First Insights on the Presence of the Unfolded Protein Response in Human Spermatozoa



Abstract
:1. Introduction
2. Results
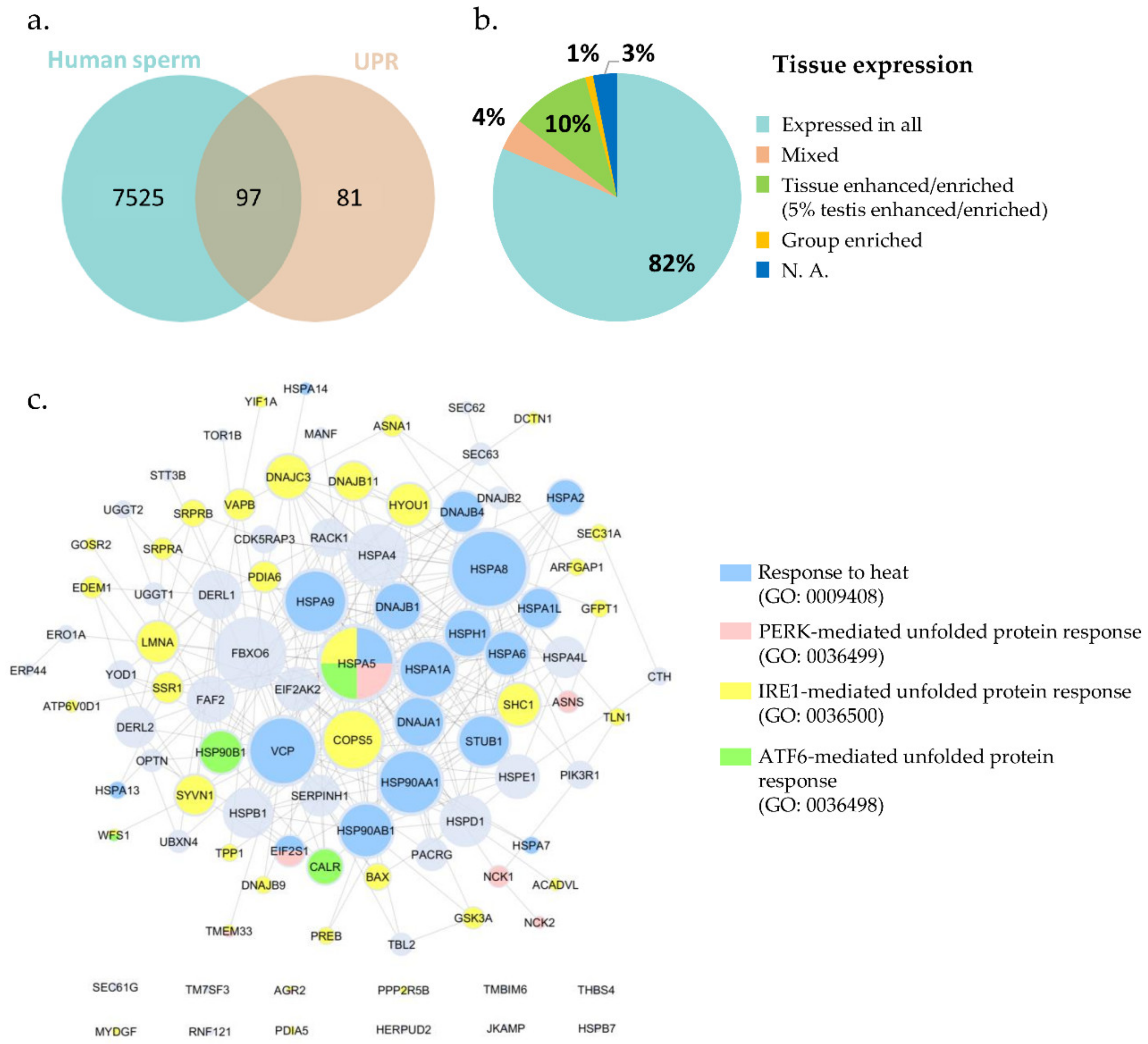
2.1. UPR-Related Proteins are Present in Ejaculated Human Spermatozoa
2.2. Identification of HSF1, GADD34, and Phosphorylated eIF2α in Human Spermatozoa for the First Time
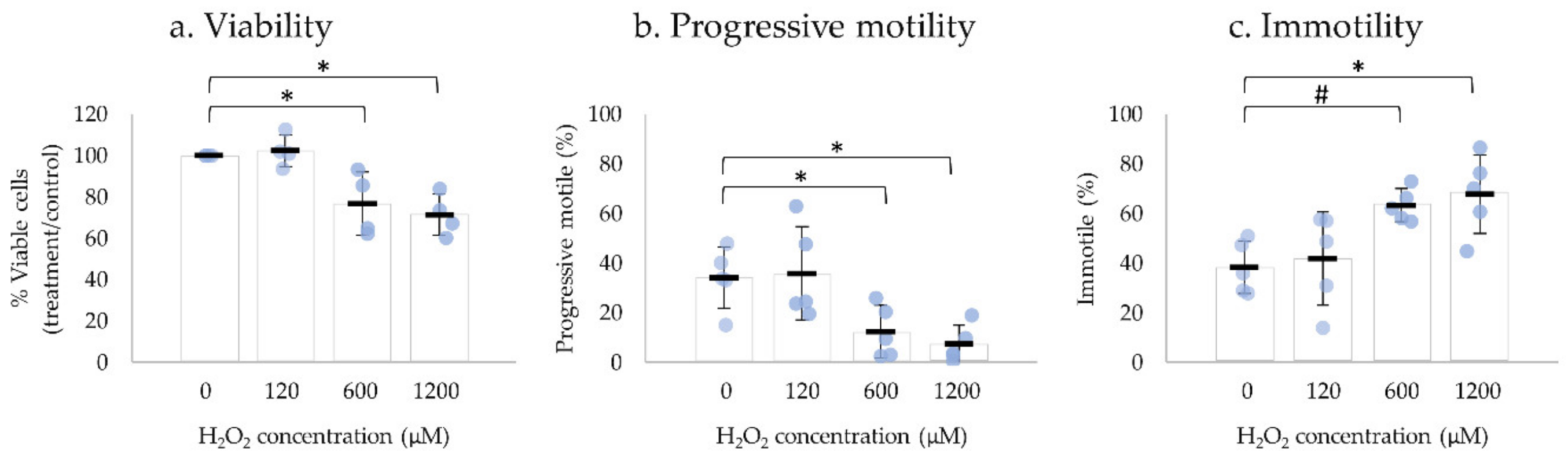
2.3. H2O2 Affects Human Sperm Viability and Motility
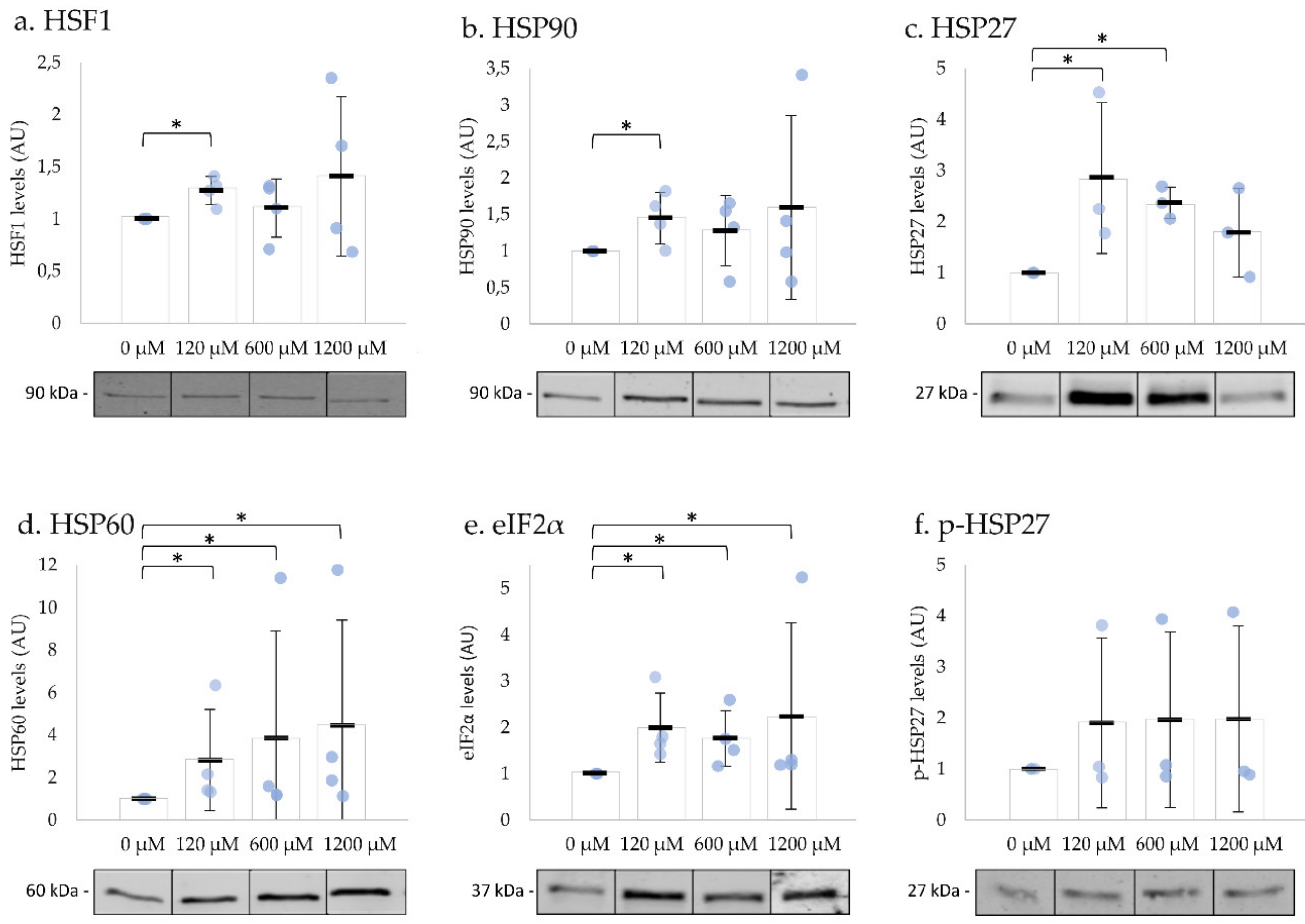
2.4. UPR-Related Proteins’ Levels Significantly Increased after Exposure to H2O2
3. Discussion
4. Materials and Methods
4.1. Identification of Proteins Involved in Response to Unfolded Proteins in Human Sperm
4.2. Bioinformatic Analysis: Gene Annotation and Involvement of UPR Proteins in Male Infertility-Related Phenotypes
4.3. Ethical Approval
4.4. Sperm Preparation
4.5. Whole Cell Lysates
4.6. Oxidative Stress Induction
4.7. Cell Viability Assays
4.8. Motility Assays
4.9. Antibodies
4.10. Western Blot
4.11. Co-Immunoprecipitation of PPP1CC2
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| UPR | Unfolded protein response |
| H2O2 | Hydrogen peroxide |
| HSF1 | Heat shock factor 1 |
| GADD34 | DNA damage-inducible protein GADD34 |
| HSP | Heat shock protein |
| eIF2α | Eukaryotic translation initiation factor-2 |
| ER | Endoplasmic reticulum |
| HSR | Heat shock response |
| UPRER | Endoplasmic reticulum unfolded protein response |
| UPRmt | Mitochondrial unfolded protein response |
| HPA | Human Protein Atlas |
| PPP1 | Serine/threonine-protein phosphatase 1 |
| ROS | Reactive oxygen species |
| PPP1CC2 | Phosphoprotein phosphatase 1 subunit gamma 2 |
| SD | Standard deviation |
| PPI | Protein–protein interaction |
References
- Saito, A.; Imaizumi, K. Unfolded protein response-dependent communication and contact among endoplasmic reticulum, mitochondria, and plasma membrane. Int. J. Mol. Sci. 2018, 19, 3215. [Google Scholar] [CrossRef] [PubMed]
- Doultsinos, D.; Avril, T.; Lhomond, S.; Dejeans, N.; Guédat, P.; Chevet, E. Control of the unfolded protein response in health and disease. SLAS Discov. Adv. Life Sci. RD 2017, 22, 787–800. [Google Scholar] [CrossRef] [PubMed]
- Lindholm, D.; Korhonen, L.; Eriksson, O.; Kõks, S. Recent insights into the role of unfolded protein response in ER stress in health and disease. Front. cell Dev. Biol. 2017, 5, 48. [Google Scholar] [CrossRef] [PubMed]
- Jovaisaite, V.; Mouchiroud, L.; Auwerx, J. The mitochondrial unfolded protein response, a conserved stress response pathway with implications in health and disease. J. Exp. Biol. 2014, 217, 137–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergmann, T.J.; Molinari, M. Three branches to rule them all? UPR signalling in response to chemically versus misfolded proteins-induced ER stress. Biol. Cell 2018, 110, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Boyce, M.; Bryant, K.F.; Jousse, C.; Long, K.; Harding, H.P.; Scheuner, D.; Kaufman, R.J.; Ma, D.; Coen, D.M.; Ron, D.; et al. A selective inhibitor of eIF2α dephosphorylation protects cells from ER stress. Science. 2005, 307. [Google Scholar] [CrossRef]
- Zhang, G.; Ling, X.; Liu, K.; Wang, Z.; Zou, P.; Gao, J.; Cao, J.; Ao, L. The p-eIF2α/ATF4 pathway links endoplasmic reticulum stress to autophagy following the production of reactive oxygen species in mouse spermatocyte-derived cells exposed to dibutyl phthalate. Free Radic. Res. 2016, 50, 698–707. [Google Scholar] [CrossRef]
- Pesch, S.; Bergmann, M. Structure of mammalian spermatozoa in respect to viability, fertility and cryopreservation. Micron 2006, 37, 597–612. [Google Scholar] [CrossRef]
- Keber, R.; Rozman, D.; Horvat, S. Sterols in spermatogenesis and sperm maturation. J. Lipid Res. 2012, 54, 20–33. [Google Scholar] [CrossRef]
- Kim, J.H.; Park, S.J.; Kim, T.S.; Park, H.J.; Park, J.; Kim, B.K.; Kim, G.R.; Kim, J.M.; Huang, S.M.; Chae, J.-I.; et al. Testicular hyperthermia induces unfolded protein response signaling activation in spermatocyte. Biochem. Biophys. Res. Commun. 2013, 434, 861–866. [Google Scholar] [CrossRef]
- Ji, Y.-L.; Wang, H.; Zhao, X.-F.; Wang, Q.; Zhang, C.; Zhang, Y.; Zhao, M.; Chen, Y.-H.; Meng, X.-H.; Xu, D.-X. Crosstalk between endoplasmic reticulum stress and mitochondrial pathway mediates cadmium-induced germ cell apoptosis in testes. Toxicol. Sci. 2011, 124, 446–459. [Google Scholar] [CrossRef] [PubMed]
- Karna, K.K.; Shin, Y.S.; Choi, B.R.; Kim, H.K.; Park, J.K. The role of endoplasmic reticulum stress response in male reproductive physiology and pathology: A Review. World J. Mens. Health 2019, 37. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Agarwal, A.; Mohanty, G.; Hamada, A.J.; Gopalan, B.; Willard, B.; Yadav, S.; du Plessis, S. Proteomic analysis of human spermatozoa proteins with oxidative stress. Reprod. Biol. Endocrinol. 2013, 11, 48. [Google Scholar] [CrossRef] [PubMed]
- Amaral, A.; Castillo, J.; Ramalho-Santos, J.; Oliva, R. The combined human sperm proteome: Cellular pathways and implications for basic and clinical science. Hum. Reprod. Update 2014, 20, 40–62. [Google Scholar] [CrossRef] [PubMed]
- Amaral, A.; Paiva, C.; Attardo Parrinello, C.; Estanyol, J.M.; Ballescà, J.L.; Ramalho-Santos, J.; Oliva, R. Identification of Proteins Involved in Human Sperm Motility Using High-Throughput Differential Proteomics. J. Proteome Res. 2014, 13, 5670–5684. [Google Scholar] [CrossRef]
- McReynolds, S.; Dzieciatkowska, M.; Stevens, J.; Hansen, K.C.; Schoolcraft, W.B.; Katz-Jaffe, M.G. Toward the identification of a subset of unexplained infertility: A sperm proteomic approach. Fertil. Steril. 2014, 102, 692–699. [Google Scholar] [CrossRef]
- Liu, Y.; Guo, Y.; Song, N.; Fan, Y.; Li, K.; Teng, X.; Guo, Q.; Ding, Z. Proteomic pattern changes associated with obesity-induced asthenozoospermia. Andrology 2015, 3, 247–259. [Google Scholar] [CrossRef]
- Liu, X.; Liu, F.J.; Jin, S.H.; Wang, Y.W.; Liu, X.X.; Zhu, P.; Wang, W.T.; Liu, J.; Wang, W.J. Comparative proteome analysis of human testis from newborn, young adult, and aged men identified spermatogenesis-associated proteins. Electrophoresis 2015. [Google Scholar] [CrossRef]
- Jumeau, F.; Com, E.; Lane, L.; Duek, P.; Lagarrigue, M.; Lavigne, R.; Guillot, L.; Rondel, K.; Gateau, A.; Melaine, N.; et al. Human Spermatozoa as a Model for Detecting Missing Proteins in the Context of the Chromosome-Centric Human Proteome Project. J. Proteome Res. 2015, 14, 3606–3620. [Google Scholar] [CrossRef]
- Wang, Y.; Wan, J.; Ling, X.; Liu, M.; Zhou, T. The human sperm proteome 2.0: An integrated resource for studying sperm functions at the level of posttranslational modification. Proteomics 2016, 16, 2597–2601. [Google Scholar] [CrossRef]
- Bogle, O.A.; Kumar, K.; Attardo-Parrinello, C.; Lewis, S.E.M.; Estanyol, J.M.; Ballescà, J.L.; Oliva, R. Identification of protein changes in human spermatozoa throughout the cryopreservation process. Andrology 2017, 5, 10–22. [Google Scholar] [CrossRef] [PubMed]
- Vandenbrouck, Y.; Lane, L.; Carapito, C.; Duek, P.; Rondel, K.; Bruley, C.; Macron, C.; Gonzalez de Peredo, A.; Couté, Y.; Chaoui, K.; et al. Looking for missing proteins in the proteome of human spermatozoa: An update. J. Proteome Res. 2016, 15, 3998–4019. [Google Scholar] [CrossRef] [PubMed]
- Garin-Muga, A.; Odriozola, L.; Martínez-Val, A.; del Toro, N.; Martínez, R.; Molina, M.; Cantero, L.; Rivera, R.; Garrido, N.; Dominguez, F.; et al. Detection of missing proteins using the PRIDE database as a source of mass spectrometry evidence. J. Proteome Res. 2016, 15, 4101–4115. [Google Scholar] [CrossRef] [PubMed]
- Intasqui, P.; Camargo, M.; Del Giudice, P.T.; Spaine, D.M.; Carvalho, V.M.; Cardozo, K.H.M.; Cedenho, A.P.; Bertolla, R.P. Unraveling the sperm proteome and post-genomic pathways associated with sperm nuclear DNA fragmentation. J. Assist. Reprod. Genet. 2013, 30, 1187–1202. [Google Scholar] [CrossRef] [Green Version]
- Dalet, A.; Argüello, R.J.; Combes, A.; Spinelli, L.; Jaeger, S.; Fallet, M.; Vu Manh, T.-P.; Mendes, A.; Perego, J.; Reverendo, M.; et al. Protein synthesis inhibition and GADD34 control IFN-β heterogeneous expression in response to dsRNA. EMBO J. 2017, 36, 761–782. [Google Scholar] [CrossRef]
- Fardilha, M.; Ferreira, M.; Pelech, S.; Vieira, S.; Rebelo, S.; Korrodi-Gregorio, L.; Sousa, M.; Barros, A.; Silva, V.; da Cruz e Silva, O.A.B.; et al. “Omics” of human sperm: Profiling protein phosphatases. OMICS 2013, 17, 460–472. [Google Scholar] [CrossRef]
- Chakrabarti, R.; Kline, D.; Lu, J.; Orth, J.; Pilder, S.; Vijayaraghavan, S. Analysis of Ppp1cc-null mice suggests a role for PP1gamma2 in sperm morphogenesis. Biol. Reprod. 2007, 76, 992–1001. [Google Scholar] [CrossRef]
- Chaki, S.; Misro, M. Assessment of human sperm function after hydrogen peroxide exposure: Development of a vaginal contraceptive. Contraception 2002, 66, 187–192. [Google Scholar] [CrossRef]
- Evdokimov, V.V.; Barinova, K.V.; Turovetskii, V.B.; Muronetz, V.I.; Schmalhausen, E.V. Low concentrations of hydrogen peroxide activate the antioxidant defense system in human sperm cells. Biochem. 2015, 80, 1178–1185. [Google Scholar] [CrossRef]
- Hayashida, N.; Inouye, S.; Fujimoto, M.; Tanaka, Y.; Izu, H.; Takaki, E.; Ichikawa, H.; Rho, J.; Nakai, A. A novel HSF1-mediated death pathway that is suppressed by heat shock proteins. EMBO J. 2006, 25, 4773–4783. [Google Scholar] [CrossRef] [Green Version]
- Tian, Y.; Merkwirth, C.; Dillin, A. Mitochondrial UPR: A double-edged sword. Trends Cell Biol. 2016, 26, 563–565. [Google Scholar] [CrossRef] [PubMed]
- Pellegrino, M.W.; Nargund, A.M.; Haynes, C.M. Signaling the mitochondrial unfolded protein response. Biochim. Biophys. Acta - Mol. Cell Res. 2013, 1833, 410–416. [Google Scholar] [CrossRef] [PubMed]
- Kus-Liśkiewicz, M.; Polańska, J.; Korfanty, J.; Olbryt, M.; Vydra, N.; Toma, A.; Widłak, W. Impact of heat shock transcription factor 1 on global gene expression profiles in cells which induce either cytoprotective or pro-apoptotic response following hyperthermia. BMC Genomics 2013, 14, 456. [Google Scholar] [CrossRef] [PubMed]
- Korfanty, J.; Stokowy, T.; Widlak, P.; Gogler-Piglowska, A.; Handschuh, L.; Podkowi??ski, J.; Vydra, N.; Naumowicz, A.; Toma-Jonik, A.; Widlak, W. Crosstalk between HSF1 and HSF2 during the heat shock response in mouse testes. Int. J. Biochem. Cell Biol. 2014, 57, 76–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, L.; Dai, Y.; Cui, Z.; Jiang, X.; Liu, W.; Han, F.; Lin, A.; Cao, J.; Liu, J. The regulation of cellular apoptosis by the ROS-triggered PERK/EIF2α/chop pathway plays a vital role in bisphenol A-induced male reproductive toxicity. Toxicol. Appl. Pharmacol. 2017, 314, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Chen, X.; Wang, Z.; Wang, D. Is transcription in sperm stationary or dynamic? J. Reprod. Dev. 2017, 63, 439–443. [Google Scholar] [CrossRef] [Green Version]
- Rossitto, M.; Philibert, P.; Poulat, F. Molecular events and signalling pathways of male germ cell differentiation in mouse. Semin. Cell Dev. Biol. 2015, 45, 84–93. [Google Scholar] [CrossRef]
- De Kretser, D.M.; Loveland, K.L.; Meinhardt, A.; Simorangkir, D.; Wreford, N. Spermatogenesis. Hum. Reprod. 1998, 13, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Melber, A.; Haynes, C.M. UPRmt regulation and output: A stress response mediated by mitochondrial-nuclear communication. Cell Res. 2018, 28, 281–295. [Google Scholar] [CrossRef]
- Rajender, S.; Rahul, P.; Mahdi, A.A. Mitochondria, spermatogenesis and male infertility. Mitochondrion 2010, 10, 419–428. [Google Scholar] [CrossRef]
- Lee, A.S. The ER chaperone and signaling regulator GRP78/BiP as a monitor of endoplasmic reticulum stress. Methods 2005, 35, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Widlak, W.; Vydra, N.; Malusecka, E.; Dudaladava, V.; Winiarski, B.; Ścieglińska, D.; Widlak, P. Heat shock transcription factor 1 down-regulates spermatocyte-specific 70 kDa heat shock protein expression prior to the induction of apoptosis in mouse testes. Genes to Cells 2007, 12, 487–499. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.; Park, K.; Hwang, D.S.; Rhee, K. Importance of eIF2α phosphorylation as a protective mechanism against heat stress in mouse male germ cells. Mol. Reprod. Dev. 2017, 84, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Somanath, P.R.; Chakrabarti, R.; Eddy, E.M.; Vijayaraghavan, S. Changes in intracellular distribution and activity of protein phosphatase PP1gamma2 and its regulating proteins in spermatozoa lacking AKAP4. Biol. Reprod. 2005, 72, 384–392. [Google Scholar] [CrossRef]
- Silva, J.V.; Yoon, S.; De Bock, P.-J.; Goltsev, A.V.; Gevaert, K.; Mendes, J.F.F.; Fardilha, M. Construction and analysis of a human testis/sperm-enriched interaction network: Unraveling the PPP1CC2 interactome. Biochim. Biophys. Acta - Gen. Subj. 2017, 1861, 375–385. [Google Scholar] [CrossRef]
- Korrodi-Gregório, L.; Ferreira, M.; Vintém, A.P.; Wu, W.; Muller, T.; Marcus, K.; Vijayaraghavan, S.; Brautigan, D.L.; Odete, A.B.; Fardilha, M.; et al. Identification and characterization of two distinct PPP1R2 isoforms in human spermatozoa. BMC Cell Biol. 2013, 14, 2–14. [Google Scholar] [CrossRef]
- Biggiogera, M.; Tanguay, R.M.; Marin, R.; Wu, Y.; Martin, T.E.; Fakan, S. Localization of Heat Shock Proteins in mouse male germ cells: An immunoelectron microscopical study. Exp. Cell Res. 1996, 229, 77–85. [Google Scholar] [CrossRef]
- Calle-Guisado, V.; Bragado, M.J.; García-Marín, L.J.; González-Fernández, L. HSP90 maintains boar spermatozoa motility and mitochondrial membrane potential during heat stress. Anim. Reprod. Sci. 2017, 0–1. [Google Scholar] [CrossRef]
- Rao, M.; Xia, W.; Yang, J.; Hu, L.X.; Hu, S.F.; Lei, H.; Wu, Y.Q.; Zhu, C.H. Transient scrotal hyperthermia affects human sperm DNA integrity, sperm apoptosis, and sperm protein expression. Andrology 2016, 4, 1054–1063. [Google Scholar] [CrossRef]
- Gur, Y.; Breitbart, H. Mammalian sperm translate nuclear-encoded proteins by mitochondrial-type ribosomes. Genes Dev. 2006, 20, 411–416. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.; Umehara, T.; Okazaki, T.; Goto, M.; Fujita, Y.; Hoque, S.A.M.; Kawai, T.; Zeng, W.; Shimada, M. Gene expression and protein synthesis in mitochondria enhance the duration of high-speed linear motility in boar sperm. Front. Physiol. 2019, 10, 252. [Google Scholar] [CrossRef] [PubMed]
- Deng, S.-L.; Sun, T.-C.; Yu, K.; Wang, Z.-P.; Zhang, B.-L.; Zhang, Y.; Wang, X.-X.; Lian, Z.-X.; Liu, Y.-X. Melatonin reduces oxidative damage and upregulates heat shock protein 90 expression in cryopreserved human semen. Free Radic. Biol. Med. 2017, 113, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Ferlin, A.; Speltra, E.; Patassini, C.; Pati, M.A.; Garolla, A.; Caretta, N.; Foresta, C. Heat shock protein and heat shock factor expression in sperm: relation to oligozoospermia and varicocele. J. Urol. 2010, 183, 1248–1252. [Google Scholar] [CrossRef]
- Jodar, M.; Sendler, E.; Krawetz, S.A. The protein and transcript profiles of human semen. Cell Tissue Res. 2016, 363, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, W.; Duan, Y. Effect of H 2 O 2 induced oxidative stress (OS) on volatile organic compounds (VOCs) and intracellular metabolism in MCF-7 breast cancer cells. J. Breath Res. 2019, 13, 036005. [Google Scholar] [CrossRef] [PubMed]
- Bardou, P.; Mariette, J.; Escudié, F.; Djemiel, C.; Klopp, C. Jvenn: An interactive Venn diagram viewer. BMC Bioinformatics 2014, 15, 293. [Google Scholar] [CrossRef] [PubMed]
- Alanis-Lobato, G.; Andrade-Navarro, M.A.; Schaefer, M.H. HIPPIE v2.0: Enhancing meaningfulness and reliability of protein–protein interaction networks. Nucleic Acids Res. 2017, 45, D408–D414. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- World Health Organization. WHO Laboratory Manual for the Examination and Processing of Human Semen; World Health Organization: Geneva, Switzerland, 2010; ISBN 9789241547789. [Google Scholar]
- Da Cruz e Silva, E.F.; Fox, C.A.; Ouimet, C.C.; Gustafson, E.; Watson, S.J.; Greengard, P. Differential expression of protein phosphatase 1 isoforms in mammalian brain. J. Neurosci. 1995, 15, 3375–3389. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| References | Total Proteins Identified in Each Study | Total Proteins Identified without Duplicates in All Studies | UPR Proteins Identified in Each Study | Total UPR Proteins Identified in Human Sperm Proteome |
|---|---|---|---|---|
| Sharma et al. 2013 [13] | 72 | 7622 | 6 | 97 |
| Amaral et al. 2014 * [14] | 6198 | 93 | ||
| Amaral et al. 2014 [15] | 1624 | 33 | ||
| McReynolds et al. 2014 [16] | 49 | 3 | ||
| Y. Liu et al. 2014 [17] | 1974 | 37 | ||
| F. Liu et al. 2015 [18] | 22 | 1 | ||
| Jumeau et al. 2015 [19] | 94 | 0 | ||
| Wang et al. 2016 * [20] | 2132 | 42 | ||
| Bogle et al. 2016 [21] | 264 | 8 | ||
| Vandenbrouk et al. 2016 [22] | 235 | 0 | ||
| Garin-Muga et al. 2016 [23] | 111 | 0 | ||
| Intasqui et al. 2017 [24] | 572 | 22 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santiago, J.; Silva, J.V.; Fardilha, M. First Insights on the Presence of the Unfolded Protein Response in Human Spermatozoa. Int. J. Mol. Sci. 2019, 20, 5518. https://doi.org/10.3390/ijms20215518
Santiago J, Silva JV, Fardilha M. First Insights on the Presence of the Unfolded Protein Response in Human Spermatozoa. International Journal of Molecular Sciences. 2019; 20(21):5518. https://doi.org/10.3390/ijms20215518
Chicago/Turabian StyleSantiago, Joana, Joana Vieira Silva, and Margarida Fardilha. 2019. "First Insights on the Presence of the Unfolded Protein Response in Human Spermatozoa" International Journal of Molecular Sciences 20, no. 21: 5518. https://doi.org/10.3390/ijms20215518
APA StyleSantiago, J., Silva, J. V., & Fardilha, M. (2019). First Insights on the Presence of the Unfolded Protein Response in Human Spermatozoa. International Journal of Molecular Sciences, 20(21), 5518. https://doi.org/10.3390/ijms20215518