Organ-Specific Analysis of Morus alba Using a Gel-Free/Label-Free Proteomic Technique

 ,
, 
Abstract
:1. Introduction
2. Results
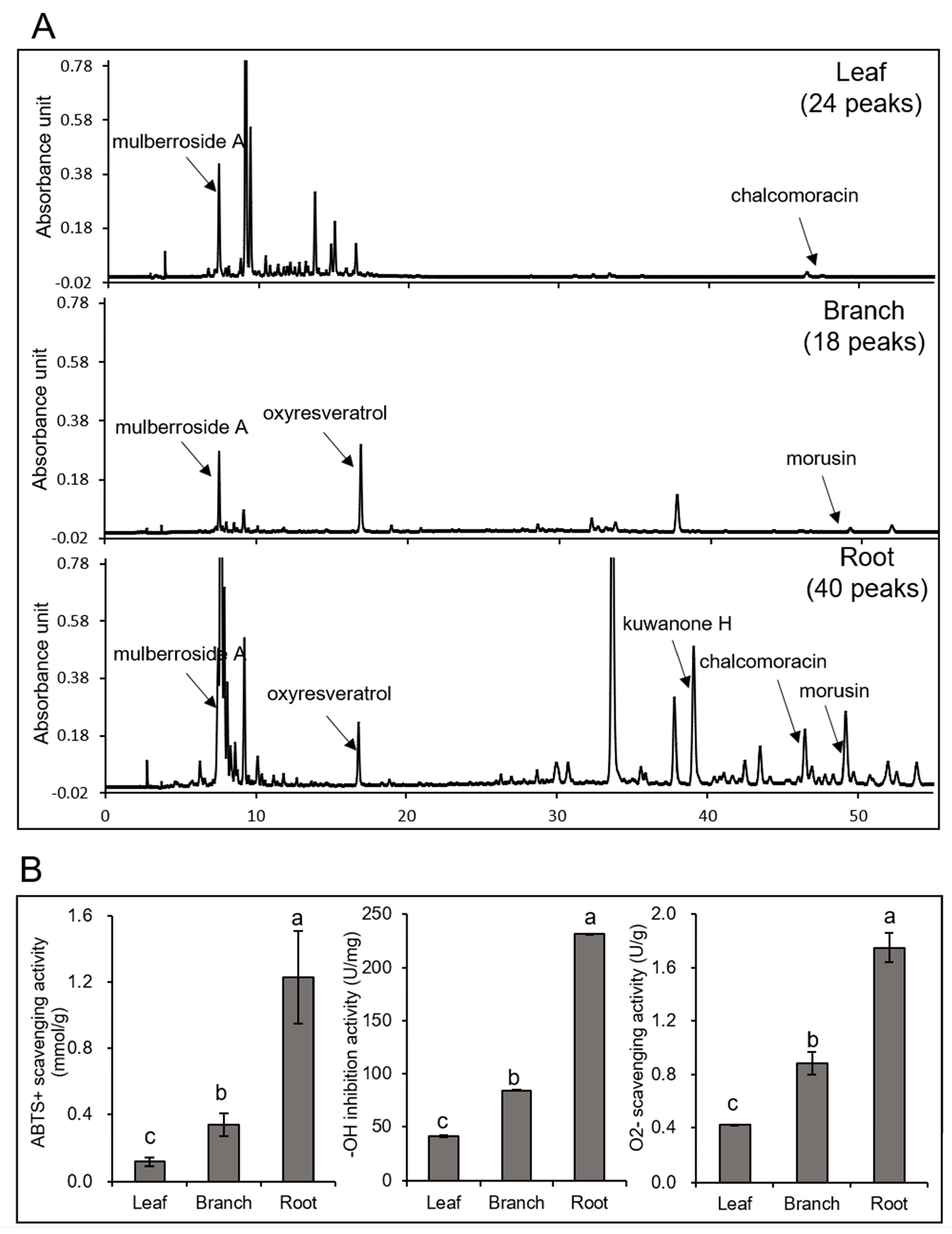
2.1. The Metabolite Contents and Antioxidant Activity in Morus Root were Higher than in Branch and Leaf
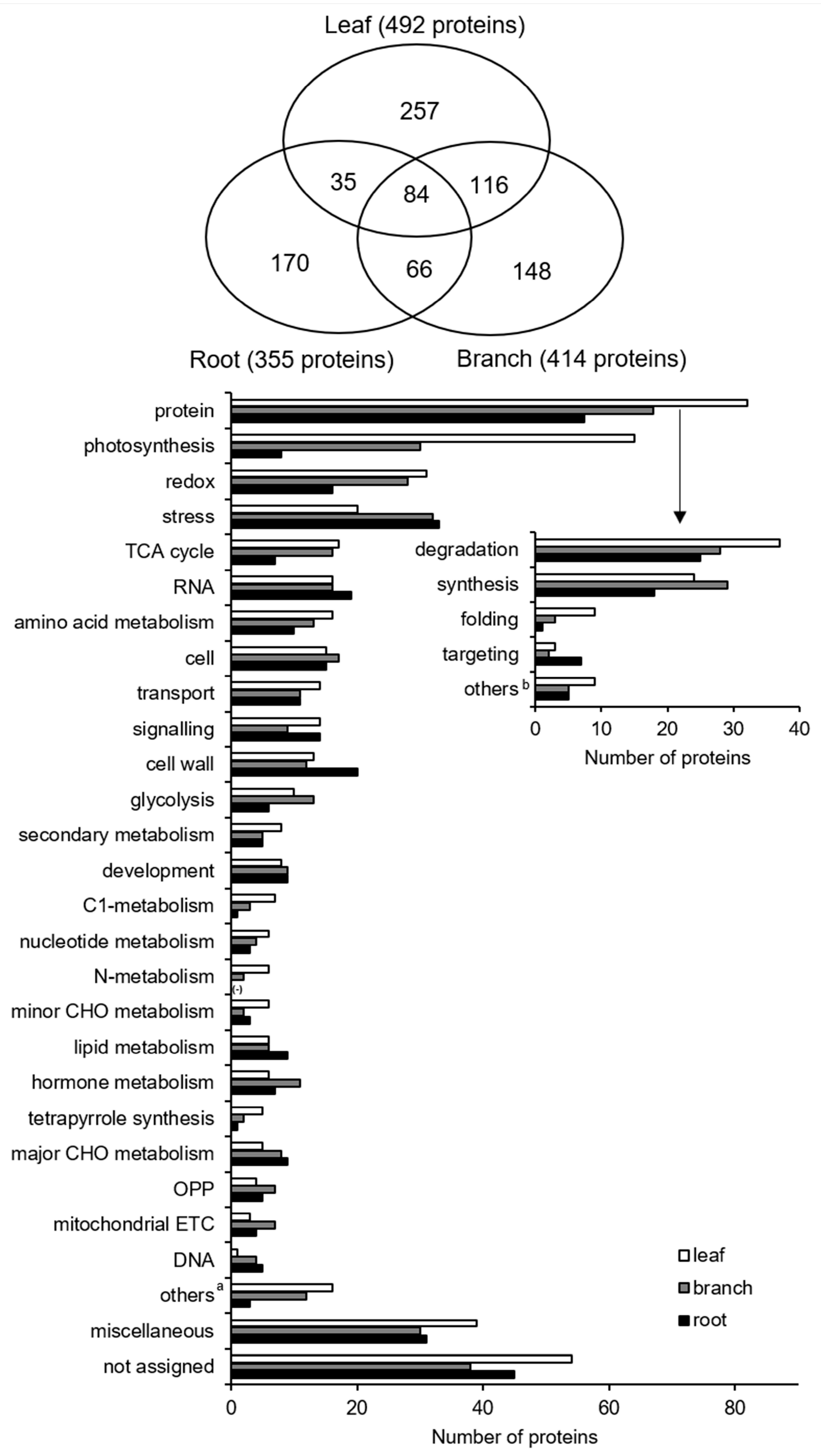
2.2. A Total of 257, 148, and 170 Proteins were Specific to Leaf, Branch, and Root in Morus, Respectively
2.3. Functional Characterization of Organ-Specific Proteins Identified from Morus
2.4. Largely Differential Common Proteins Were Identified among Three Organs in Morus
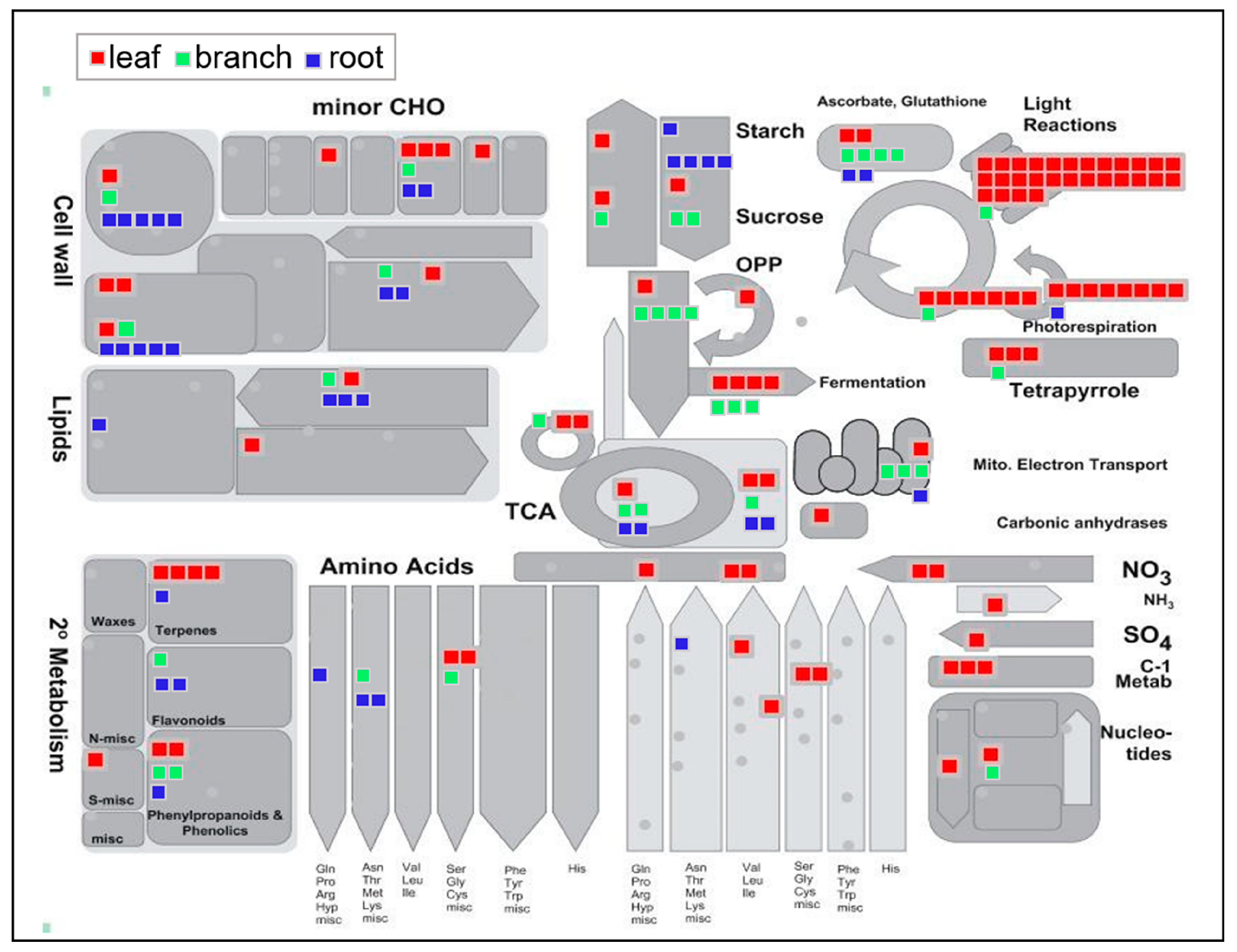
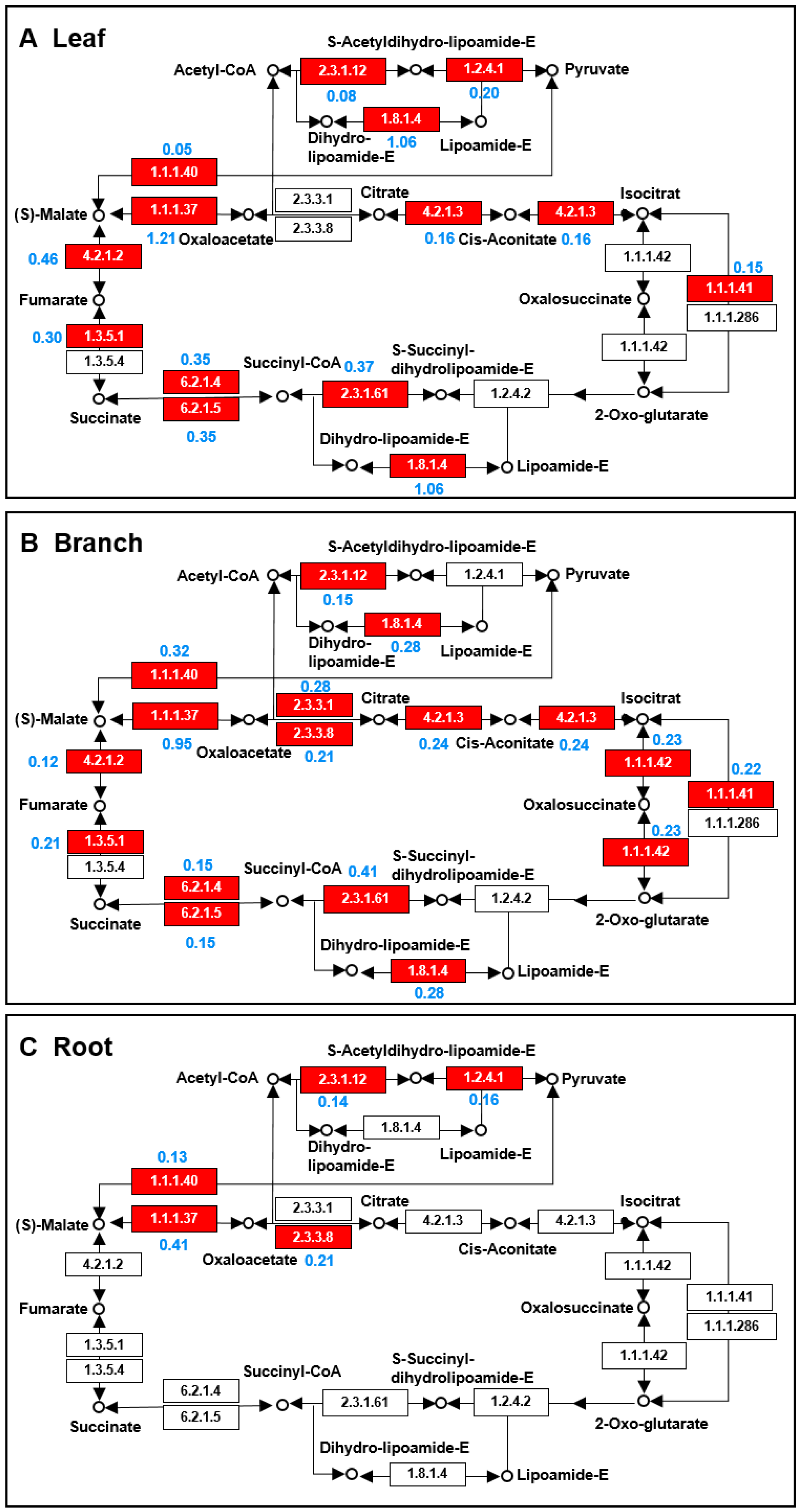
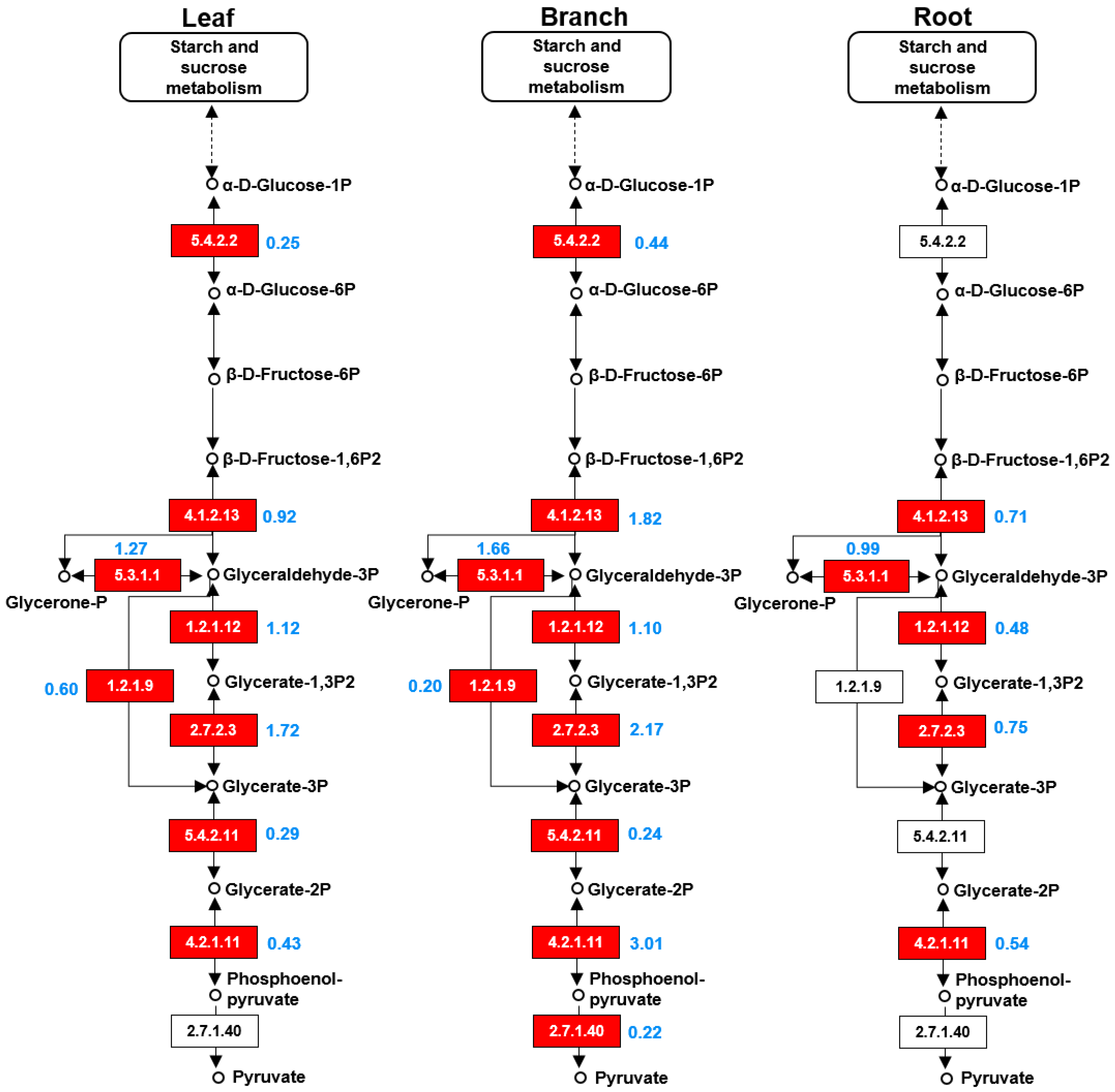
2.5. TCA Cycle and Glycolysis Pathways Largely Differed among the Three Organs
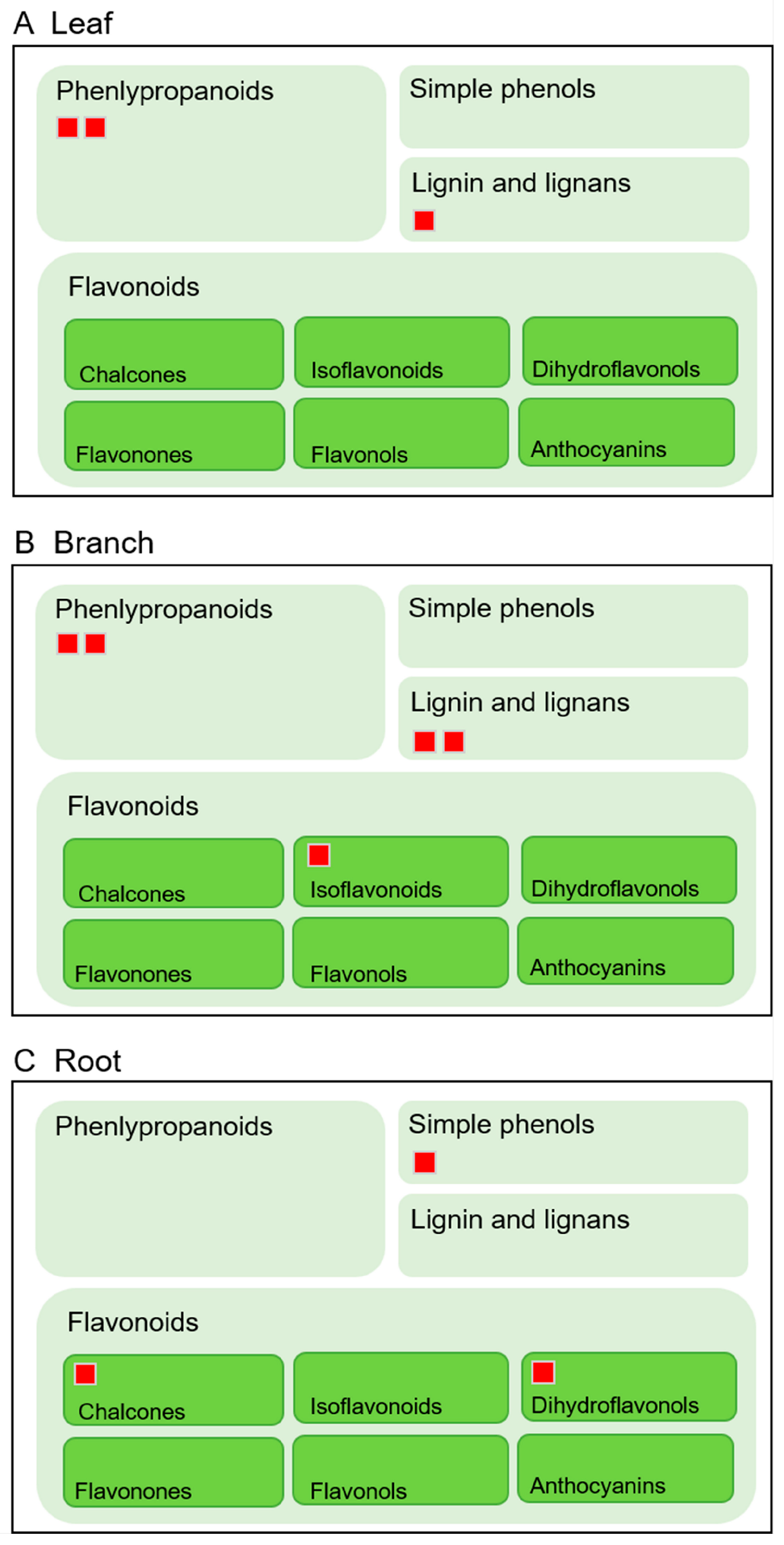
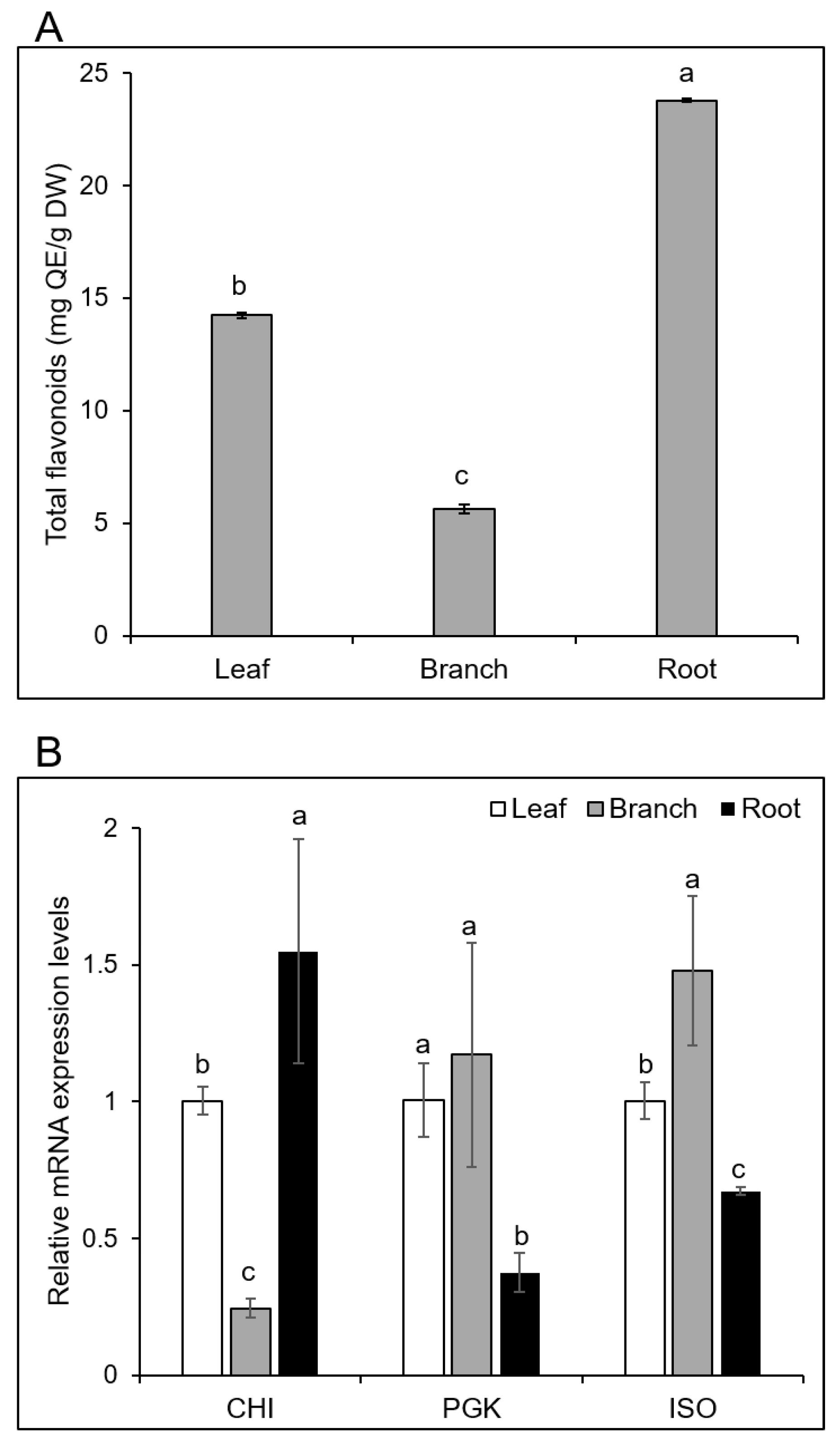
2.6. Total Flavonoid Contents were the Highest in the Roots
2.7. Expression of Genes Related to Root- and Branch-Specific Proteins
3. Discussion
3.1. The Secondary Metabolites and Total Flavonoid Contents are Different in Morus Leaf, Branch, and Root
3.2. Anaylses of Enzymes Involved in the Glycolysis and Isoflavonoid Biosynthetic Pathway in Morus
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Protein Extraction
4.3. Purification and Digestion of Proteins for Mass Spectrometry Analysis
4.4. Nanoliquid Chromatography-Tandem Mass Spectrometry Analysis
4.5. Protein Identification from the Mass Spectrometry Data
4.6. Functional Analysis of Identified Proteins
4.7. Quantitative Analysis of Metabolites from Morus
4.8. In Vitro Antioxidant Activity Analysis of Mulberry Leaf, Branch, and Root
4.9. RNA Extraction and Quantitative Reverse Transcription-Polymerase Chain Reaction Analysis
4.10. Quantitative Analysis of the Total Flavonoids in Three Morus Organs
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ABTS | 2,2’-Azino-bis (3-ethylbenzothiazoline-6-sulfonic acid) |
| CHAPS | 3-[(3-Cholamidopropyl) dimethylammonio] propanesulfonate |
| CHI | Chalcone flavanone isomerase |
| emPAI | Exponentially-modified protein abundance index |
| HPLC | High-performance liquid chromatography |
| ISO | Isoflavone reductase |
| IFRh | Isoflavonoid reductase homolog |
| MS | Mass spectrometry |
| MVA | Mevalonate |
| PGK | Phosphoglycerate kinase |
| POD | Peroxidase |
| qRT-PCR | Quantitative reverse transcription-polymerase chain reaction |
| SOD | Superoxide dismutase |
| TCA | Tricarboxylic acid |
| TEAC | Trolox-equivalent antioxidant capacity |
References
- Umate, P. Mulberry improvements via plastid transformation and tissue culture engineering. Plant Signal. Behav. 2010, 5, 785–787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, J.; He, N.J.; Huang, X.Z.; Xiang, Z.H. Development of mulberry ecological industry and sericulture. Sci. Seric. 2010, 36, 984–989. (In Chinese) [Google Scholar]
- Liu, Y.; Willison, J.H.M. Prospects for cultivating white mulberry (Morus alba) in the drawdown zone of the Three Gorges Reservoir, China. Environ. Sci. Pollut. Res. 2013, 20, 7142–7151. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.G.; Chen, R.Y.; Yu, D.Q. Structures of two new benzofuran derivatives from the bark of mulberry tree (Morus macroura Miq.). J. Asian Nat. Prod. Res. 2001, 3, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Carrizzo, A.; Ambrosio, M.; Damato, A.; Madonna, M.; Storto, M.; Capocci, L.; Campiglia, P.; Sommella, E.; Trimarco, V.; Rozza, F.; et al. Morus alba extract modulates blood pressure homeostasis through eNOS signaling. Mol. Nutr. Food Res. 2016, 60, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Q.; Xie, Y.; Wang, W.; Yan, Y.; Ye, H.; Jabbar, S.; Zeng, X. Extraction iptimization, characterization and antioxidant activity in vitro of polysaccharides from mulberry (Morus alba L.) leaves. Carbohyd. Polym. 2015, 128, 52–62. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.Y.; Wang, J.; Ma, J.; Zhang, Y.Q. Interference effect of oral administration of mulberry branch bark powder on the incidence of type II diabetes in mice induced by streptozotocin. Food Nutr. Res. 2016, 60, 31606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, S.Z.; Tuo, Z.D.; Li, J.L.; Xing, S.S.; Li, B.B.; Zhang, L.; Lee, H.S.; Chen, J.G.; Cui, L. Effects of Morus root bark extract and active constituents on blood lipids in hyperlipidemia rats. J. Ethnopharmacol. 2016, 180, 54–59. [Google Scholar] [CrossRef]
- Shang, X.F.; He, X.R.; He, X.Y.; Li, M.X.; Zhang, R.X.; Fan, P.C.; Zhang, Q.L.; Jia, Z.P. The genus Scutellaria an ethnopharmacological and phytochemical review. J. Ethnopharmacol. 2010, 128, 279–313. [Google Scholar] [CrossRef] [PubMed]
- Lam, S.C.; Liu, X.; Chen, X.Q.; Hu, D.J.; Zhao, J.; Long, Z.R.; Fan, B.; Li, S.P. Chemical characteristics of different parts of Coreopsis tinctorial in China using microwave-assisted extraction and high-performance liquid chromatography followed by chemometric analysis. J. Sep. Sci. 2016, 39, 2919–2927. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Rahman, A.A.; Islam, S.; Khandokhar, P.; Parvin, S.; Islam, M.B.; Hossain, M.; Rashid, M.; Sadik, G.; Nasrin, S.; Mollah, M.N.H.; Alam, A.H.M.K. A comparative study on the antioxidant activity of methanolic extracts from different parts of Morus alba L. (Moraceae). BMC Research Notes 2013, 6, 24. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zu, Y.; Fu, Y.; Efferth, T. In vitro antioxidant and antimicrobial activity of extracts from Morus alba L. leaves, stems and fruits. Am. J. Chin. Med. 2012, 40, 349–356. [Google Scholar] [CrossRef]
- Chang, L.W.; Juang, L.J.; Wang, B.S.; Wang, M.Y.; Tai, H.M.; Hung, W.J.; Chen, Y.J.; Huang, M.H. Antioxidant and antityrosinase activity of mulberry (Morus alba L.) twigs and root bark. Food Chem. Toxicol. 2011, 49, 785–790. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Zhang, L.; Tai, Y.; Wang, X.; Ho, C.T.; Wan, X. Gene discovery of characteristic metabolic pathways in the tea plant (Camellia sinensis) using ‘Omics’-based network approaches: A future perspective. Front. Plant Sci. 2018, 9, 480. [Google Scholar] [CrossRef] [PubMed]
- García-Sevillano, M.A.; García-Barrera, T.; Abril, N.; Pueyo, C.; Lopez-Barea, J.; Gomez-Ariza, J.L. Omics technologies and their applications to evaluate metal toxicity in mice M. spretus as a bioindicator. J. Proteom. 2014, 104, 4–23. [Google Scholar] [CrossRef]
- Zhu, W.; Hu, J.; Wang, X.; Tian, J.K.; Komatsu, S. Organ-specific analysis of Mahonia using gel-free/label-free proteomic technique. J. Proteome Res. 2015, 14, 2669–2685. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.L.; Gai, Y.P.; Zheng, C.C.; Mu, Z.M. Comparative proteomic analysis provides new insights into mulberry dwarf responses in mulberry (Morus alba L.). Proteomics 2009, 9, 5328–5339. [Google Scholar] [CrossRef]
- Ma, X.; Iwanaka, N.; Masuda, S.; Karaike, K.; Egawa, T.; Hamada, T.; Toyoda, T.; Myamoto, L.; Nakao, K.; Hayashi, T. Morus alba leaf extract stimulates 5’-AMP-activated protein kinase in isolated rat skeletal muscle. J. Ethnopharmacol. 2009, 122, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.W.; Ko, W.M.; Park, J.H.; Seo, K.H.; Oh, E.J.; Lee, D.Y.; Lee, D.S.; Kim, Y.C.; Lim, D.W.; Han, D.; Baek, N.I. Isoprenylated flavonoids from the root bark of Morus alba and their hepatoprptective and neuroprotective activities. Arch. Pharm. Res. 2015, 38, 2066–2075. [Google Scholar] [CrossRef]
- Eo, H.J.; Park, G.H.; Jeong, J.B. The involvement of cyclin D1 degradation through GSK3β-mediated threonine-286 phosphorylation-dependent nuclear export in anti-cancer activity of mulberry root bark extracts. Phytomedicine 2016, 23, 105–113. [Google Scholar] [CrossRef]
- Chahid, K.; Laglaoui, A.; Zantar, S.; Ennabili, A. Antioxidant-enzyme reaction to the oxidative stress due to alpha-cypermethrin, chlorpyriphos, and pirimicarb in tomato (Lycopersicon esculentum Mill.). Environ. Sci. Pollut. Res. Int. 2015, 22, 18115–18126. [Google Scholar] [CrossRef] [PubMed]
- Qin, R.; Jiao, Y.; Zhang, S.; Jiang, W.; Liu, D. Effects of aluminum on nucleoli in root tip cells and selected physiological and biochemical characters in Allium cepa var. agrogarum L. BMC Plant Biol. 2010, 10, 225. [Google Scholar]
- Prashanth, S.R.; Sadhasivam, V.; Parida, A. Over expression of cytosolic copper/zine superoxide dismutase from a mangrove plant Avicennia marina in indica rice var: Pusa basmati-1 confers abiotic stress tolerance. Transgenic Res. 2008, 17, 281–291. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Chan, E.W.C.; Lye, P.Y.; Wang, S.K. Phytochemistry, pharmacology, and clinical trials of Morus alba. Chin. J. Nat. Med. 2016, 14, 17–30. [Google Scholar] [PubMed]
- Chen, Z.Y.; Du, X.; Yang, Y.Y.; Cui, X.M.; Zhang, Z.J.; Li, Y. Comparative study of chemical composition and active components against α-glucosidase of various medicinal parts of Morus alba L. Biomed. Chromatogr. 2018, 32, e4328. [Google Scholar] [CrossRef] [PubMed]
- Dastmalchi, M.; Dhaubhadel, S. Soybean chalcone isomerase: Evolution of the fold, and the differential expression and localization of the gene family. Planta 2015, 241, 507–523. [Google Scholar] [CrossRef]
- Bogs, J.; Ebadi, A.; McDavid, D.; Robinson, S.P. Identification of the flavonoid hydroxylases from grapevine and their regulation during fruit development. Plant Physiol. 2006, 140, 279–291. [Google Scholar] [CrossRef]
- Liu, S.H.; Ju, J.F.; Xia, G.M. Identification of the flavonoid 3’-hydroxylase and flavonoid 3’,5’-hydroxylase genes from Antarctic moss and their regulation during abiotic stress. Gene 2014, 543, 145–152. [Google Scholar] [CrossRef]
- Fini, A.; Brunetti, C.; Ferdinando, M.D.; Ferrini, F.; Tattini, M. Stress-induced flavonoid biosynthesis and the antioxidant machinery of plants. Plant Signal. Behav. 2011, 5, 709–711. [Google Scholar] [CrossRef]
- Zgiby, S.M.; Thomson, G.J.; Qamar, S.; Berry, A. Exploring substrate binding and discrimination in fructose 1,6-bisphosphate and tagatose 1,6-bisphosphate aldolases. Eur. J. Biochem. 2000, 267, 1858–1868. [Google Scholar] [CrossRef] [PubMed]
- Watson, H.C.; Walker, N.P.; Shaw, P.J.; Bryant, T.N.; Wendell, P.L.; Fothergill, L.A.; Perkins, R.E.; Conroy, S.C.; Dobson, M.J.; Tuite, M.F. Sequence and structure of yeast phosphoglycerate kinase. EMBO J. 1982, 1, 1635–1640. [Google Scholar] [CrossRef] [PubMed]
- Hoom, R.K.; Flickweert, J.P.; Staal, G.E. Purification and properties of enolase of human erythroctyes. Int. J. Biochem. 1974, 5, 845–852. [Google Scholar]
- Cramer, G.R.; Sluyter, S.C.V.; Hopper, D.W.; Pascovici, D.; Keighley, T.; Haynes, P.A. Proteomic analysis indicates massive changes in metabolism prior to the inhibition of growth and photosynthesis of grapevine (Vitis vinifera L.) in response to water deficit. BMC Plant Biol. 2013, 13, 49. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Dong, Z.; Zhang, Y.; Guo, K.; Guo, P.; Zhao, P.; Xia, Q. Proteomics provides insight into the interaction between mulberry and silkworm. J. Proteome Res. 2017, 16, 2477–2480. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Ma, Z.F.; Luo, X.; Li, X. Effects of mulberry fruit (Morus alba L.) consumption on health outcomes: A mini-review. Antioxidants 2018, 7, 69. [Google Scholar] [CrossRef]
- Qiu, F.; He, T.Z.; Zhang, Y.Q. The isolation and the characterization of two polysaccharides from the branch bark of mulberry (Morus alba L.). Arch. Pharm. Res. 2016, 39, 887–896. [Google Scholar] [CrossRef]
- Wang, S.; Fang, M.; Ma, Y.L.; Zhang, Y.Q. Preparation of the branch bark ethanol extract in mulberry Morus alba, its antioxidation, and antihyperglycemic activity in vivo. Evid-Based Complement. Altern. 2014, 2014, 569652. [Google Scholar]
- Graham, T.L.; Kim, J.E.; Graham, M.Y. Role of constitutive isoflavone conjugates in the accumulation of glyceollin in soybean infected with Phytophthora megasperma. Mol. Plant Microbe Interact. 1990, 3, 157–166. [Google Scholar] [CrossRef]
- Guo, L.; Dixon, R.A.; Paiva, N.L. Conversion of vestitone to medicarpin in Alfalfa (Medicago sativa L.) is catalyzed by two independent enzymes. J. Biol. Chem. 1994, 269, 22372–22378. [Google Scholar]
- Cooper, J.D.; Qiu, F.; Paiva, N.L. Biotransformation of an exogenously supplied isoflavonoid by transgenic tobacco cells expressing alfalfa isoflavone reductase. Plant Cell Rep. 2002, 20, 876–884. [Google Scholar]
- Cheng, Q.; Li, N.; Dong, L.; Zhang, D.; Fang, S.; Jiang, L.; Wang, X.; Xu, P.; Zhang, S. Overexpression of soybean isoflavone reductase (GmIFR) enhances resistance to Phytophthora sojae in soybean. Front. Plant. Sci. 2015, 6, 1024. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of proteindye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Nanjo, Y.; Skultety, L.; Uvackova, L.; Klubicova, K.; Hajduch, M.; Komatsu, S. Mass spectrometry-based analysis of proteomic changes in the root tips of flooded soybean seedlings. J. Proteome Res. 2012, 11, 372–385. [Google Scholar] [CrossRef] [PubMed]
- Olsen, J.V.; de Godoy, L.M.F.; Li, G.Q.; Macek, B.; Mortensen, P.; Pesch, R.; Makarov, A.; Lange, O.; Horning, S.; Mann, M. Parts per million mass accuracy on an Orbitrap mass spectrometer via lock mass injection into a C-trap. Mol. Cell. Proteom. 2005, 4, 2010–2021. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wen, Z.H.; Washburn, M.P.; Florens, L. Effect of dynamic exclusion duration on spectral count based quantitative proteomics. Anal. Chem. 2009, 81, 6317–6326. [Google Scholar] [CrossRef] [PubMed]
- Brosch, M.; Yu, L.; Hubbard, T.; Choudhary, J. Accurate and sensitive peptide identification with MASCOT percolator. J. Proteome Res. 2009, 8, 3176–3181. [Google Scholar] [CrossRef] [PubMed]
- Shinoda, K.; Tomita, M.; Ishihama, Y. emPAI Calc-for the estimation of protein abundance from large-scale identification data by liquid chromatography-tandem mass spectrometry. Bioinfomatics 2010, 26, 576–577. [Google Scholar] [CrossRef] [PubMed]
- Usadel, B.; Nagel, A.; Thimm, O.; Redestig, H.; Blaesing, O.E.; Natalia, P.R.; Selbig, J.; Hannemann, J.; Piques, M.C.; Steinhauser, D.; et al. Extension of the visualization tool MapMan to allow statistical analysis of arrays, display of corresponding genes, and comparison with known responses. Plant Physiol. 2005, 138, 1195–1204. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Fenton, H.J.H. Oxidation of tartaric acid in presence of iron. J. Chem. Soc. Trans. 1894, 65, 899–910. [Google Scholar] [CrossRef]
- Guan, Q.J.; Yu, J.J.; Zhu, W.; Yang, B.X.; Li, Y.H.; Zhang, L.; Tian, J.K. RNA-Seq transcriptome analysis of the Morus alba L. leaves exposed to high-level UV-B with and without dark treatment. Gene 2018, 645, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Li, Y.; Bao, T.; Gowd, V. Mulberry fruit extract affords protection against ethyl carbamate-induced cytotoxicity and oxidative stress. Oxid. Med. Cell. Longev. 2017, 2017, 1594963. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mulberroside A (mg/g DW) | Oxyresveratrol (mg/g DW) | Kuwanone H (mg/g DW) | Chalcomoracin (mg/g DW) | Morusin (mg/g DW) | |
|---|---|---|---|---|---|
| Leaf | 0.917 ± 0.015 | n.d. * | n.d. | 0.045 ± 0.005 | n.d. |
| Branch | 0.451 ± 0.012 | 0.453 ± 0.008 | n.d. | n.d. | 0.043 ± 0.008 |
| Root | 24.206 ± 0.688 | 0.345 ± 0.022 | 5.551 ± 0.226 | 0.610 ± 0.051 | 2.874 ± 0.158 |
| No. | Protein ID a | Description | M.P. b | Score | Mass (Da) | Function c | Mol (%) d | ||
|---|---|---|---|---|---|---|---|---|---|
| leaf | stem | root | |||||||
| 1 | Morus009492.p1 | Macrophage migration inhibitory factor homolog | 8 | 203 | 8961 | not assigned | 3.10 | 2.43 | 3.69 |
| 2 | Morus001961.p1 | Peroxidase 12 | 16 | 244 | 38,426 | misc | 0.57 | 1.29 | 3.62 |
| 3 | Morus018316.p1 | Superoxide dismutase 1 copper chaperone | 9 | 339 | 11,171 | metal handling | 2.11 | 1.69 | 2.48 |
| 4 | Morus009000.p1 | 60S acidic ribosomal protein P2B | 12 | 110 | 11,673 | protein | 1.62 | 2.32 | 2.32 |
| 5 | Morus023628.p1 | Tubulin beta-1 chain | 22 | 578 | 51,015 | cell | 1.11 | 1.83 | 1.99 |
| 6 | Morus017847.p1 | Ribonuclease UK114 | 16 | 266 | 19,960 | RNA | 1.08 | 0.27 | 1.80 |
| 7 | Morus017207.p1 | Proteasome subunit alpha type-4 | 6 | 173 | 27,440 | protein | 0.24 | 0.34 | 1.58 |
| 8 | Morus003952.p1 | Lipoxygenase homology domain-containing protein 1 | 6 | 154 | 21,171 | not assigned | 0.50 | 0.95 | 1.44 |
| 9 | Morus022430.p1 | Proteasome subunit beta type-1 | 11 | 313 | 24,861 | protein | 0.68 | 0.56 | 1.44 |
| 10 | Morus022592.p1 | Thaumatin-like protein 1a | 18 | 362 | 26,984 | stress | 1.02 | 0.44 | 1.41 |
| 11 | Morus022525.p1 | Calmodulin | 9 | 250 | 16,894 | signaling | 1.03 | 1.12 | 1.31 |
| 12 | Morus015082.p1 | Auxin-repressed 12.5 kDa protein | 3 | 89 | 13,355 | development | 0.49 | 1.99 | 1.30 |
| 13 | Morus003616.p1 | Fructokinase-2 | 10 | 137 | 35,370 | major CHO metabolism | 0.42 | 0.79 | 1.25 |
| 14 | Morus017382.p1 | Calcium-binding protein CML27 | 4 | 116 | 18,705 | signaling | 0.26 | 0.29 | 1.05 |
| 15 | Morus008669.p1 | Allene oxide cyclase 2, chloroplastic | 6 | 168 | 27,569 | hormone metabolism | 0.60 | 1.24 | 0.99 |
| 16 | Morus010743.p1 | Triosephosphate isomerase, cytosolic | 19 | 263 | 27,548 | glycolysis | 1.27 | 1.66 | 0.99 |
| 17 | Morus004210.p1 | Glucan endo-1,3-beta-glucosidase, basic vacuolar isoform | 20 | 178 | 39,002 | misc | 0.63 | 0.34 | 0.94 |
| 18 | Morus011779.p1 | Superoxide dismutase [Cu-Zn], chloroplastic | 22 | 579 | 29,603 | redox | 0.74 | 0.31 | 0.90 |
| 19 | Morus001936.p1 | Peroxiredoxin-2B | 17 | 350 | 17,391 | redox | 1.22 | 1.48 | 0.90 |
| 20 | Morus004201.p1 | Universal stress protein A-like protein | 9 | 223 | 18,591 | stress | 0.58 | 0.58 | 0.82 |
| 21 | Morus001634.p1 | Nucleoside diphosphate kinase 1 | 11 | 215 | 16,322 | nucleotide metabolism | 1.35 | 0.78 | 0.78 |
| 22 | Morus003013.p1 | Phosphoglycerate kinase, cytosolic | 59 | 1143 | 42,729 | glycolysis | 1.72 | 2.17 | 0.75 |
| 23 | Morus028068.p1 | Polygalacturonase inhibitor 1 | 10 | 306 | 37,677 | cell wall | 0.86 | 0.73 | 0.73 |
| 24 | Morus013807.p1 | Fructose-bisphosphate aldolase, cytoplasmic isozyme | 23 | 584 | 38,459 | glycolysis | 0.92 | 1.82 | 0.71 |
| 25 | Morus020532.p1 | Glutaredoxin | 5 | 244 | 15,307 | redox | 0.75 | 0.69 | 0.69 |
| 26 | Morus023908.p1 | Uncharacterized protein | 15 | 271 | 57,888 | protein | 0.50 | 1.55 | 0.69 |
| 27 | Morus018475.p1 | Peroxidase 54 | 5 | 128 | 36,921 | misc | 0.13 | 0.47 | 0.67 |
| 28 | Morus025517.p1 | Tubulin alpha chain | 18 | 426 | 49,920 | cell | 0.68 | 0.72 | 0.66 |
| 29 | Morus002489.p1 | Nascent polypeptide-associated complex subunit alpha-like protein 1 | 11 | 285 | 22,279 | protein | 1.01 | 0.34 | 0.65 |
| 30 | Morus014304.p1 | Plastocyanin, chloroplastic | 23 | 661 | 16,620 | photosynthesis | 1.16 | 0.62 | 0.62 |
| 31 | Morus024265.p1 | Aquaporin PIP1-3 | 5 | 67 | 30,856 | transport | 0.22 | 0.37 | 0.59 |
| 32 | Morus008884.p1 | Cysteine proteinase RD21a | 10 | 350 | 52,217 | protein | 0.35 | 0.32 | 0.58 |
| 33 | Morus025862.p1 | ATP synthase subunit beta, mitochondrial | 33 | 1017 | 59,400 | mitochondrial electron transport | 1.06 | 1.43 | 0.54 |
| 34 | Morus026982.p1 | Allene oxide synthase, chloroplastic | 11 | 164 | 56,861 | hormone metabolism | 0.30 | 0.91 | 0.54 |
| 35 | Morus007342.p1 | Peroxiredoxin-2F, mitochondrial | 12 | 178 | 22,580 | redox | 0.77 | 1.03 | 0.52 |
| 36 | Morus009738.p1 | ATP-dependent Clp protease proteolytic subunit 5, chloroplastic | 4 | 146 | 34,203 | protein | 0.13 | 0.21 | 0.52 |
| 37 | Morus009210.p1 | 60S acidic ribosomal protein P3-2 | 2 | 71 | 12,022 | protein | 0.42 | 0.48 | 0.48 |
| 38 | Morus007901.p1 | Actin-7 | 30 | 717 | 41,897 | cell | 1.02 | 1.20 | 0.47 |
| 39 | Morus007352.p1 | Stem-specific protein TSJT1 | 7 | 96 | 25,521 | metal handling | 0.18 | 0.45 | 0.45 |
| 40 | Morus026327.p1 | Heat shock cognate 70 kDa protein 1 | 41 | 791 | 71,553 | stress | 0.91 | 0.67 | 0.44 |
| 41 | Morus021433.p1 | Malate dehydrogenase, cytoplasmic | 20 | 463 | 35,912 | TCA | 0.57 | 0.95 | 0.43 |
| 42 | Morus006184.p1 | Cysteine synthase | 33 | 572 | 34,400 | amino acid metabolism | 1.77 | 0.59 | 0.39 |
| 43 | Morus018842.p1 | 2-Cys peroxiredoxin BAS1-like, chloroplastic | 23 | 321 | 29,121 | redox | 0.81 | 0.65 | 0.39 |
| 44 | Morus022454.p1 | Fasciclin-like arabinogalactan protein 8 | 6 | 120 | 43,455 | cell wall | 0.22 | 0.16 | 0.39 |
| 45 | Morus008883.p1 | Uncharacterized protein | 6 | 104 | 49,487 | signaling | 0.17 | 0.34 | 0.38 |
| 46 | Morus000210.p1 | Calvin cycle protein CP12 | 8 | 288 | 14,542 | photosynthesis | 0.34 | 0.38 | 0.38 |
| 47 | Morus002920.p1 | Thioredoxin M-type 4, chloroplastic | 6 | 164 | 20,233 | redox | 0.42 | 0.26 | 0.37 |
| 48 | Morus018564.p1 | Isoflavone reductase homolog P3 | 12 | 102 | 45,171 | secondary metabolism | 0.33 | 1.17 | 0.37 |
| 49 | Morus013051.p1 | Adenosine kinase 2 | 7 | 216 | 37,797 | nucleotide metabolism | 0.65 | 0.41 | 0.35 |
| 50 | Morus000836.p1 | Ribulose bisphosphate carboxylase large chain (Fragment) | 290 | 4805 | 61,599 | photosynthesis | 1.18 | 0.63 | 0.33 |
| 51 | Morus015202.p1 | Uncharacterized protein | 11 | 234 | 33,994 | not assigned | 0.46 | 0.39 | 0.32 |
| 52 | Morus014140.p1 | Plastid-lipid-associated protein, chloroplastic | 10 | 281 | 35,137 | cell | 0.52 | 0.31 | 0.31 |
| 53 | Morus025784.p1 | Phospholipase D alpha 1 | 4 | 47 | 92,059 | lipid metabolism | 0.05 | 0.37 | 0.30 |
| 54 | Morus018550.p1 | Glycine-rich RNA-binding protein GRP1A | 5 | 165 | 18,416 | RNA | 0.39 | 0.29 | 0.29 |
| 55 | Morus014011.p1 | Glycerophosphoryl diester phosphodiesterase 2 | 6 | 295 | 81,816 | lipid metabolism | 0.16 | 0.12 | 0.29 |
| 56 | Morus019087.p1 | Putative mitochondrial 2-oxoglutarate/malate carrier protein | 12 | 194 | 32,224 | transport | 0.98 | 0.29 | 0.28 |
| 57 | Morus002874.p1 | Leucine aminopeptidase 3, chloroplastic | 39 | 981 | 60,563 | protein | 0.96 | 1.09 | 0.27 |
| 58 | Morus010230.p1 | Superoxide dismutase [Cu-Zn] | 2 | 63 | 20,420 | redox | 0.34 | 0.37 | 0.26 |
| 59 | Morus019413.p1 | Cysteine proteinase 15A | 9 | 259 | 41,574 | protein | 0.23 | 0.12 | 0.26 |
| 60 | Morus015818.p1 | Probable glucan endo-1,3-beta-glucosidase A6 | 3 | 111 | 52,145 | misc | 0.15 | 0.17 | 0.24 |
| 61 | Morus020384.p1 | Cysteine synthase, chloroplastic/chromoplastic | 21 | 258 | 43,997 | amino acid metabolism | 0.52 | 0.11 | 0.24 |
| 62 | Morus013361.p1 | Protein disulfide-isomerase | 15 | 432 | 56,492 | redox | 0.66 | 0.92 | 0.22 |
| 63 | Morus007114.p1 | Glycine-rich RNA-binding protein 2 | 3 | 170 | 27,802 | RNA | 0.17 | 0.19 | 0.19 |
| 64 | Morus011198.p1 | L-ascorbate peroxidase, cytosolic | 26 | 392 | 27,414 | redox | 1.05 | 1.13 | 0.19 |
| 65 | Morus008123.p1 | IAA-amino acid hydrolase ILR1-like 5 | 5 | 89 | 47,707 | hormone metabolism | 0.15 | 0.50 | 0.18 |
| 66 | Morus024851.p1 | Catalase isozyme 1 | 19 | 137 | 57,208 | redox | 0.62 | 0.32 | 0.18 |
| 67 | Morus017351.p1 | Serine carboxypeptidase-like 50 | 5 | 55 | 49,604 | protein | 0.15 | 0.14 | 0.17 |
| 68 | Morus014667.p1 | Alpha-xylosidase | 15 | 257 | 103,539 | misc | 0.17 | 0.45 | 0.15 |
| 69 | Morus017174.p1 | Predicted protein | 8 | 193 | 33,060 | signaling | 0.20 | 0.27 | 0.15 |
| 70 | Morus024951.p1 | Triosephosphate isomerase, chloroplastic | 42 | 655 | 34,813 | photosynthesis | 1.66 | 0.99 | 0.15 |
| 71 | Morus008661.p1 | 14-3-3-like protein A | 15 | 283 | 81,889 | cell | 0.26 | 0.37 | 0.14 |
| 72 | Morus016271.p1 | Elongation factor 2 | 14 | 106 | 99,403 | protein | 0.11 | 0.44 | 0.14 |
| 73 | Morus001657.p1 | 6-phosphogluconolactonase 4, chloroplastic | 6 | 108 | 35,151 | OPP | 0.18 | 0.97 | 0.14 |
| 74 | Morus009365.p1 | 5-methyltetrahydropteroyltriglutamate--homocysteine methyltransferase | 13 | 188 | 84,904 | amino acid metabolism | 0.17 | 0.46 | 0.14 |
| 75 | Morus007784.p1 | UTP--glucose-1-phosphate uridylyltransferase | 26 | 372 | 76,133 | glycolysis | 0.48 | 0.80 | 0.13 |
| 76 | Morus017695.p1 | 31 kDa ribonucleoprotein, chloroplastic | 10 | 230 | 38,128 | RNA | 0.17 | 0.13 | 0.13 |
| 77 | Morus007494.p1 | RuBisCO large subunit-binding protein subunit alpha, chloroplastic | 30 | 964 | 62,000 | photosynthesis | 1.29 | 0.33 | 0.11 |
| 78 | Morus011664.p1 | L-ascorbate oxidase homolog | 4 | 53 | 60,522 | not assigned | 0.13 | 0.37 | 0.11 |
| 79 | Morus006060.p1 | V-type proton ATPase subunit B2 | 10 | 400 | 63,333 | transport | 0.41 | 0.16 | 0.11 |
| 80 | Morus004111.p1 | Calreticulin | 7 | 266 | 50,196 | signaling | 0.32 | 0.56 | 0.10 |
| 81 | Morus013778.p1 | Monodehydroascorbate reductase | 13 | 142 | 49,982 | redox | 0.30 | 0.33 | 0.10 |
| 82 | Morus024141.p1 | Beta-D-xylosidase 4 | 6 | 187 | 84,604 | cell wall | 0.14 | 0.10 | 0.10 |
| 83 | Morus007961.p1 | Hypothetical protein | 7 | 62 | 95,561 | not assigned | 0.05 | 0.07 | 0.09 |
| 84 | Morus025925.p1 | Alpha-glucosidase | 6 | 207 | 93,365 | misc | 0.12 | 0.25 | 0.07 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, W.; Zhong, Z.; Liu, S.; Yang, B.; Komatsu, S.; Ge, Z.; Tian, J. Organ-Specific Analysis of Morus alba Using a Gel-Free/Label-Free Proteomic Technique. Int. J. Mol. Sci. 2019, 20, 365. https://doi.org/10.3390/ijms20020365
Zhu W, Zhong Z, Liu S, Yang B, Komatsu S, Ge Z, Tian J. Organ-Specific Analysis of Morus alba Using a Gel-Free/Label-Free Proteomic Technique. International Journal of Molecular Sciences. 2019; 20(2):365. https://doi.org/10.3390/ijms20020365
Chicago/Turabian StyleZhu, Wei, Zhuoheng Zhong, Shengzhi Liu, Bingxian Yang, Setsuko Komatsu, Zhiwei Ge, and Jingkui Tian. 2019. "Organ-Specific Analysis of Morus alba Using a Gel-Free/Label-Free Proteomic Technique" International Journal of Molecular Sciences 20, no. 2: 365. https://doi.org/10.3390/ijms20020365
APA StyleZhu, W., Zhong, Z., Liu, S., Yang, B., Komatsu, S., Ge, Z., & Tian, J. (2019). Organ-Specific Analysis of Morus alba Using a Gel-Free/Label-Free Proteomic Technique. International Journal of Molecular Sciences, 20(2), 365. https://doi.org/10.3390/ijms20020365