Deletion of a Seminal Gene Cluster Reinforces a Crucial Role of SVS2 in Male Fertility

 , , ,
, , ,  , , and
, , and 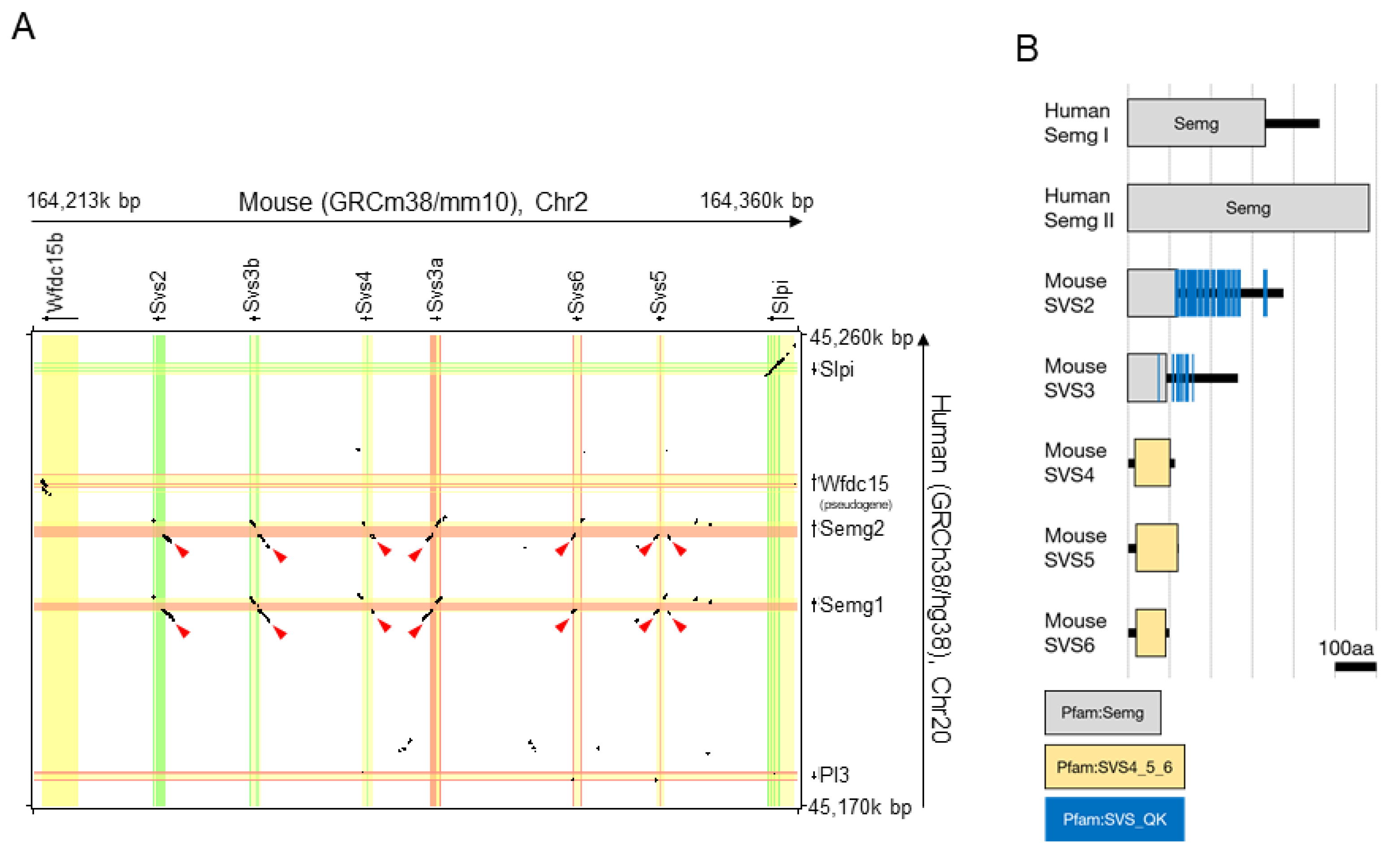{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Designing gRNA for Deletion of a Cluster of Svs2–6 Genes
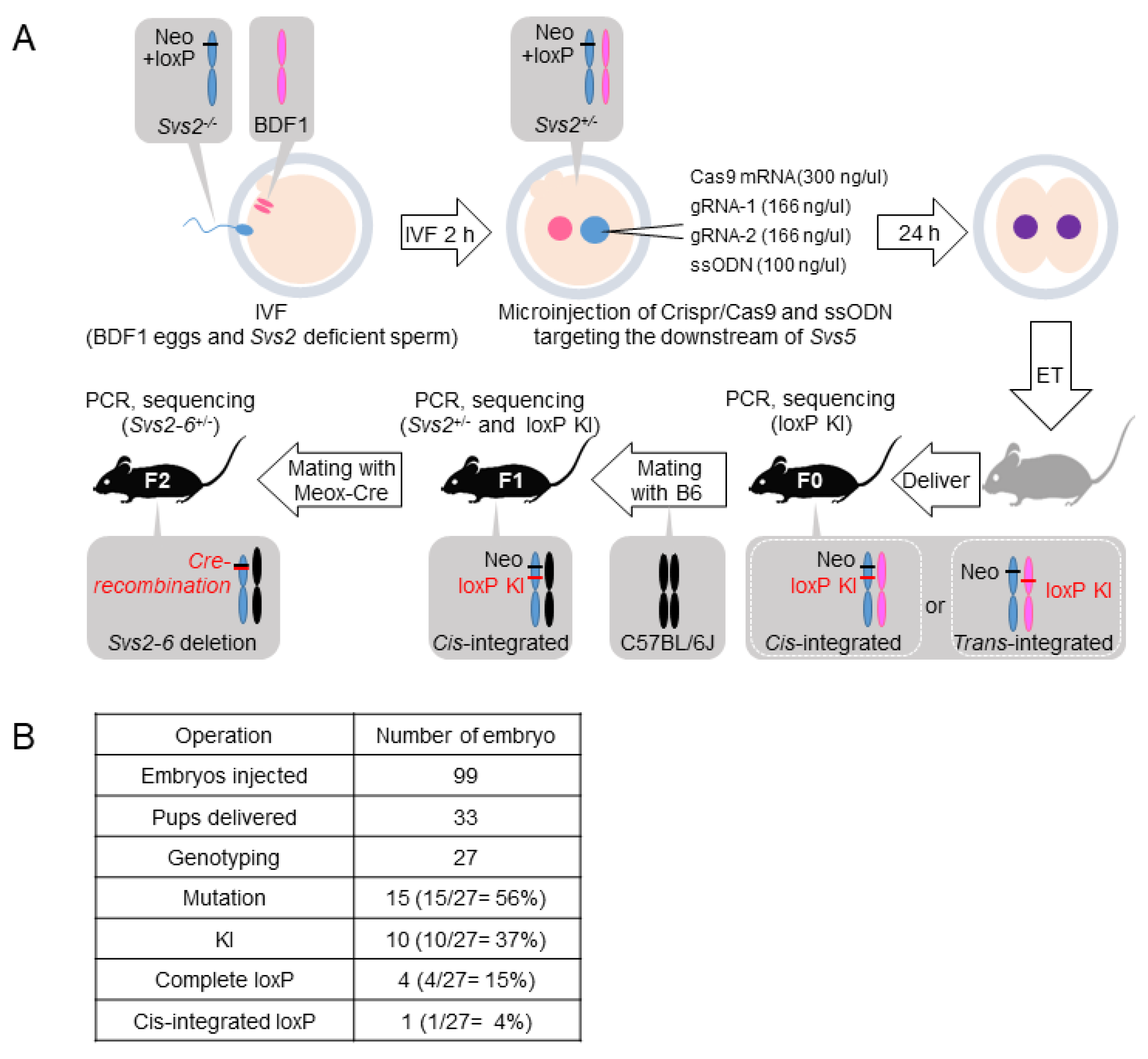
2.2. Generation of Svs2–6−/− Mice
2.3. Male Fertility of Svs2–6−/− Mice
3. Discussion
4. Materials and Methods
4.1. Production of Knockout Mice
4.2. Preparation of Seminal Vesicle Secretions
4.3. Male Fertility In Vivo and In Vitro
Author Contributions
Funding
Conflicts of Interest
References
- Ferrier, D.E.; Holland, P.W. Ancient origin of the Hox gene cluster. Nat. Rev. Genet. 2001, 2, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, J. Selfish operons: The evolutionary impact of gene clustering in prokaryotes and eukaryotes. Curr. Opin. Genet. Dev. 1999, 9, 642–648. [Google Scholar] [CrossRef]
- Swanson, W.J.; Vacquier, V.D. The rapid evolution of reproductive proteins. Nat. Rev. Genet. 2002, 3, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Karn, R.C.; Clark, N.L.; Nguyen, E.D.; Swanson, W.J. Adaptive Evolution in Rodent Seminal Vesicle Secretion Proteins. Mol. Biol. Evol. 2008, 25, 2301–2310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramm, S.A.; McDonald, L.; Hurst, J.L.; Beynon, R.J.; Stockley, P. Comparative Proteomics Reveals Evidence for Evolutionary Diversification of Rodent Seminal Fluid and Its Functional Significance in Sperm Competition. Mol. Biol. Evol. 2009, 26, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Claw, K.G.; George, R.D.; MacCoss, M.J.; Swanson, W.J. Quantitative evolutionary proteomics of seminal fluid from primates with different mating systems. BMC Genom. 2018, 19, 488. [Google Scholar] [CrossRef] [PubMed]
- Robert, M.; Gagnon, C. Semenogelin I: A coagulum forming, multifunctional seminal vesicle protein. Cell. Mol. Life Sci. CMLS 1999, 55, 944–960. [Google Scholar] [CrossRef] [PubMed]
- Dorus, S.; Evans, P.D.; Wyckoff, G.J.; Choi, S.S.; Lahn, B.T. Rate of molecular evolution of the seminal protein gene SEMG2 correlates with levels of female promiscuity. Nat. Genet. 2004, 36, 1326–1329. [Google Scholar] [CrossRef]
- Clauss, A.; Lilja, H.; Lundwall, A. A locus on human chromosome 20 contains several genes expressing protease inhibitor domains with homology to whey acidic protein. Biochem. J. 2002, 368, 233–242. [Google Scholar] [CrossRef] [Green Version]
- Clauss, A.; Lilja, H.; Lundwall, A. The evolution of a genetic locus encoding small serine proteinase inhibitors. Biochem. Biophys. Res. Commun. 2005, 333, 383–389. [Google Scholar] [CrossRef] [Green Version]
- Kawano, N.; Yoshida, M. Semen-coagulating protein, SVS2, in mouse seminal plasma controls sperm fertility. Biol. Reprod. 2007, 76, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Araki, N.; Trencsenyi, G.; Krasznai, Z.T.; Nizsaloczki, E.; Sakamoto, A.; Kawano, N.; Miyado, K.; Yoshida, K.; Yoshida, M. Seminal Vesicle Secretion 2 Acts as a Protectant of Sperm Sterols and Prevents Ectopic Sperm Capacitation in Mice. Biol. Reprod. 2015, 92, 8. [Google Scholar] [CrossRef] [PubMed]
- Kawano, N.; Araki, N.; Yoshida, K.; Hibino, T.; Ohnami, N.; Makino, M.; Kanai, S.; Hasuwa, H.; Yoshida, M.; Miyado, K.; et al. Seminal vesicle protein SVS2 is required for sperm survival in the uterus. Proc. Natl. Acad. Sci. USA 2014, 141, 4145–4150. [Google Scholar] [CrossRef] [PubMed]
- Miyado, M.; Yoshida, K.; Miyado, K.; Katsumi, M.; Saito, K.; Nakamura, S.; Ogata, T.; Fukami, M. Knockout of Murine Mamld1 Impairs Testicular Growth and Daily Sperm Production but Permits Normal Postnatal Androgen Production and Fertility. Int. J. Mol. Sci. 2017, 18, 1300. [Google Scholar] [CrossRef] [PubMed]
- Amelar, R.D. Coagulation, liquefaction and viscosity of human semen. J. Urol. 1962, 87, 187–190. [Google Scholar] [CrossRef]
- Jensen-Seaman, M.I.; Li, W.-H. Evolution of the hominoid semenogelin genes, the major proteins of ejaculated semen. J. Mol. Evol. 2003, 57, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Edstrom, A.M.L.; Malm, J.; Frohm, B.; Martellini, J.A.; Giwercman, A.; Morgelin, M.; Cole, A.M.; Sorensen, O.E. The major bactericidal activity of human seminal plasma is zinc-dependent and derived from fragmentation of the semenogelins. J. Immunol. 2008, 181, 3413–3421. [Google Scholar] [CrossRef]
- Iwamoto, T.; Tsang, A.; Luterman, M.; Dickson, J.; Delamirande, E.; Okuno, M.; Mohri, H.; Gagnon, C. Purification and Characterization of a Sperm Motility-Dynein Atpase Inhibitor from Boar Seminal Plasma. Mol. Reprod. Dev. 1992, 31, 55–62. [Google Scholar] [CrossRef]
- Lundwall, A. The cloning of a rapidly evolving seminal-vesicle-transcribed gene encoding the major clot-forming protein of mouse semen. Eur. J. Biochem. 1996, 235, 424–430. [Google Scholar] [CrossRef]
- Hughes, A.L.; Yeager, M. Natural selection at major histocompatibility complex loci of vertebrates. Annu. Rev. Genet. 1998, 32, 415–435. [Google Scholar] [CrossRef]
- Ota, Y.; Tamegai, H.; Kudo, F.; Kuriki, H.; Koike-Takeshita, A.; EOUCHI, T.; Kakinuma, K. Butirosin-biosynthetic gene cluster from Bacillus circulans. J. Antibiot. 2000, 53, 1158–1167. [Google Scholar] [CrossRef] [PubMed]
- Semple, C.A.M.; Rolfe, M.; Dorin, J.R. Duplication and selection in the evolution of primate beta-defensin genes. Genome. Biol. 2003, 4. [Google Scholar] [CrossRef]
- Fawcett, J.; Innan, H. Neutral and non-neutral evolution of duplicated genes with gene conversion. Genes 2011, 2, 191–209. [Google Scholar] [CrossRef] [PubMed]
- Jonjić, S.; Polić, B.; Krmpotić, A. Viral inhibitors of NKG2D ligands: Friends or foes of immune surveillance? Eur. J. Immunol. 2008, 38, 2952–2956. [Google Scholar] [CrossRef] [PubMed]
- Kondo, M.; Maruoka, T.; Otsuka, N.; Kasamatsu, J.; Fugo, K.; Hanzawa, N.; Kasahara, M. Comparative genomic analysis of mammalian NKG2D ligand family genes provides insights into their origin and evolution. Immunogenetics 2010, 62, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Elizondo, L.I.; Jafar-Nejad, P.; Clewing, J.M.; Boerkoel, C.F. Gene clusters, molecular evolution and disease: A speculation. Curr. Genom. 2009, 10, 64–75. [Google Scholar] [CrossRef] [PubMed]
- Inui, M.; Mokuda, S.; Sato, T.; Tamano, M.; Takada, S.; Asahara, H. Dissecting the roles of miR-140 and its host gene. Nat. Cell. Biol. 2018, 20, 516. [Google Scholar] [CrossRef]
- Tallquist, M.D.; Soriano, P. Epiblast-restricted Cre expression in MORE mice: A tool to distinguish embryonic vs. extra-embryonic gene function. Genesis 2000, 26, 113–115. [Google Scholar] [CrossRef]
- Iwai, M.; Harada, Y.; Miyabayashi, R.; Kang, W.; Nakamura, A.; Kawano, N.; Miyamoto, Y.; Yamada, M.; Hamatani, T.; Miyado, M.; et al. Chemotactic behavior of egg mitochondria in response to sperm fusion in mice. Heliyon 2018, 4, e00944. [Google Scholar] [CrossRef]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shindo, M.; Inui, M.; Kang, W.; Tamano, M.; Tingwei, C.; Takada, S.; Hibino, T.; Yoshida, M.; Yoshida, K.; Okada, H.; et al. Deletion of a Seminal Gene Cluster Reinforces a Crucial Role of SVS2 in Male Fertility. Int. J. Mol. Sci. 2019, 20, 4557. https://doi.org/10.3390/ijms20184557
Shindo M, Inui M, Kang W, Tamano M, Tingwei C, Takada S, Hibino T, Yoshida M, Yoshida K, Okada H, et al. Deletion of a Seminal Gene Cluster Reinforces a Crucial Role of SVS2 in Male Fertility. International Journal of Molecular Sciences. 2019; 20(18):4557. https://doi.org/10.3390/ijms20184557
Chicago/Turabian StyleShindo, Miyuki, Masafumi Inui, Woojin Kang, Moe Tamano, Cai Tingwei, Shuji Takada, Taku Hibino, Manabu Yoshida, Kaoru Yoshida, Hiroshi Okada, and et al. 2019. "Deletion of a Seminal Gene Cluster Reinforces a Crucial Role of SVS2 in Male Fertility" International Journal of Molecular Sciences 20, no. 18: 4557. https://doi.org/10.3390/ijms20184557
APA StyleShindo, M., Inui, M., Kang, W., Tamano, M., Tingwei, C., Takada, S., Hibino, T., Yoshida, M., Yoshida, K., Okada, H., Iwamoto, T., Miyado, K., & Kawano, N. (2019). Deletion of a Seminal Gene Cluster Reinforces a Crucial Role of SVS2 in Male Fertility. International Journal of Molecular Sciences, 20(18), 4557. https://doi.org/10.3390/ijms20184557