Chromosome Translocations as a Driver of Diversification in Mole Voles Ellobius (Rodentia, Mammalia)

 ,
,  and
and 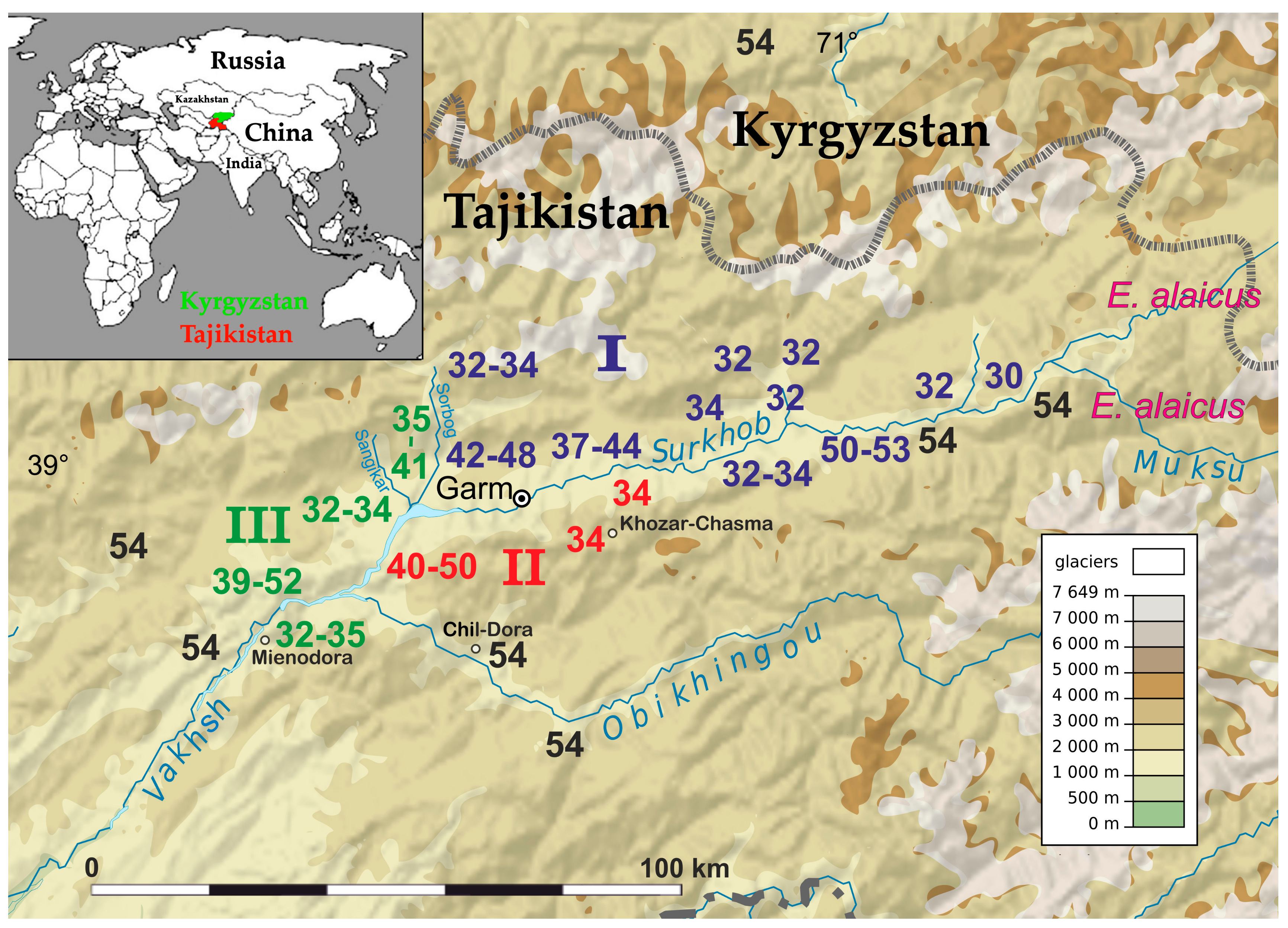{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
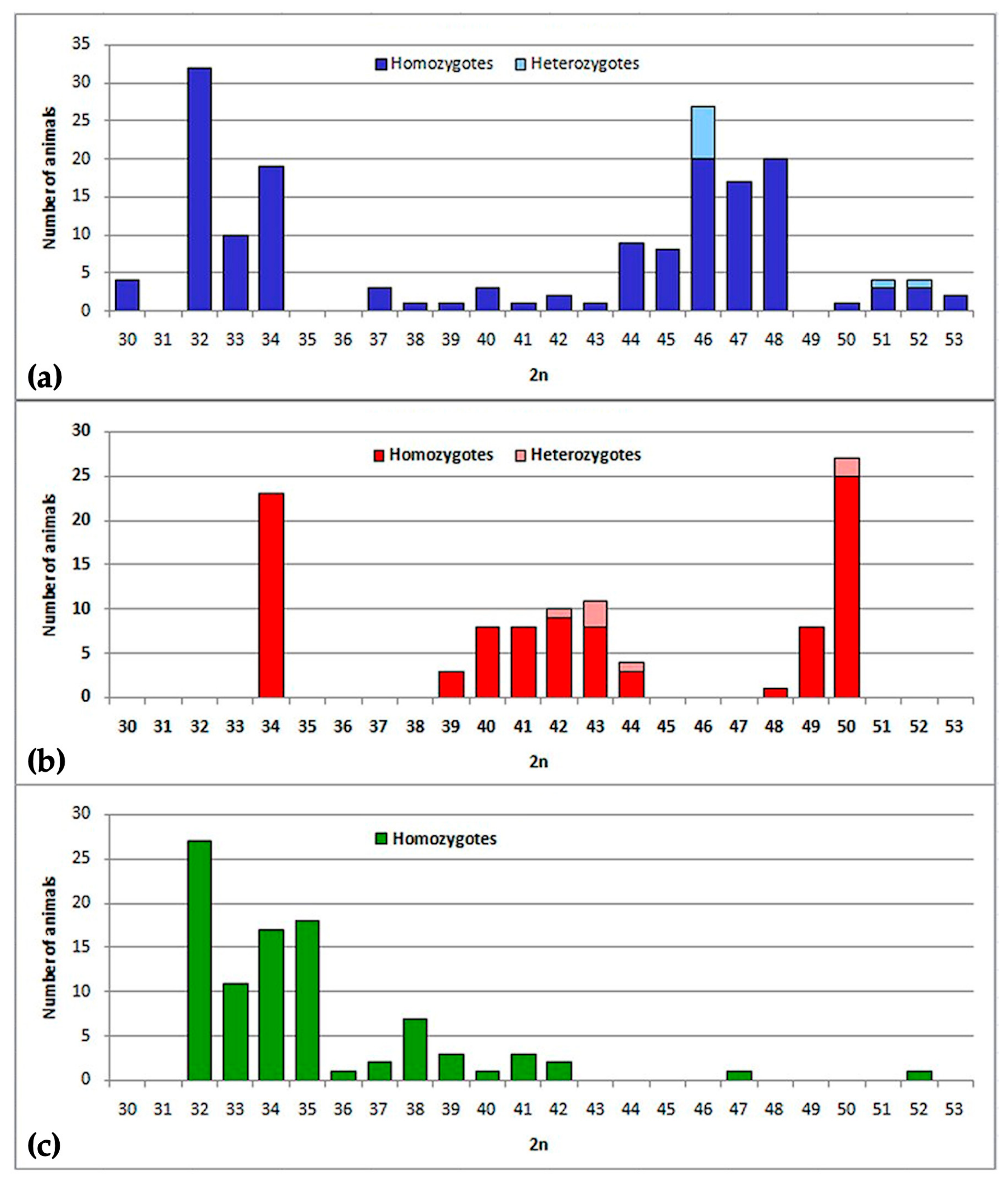
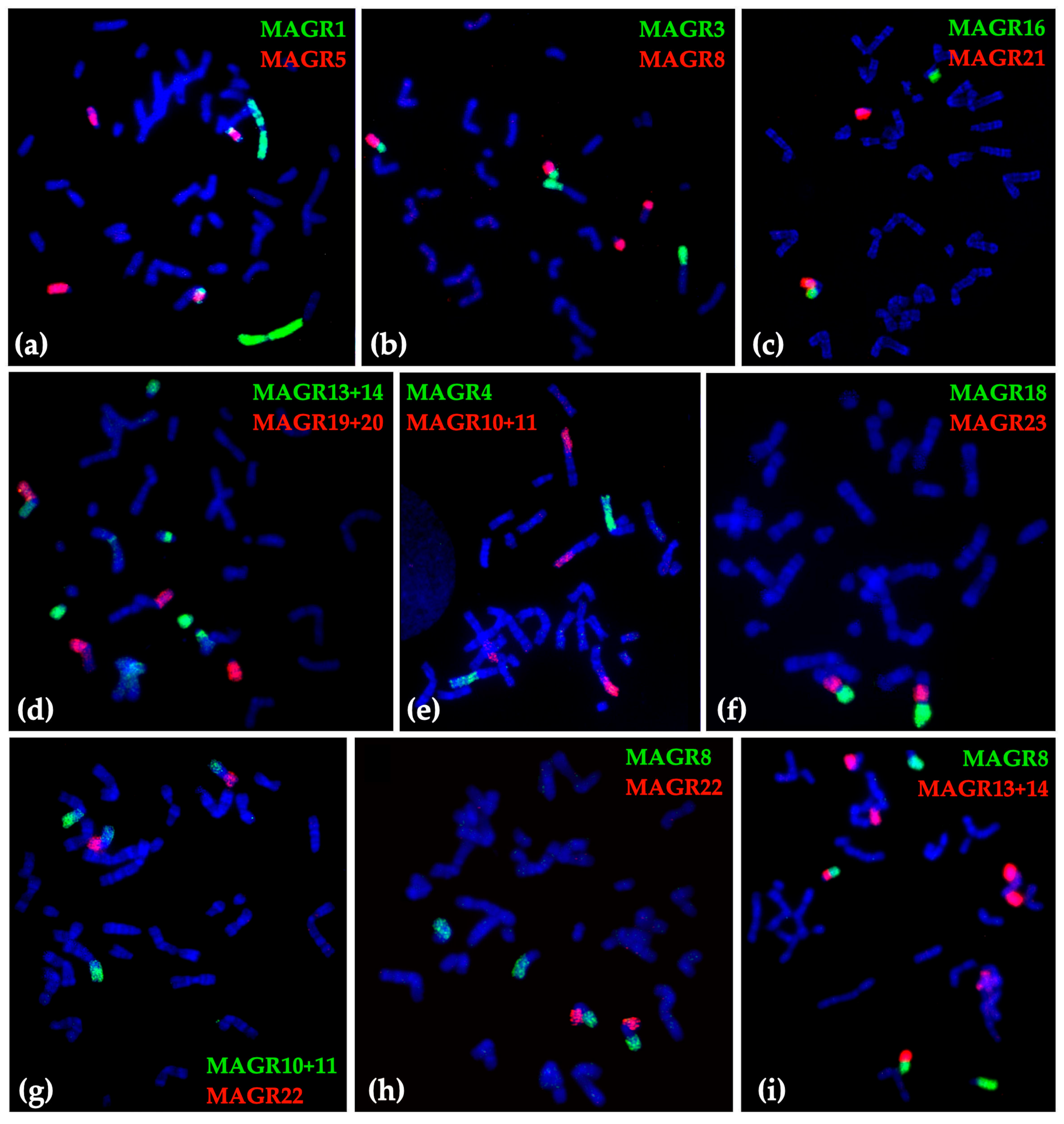
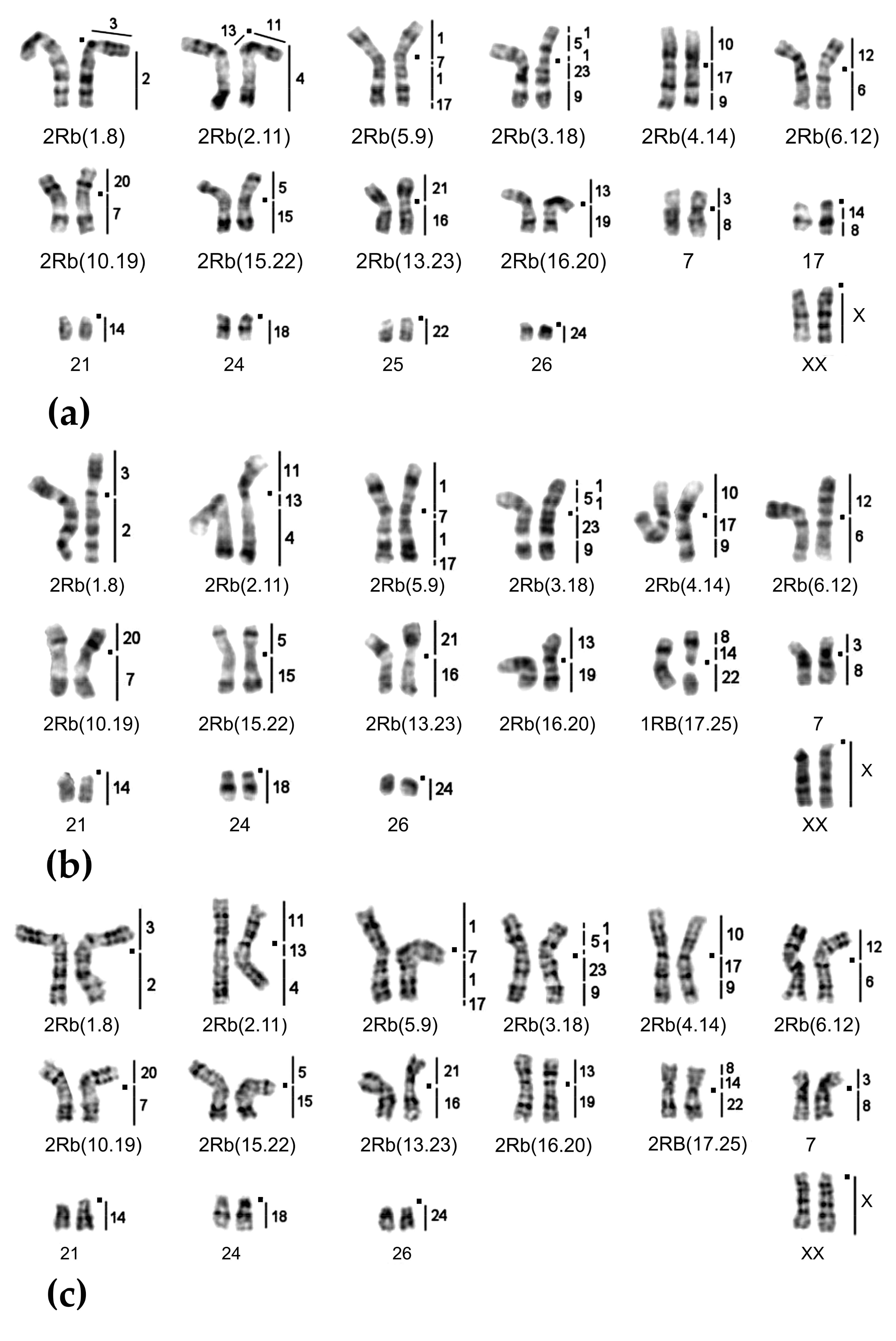
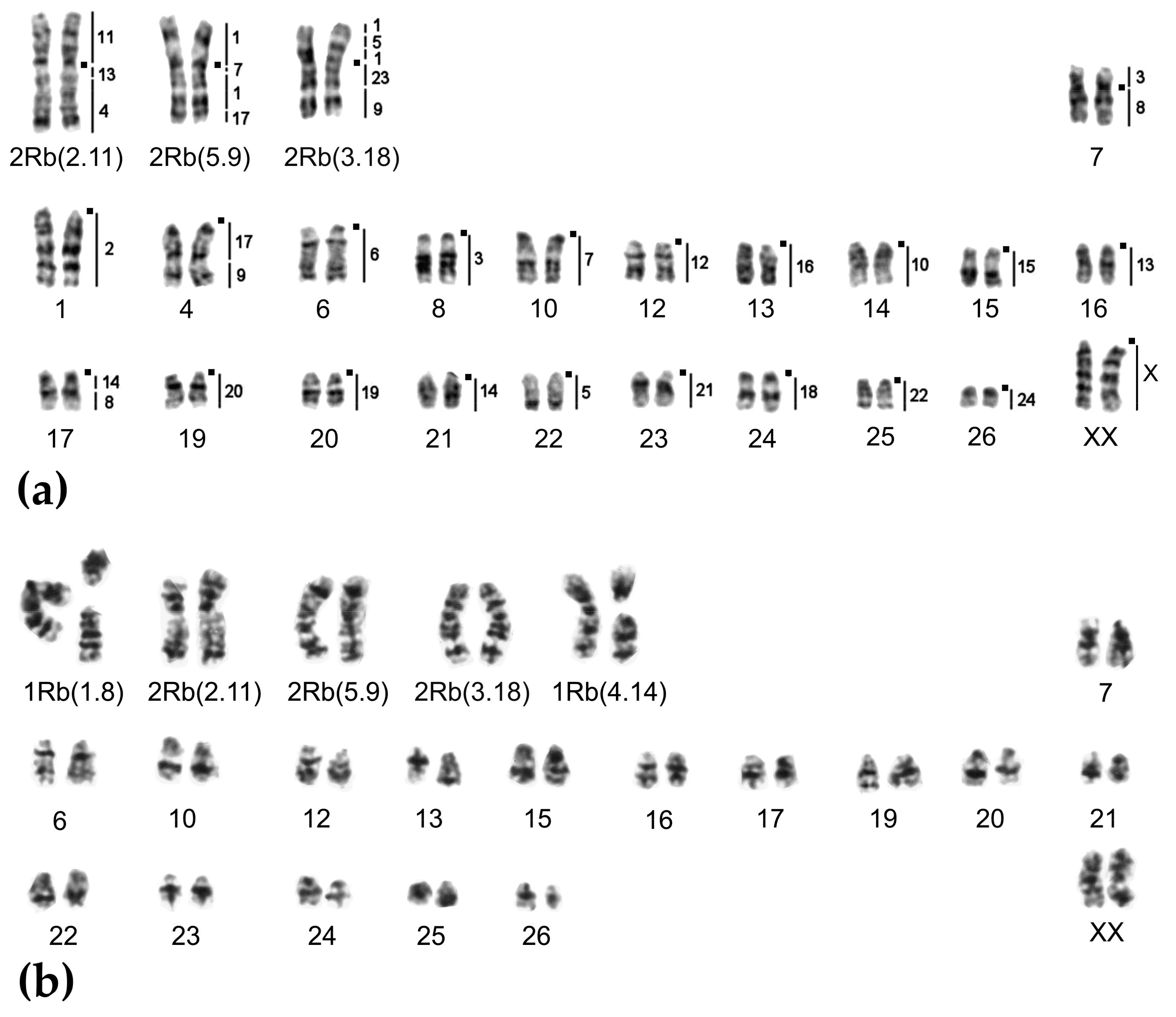
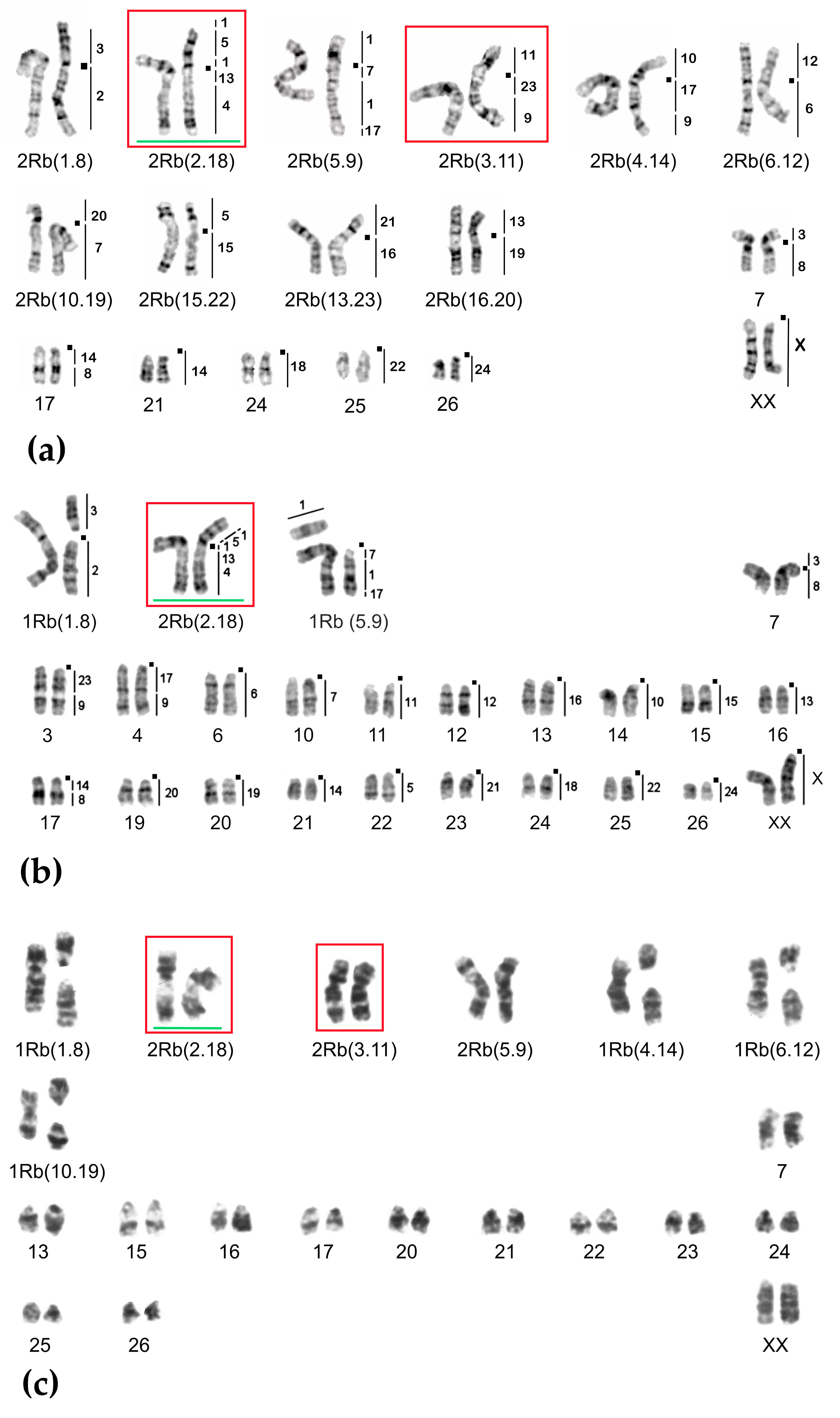
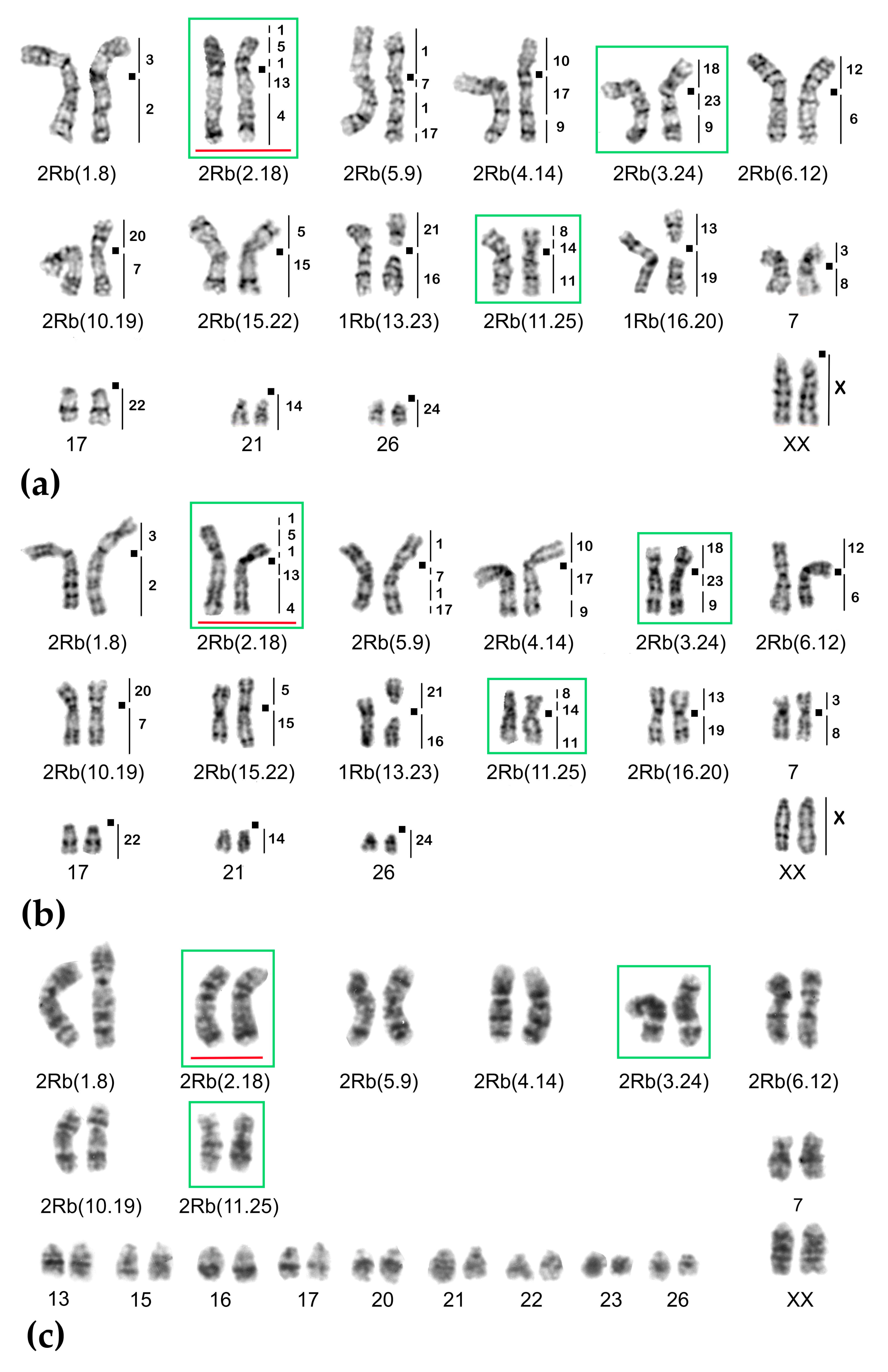
2.1. Chromosome Form I
2.2. Chromosome Form II
2.3. Chromosome Form III
3. Discussion
4. Materials and Methods
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vorontsov, N.N. Species of Palaearctic hamsters (Cricetinae Rodentia) in statu nascendi. Dokl. Acad. Nauk. SSSR Biol. Sci. 1960, 132, 491–493. [Google Scholar]
- White, M.J.D. Modes of Speciation; W.H. Freeman and Co.: New York, NY, USA, 1978. [Google Scholar]
- King, M. Species Evolution: The Role of Chromosome Change; Cambridge University Press: Cambridge, UK, 1993. [Google Scholar]
- Faria, R.; Navarro, A. Chromosomal speciation revisited: Rearranging theory with pieces of evidence. Trends Ecol. Evol. 2010, 25, 660–669. [Google Scholar] [CrossRef] [PubMed]
- Baklolushinskaya, I.Y. Chromosomal rearrangements, genome reorganization, and speciation. Biol. Bull. 2016, 43, 759–775. [Google Scholar] [CrossRef]
- Dobigny, G.; Britton-Davidian, J.; Robinson, T.J. Chromosomal polymorphism in mammals: an evolutionary perspective. Biol.l Rev. 2017, 92, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Berríos, S.; Manieu, C.; López-Fenner, J.; Ayarza, E.; Page, J.; González, M.; Manterola, M.; Fernández-Donoso, R. Robertsonian chromosomes and the nuclear architecture of mouse meiotic prophase spermatocytes. Biol. Res. 2014, 47, 16. [Google Scholar] [CrossRef]
- Farré, M.; Robinson, T.J.; Ruiz-Herrera, A. An Integrative Breakage Model of genome architecture, reshuffling and evolution: The Integrative Breakage Model of genome evolution, a novel multidisciplinary hypothesis for the study of genome plasticity. BioEssays 2015, 37, 479–488. [Google Scholar] [CrossRef]
- Capilla, L.; Sanchez-Guillen, R.A.; Farre, M.; Paytuví-Gallart, A.; Malinverni, R.; Ventura, J.; Larkin, D.M.; Ruiz-Herrera, A. Mammalian comparative genomics reveals genetic and epigenetic features associated with genome reshuffling in Rodentia. Genome Biol. Evol. 2016, 8, 3703–3717. [Google Scholar] [CrossRef] [Green Version]
- Nunes, A.C.; Catalan, J.; Lopez, J.; Da Graça Ramalhinho, M.; Da Luz Mathias, M.; Britton-Davidian, J. Fertility assessment in hybrids between monobrachially homologous Rb races of the house mouse from the island of Madeira: Implications for modes of chromosomal evolution. Heredity 2011, 106, 348–356. [Google Scholar] [CrossRef]
- Garagna, S.; Page, J.; Fernandez-Donoso, R.; Zuccotti, M.; Searle, J.B. The Robertsonian phenomenon in the house mouse: mutation, meiosis and speciation. Chromosoma 2014, 123, 529–544. [Google Scholar] [CrossRef]
- Sotero-Caio, C.; Baker, R.; Volleth, M. Chromosomal evolution in Chiroptera. Genes 2017, 8, 272. [Google Scholar] [CrossRef]
- Deakin, J. Chromosome evolution in marsupials. Genes 2018, 9, 72. [Google Scholar] [CrossRef] [PubMed]
- Matthey, R. La formule chromosomique et le problème de la détermination sexuelle chez Ellobius lutescens (Rodentia-Muridae-Microtinae). Arch. Julius Klaus-Stift Vererb. Forsch. 1953, 28, 65–73. [Google Scholar]
- Lyapunova, E.A.; Vorontsov, N.N. Genetics of Ellobius (Rodentia). I. Karyological characteristics of four Ellobius species. Genetika 1978, 14, 2012–2024. (In Russian) [Google Scholar]
- Vorontsov, N.N.; Lyapunova, E.A.; Borissov, Y.M.; Dovgal, V.E. Variability of sex chromosomes in mammals. Genetica 1980, 52/53, 361–372. [Google Scholar] [CrossRef]
- Matveevsky, S.; Kolomiets, O.; Bogdanov, A.; Hakhverdyan, M.; Bakloushinskaya, I. Chromosomal evolution in mole voles Ellobius (Cricetidae, Rodentia): bizarre sex chromosomes, variable autosomes and meiosis. Genes 2017, 8, 306. [Google Scholar] [CrossRef] [PubMed]
- Topačevsky, V.A.; Rekovets, L.I. New materials for taxonomy and evolution of Ellobius s. str. (Rodentia, Cricetidae). Vestnik Zoologii 1982, 5, 47–54. (In Russian) [Google Scholar]
- Yakimenko, L.V.; Vorontsov, N.N. The morphotypical variety of karyologycally different populations of mole-vole superspecies Ellobius talpinus. In Phenetics of Populations; Yablokov, A.V., Ed.; Nauka: Moscow, Russia, 1982; pp. 276–289, [in Russian]. [Google Scholar]
- Lyapunova, E.A.; Vorontsov, N.N.; Korobitsina, K.V.; Ivanitskaya, E.Y.; Borisov, Y.M.; Yakimenko, L.V.; Dovgal, V.Y. A Robertsonian fan in Ellobius talpinus. Genetica 1980, 52/53, 239–247. [Google Scholar] [CrossRef]
- Bogdanov, Y.F.; Kolomiets, O.L.; Lyapunova, E.A.; Yanina, I.Y.; Mazurova, T.F. Synaptonemal complexes and chromosome chains in the rodent Ellobius talpinus heterozygous for ten Robertsonian translocations. Chromosoma 1986, 94, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Bakloushinskaya, I.Y.; Matveevsky, S.N.; Romanenko, S.A.; Serdukova, N.A.; Kolomiets, O.L.; Spangenberg, V.E.; Lyapunova, E.A.; Graphodatsky, A.S. A comparative analysis of the mole vole sibling species Ellobius tancrei and E. talpinus (Cricetidae, Rodentia) through chromosome painting and examination of synaptonemal complex structures in hybrids. Cytogenet. Genome Res. 2012, 136, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Bogdanov, A.S.; Lebedev, V.S.; Zykov, A.E.; Bakloushinskaya, I.Y. Variability of cytochrome b gene and adjacent part of tRNA-Thr gene of mitochondrial DNA in the northern mole vole Ellobius talpinus (Mammalia, Rodentia). Russ. J. Genet. 2015, 51, 1243–1248. [Google Scholar] [CrossRef]
- Lyapunova, E.A.; Ivnitskii, S.B.; Korablev, V.P.; Yanina, I.Y. Complete Robertsonian fan of the chromosomal forms in the mole-vole superspecies Ellobius talpinus. Proc. USSR Acad. Sci. 1984, 274, 1209–1213. (In Russian) [Google Scholar]
- Lyapunova, E.A.; Bakloushinskaya, I.Y.; Saidov, A.S.; Saidov, K.K. Dynamics of chromosome variation in mole voles Ellobius tancrei (Mammalia, Rodentia) in Pamiro-Alay in the period from 1982 to 2008. Russ. J. Genet. 2010, 45, 566–571. [Google Scholar] [CrossRef]
- Bakloushinskaya, I.; Romanenko, S.; Serdukova, N.; Graphodatsky, A.; Lyapunova, E. A new form of the mole vole Ellobius tancrei Blasius, 1884 (Mammalia, Rodentia) with the lowest chromosome number. Comp. Cytogenet. 2013, 7, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Romanenko, S.A.; Sitnikova, N.A.; Serdukova, N.A.; Perelman, P.L.; Rubtsova, N.V.; Bakloushinskaya, I.Y.; Lyapunova, E.A.; Just, W.; Ferguson-Smith, M.A.; Yang, F.; et al. Chromosomal evolution of Arvicolinae (Cricetidae, Rodentia). II. The genome homology of two mole voles (genus Ellobius), the field vole and golden hamster revealed by comparative chromosome painting. Chromosome Res. 2007, 15, 891–897. [Google Scholar] [CrossRef] [PubMed]
- Bakloushinskya, I.Y.; Romanenko, S.A.; Graphodatsky, A.S.; Matveevsky, S.N.; Lyapunova, E.A.; Kolomiets, O.L. The role of chromosome rearrangements in the evolution of mole voles of the genus Ellobius (Rodentia, Mammalia). Russ. J. Genet. 2010, 46, 1143–1145. [Google Scholar] [CrossRef]
- Matveevsky, S.; Bakloushinskaya, I.; Tambovtseva, V.; Romanenko, S.; Kolomiets, O. Analysis of meiotic chromosome structure and behavior in Robertsonian heterozygotes of Ellobius tancrei (Rodentia, Cricetidae): A case of monobrachial homology. Comp. Cytogenet. 2015, 9, 691–706. [Google Scholar] [CrossRef]
- Bakloushinskaya, I.; Lyapunova, E.A.; Saidov, A.S.; Romanenko, S.A.; O’Brien, P.C.; Serdyukova, N.A.; Ferguson-Smith, M.A.; Matveevsky, S.; Bogdanov, A.S. Rapid chromosomal evolution in enigmatic mammal with XX in both sexes, the Alay mole vole Ellobius alaicus Vorontsov et al., 1969 (Mammalia, Rodentia). Comp. Cytogenet. 2019, 13, 147–177. [Google Scholar] [CrossRef]
- Winking, H. Some aspects of Robertsonian karyotype variation in European wild mice. Curr. Topics. Microbiol. Immunol. 1986, 127, 68–74. [Google Scholar]
- Catalan, J.; Auffray, J.C.; Pellestor, F.; Britton-Davidian, J. Spontaneous occurrence of a Robertsonian fusion involving chromosome 19 by single whole-arm reciprocal translocation (WART) in wild-derived house mice. Chromosome Res. 2000, 8, 593–601. [Google Scholar] [CrossRef]
- Searle, J.B.; Polly, P.D.; Zima, J. (Eds.) Shrews, Chromosomes and Speciation. Cambridge University Press: Cambridge, UK, 2019; p. 488. [Google Scholar]
- Hauffe, H.C.; Piálek, J. Evolution of the chromosomal races of Mus musculus domesticus in the Rhaetian Alps: the roles of whole-arm reciprocal translocation and zonal raciation. Biol. J. Linn. Soc. 1997, 62, 255–278. [Google Scholar] [CrossRef]
- Britton-Davidian, J.; Catalan, J.; Ramalhinho, M.-G.; Ganem, G.; Auffray, J.-C.; Capela, R.; Biscoito, M.; Searle, J.B.; Da Luz Mathiaset, M. Rapid chromosomal evolution in island mice. Nature 2000, 403, 158. [Google Scholar] [CrossRef] [PubMed]
- Veyrunes, F.; Catalan, J.; Tatard, C.; Cellier-Holzem, E.; Watson, J.; Chevret, P.; Robinson, T.J.; Britton-Davidian, J. Mitochondrial and chromosomal insights into karyotypic evolution of the pygmy mouse, Mus minutoides, in South Africa. Chromosome Res. 2010, 18, 563–574. [Google Scholar] [CrossRef] [PubMed]
- Castiglia, R.; Capanna, E.; Bezerra, A.M.; Bizzoco, D.; Zambigli, E.; Solano, E. New metacentric populations and phylogenetic hypotheses involving Whole-Arm Reciprocal Translocation in Mus musculus domesticus from Sicily, Southern Italy. Cytogenet. Genome Res. 2015, 146, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Tambovtseva, V.G.; Matveevsky, S.N.; Kashintsova, A.A.; Tretiakov, A.V.; Kolomiets, O.L.; Bakloushinskaya, I.Y. A meiotic mystery in experimental hybrids of the eastern mole vole (Ellobius tancrei, Mammalia, Rodentia). Vavilov J. Genetics Breed. 2019, 23, 239–243. [Google Scholar]
- Vorontsov, N.N.; Lyapunova, E.A. Explosive chromosomal speciation in seismic active regions. Chromosomes Today 1984, 8, 279–294. [Google Scholar]
- Kufner, S.K.; Schurr, B.; Ratschbacher, L.; Murodkulov, S.; Abdulhameed, S.; Ischuk, A.; Metzger, S.; Kakar, N. Seismotectonics of the Tajik basin and surrounding mountain ranges. Tectonics 2018, 37, 2404–2424. [Google Scholar] [CrossRef]
- Bush, G.L.; Case, S.M.; Wilson, A.S.; Patton, J.L. Rapid speciation and chromosomal evolution in mammals. Proc. Natl. Acad. Sci. USA 1977, 74, 3942–3946. [Google Scholar] [CrossRef]
- Baker, R.J.; Bickham, J.W. Speciation by monobrachial centric fusions. Proc. Natl. Acad. Sci. USA 1986, 83, 8245–8248. [Google Scholar] [CrossRef] [Green Version]
- Stanyon, R.; Galleni, L. A rapid fibroblast culture technique for high resolution karyotypes. Italian J. Zool. 1991, 58, 81–83. [Google Scholar] [CrossRef] [Green Version]
- Romanenko, S.A.; Biltueva, L.S.; Serdyukova, N.A.; Kulemzina, A.I.; Beklemisheva, V.R.; Gladkikh, O.L.; Lemskaya, N.A.; Interesova, E.A.; Korentovich, M.A.; Vorobieva, N.V.; et al. Segmental paleotetraploidy revealed in sterlet (Acipenser ruthenus) genome by chromosome painting. Mol. Cytogenet. 2015, 8, 90. [Google Scholar] [CrossRef]
- Sitnikova, N.A.; Romanenko, S.A.; O’Brien, P.C.; Perelman, P.L.; Fu, B.; Rubtsova, N.V.; Serdukova, N.A.; Golenishchev, F.N.; Trifonov, V.A.; Ferguson-Smith, M.A.; et al. Chromosomal evolution of Arvicolinae (Cricetidae, Rodentia). I. The genome homology of tundra vole, field vole, mouse and golden hamster revealed by comparative chromosome painting. Chromosome Res. 2007, 15, 447–456. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; O’Brien, P.C.M.; Milne, B.S.; Graphodatsky, A.S.; Solanky, N.; Trifonov, V.; Rens, W.; Sargan, D.; Ferguson-Smith, M.A. A complete comparative chromosome map for the dog, red fox, and human and its integration with canine genetic maps. Genomics 1999, 62, 189–202. [Google Scholar] [CrossRef] [PubMed]
- Graphodatsky, A.S.; Yang, F.; O’Brien, P.C.M.; Serdukova, N.; Milne, B.S.; Trifonov, V.; Ferguson-Smith, M.A. A comparative chromosome map of the Arctic fox, red fox and dog defined by chromosome painting and high resolution G-banding. Chromosome Res. 2000, 8, 253–263. [Google Scholar] [CrossRef] [PubMed]
- Seabright, M. A rapid banding technique for human chromosomes. Lancet 1971, 2, 971–972. [Google Scholar] [CrossRef]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Romanenko, S.A.; Lyapunova, E.A.; Saidov, A.S.; O’Brien, P.C.M.; Serdyukova, N.A.; Ferguson-Smith, M.A.; Graphodatsky, A.S.; Bakloushinskaya, I. Chromosome Translocations as a Driver of Diversification in Mole Voles Ellobius (Rodentia, Mammalia). Int. J. Mol. Sci. 2019, 20, 4466. https://doi.org/10.3390/ijms20184466
Romanenko SA, Lyapunova EA, Saidov AS, O’Brien PCM, Serdyukova NA, Ferguson-Smith MA, Graphodatsky AS, Bakloushinskaya I. Chromosome Translocations as a Driver of Diversification in Mole Voles Ellobius (Rodentia, Mammalia). International Journal of Molecular Sciences. 2019; 20(18):4466. https://doi.org/10.3390/ijms20184466
Chicago/Turabian StyleRomanenko, Svetlana A., Elena A. Lyapunova, Abdusattor S. Saidov, Patricia C.M. O’Brien, Natalia A. Serdyukova, Malcolm A. Ferguson-Smith, Alexander S. Graphodatsky, and Irina Bakloushinskaya. 2019. "Chromosome Translocations as a Driver of Diversification in Mole Voles Ellobius (Rodentia, Mammalia)" International Journal of Molecular Sciences 20, no. 18: 4466. https://doi.org/10.3390/ijms20184466
APA StyleRomanenko, S. A., Lyapunova, E. A., Saidov, A. S., O’Brien, P. C. M., Serdyukova, N. A., Ferguson-Smith, M. A., Graphodatsky, A. S., & Bakloushinskaya, I. (2019). Chromosome Translocations as a Driver of Diversification in Mole Voles Ellobius (Rodentia, Mammalia). International Journal of Molecular Sciences, 20(18), 4466. https://doi.org/10.3390/ijms20184466