and is associated with differentiated phenotype in early breast cancer but predisposes to invasion and metastasis in advanced disease. Am. J. Pathol. 2012, 180, 904–916. [Google Scholar] [CrossRef]
- Piggott, N.H.; Henry, J.A.; May, F.E.; Westley, B.R. Antipeptide antibodies against the pNR-2 oestrogen-regulated protein of human breast cancer cells and detection of pNR-2 expression in normal tissues by immunohistochemistry. J. Pathol. 1991, 163, 95–104. [Google Scholar] [CrossRef]
- May, F.E.B.; Westley, B.R. TFF3 is a valuable predictive biomarker of endocrine response in metastatic breast cancer. Endocr.-Relat. Cancer 2015, 22, 465–479. [Google Scholar] [CrossRef] [Green Version]
- May, F.E.; Semple, J.I.; Prest, S.J.; Westley, B.R. Expression and motogenic activity of TFF2 in human breast cancer cells. Peptides 2004, 25, 865–872. [Google Scholar] [CrossRef] [PubMed]
- Ruchaud-Sparagano, M.H.; Westley, B.R.; May, F.E. The trefoil protein TFF1 is bound to MUC5AC in human gastric mucosa. Cell. Mol. Life Sci. 2004, 61, 1946–1954. [Google Scholar] [CrossRef] [PubMed]
- Hanby, A.M.; Poulsom, R.; Elia, G.; Singh, S.; Longcroft, J.M.; Wright, N.A. The expression of the trefoil peptides pS2 and human spasmolytic polypeptide (hSP) in ‘gastric metaplasia’ of the proximal duodenum: Implications for the nature of ‘gastric metaplasia’. J. Pathol. 1993, 169, 355–360. [Google Scholar] [CrossRef] [PubMed]
- May, F.E.; Griffin, S.M.; Westley, B.R. The trefoil factor interacting protein TFIZ1 binds the trefoil protein TFF1 preferentially in normal gastric mucosal cells but the co-expression of these proteins is deregulated in gastric cancer. Int. J. Biochem. Cell. Biol. 2009, 41, 632–640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newton, J.L.; Allen, A.; Westley, B.R.; May, F.E. The human trefoil peptide, TFF1, is present in different molecular forms that are intimately associated with mucus in normal stomach. Gut 2000, 46, 312–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westley, B.R.; Griffin, S.M.; May, F.E. Interaction between TFF1, a gastric tumor suppressor trefoil protein, and TFIZ1, a brichos domain-containing protein with homology to sp-c. Biochemistry 2005, 44, 7967–7975. [Google Scholar] [CrossRef] [PubMed]
- Menheniott, T.R.; Peterson, A.J.; O’Connor, L.; Lee, K.S.; Kalantzis, A.; Kondova, I.; Bontrop, R.E.; Bell, K.M.; Giraud, A.S. A novel gastrokine, GKN3, marks gastric atrophy and shows evidence of adaptive gene loss in humans. Gastroenterology 2010, 138, 1823–1835. [Google Scholar] [CrossRef] [PubMed]
- Clyne, M.; Dillon, P.; Daly, S.; O’Kennedy, R.; May, F.E.; Westley, B.R.; Drumm, B. Helicobacter pylori interacts with the human single-domain trefoil protein TFF1. Proc. Natl. Acad. Sci. USA 2004, 101, 7409–7414. [Google Scholar] [CrossRef]
- Jonsson, U.; Fagerstam, L.; Ivarsson, B.; Johnsson, B.; Karlsson, R.; Lundh, K.; Lofas, S.; Persson, B.; Roos, H.; Ronnberg, I.; et al. Real-time biospecific interaction analysis using surface plasmon resonance and a sensor chip technology. Biotechniques 1991, 11, 620–627. [Google Scholar]
- Josenhans, C.; Labigne, A.; Suerbaum, S. Comparative ultrastructural and functional studies of Helicobacter pylori and Helicobacter mustelae flagellin mutants: Both flagellin subunits, FlaA and FlaB, are necessary for full motility in Helicobacter species. J. Bacteriol. 1995, 177, 3010–3020. [Google Scholar] [CrossRef]
- Aspinall, G.O.; Monteiro, M.A. Lipopolysaccharides of Helicobacter pylori strains P466 and MO19: Structures of the O antigen and core oligosaccharide regions. Biochemistry 1996, 35, 2498–2504. [Google Scholar] [CrossRef]
- Aspinall, G.O.; Monteiro, M.A.; Pang, H.; Walsh, E.J.; Moran, A.P. Lipopolysaccharide of the Helicobacter pylori type strain NCTC 11637 (ATCC 43504): Structure of the o antigen chain and core oligosaccharide regions. Biochemistry 1996, 35, 2489–2497. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, M.A.; Chan, K.H.; Rasko, D.A.; Taylor, D.E.; Zheng, P.Y.; Appelmelk, B.J.; Wirth, H.P.; Yang, M.; Blaser, M.J.; Hynes, S.O.; et al. Simultaneous expression of type 1 and type 2 Lewis blood group antigens by Helicobacter pylori lipopolysaccharides. Molecular mimicry between H. pylori lipopolysaccharides and human gastric epithelial cell surface glycoforms. J. Biol. Chem. 1998, 273, 11533–11543. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, M.A.; Rasko, D.; Taylor, D.E.; Perry, M.B. Glucosylated n-acetyllactosamine o-antigen chain in the lipopolysaccharide from Helicobacter pylori strainUA861. Glycobiology 1998, 8, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Moran, A.P.; Lindner, B.; Walsh, E.J. Structural characterization of the Lipid A component of Helicobacter pylori rough- and smooth-form lipopolysaccharides. J. Bacteriol. 1997, 179, 6453–6463. [Google Scholar] [CrossRef] [PubMed]
- Cullen, T.W.; Giles, D.K.; Wolf, L.N.; Ecobichon, C.; Boneca, I.G.; Trent, M.S. Helicobacter pylori versus the host: Remodeling of the bacterial outer membrane is required for survival in the gastric mucosa. PLoS Pathog. 2011, 7, e1002454. [Google Scholar] [CrossRef] [PubMed]
- Moran, A.P. Relevance of fucosylation and Lewis antigen expression in the bacterial gastroduodenal pathogen Helicobacter pylori. Carbohydr. Res. 2008, 343, 1952–1965. [Google Scholar] [CrossRef]
- Edwards, N.J.; Monteiro, M.A.; Faller, G.; Walsh, E.J.; Moran, A.P.; Roberts, I.S.; High, N.J. Lewis x structures in the O antigen side-chain promote adhesion of Helicobacter pylori to the gastric epithelium. Mol. Microbiol. 2000, 35, 1530–1539. [Google Scholar] [CrossRef]
- Li, H.; Liao, T.; Debowski, A.W.; Tang, H.; Nilsson, H.O.; Stubbs, K.A.; Marshall, B.J.; Benghezal, M. Lipopolysaccharide structure and biosynthesis in Helicobacter pylori. Helicobacter 2016, 21, 445–461. [Google Scholar] [CrossRef]
- Reeves, E.P.; Ali, T.; Leonard, P.; Hearty, S.; O’Kennedy, R.; May, F.E.; Westley, B.R.; Josenhans, C.; Rust, M.; Suerbaum, S.; et al. Helicobacter pylori lipopolysaccharide interacts with TFF1 in a pH-dependent manner. Gastroenterology 2008, 135, 2043–2054, 2054 e2041–e2042. [Google Scholar] [CrossRef]
- Moran, A.P.; Helander, I.M.; Kosunen, T.U. Compositional analysis of Helicobacter pylori rough-form lipopolysaccharides. J. Bacteriol. 1992, 174, 1370–1377. [Google Scholar] [CrossRef] [PubMed]
- Leker, K.; Lozano-Pope, I.; Bandyopadhyay, K.; Choudhury, B.P.; Obonyo, M. Comparison of lipopolysaccharides composition of two different strains of Helicobacter pylori. BMC Microbiol. 2017, 17, 226. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.; O’Rourke, J.; de Ungria, M.C.; Robertson, B.; Daskalopoulos, G.; Dixon, M.F. A standardized mouse model of Helicobacter pylori infection: Introducing the Sydney strain. Gastroenterology 1997, 112, 1386–1397. [Google Scholar] [CrossRef]
- Covacci, A.; Censini, S.; Bugnoli, M.; Petracca, R.; Burroni, D.; Macchia, G.; Massone, A.; Papini, E.; Xiang, Z.; Figura, N.; et al. Molecular characterization of the 128-kda immunodominant antigen of Helicobacter pylori associated with cytotoxicity and duodenal ulcer. Proc. Natl. Acad. Sci. USA 1993, 90, 5791–5795. [Google Scholar] [CrossRef] [PubMed]
- Dunne, C.; Naughton, J.; Duggan, G.; Loughrey, C.; Kilcoyne, M.; Joshi, L.; Carrington, S.; Earley, H.; Backert, S.; Robbe Masselot, C.; et al. Binding of Helicobacter pylori to human gastric mucins correlates with binding of TFF1. Microorganisms 2018, 6, 44. [Google Scholar] [CrossRef] [PubMed]
- Polshakov, V.I.; Williams, M.A.; Gargaro, A.R.; Frenkiel, T.A.; Westley, B.R.; Chadwick, M.P.; May, F.E.; Feeney, J. High-resolution solution structure of human pNR-2/pS2: A single trefoil motif protein. J. Mol. Biol. 1997, 267, 418–432. [Google Scholar] [CrossRef]
- Muskett, F.W.; May, F.E.; Westley, B.R.; Feeney, J. Solution structure of the disulfide-linked dimer of human intestinal trefoil factor (TFF3): The intermolecular orientation and interactions are markedly different from those of other dimeric trefoil proteins. Biochemistry 2003, 42, 15139–15147. [Google Scholar] [CrossRef] [PubMed]
- Williams, M.A.; Westley, B.R.; May, F.E.; Feeney, J. The solution structure of the disulphide-linked homodimer of the human trefoil protein TFF1. FEBS Lett. 2001, 493, 70–74. [Google Scholar] [CrossRef]
- Schreiber, S.; Bucker, R.; Groll, C.; Azevedo-Vethacke, M.; Garten, D.; Scheid, P.; Friedrich, S.; Gatermann, S.; Josenhans, C.; Suerbaum, S. Rapid loss of motility of Helicobacter pylori in the gastric lumen in vivo. Infect. Immun. 2005, 73, 1584–1589. [Google Scholar] [CrossRef]
- Dolan, B.; Naughton, J.; Tegtmeyer, N.; May, F.E.; Clyne, M. The interaction of Helicobacter pylori with the adherent mucus gel layer secreted by polarized HT29-MTX-E12 cells. PLoS ONE 2012, 7, e47300. [Google Scholar] [CrossRef]
- Behrens, I.; Stenberg, P.; Artursson, P.; Kissel, T. Transport of lipophilic drug molecules in a new mucus-secreting cell culture model based on HT29-MTX cells. Pharm. Res. 2001, 18, 1138–1145. [Google Scholar] [CrossRef] [PubMed]
- Duraj-Thatte, A.M.; Praveschotinunt, P.; Nash, T.R.; Ward, F.R.; Joshi, N.S. Modulating bacterial and gut mucosal interactions with engineered biofilm matrix proteins. Sci. Rep. 2018, 8, 3475. [Google Scholar] [CrossRef] [PubMed]
- Skoglund, A.; Backhed, H.K.; Nilsson, C.; Bjorkholm, B.; Normark, S.; Engstrand, L. A changing gastric environment leads to adaptation of lipopolysaccharide variants in Helicobacter pylori populations during colonization. PLoS ONE 2009, 4, e5885. [Google Scholar] [CrossRef] [PubMed]
- Heneghan, M.A.; McCarthy, C.F.; Moran, A.P. Relationship of blood group determinants on Helicobacter pylori lipopolysaccharide with host Lewis phenotype and inflammatory response. Infect. Immun. 2000, 68, 937–941. [Google Scholar] [CrossRef] [PubMed]
- Rasko, D.A.; Keelan, M.; Wilson, T.J.; Taylor, D.E. Lewis antigen expression by Helicobacter pylori. J. Infect. Dis. 2001, 184, 315–321. [Google Scholar] [CrossRef] [PubMed]
- Soutto, M.; Chen, Z.; Katsha, A.M.; Romero-Gallo, J.; Krishna, U.S.; Piazuelo, M.B.; Washington, M.K.; Peek, R.M., Jr.; Belkhiri, A.; El-Rifai, W.M. Trefoil factor 1 expression suppresses Helicobacter pylori-induced inflammation in gastric carcinogenesis. Cancer 2015, 121, 4348–4358. [Google Scholar] [CrossRef]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Clyne, M.; May, F.E.B. The Interaction of Helicobacter pylori with TFF1 and Its Role in Mediating the Tropism of the Bacteria Within the Stomach. Int. J. Mol. Sci. 2019, 20, 4400. https://doi.org/10.3390/ijms20184400
Clyne M, May FEB. The Interaction of Helicobacter pylori with TFF1 and Its Role in Mediating the Tropism of the Bacteria Within the Stomach. International Journal of Molecular Sciences. 2019; 20(18):4400. https://doi.org/10.3390/ijms20184400
Chicago/Turabian StyleClyne, Marguerite, and Felicity E. B. May. 2019. "The Interaction of Helicobacter pylori with TFF1 and Its Role in Mediating the Tropism of the Bacteria Within the Stomach" International Journal of Molecular Sciences 20, no. 18: 4400. https://doi.org/10.3390/ijms20184400
APA StyleClyne, M., & May, F. E. B. (2019). The Interaction of Helicobacter pylori with TFF1 and Its Role in Mediating the Tropism of the Bacteria Within the Stomach. International Journal of Molecular Sciences, 20(18), 4400. https://doi.org/10.3390/ijms20184400