Overview of the Anticancer Profile of Avenanthramides from Oat




Abstract
:1. Introduction
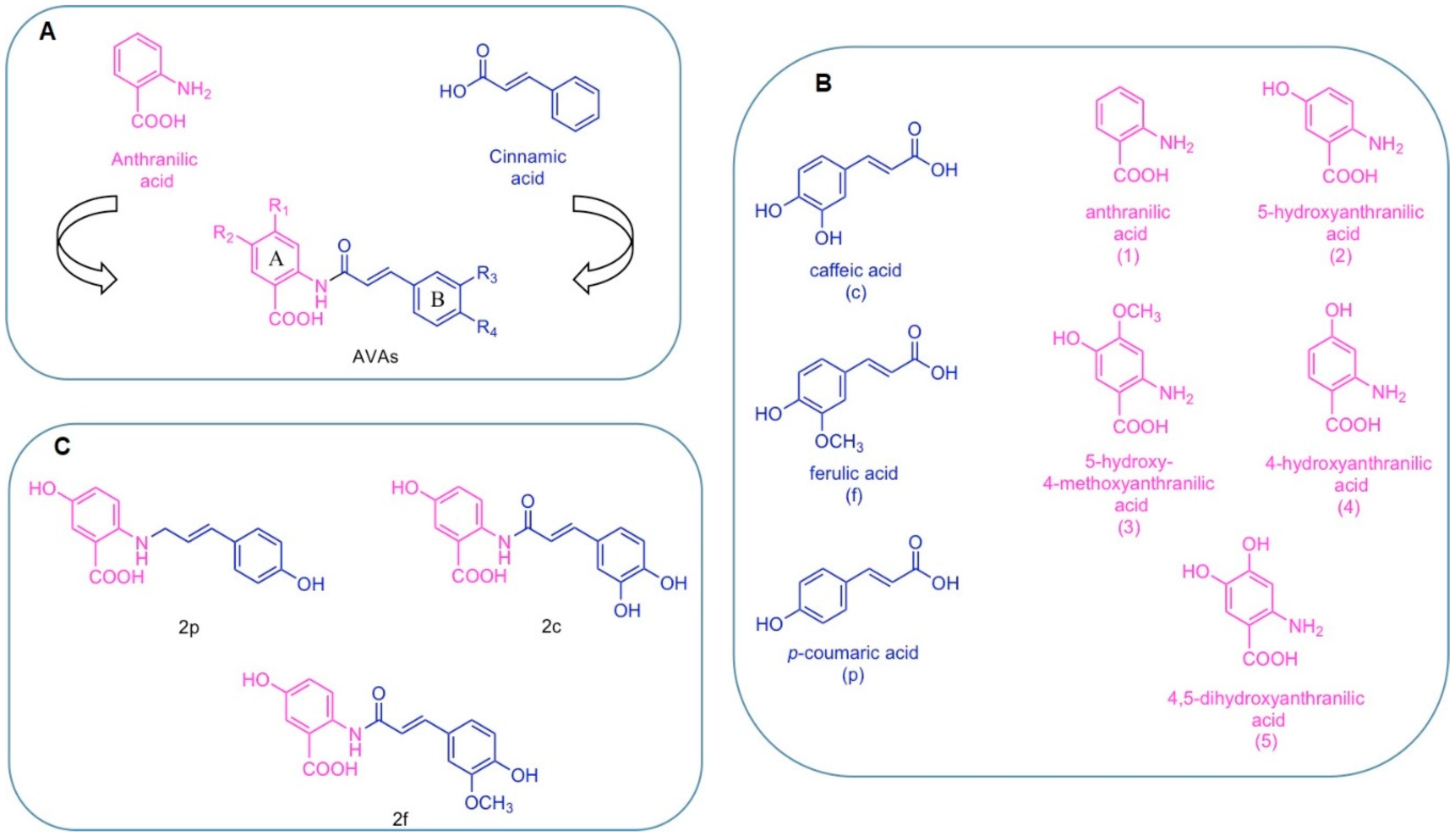
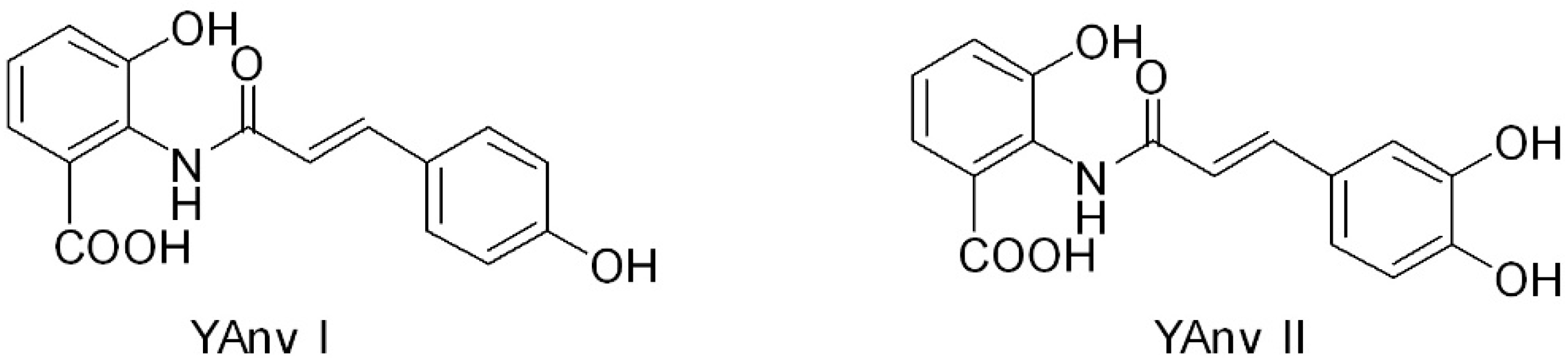
2. Chemical Structure of AVAs and Their Derivatives
3. Pharmacokinetics
4. Epidemiological Studies
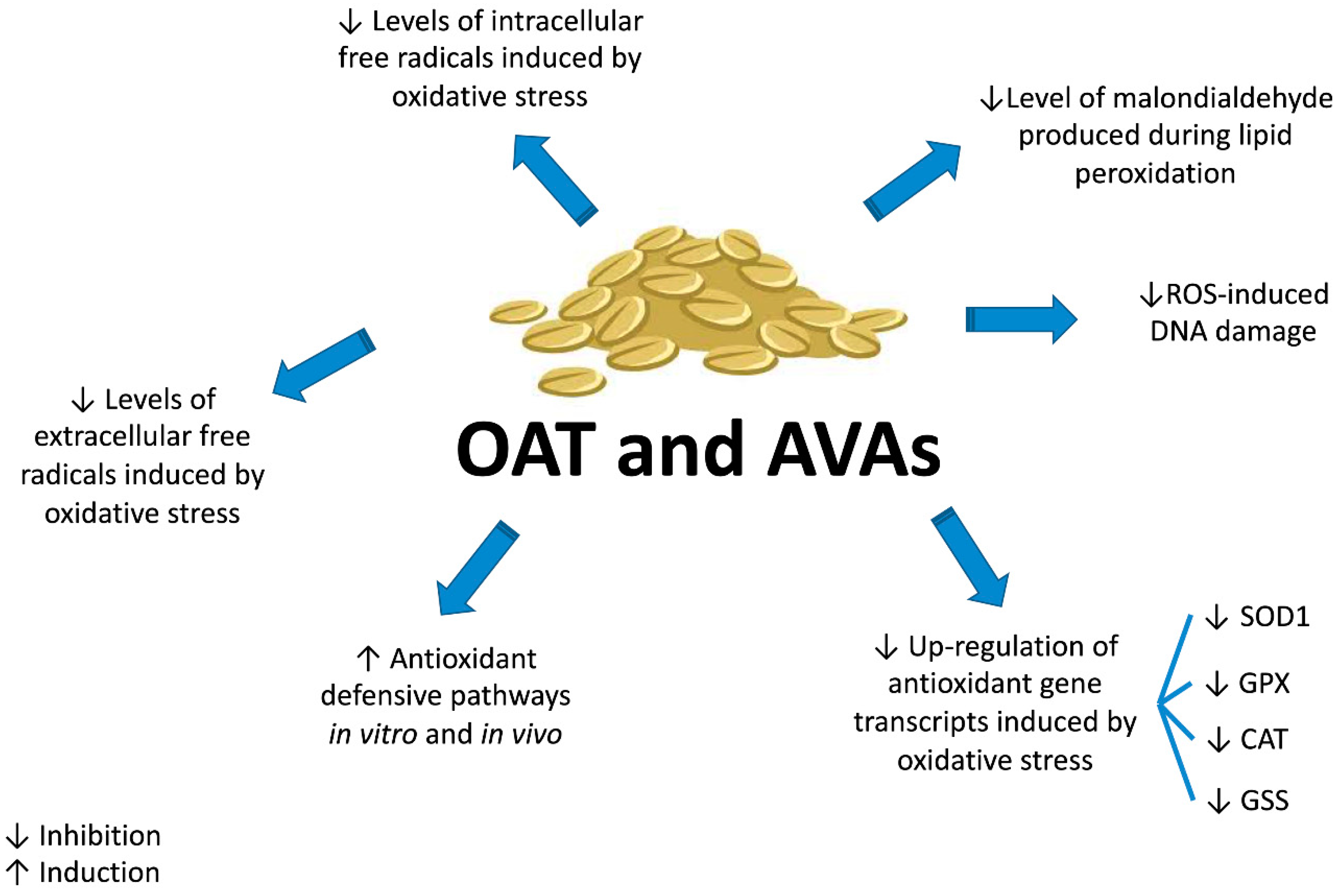
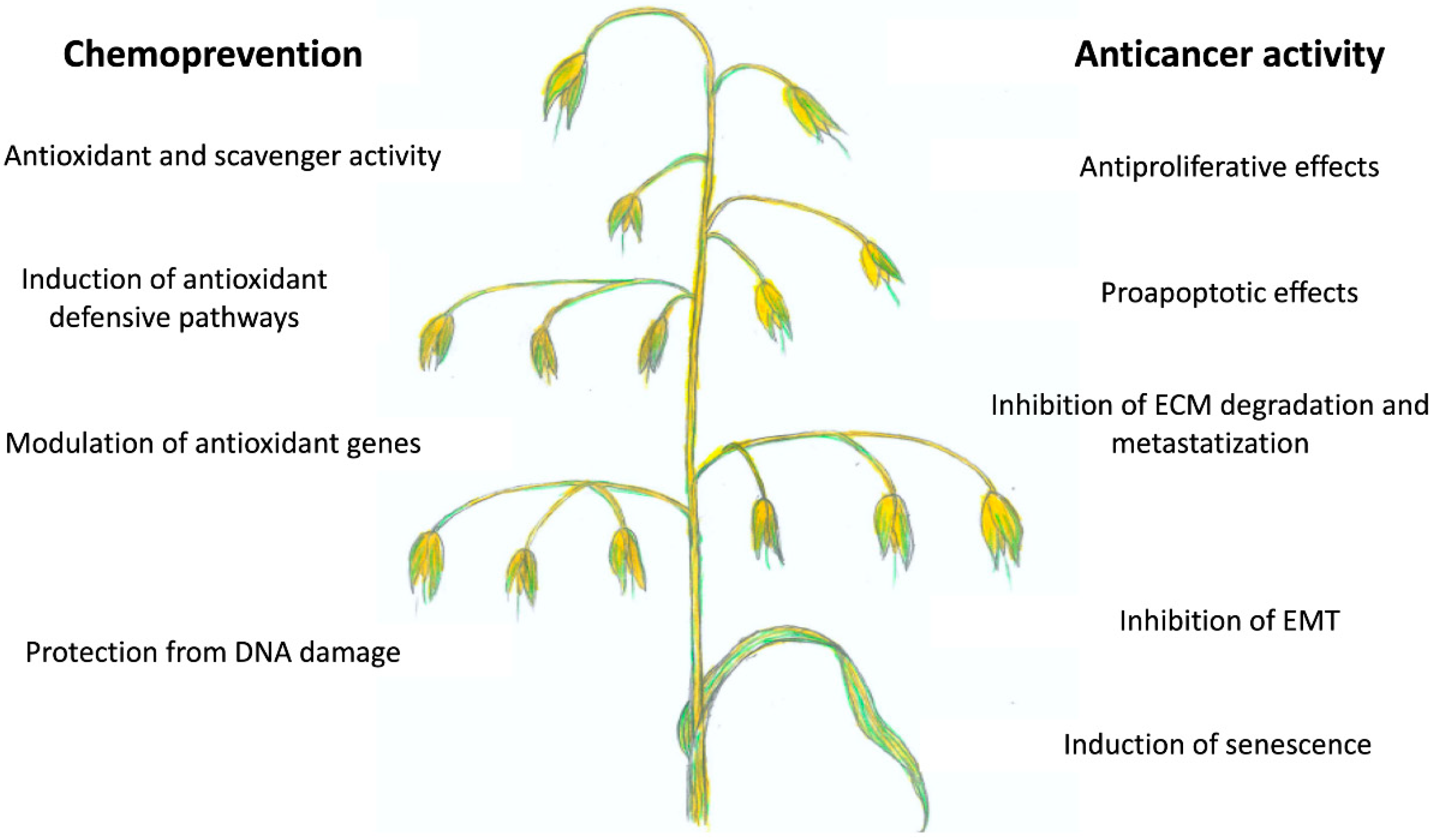
5. Cancer Chemopreventive Activity of AVAs
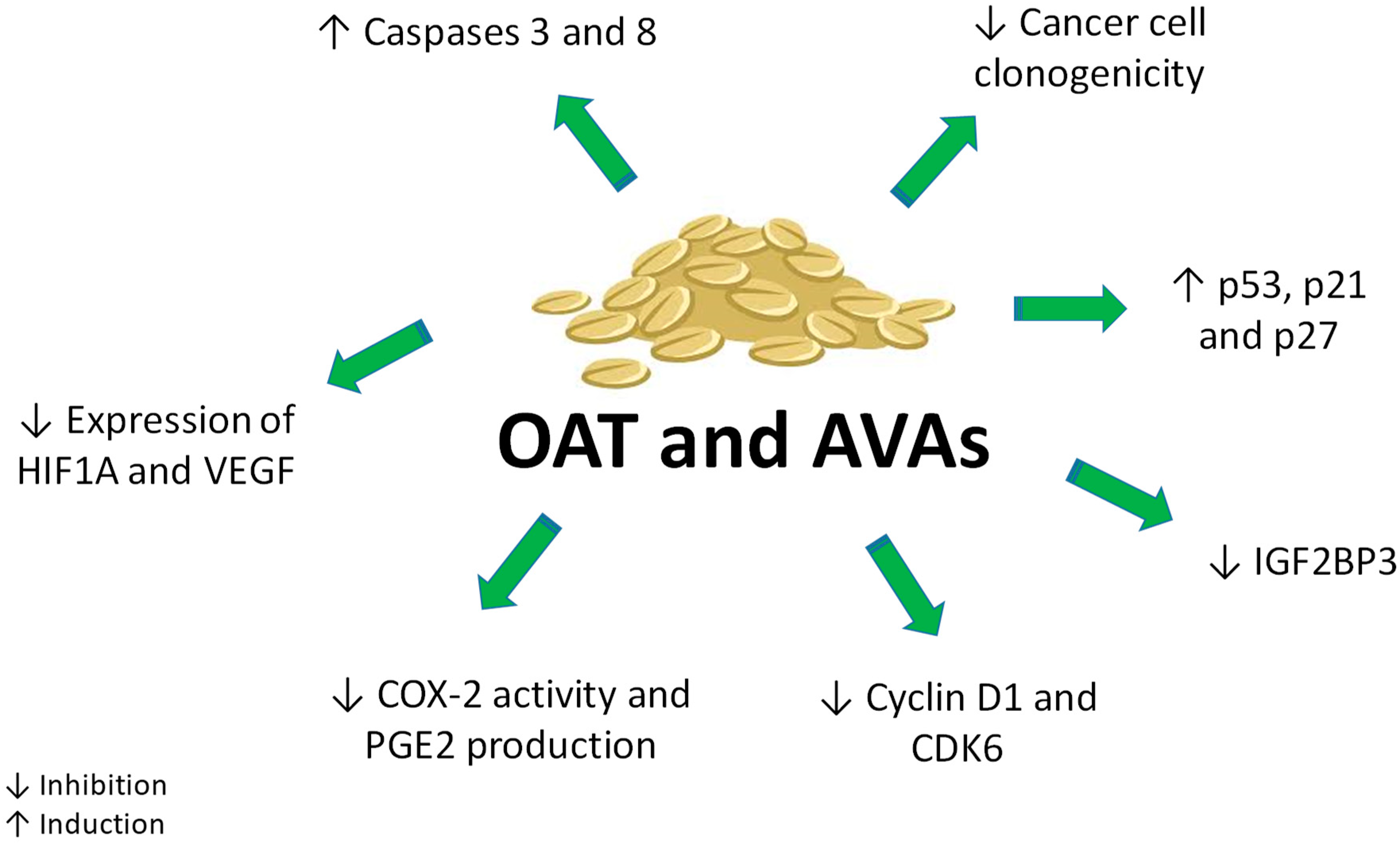
6. Anticancer Effects of AVAs
6.1. In Vitro Studies
6.1.1. Antiproliferative Effects and Induction of Apoptosis
6.1.2. Induction of Senescence
6.1.3. Inhibition of Epithelial-Mesenchymal Transition and Metastatization
6.2. In Vivo Studies
7. Selectivity of AVAs towards Cancer Cells
8. Conclusions
Supplementary Materials
Funding
Conflicts of Interest
Abbreviations
| AEM | AVA-enriched mixture |
| AFP | alpha-fetoprotein |
| AOM/DSS | azoxymethane/dextran sodium sulfate |
| AP-1 | activator protein-1 |
| ARE | AVAs-rich extract |
| AVA 2c-O-Glc | AVA 2c glycosylated |
| AVAs | Avenanthramides |
| AvExO | AVAs enriched extract |
| b.w. | body weight |
| BHT | butylated hydroxytoluene |
| CAA | cellular antioxidant activity |
| CAT | Catalase |
| CDK6 | cyclin-dependent kinase 6 |
| CEA | carcinoembryonic antigen |
| COX-2 | cyclooxygenase-2 |
| DCFH-DA | dichloro-dihydro-fluorescein diacetate |
| DH 2c | dihydroavenanthramide 2c |
| DH 2f | dihydroavenanthramide 2f |
| DHAvD | dihydroavenanthramide D |
| DPPH | 2,2-dyphenyl-1-picryldrazyl |
| ECM | extracellular matrix |
| EMT | epithelial-mesenchymal transition |
| FFQs | food frequency questionnaires |
| FRAP | ferric reducing antioxidant power |
| GPX | glutathione peroxidase 1 |
| GSH | Glutathione |
| GSS | glutathione synthetase |
| H-AVA | high amount of AVAs |
| HMOX1 | heme oxygenase 1 |
| HO-1 | heme oxygenase 1 |
| HORAC | hydroxyl radical absorption capacity |
| IGF2BP3 | insulin like growth factor 2 mRNA binding protein 3 |
| Keap1 | Kelch-like ECH associating protein 1 |
| L-AVA | low amount of AVAs |
| LEF-1 | lymphoid-enhancing factor-1 |
| MAPK | mitogen-activated protein kinase |
| MDA | Malondialdehyde |
| MEK1 | mitogen-activated protein kinase kinase |
| MiRNA | microRNA |
| MMP | matrix metalloproteinase |
| MTS | (3-[4,5-dimethylthiazol-2-yl]-5-[3-carboxymethoxyphenyl]-2-[4-sulfophenyl]-2H-tetrazolium) |
| MTT | (3-[4,5-dimethylthiazol-2-yl]-2,5 diphenyl tetrazolium bromide) |
| NAC | N-acetylcysteine |
| NF-κB | nuclear factor-kappa B |
| NORAC | peroxynitrite absorption capacity |
| Nrf2 | nuclear factor erythroid related factor 2 |
| ORAC | oxygen radical absorption capacity |
| PCNA | proliferating cell nuclear antigen |
| PGE2 | prostaglandin-E2 |
| PI3K | phosphoinositide 3-kinase |
| Pirh2 | E3 ubiquitin ligase p53-induced RING-H2 |
| ROS | reactive oxygen species |
| SOAC | singlet oxygen absorption capacity |
| SOD | superoxide dismutase |
| SORAC | superoxide anion absorption capacity |
| TNF | tumor necrosis factor |
| TPA | 12-O-tetradecanoylphorbol-13-acetate |
| Trolox | 6-hydroxy-2,5,7,8-tetramethylchromane-2-carboxylic acid |
| UPLC – MS | ultra-high-performance liquid chromatography – mass spectrometry |
| YAvn I | N-(4′-hydroxycinnamoyl)-3-hydroxyanthranilic acid |
| YAvn II | N-(3′-4′-dihydroxycinnamoyl)-3-hydroxyanthranilic acid |
References
- Fardet, A. New hypotheses for the health-protective mechanisms of whole-grain cereals: What is beyond fibre? Nutr. Res. Rev. 2010, 23, 65–134. [Google Scholar] [CrossRef] [PubMed]
- Aune, D.; Keum, N.; Giovannucci, E.; Fadnes, L.T.; Boffetta, P.; Greenwood, D.C.; Tonstad, S.; Vatten, L.J.; Riboli, E.; Norat, T. Whole grain consumption and risk of cardiovascular disease, cancer, and all cause and cause specific mortality: Systematic review and dose-response meta-analysis of prospective studies. BMJ 2016, 353, i2716. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Cancer. Available online: https://www.who.int/news-room/fact-sheets/detail/cancer (accessed on 26 August 2019).
- Pritchard, J.R.; Lauffenburger, D.A.; Hemann, M.T. Understanding resistance to combination chemotherapy. Drug Resist. Updates 2012, 15, 249–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fimognari, C.; Ferruzzi, L.; Turrini, E.; Carulli, G.; Lenzi, M.; Hrelia, P.; Cantelli-Forti, G. Metabolic and toxicological considerations of botanicals in anticancer therapy. Expert Opin. Drug Metab. Toxicol. 2012, 8, 819–832. [Google Scholar] [CrossRef] [PubMed]
- Panda, A.K.; Chakraborty, D.; Sarkar, I.; Khan, T.; Sa, G. New insights into therapeutic activity and anticancer properties of curcumin. J. Exp. Pharmacol. 2017, 9, 31–45. [Google Scholar] [CrossRef]
- Turrini, E.; Catanzaro, E.; Ferruzzi, L.; Guerrini, A.; Tacchini, M.; Sacchetti, G.; Paganetto, G.; Maffei, F.; Pellicioni, V.; Poli, F.; et al. Hemidesmus indicus induces apoptosis via proteasome inhibition and generation of reactive oxygen species. Sci. Rep. 2019, 9, 7199. [Google Scholar] [CrossRef]
- Cragg, G.M.; Pezzuto, J.M. Natural Products as a Vital Source for the Discovery of Cancer Chemotherapeutic and Chemopreventive Agents. Med. Princ. Pract. 2016, 25 (Suppl. 2), 41–59. [Google Scholar] [CrossRef]
- Rasane, P.; Jha, A.; Sabikhi, L.; Kumar, A.; Unnikrishnan, V.S. Nutritional advantages of oats and opportunities for its processing as value added foods—A review. J. Food Sci. Technol. 2015, 52, 662–675. [Google Scholar] [CrossRef]
- Perrelli, A.; Goitre, L.; Salzano, A.M.; Moglia, A.; Scaloni, A.; Retta, S.F. Biological Activities, Health Benefits, and Therapeutic Properties of Avenanthramides: From Skin Protection to Prevention and Treatment of Cerebrovascular Diseases. Oxid. Med. Cell Longev. 2018, 2018, 6015351. [Google Scholar] [CrossRef]
- Johnsen, N.F.; Frederiksen, K.; Christensen, J.; Skeie, G.; Lund, E.; Landberg, R.; Johansson, I.; Nilsson, L.M.; Halkjaer, J.; Olsen, A.; et al. Whole-grain products and whole-grain types are associated with lower all-cause and cause-specific mortality in the Scandinavian HELGA cohort. Br. J. Nutr. 2015, 114, 608–623. [Google Scholar] [CrossRef] [Green Version]
- Collins, F.W. Oat phenolics: Avenanthramides, novel substituted N-cinnamoylanthranilate alkaloids from oat groats and hulls. J. Agric. Food Chem. 1989, 37, 60–66. [Google Scholar] [CrossRef]
- Okazaki, Y.; Isobe, T.; Iwata, Y.; Matsukawa, T.; Matsuda, F.; Miyagawa, H.; Ishihara, A.; Nishioka, T.; Iwamura, H. Metabolism of avenanthramide phytoalexins in oats. Plant J. 2004, 39, 560–572. [Google Scholar] [CrossRef] [PubMed]
- Meydani, M. Potential health benefits of avenanthramides of oats. Nutr. Rev. 2009, 67, 731–735. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, V.; Singh, A.; Ashraf, M.T. Avenanthramides of oats: Medicinal importance and future perspectives. Pharmacogn. Rev. 2018, 12, 66. [Google Scholar] [CrossRef]
- Wu, W.; Tang, Y.; Yang, J.; Idehen, E.; Sang, S. Avenanthramide Aglycones and Glucosides in Oat Bran: Chemical Profile, Levels in Commercial Oat Products, and Cytotoxicity to Human Colon Cancer Cells. J. Agric. Food Chem. 2018, 66, 8005–8014. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Ou, B.; Wise, M.L.; Chu, Y. In vitro total antioxidant capacity and anti-inflammatory activity of three common oat-derived avenanthramides. Food Chem. 2014, 160, 338–345. [Google Scholar] [CrossRef] [PubMed]
- Finetti, F.; Moglia, A.; Schiavo, I.; Donnini, S.; Berta, G.N.; Di Scipio, F.; Perrelli, A.; Fornelli, C.; Trabalzini, L.; Retta, S.F. Yeast-Derived Recombinant Avenanthramides Inhibit Proliferation, Migration and Epithelial Mesenchymal Transition of Colon Cancer Cells. Nutrients 2018, 10, 1159. [Google Scholar] [CrossRef] [PubMed]
- Moglia, A.; Goitre, L.; Gianoglio, S.; Baldini, E.; Trapani, E.; Genre, A.; Scattina, A.; Dondo, G.; Trabalzini, L.; Beekwilder, J.; et al. Evaluation of the bioactive properties of avenanthramide analogs produced in recombinant yeast. Biofactors 2015, 41, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Sim, G.Y.; Kang, H.; Yeo, W.S.; Kim, B.G.; Ahn, J.H. Synthesis of avenanthramides using engineered Escherichia coli. Microb. Cell Fact. 2018, 17, 46. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.M.; Noh, E.M.; Kwon, K.B.; Hwang, B.M.; Hwang, J.K.; You, Y.O.; Kim, M.S.; Lee, W.; Lee, J.H.; Kim, H.J.; et al. Dihydroavenanthramide D prevents UV-irradiated generation of reactive oxygen species and expression of matrix metalloproteinase-1 and -3 in human dermal fibroblasts. Exp. Dermatol. 2013, 22, 759–761. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.R.; Noh, E.M.; Oh, H.J.; Hur, H.; Kim, J.M.; Han, J.H.; Hwang, J.K.; Park, B.H.; Park, J.W.; Youn, H.J.; et al. Dihydroavenanthramide D inhibits human breast cancer cell invasion through suppression of MMP-9 expression. Biochem. Biophys. Res. Commun. 2011, 405, 552–557. [Google Scholar] [CrossRef] [PubMed]
- Lv, N.; Song, M.Y.; Lee, Y.R.; Choi, H.N.; Kwon, K.B.; Park, J.W.; Park, B.H. Dihydroavenanthramide D protects pancreatic beta-cells from cytokine and streptozotocin toxicity. Biochem. Biophys. Res. Commun. 2009, 387, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Nie, L.; Wu, D.; Wise, M.L.; Collins, F.W.; Meydani, S.N.; Meydani, M. Avenanthramides inhibit proliferation of human colon cancer cell lines in vitro. Nutr. Cancer 2010, 62, 1007–1016. [Google Scholar] [CrossRef] [PubMed]
- Fagerlund, A.; Sunnerheim, K.; Dimberg, L.H. Radical-scavenging and antioxidant activity of avenanthramides. Food Chem. 2009, 113, 550–556. [Google Scholar] [CrossRef]
- Bohn, T.; McDougall, G.J.; Alegria, A.; Alminger, M.; Arrigoni, E.; Aura, A.M.; Brito, C.; Cilla, A.; El, S.N.; Karakaya, S.; et al. Mind the gap-deficits in our knowledge of aspects impacting the bioavailability of phytochemicals and their metabolites--a position paper focusing on carotenoids and polyphenols. Mol. Nutr. Food Res. 2015, 59, 1307–1323. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Y.; Milbury, P.E.; Kwak, H.K.; Collins, F.W.; Samuel, P.; Blumberg, J.B. Avenanthramides and phenolic acids from oats are bioavailable and act synergistically with vitamin C to enhance hamster and human LDL resistance to oxidation. J. Nutr. 2004, 134, 1459–1466. [Google Scholar] [CrossRef]
- Koenig, R.T.; Dickman, J.R.; Wise, M.L.; Ji, L.L. Avenanthramides are bioavailable and accumulate in hepatic, cardiac, and skeletal muscle tissue following oral gavage in rats. J. Agric. Food Chem. 2011, 59, 6438–6443. [Google Scholar] [CrossRef]
- Chen, C.Y.; Milbury, P.E.; Collins, F.W.; Blumberg, J.B. Avenanthramides are bioavailable and have antioxidant activity in humans after acute consumption of an enriched mixture from oats. J. Nutr. 2007, 137, 1375–1382. [Google Scholar] [CrossRef]
- Zhang, T.; Shao, J.; Gao, Y.; Chen, C.; Yao, D.; Chu, Y.F.; Johnson, J.; Kang, C.; Yeo, D.; Ji, L.L. Absorption and Elimination of Oat Avenanthramides in Humans after Acute Consumption of Oat Cookies. Oxid. Med. Cell Longev. 2017, 2017, 2056705. [Google Scholar] [CrossRef]
- Sonnenburg, J.L.; Backhed, F. Diet-microbiota interactions as moderators of human metabolism. Nature 2016, 535, 56–64. [Google Scholar] [CrossRef]
- Kawabata, K.; Yoshioka, Y.; Terao, J. Role of Intestinal Microbiota in the Bioavailability and Physiological Functions of Dietary Polyphenols. Molecules 2019, 24, 370. [Google Scholar] [CrossRef] [PubMed]
- Williamson, G.; Clifford, M.N. Role of the small intestine, colon and microbiota in determining the metabolic fate of polyphenols. Biochem. Pharmacol. 2017, 139, 24–39. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Chen, H.; Zhu, Y.; McBride, J.; Fu, J.; Sang, S. Oat avenanthramide-C (2c) is biotransformed by mice and the human microbiota into bioactive metabolites. J. Nutr. 2015, 145, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Sang, S.; Chu, Y. Whole grain oats, more than just a fiber: Role of unique phytochemicals. Mol. Nutr. Food Res. 2017, 61. [Google Scholar] [CrossRef] [PubMed]
- Dall’Asta, M.; Calani, L.; Tedeschi, M.; Jechiu, L.; Brighenti, F.; Del Rio, D. Identification of microbial metabolites derived from in vitro fecal fermentation of different polyphenolic food sources. Nutrition 2012, 28, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Schar, M.Y.; Corona, G.; Soycan, G.; Dine, C.; Kristek, A.; Alsharif, S.N.S.; Behrends, V.; Lovegrove, A.; Shewry, P.R.; Spencer, J.P.E. Excretion of Avenanthramides, Phenolic Acids and their Major Metabolites Following Intake of Oat Bran. Mol. Nutr. Food Res. 2018, 62. [Google Scholar] [CrossRef] [PubMed]
- Boffetta, P.; Thies, F.; Kris-Etherton, P. Epidemiological studies of oats consumption and risk of cancer and overall mortality. Br. J. Nutr. 2014, 112 (Suppl. 2), S14–S18. [Google Scholar] [CrossRef] [Green Version]
- Murphy, N.; Moreno, V.; Hughes, D.J.; Vodicka, L.; Vodicka, P.; Aglago, E.K.; Gunter, M.J.; Jenab, M. Lifestyle and dietary environmental factors in colorectal cancer susceptibility. Mol. Asp. Med. 2019. [Google Scholar] [CrossRef] [PubMed]
- Kyro, C.; Skeie, G.; Loft, S.; Landberg, R.; Christensen, J.; Lund, E.; Nilsson, L.M.; Palmqvist, R.; Tjonneland, A.; Olsen, A. Intake of whole grains from different cereal and food sources and incidence of colorectal cancer in the Scandinavian HELGA cohort. Cancer Causes Control 2013, 24, 1363–1374. [Google Scholar] [CrossRef] [PubMed]
- Kyro, C.; Skeie, G.; Loft, S.; Overvad, K.; Christensen, J.; Tjonneland, A.; Olsen, A. Adherence to a healthy Nordic food index is associated with a lower incidence of colorectal cancer in women: The Diet, Cancer and Health cohort study. Br. J. Nutr. 2013, 109, 920–927. [Google Scholar] [CrossRef] [PubMed]
- Skeie, G.; Braaten, T.; Olsen, A.; Kyro, C.; Tjonneland, A.; Landberg, R.; Nilsson, L.M.; Wennberg, M.; Overvad, K.; Asli, L.A.; et al. Intake of whole grains and incidence of oesophageal cancer in the HELGA Cohort. Eur. J. Epidemiol. 2016, 31, 405–414. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.M.; Wang, F.; Holly, E.A. Whole grains and risk of pancreatic cancer in a large population-based case-control study in the San Francisco Bay Area, California. Am. J. Epidemiol. 2007, 166, 1174–1185. [Google Scholar] [CrossRef] [PubMed]
- Egeberg, R.; Olsen, A.; Loft, S.; Christensen, J.; Johnsen, N.F.; Overvad, K.; Tjonneland, A. Intake of whole grain products and risk of breast cancer by hormone receptor status and histology among postmenopausal women. Int. J. Cancer 2009, 124, 745–750. [Google Scholar] [CrossRef] [PubMed]
- Gil, A.; Ortega, R.M.; Maldonado, J. Wholegrain cereals and bread: A duet of the Mediterranean diet for the prevention of chronic diseases. Public Health Nutr. 2011, 14, 2316–2322. [Google Scholar] [CrossRef] [PubMed]
- Haraldsdottir, A.; Torfadottir, J.E.; Valdimarsdottir, U.A.; Adami, H.O.; Aspelund, T.; Tryggvadottir, L.; Thordardottir, M.; Birgisdottir, B.E.; Harris, T.B.; Launer, L.J.; et al. Dietary habits in adolescence and midlife and risk of breast cancer in older women. PLoS ONE 2018, 13, e0198017. [Google Scholar]
- Rawla, P. Epidemiology of Prostate Cancer. World J. Oncol. 2019, 10, 63–89. [Google Scholar] [CrossRef] [Green Version]
- Egeberg, R.; Olsen, A.; Christensen, J.; Johnsen, N.F.; Loft, S.; Overvad, K.; Tjonneland, A. Intake of whole-grain products and risk of prostate cancer among men in the Danish Diet, Cancer and Health cohort study. Cancer Causes Control 2011, 22, 1133–1139. [Google Scholar] [CrossRef]
- Torfadottir, J.E.; Valdimarsdottir, U.A.; Mucci, L.; Stampfer, M.; Kasperzyk, J.L.; Fall, K.; Tryggvadottir, L.; Aspelund, T.; Olafsson, O.; Harris, T.B.; et al. Rye bread consumption in early life and reduced risk of advanced prostate cancer. Cancer Causes Control 2012, 23, 941–950. [Google Scholar] [CrossRef] [Green Version]
- Peterson, D.M.; Hahn, M.J.; Emmons, C.L. Oat avenanthramides exhibit antioxidant activities in vitro. Food Chem. 2002, 79, 473–478. [Google Scholar] [CrossRef]
- Surh, Y. Molecular mechanisms of chemopreventive effects of selected dietary and medicinal phenolic substances. Mutat. Res. 1999, 428, 305–327. [Google Scholar] [CrossRef]
- Chu, Y.F.; Wise, M.L.; Gulvady, A.A.; Chang, T.; Kendra, D.F.; Jan-Willem van Klinken, B.; Shi, Y.; O’Shea, M. In vitro antioxidant capacity and anti-inflammatory activity of seven common oats. Food Chem. 2013, 139, 426–431. [Google Scholar] [CrossRef] [PubMed]
- Bratt, K.; Sunnerheim, K.; Bryngelsson, S.; Fagerlund, A.; Engman, L.; Andersson, R.E.; Dimberg, L.H. Avenanthramides in oats (Avena sativa L.) and structure-antioxidant activity relationships. J. Agric. Food Chem. 2003, 51, 594–600. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Wang, L.; Wang, R.; Luo, X.; Li, Y.; Li, J.; Chen, Z. Phenolic contents, cellular antioxidant activity and antiproliferative capacity of different varieties of oats. Food Chem. 2018, 239, 260–267. [Google Scholar] [CrossRef] [PubMed]
- Antonini, E.; Diamantini, G.; Ninfali, P. The effect of mechanical processing on avenanthramide and phenol levels in two organically grown Italian oat cultivars. J. Food Sci. Technol. 2017, 54, 2279–2287. [Google Scholar] [CrossRef] [PubMed]
- Scarpa, E.S.; Antonini, E.; Palma, F.; Mari, M.; Ninfali, P. Antiproliferative activity of vitexin-2-O-xyloside and avenanthramides on CaCo-2 and HepG2 cancer cells occurs through apoptosis induction and reduction of pro-survival mechanisms. Eur. J. Nutr. 2018, 57, 1381–1395. [Google Scholar] [CrossRef] [PubMed]
- Lee-Manion, A.M.; Price, R.K.; Strain, J.J.; Dimberg, L.H.; Sunnerheim, K.; Welch, R.W. In vitro antioxidant activity and antigenotoxic effects of avenanthramides and related compounds. J. Agric. Food Chem. 2009, 57, 10619–10624. [Google Scholar] [CrossRef] [PubMed]
- Antonini, E.; Iori, R.; Ninfali, P.; Scarpa, E.S. A Combination of Moringin and Avenanthramide 2f Inhibits the Proliferation of Hep3B Liver Cancer Cells Inducing Intrinsic and Extrinsic Apoptosis. Nutr. Cancer 2018, 70, 1159–1165. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Eskiw, C.H. Cytoprotective effects of Avenathramide C against oxidative and inflammatory stress in normal human dermal fibroblasts. Sci. Rep. 2019, 9, 2932. [Google Scholar] [CrossRef] [PubMed]
- Cao, G.; Prior, R.L. Measurement of oxygen radical absorbance capacity in biological samples. Methods Enzymol. 1999, 299, 50–62. [Google Scholar]
- Catanzaro, E.; Calcabrini, C.; Turrini, E.; Sestili, P.; Fimognari, C. Nrf2: A potential therapeutic target for naturally occurring anticancer drugs? Expert Opin. Ther. Targets 2017, 21, 781–793. [Google Scholar] [CrossRef]
- Fu, J.; Zhu, Y.; Yerke, A.; Wise, M.L.; Johnson, J.; Chu, Y.; Sang, S. Oat avenanthramides induce heme oxygenase-1 expression via Nrf2-mediated signaling in HK-2 cells. Mol. Nutr. Food Res. 2015, 59, 2471–2479. [Google Scholar] [CrossRef] [PubMed]
- Martin, D.; Rojo, A.I.; Salinas, M.; Diaz, R.; Gallardo, G.; Alam, J.; De Galarreta, C.M.; Cuadrado, A. Regulation of heme oxygenase-1 expression through the phosphatidylinositol 3-kinase/Akt pathway and the Nrf2 transcription factor in response to the antioxidant phytochemical carnosol. J. Biol. Chem. 2004, 279, 8919–8929. [Google Scholar] [CrossRef] [PubMed]
- Jena, N.R. DNA damage by reactive species: Mechanisms, mutation and repair. J. Biosci. 2012, 37, 503–517. [Google Scholar] [CrossRef] [PubMed]
- Hassanein, K.M.; El-Amir, Y.O. Protective effects of thymoquinone and avenanthramides on titanium dioxide nanoparticles induced toxicity in Sprague-Dawley rats. Pathol. Res. Pract. 2017, 213, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Ji, L.L.; Lay, D.; Chung, E.; Fu, Y.; Peterson, D.M. Effects of avenanthramides on oxidant generation and antioxidant enzyme activity in exercised rats. Nutr. Res. 2003, 23, 1579–1590. [Google Scholar] [CrossRef]
- Ren, Y.; Yang, X.; Niu, X.; Liu, S.; Ren, G. Chemical characterization of the avenanthramide-rich extract from oat and its effect on D-galactose-induced oxidative stress in mice. J. Agric. Food Chem. 2011, 59, 206–211. [Google Scholar] [CrossRef] [PubMed]
- Gawel, S.; Wardas, M.; Niedworok, E.; Wardas, P. Malondialdehyde (MDA) as a lipid peroxidation marker. Wiad. Lek. 2004, 57, 453–455. [Google Scholar] [PubMed]
- Hastings, J.; Kenealey, J. Avenanthramide-C reduces the viability of MDA-MB-231 breast cancer cells through an apoptotic mechanism. Cancer Cell Int. 2017, 17, 93. [Google Scholar] [CrossRef] [PubMed]
- Fu, R.; Yang, P.; Amin, S.; Li, Z. Avenanthramide A induces cellular senescence via miR-129-3p/Pirh2/p53 signaling pathway to suppress colon cancer growth. J. Agric. Food Chem. 2019, 67, 4808–4816. [Google Scholar] [CrossRef]
- Wang, D.; Wise, M.L.; Li, F.; Dey, M. Phytochemicals attenuating aberrant activation of beta-catenin in cancer cells. PLoS ONE 2012, 7, e50508. [Google Scholar]
- Balkwill, F.R.; Mantovani, A. Cancer-related inflammation: Common themes and therapeutic opportunities. Semin. Cancer Biol. 2012, 22, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Muller-Decker, K.; Furstenberger, G. The cyclooxygenase-2-mediated prostaglandin signaling is causally related to epithelial carcinogenesis. Mol. Carcinog. 2007, 46, 705–710. [Google Scholar] [CrossRef] [PubMed]
- Cianchi, F.; Cortesini, C.; Bechi, P.; Fantappie, O.; Messerini, L.; Vannacci, A.; Sardi, I.; Baroni, G.; Boddi, V.; Mazzanti, R.; et al. Up-regulation of cyclooxygenase 2 gene expression correlates with tumor angiogenesis in human colorectal cancer. Gastroenterology 2001, 121, 1339–1347. [Google Scholar] [CrossRef] [PubMed]
- Fulda, S.; Debatin, K.M. Extrinsic versus intrinsic apoptosis pathways in anticancer chemotherapy. Oncogene 2006, 25, 4798–4811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, M.E.; Howerth, E.W. Survivin: A bifunctional inhibitor of apoptosis protein. Vet. Pathol. 2004, 41, 599–607. [Google Scholar] [CrossRef] [PubMed]
- Gorrini, C.; Harris, I.S.; Mak, T.W. Modulation of oxidative stress as an anticancer strategy. Nat. Rev. Drug Discov. 2013, 12, 931–947. [Google Scholar] [CrossRef] [PubMed]
- Collado, M.; Serrano, M. Senescence in tumours: Evidence from mice and humans. Nat. Rev. Cancer 2010, 10, 51–57. [Google Scholar] [CrossRef]
- Visone, R.; Croce, C.M. MiRNAs and cancer. Am. J. Pathol. 2009, 174, 1131–1138. [Google Scholar] [CrossRef] [PubMed]
- Leng, R.P.; Lin, Y.; Ma, W.; Wu, H.; Lemmers, B.; Chung, S.; Parant, J.M.; Lozano, G.; Hakem, R.; Benchimol, S. Pirh2, a p53-induced ubiquitin-protein ligase, promotes p53 degradation. Cell 2003, 112, 779–791. [Google Scholar] [CrossRef]
- Kouhkan, F.; Mobarra, N.; Soufi-Zomorrod, M.; Keramati, F.; Hosseini Rad, S.M.; Fathi-Roudsari, M.; Tavakoli, R.; Hajarizadeh, A.; Ziaei, S.; Lahmi, R.; et al. MicroRNA-129-1 acts as tumour suppressor and induces cell cycle arrest of GBM cancer cells through targeting IGF2BP3 and MAPK1. J. Med. Genet. 2016, 53, 24–33. [Google Scholar] [CrossRef]
- Marcuzzi, E.; Angioni, R.; Molon, B.; Cali, B. Chemokines and Chemokine Receptors: Orchestrating Tumor Metastasization. Int. J. Mol. Sci. 2018, 20, 96. [Google Scholar] [CrossRef] [PubMed]
- Prieto-Garcia, E.; Diaz-Garcia, C.V.; Garcia-Ruiz, I.; Agullo-Ortuno, M.T. Epithelial-to-mesenchymal transition in tumor progression. Med. Oncol. 2017, 34, 122. [Google Scholar] [CrossRef] [PubMed]
- Mittal, V. Epithelial Mesenchymal Transition in Tumor Metastasis. Annu. Rev. Pathol. 2018, 13, 395–412. [Google Scholar] [CrossRef] [PubMed]
- Onder, T.T.; Gupta, P.B.; Mani, S.A.; Yang, J.; Lander, E.S.; Weinberg, R.A. Loss of E-cadherin promotes metastasis via multiple downstream transcriptional pathways. Cancer Res. 2008, 68, 3645–3654. [Google Scholar] [CrossRef] [PubMed]
- Jablonska-Trypuc, A.; Matejczyk, M.; Rosochacki, S. Matrix metalloproteinases (MMPs), the main extracellular matrix (ECM) enzymes in collagen degradation, as a target for anticancer drugs. J. Enzym. Inhib. Med. Chem. 2016, 31, 177–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.O.; Jeong, Y.J.; Kim, M.; Kim, C.H.; Lee, I.S. Suppression of PMA-induced tumor cell invasion by capillarisin via the inhibition of NF-kappaB-dependent MMP-9 expression. Biochem. Biophys. Res. Commun. 2008, 366, 1019–1024. [Google Scholar] [CrossRef]
- Woo, M.S.; Jung, S.H.; Kim, S.Y.; Hyun, J.W.; Ko, K.H.; Kim, W.K.; Kim, H.S. Curcumin suppresses phorbol ester-induced matrix metalloproteinase-9 expression by inhibiting the PKC to MAPK signaling pathways in human astroglioma cells. Biochem. Biophys. Res. Commun. 2005, 335, 1017–1025. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Xiong, S.; Klos, K.; Nguyen, N.; Grijalva, R.; Li, P.; Yu, D. Multiple signaling pathways involved in activation of matrix metalloproteinase-9 (MMP-9) by heregulin-beta1 in human breast cancer cells. Oncogene 2001, 20, 8066–8074. [Google Scholar] [CrossRef]
- Guo, W.; Wise, M.L.; Collins, F.W.; Meydani, M. Avenanthramides, polyphenols from oats, inhibit IL-1beta-induced NF-kappaB activation in endothelial cells. Free Radic Biol. Med. 2008, 44, 415–429. [Google Scholar] [CrossRef]
- Eberhardt, W.; Huwiler, A.; Beck, K.F.; Walpen, S.; Pfeilschifter, J. Amplification of IL-1 beta-induced matrix metalloproteinase-9 expression by superoxide in rat glomerular mesangial cells is mediated by increased activities of NF-kappa B and activating protein-1 and involves activation of the mitogen-activated protein kinase pathways. J. Immunol. 2000, 165, 5788–5797. [Google Scholar]
- Anastas, J.N.; Moon, R.T. WNT signalling pathways as therapeutic targets in cancer. Nat. Rev. Cancer 2013, 13, 11–26. [Google Scholar] [CrossRef] [PubMed]
- Aldubayan, M.A.; Elgharabawy, R.M.; Ahmed, A.S.; Tousson, E. Antineoplastic Activity and Curative Role of Avenanthramides against the Growth of Ehrlich Solid Tumors in Mice. Oxid. Med. Cell Longev. 2019, 2019, 5162687. [Google Scholar] [CrossRef] [PubMed]
- Babbar, N.; Casero, R.A., Jr. Tumor necrosis factor-alpha increases reactive oxygen species by inducing spermine oxidase in human lung epithelial cells: A potential mechanism for inflammation-induced carcinogenesis. Cancer Res. 2006, 66, 11125–11130. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.C. PCNA: A silent housekeeper or a potential therapeutic target? Trends Pharmacol. Sci. 2014, 35, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Xia, L.; Tan, S.; Zhou, Y.; Lin, J.; Wang, H.; Oyang, L.; Tian, Y.; Liu, L.; Su, M.; Cao, D.; et al. Role of the NFkappaB-signaling pathway in cancer. Onco. Targets Ther. 2018, 11, 2063–2073. [Google Scholar] [CrossRef]
- McKay, D.; Chen, C.O.; Collins, F.W.; Blumberg, J. Avenanthramide-enriched oats have an anti-inflammatory action: A pilot clinical trial. FASEB J. 2015, 29, 922.18. [Google Scholar]
- Laekeman, G.; Vlietinck, A. Assessment Report on Avena sativa L., Herba and Avena sativa L., Fructus; European Medicines Agency: London, UK, 2008; pp. 1–21. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Product Tested | Assay | Quantification of Antioxidant Activity | Phenolic/AVAs Content | Ref. |
|---|---|---|---|---|
| 7 cultivars of whole groats | ORAC | 11–28 µmol Trolox equivalents/g | Phenolic content (mg/g gallic acid equivalent): 0.57–0.94 | [52] |
| Total AVAs content (mg/kg): 9.44–163.36 | ||||
| Flour or bran from different Chinese varieties | ORAC CAA | 18.02–25.62 µmol Trolox equivalents/g | Total phenolic content (mg/g dry weight): 52.82–81.20 | [54] |
| 25.31–33.38 µmol of quercetin equivalents/100 g | Total AVAs content (mg/g dry weight): 5.01–214.26 | |||
| Two varieties cultivated in different soil types | ORAC | Free phenols (FPs): 46.3–195.3 µmol Trolox equivalents/g dry weight | Free phenols (FPs) content: 1.27–1.99 mg/g dry weight | [55] |
| Bound phenols (BPs) content: 2.02–2.71 mg/g dry weight | ||||
| AVAs content: 68.5–417.4 µg/g dry weight | ||||
| P3 fraction P4 fraction | ORACCAA DCFH-DA a | P3: 6547 µmol Trolox equivalents/g; P4: 19,079 µmol Trolox equivalents/g | AVAs average percentages present in each fraction: P3: 95% AVA 2f; P4: mixture of 37% AVA 2c, 8% 2f and 35% 2p | [56] |
| CAA: P4 > P3 in both HepG2 and Caco2 | ||||
| DCFH-DA: P4 > P3 | ||||
| AVA 2c, 2f, 2p | ORAC | 17,860–36,818 µmol Trolox equivalents/g | [17] | |
| HORAC | 16,240–19,915 µmol Trolox equivalents/g | |||
| NORAC | 1044–3038 µmol Trolox equivalents/g | |||
| SORAC | 8334–47,729 µmol Trolox equivalents/g | |||
| SOAC | 5059–20,089 µmol Trolox equivalents/g | |||
| AVA 2c, 2f, 2p | DPPH assay β-carotene assay | EC50 c (µmol): AVA2c: 0.074; AVA 2f: 0.105; AVA 2p: 0.198 | Phenolic content (gallic acid equivalent, mol/mol): AVA 2c: 1.89; AVA 2f: 1.09; AVA 2p: 0.93 | [50] |
| EC50 (µmol): AVA2c: 0.0029; AVA 2f: 0.018; AVA 2p: 0.074 | ||||
| AVA 2c, 2p, 2f | DPPH assay FRAP b assay | AVA 2c: 6.1 µmol; AVA 2p: 5.7 µmol; AVA 2f: 3.3 µmol | [57] | |
| EC1 d, (µmol/L): AVA 2c: 275; AVA 2p: 343; AVA 2f: 422 | ||||
| AVA 2f | DCFH-DA | Reduction of H2O2-induced ROS | [58] | |
| DHAvD | DCFH-DA | Reduction of UVB-induced ROS | [21] | |
| AVA 2c | Mitotracker Orange | Reduction of H2O2-induced oxidative stress | [59] |
| Product Tested | Cell Line | Assay (Treatment Time) | Range of Concentrations, EC50 a or IC50 b | Anticancer Effects | Ref. |
|---|---|---|---|---|---|
| AvExO, AVA 2c, CH3-AVA 2c | Caco-2, HT29, LS174, HCT116 | MTT c assay (48 h) | AvExO (40–160 µM); AVA 2c (40–160 µM); CH3-AVA 2c (1–80 µM) | Antiproliferative effects | [24] |
| Flour or bran from different Chinese varieties | HepG2 | MTT assay (72 h) | EC50: 167.31–233.42 mg/mL | Antiproliferative effects | [54] |
| P3 fraction P4 fraction | Caco-2, HepG2 | Sulforhodamine B assay and caspase activation (48 h) | IC50 Caco2 (µM): P3: 126.5 ± 12.5; P4: 114.6 ± 5.5 IC50 HepG2 (µM): P3: 182.7 ± 18.1; P4: 39.9 ± 4.1 | Apoptosis via activation of caspases 8 and 3 Down-regulation of VEGF and HIF1A. | [56] |
| AVA 2f | Hep3B | Sulforhodamine B assay and caspase activation (48 h) | IC50: 240 ± 10 µM | Apoptosis via activation of caspase-8 | [58] |
| AVA 2c, DH 2c, AVA 2f, DH 2f | HCT116 | MTT assay (24 h) | IC50 (µM): AVA 2c: 363; DH 2c: 158; AVA 2f: >400; DH 2f: 257 | Antiproliferative effects Apoptosis induction by DH 2c | [34] |
| AVA 2c, AVA 2c-O-glc | HCT116, HT29 | MTT assay (48 h) | IC50 HCT116 (µM): AVA 2c: 319.7; AVA 2c-O-glc: 301.1 IC50 HT29 (µM): AVA 2c: 326.8; AVA 2c-O-glc: 389.9 | Antiproliferative effects | [16] |
| AVA 2c, AVA 2f, AVA 2p | MDA-MB-231 | MTT assay; caspase activation (48–96 h) | 50–400 µM | Antiproliferative effects Apoptosis activation via caspase-3/7 (AVA 2c) | [69] |
| AVA 2c, AVA 2p, YAvn I, YAvn II | HT29, WiDr | Colony formation assay; MTT assay; gene and protein expression (72 h) | 50–200 µM | Antiproliferative effects Up-regulation of p53, p21, p27 Inhibition of EMT and tumor migration Down-regulation of E-cadherin by YAvn I and II via Snail1 and LEF-1 down-regulation | [18] |
| AVA 2f, YAvn I, YAvn II | Hela | MTT assay; cyclin D1 expression (24 h) | 25–150 µM | No cytotoxic effect up to 150 µM Decrease in cyclin D1 expression | [19] |
| AVA 2p | HCT116, HCT8 | Gene and protein expression (3, 5, 7 days) | 7.5 or 15 µM | Senescence induction via the activation of miR-129–3p/Pirh2/p53 signaling pathway Down-regulation of IGF2BP3 and CDK6 | [70] |
| DHAvD | MCF7 | MTT assay (24 h) | 1–20 µM | MMP-9 down-regulationNF-κB inhibitionInhibition of metastasis and invasion | [22] |
| AVA 2p, AVA 2f | HeLa | MTS d assay (48 h) | 20–160 µM | Antiproliferative and pro-apoptotic effects of AVA 2p through abrogation of aberrant β-catenin signalingc-Myc down-regulation | [71] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Turrini, E.; Maffei, F.; Milelli, A.; Calcabrini, C.; Fimognari, C. Overview of the Anticancer Profile of Avenanthramides from Oat. Int. J. Mol. Sci. 2019, 20, 4536. https://doi.org/10.3390/ijms20184536
Turrini E, Maffei F, Milelli A, Calcabrini C, Fimognari C. Overview of the Anticancer Profile of Avenanthramides from Oat. International Journal of Molecular Sciences. 2019; 20(18):4536. https://doi.org/10.3390/ijms20184536
Chicago/Turabian StyleTurrini, Eleonora, Francesca Maffei, Andrea Milelli, Cinzia Calcabrini, and Carmela Fimognari. 2019. "Overview of the Anticancer Profile of Avenanthramides from Oat" International Journal of Molecular Sciences 20, no. 18: 4536. https://doi.org/10.3390/ijms20184536
APA StyleTurrini, E., Maffei, F., Milelli, A., Calcabrini, C., & Fimognari, C. (2019). Overview of the Anticancer Profile of Avenanthramides from Oat. International Journal of Molecular Sciences, 20(18), 4536. https://doi.org/10.3390/ijms20184536