Endometrial Immune Dysfunction in Recurrent Pregnancy Loss



Abstract
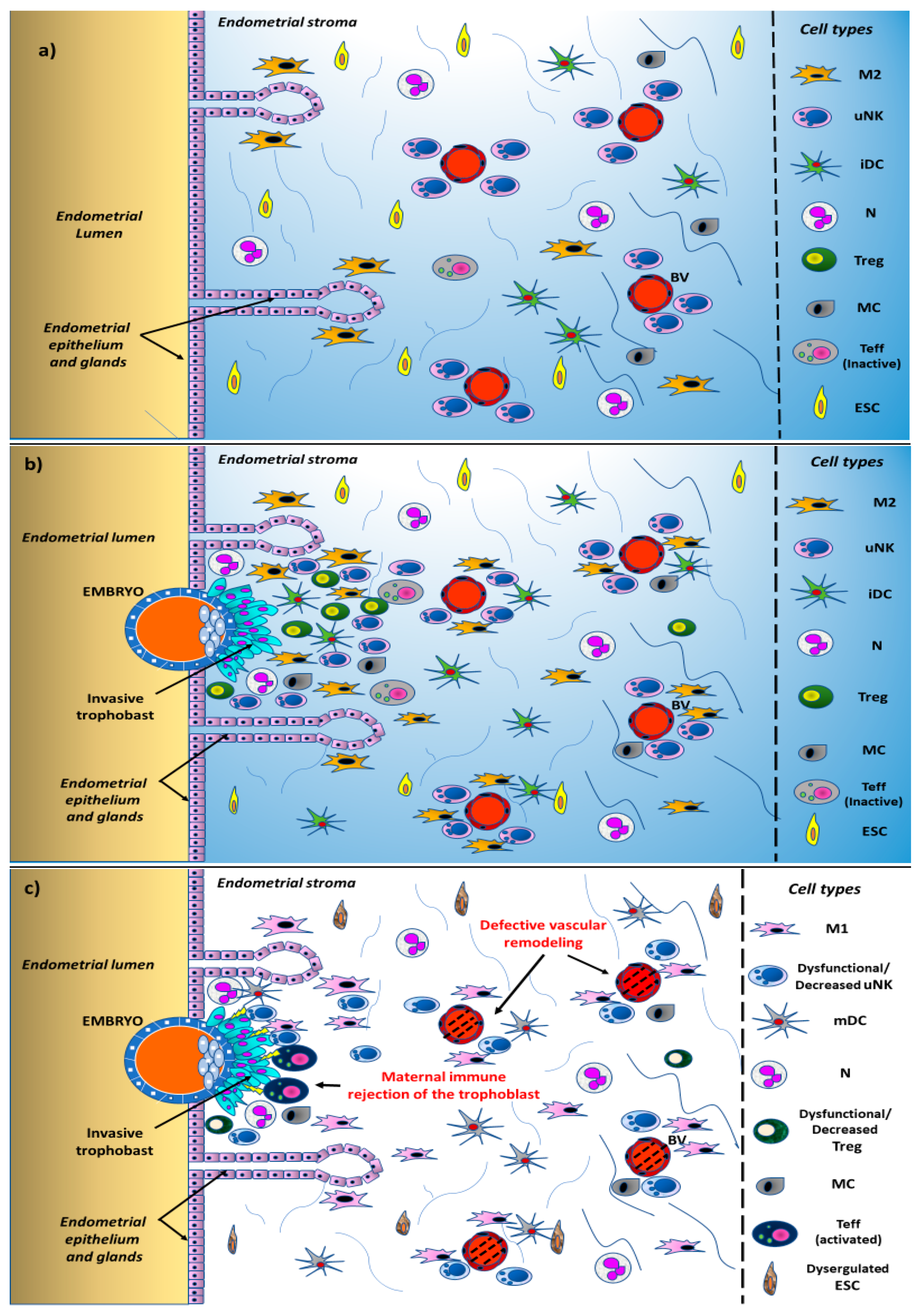
:1. Introduction
2. Physiological Endometrial Function and the Immune System
2.1. Endometrial Remodeling and Decidualization
2.2. Immune Cells and Their Functions in the Normal Endometrium and Decidua
2.2.1. Macrophages
2.2.2. NK Cells
2.2.3. DCs
2.2.4. Tregs
2.2.5. Other Components of the Immune System
2.3. Cytokine/Chemokine Network and the Maternal–Fetal Immune Cross-Talk
3. Immune Dysregulation in the Endometrium and Decidua in RPL
3.1. Endometrial Cells in RPL with Specific Application to the Regulation of the Local Immune Function
3.2. Immune Cells in the Endometrium in RPL
3.2.1. Macrophages
3.2.2. uNKs
3.2.3. Uterine Dendritic Cells (uDCs)
3.2.4. Tregs
3.3. Endometrial Cytokine Imbalance in RPL
4. Conclusions and Future Directions
Author Contributions
Funding
Conflicts of Interest
References
- Recurrent Pregnancy Loss: Guideline of the European Society of Human Reproduction and Embryology. ESHRE Early Pregnancy Guidline Development Group. pp. 1–153. Available online: https://www.eshre.eu/Guidelines-and-Legal/Guidelines/Recurrent-pregnancy-loss.aspx (accessed on 26 November 2017).
- Practice Committee of the American Society for Reproductive Medicine. Definitions of infertility and Recurrent Pregnancy Loss: A Committee Opinion. Fertil Steril. 2013, 99, 63. [Google Scholar] [CrossRef] [PubMed]
- El Hachem, H.; Crepaux, V.; May-Panloup, P.; Descamps, P.; Legendre, G.; Bouet, P.E. Recurrent Pregnancy Loss: Current Perspectives. Int. J. Womens Health 2017, 9, 331–345. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Gimenez, C.; Alijotas-Reig, J. Recurrent Miscarriage: Causes, Evaluation and Management. Postgrad. Med. J. 2015, 91, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Jeve, Y.B.; Davies, W. Evidence-Based Management of Recurrent Miscarriages. J. Hum. Reprod. Sci. 2014, 7, 159–169. [Google Scholar] [CrossRef]
- Wang, N.F.; Kolte, A.M.; Larsen, E.C.; Nielsen, H.S.; Christiansen, O.B. Immunologic Abnormalities, Treatments, and Recurrent Pregnancy Loss: What Is Real and What Is Not? Clin. Obstet Gynecol. 2016, 59, 509–523. [Google Scholar] [CrossRef]
- Gleicher, N.; Kushnir, V.A.; Barad, D.H. Redirecting Reproductive Immunology Research Toward Pregnancy as a Period of Temporary Immune Tolerance. J. Assist. Reprod. Genet. 2017, 34, 425–430. [Google Scholar] [CrossRef]
- Bazer, F.W.; Spencer, T.E.; Johnson, G.A.; Burghardt, C.; Wu, G. Comparative Aspects of Implantation. Reproduction 2009, 138, 195–209. [Google Scholar] [CrossRef]
- Gellersen, B.; Brosens, J.J. Cyclic Decidualization of the Human Endometrium in Reproductive Health and Failure. Endocr. Rev. 2014, 35, 851–905. [Google Scholar] [CrossRef]
- Okada, H.; Tsuzuki, T.; Murata, H. Decidualization of the Human Endometrium. Reprod. Med. Biol. 2018, 17, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Makieva, S.; Giacomini, E.; Ottolina, J.; Sanchez, A.M.; Papaleo, E.; Vigano, P. Inside the Endometrial Cell Signaling Subway: Mind the Gap(s). Int. J. Mol. Sci. 2018, 19, 2477. [Google Scholar] [CrossRef]
- Su, R.W.; Fazleabas, A.T. Implantation and Establishment of Pregnancy in Human and Nonhuman Primates. Adv. Anat. Embryol. Cell. Biol. 2015, 216, 189–213. [Google Scholar] [PubMed] [Green Version]
- Ewington, L.J.; Tewary, S.; Brosens, J.J. New Insights into the Mechanisms Underlying Recurrent Pregnancy Loss. J. Obstet. Gynaecol. Res. 2019, 45, 258–265. [Google Scholar] [CrossRef] [PubMed]
- Emera, D.; Romero, R.; Wagner, G. The Evolution of Menstruation: A New Model for Genetic Assimilation: Explaining Molecular Origins of Maternal Responses to Fetal Invasiveness. Bioessays 2012, 34, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Lucas, E.S.; Dyer, N.P.; Fishwick, K.; Ott, S.; Brosens, J.J. Success after Failure: The Role of Endometrial Stem Cells in Recurrent Miscarriage. Reproduction 2016, 152, R159–R166. [Google Scholar] [CrossRef]
- Brosens, J.J.; Salker, M.S.; Teklenburg, G.; Nautiyal, J.; Salter, S.; Lucas, E.S.; Steel, J.H.; Christian, M.; Chan, Y.W.; Boomsma, C.M.; et al. Uterine Selection of Human Embryos at Implantation. Sci. Rep. 2014, 4, 3894. [Google Scholar] [CrossRef]
- Macklon, N.S.; Brosens, J.J. The Human Endometrium as a Sensor of Embryo Quality. Biol. Reprod. 2014, 91, 98. [Google Scholar] [CrossRef]
- Quenby, S.; Brosens, J.J. Human Implantation: A tale of Mutual Maternal and Fetal Attraction. Biol. Reprod. 2013, 88, 81. [Google Scholar] [CrossRef]
- Lucas, E.S.; Dyer, N.P.; Murakami, K.; Lee, Y.H.; Chan, Y.W.; Grimaldi, G.; Muter, J.; Brighton, P.J.; Moore, J.D.; Patel, G.; et al. Loss of Endometrial Plasticity in Recurrent Pregnancy Loss. Stem. Cells 2016, 34, 346–356. [Google Scholar] [CrossRef]
- Zenclussen, A.C.; Hammerling, G.J. Cellular Regulation of the Uterine Microenvironment That Enables Embryo Implantation. Front. Immunol. 2015, 6, 321. [Google Scholar] [CrossRef]
- Schumacher, A.; Sharkey, D.J.; Robertson, S.A.; Zenclussen, A.C. Immune Cells at the Fetomaternal Interface: How the Microenvironment Modulates Immune Cells To Foster Fetal Development. J. Immunol. 2018, 201, 325–334. [Google Scholar] [CrossRef]
- Racicot, K.; Kwon, J.Y.; Aldo, P.; Silasi, M.; Mor, G. Understanding the Complexity of the Immune System During Pregnancy. Am. J. Reprod. Immunol. 2014, 72, 107–116. [Google Scholar] [CrossRef] [PubMed]
- PrabhuDas, M.; Bonney, E.; Caron, K.; Dey, S.; Erlebacher, A.; Fazleabas, A.; Fisher, S.; Golos, T.; Matzuk, M.; McCune, J.M.; et al. Immune Mechanisms at the Maternal-Fetal Interface: Perspectives and Challenges. Nat. Immunol. 2015, 16, 328–334. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.K.; Kim, C.J.; Kim, D.J.; Kang, J.H. Immune Cells in the Female Reproductive Tract. Immune. Netw. 2015, 15, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Mori, M.; Bogdan, A.; Balassa, T.; Csabai, T.; Szekeres-Bartho, J. The Decidua-The Maternal Bed Embracing the Embryo-Maintains the Pregnancy. In Seminars and Immunopathol; Springer: Berlin, Germany, 2016; Volume 38, pp. 635–649. [Google Scholar]
- Drury, J.A.; Parkin, K.L.; Coyne, L.; Giuliani, E.; Fazleabas, A.T.; Hapangama, D.K. The Dynamic Changes in the Number of Uterine Natural Killer Cells are Specific to the Eutopic but not to the Ectopic Endometrium in Women and in a Baboon Model of Endometriosis. Reprod. Biol. Endocrinol. 2018, 16, 67. [Google Scholar] [CrossRef] [PubMed]
- Manaster, I.; Mizrahi, S.; Goldman-Wohl, D.; Sela, H.Y.; Stern-Ginossar, N.; Lankry, D.; Gruda, R.; Hurwitz, A.; Bdolah, Y.; Haimov-Kochman, R.; et al. Endometrial NK Cells are Special Immature Cells that Await Pregnancy. J. Immunol. 2008, 181, 1869–1876. [Google Scholar] [CrossRef] [PubMed]
- Fukui, A.; Funamizu, A.; Fukuhara, R.; Shibahara, H. Expression of Natural Cytotoxicity Receptors and Cytokine Production on Endometrial Natural Killer Cells in Women with Recurrent Pregnancy Loss or Implantation Failure, and the Expression of Natural Cytotoxicity Receptors on Peripheral Blood Natural Killer Cells in Pregnant Women with a History of Recurrent Pregnancy Loss. J. Obstet. Gynaecol. Res. 2017, 43, 1678–1686. [Google Scholar]
- Lee, J.Y.; Lee, M.; Lee, S.K. Role of Endometrial Immune Cells in Implantation. Clin. Exp. Reprod. Med. 2011, 38, 119–125. [Google Scholar] [CrossRef]
- Faas, M.M.; De Vos, P. Innate Immune Cells in the Placental Bed in Healthy Pregnancy and Preeclampsia. Placenta 2018, 69, 125–133. [Google Scholar] [CrossRef]
- Tsao, F.Y.; Wu, M.Y.; Chang, Y.L.; Wu, C.T.; Ho, H.N. M1 Macrophages Decrease in the Deciduae from Normal Pregnancies but not from Spontaneous Abortions or Unexplained Recurrent Spontaneous Abortions. J. Formos. Med. Assoc. 2018, 117, 204–211. [Google Scholar] [CrossRef]
- Erlebacher, A. Immunology of the Maternal-Fetal Interface. Annu. Rev. Immunol. 2013, 31, 387–411. [Google Scholar] [CrossRef]
- Robertson, S.A.; Moldenhauer, L.M. Immunological Determinants of Implantation Success. Int. J. Dev. Biol. 2014, 58, 205–217. [Google Scholar] [CrossRef] [PubMed]
- Care, A.S.; Diener, K.R.; Jasper, M.J.; Brown, H.M.; Ingman, W.V.; Robertson, S.A. Macrophages Regulate Corpus Luteum Development During Embryo Implantation in Mice. J. Clin. Invest. 2013, 123, 3472–3487. [Google Scholar] [CrossRef] [PubMed]
- Renaud, S.J.; Graham, C.H. The Role of Macrophages in Utero-Placental Interactions During Normal and Pathological Pregnancy. Immunol. Invest. 2008, 37, 535–564. [Google Scholar] [CrossRef] [PubMed]
- Jeziorska, M.; Salamonsen, L.A.; Woolley, D.E. Mast Cell and Eosinophil Distribution and Activation in Human Endometrium Throughout the Menstrual Cycle. Biol. Reprod. 1995, 53, 312–320. [Google Scholar] [CrossRef]
- De Leo, B.; Esnal-Zufiaurre, A.; Collins, F.; Critchley, H.O.D.; Saunders, P.T.K. Immunoprofiling of Human Uterine Mast Cells Identifies Three Phenotypes and Expression of ERβ and Glucocorticoid Receptor. F1000Res. 2017, 6, 667. [Google Scholar] [CrossRef]
- King, A. Uterine Leukocytes and Decidualization. Hum. Reprod. Update 2000, 6, 28–36. [Google Scholar] [CrossRef]
- Schulke, L.; Manconi, F.; Markham, R.; Fraser, I.S. Endometrial Dendritic Cell Populations During the Normal Menstrual Cycle. Hum. Reprod. 2008, 23, 1574–1580. [Google Scholar] [CrossRef]
- Ramhorst, R.; Grasso, E.; Paparini, D.; Hauk, V.; Gallino, L.; Calo, G.; Vota, D.; Perez Leiros, C. Decoding the Chemokine Network that Links Leukocytes with Decidual Cells and the Trophoblast During Early Implantation. Cell Adh. Migr. 2016, 10, 197–207. [Google Scholar] [CrossRef]
- Rieger, L.; Honig, A.; Sutterlin, M.; Kapp, M.; Dietl, J.; Ruck, P.; Kammerer, U. Antigen-Presenting Cells in Human Endometrium During the Menstrual Cycle Compared to Early Pregnancy. J. Soc. Gynecol. Invest. 2004, 11, 488–493. [Google Scholar] [CrossRef]
- Maridas, D.E.; Hey-Cunningham, A.J.; Ng, C.H.M.; Markham, R.; Fraser, I.S.; Berbic, M. Peripheral and Endometrial Dendritic Cell Populations During the Normal Cycle and in the Presence of Endometriosis. J. Endometr. Pelvic Pain Disord. 2014, 6, 67–119. [Google Scholar] [CrossRef]
- Zenclussen, A.C.; Olivieri, D.N.; Dustin, M.L.; Tadokoro, C.E. In Vivo Multiphoton Microscopy Technique to Reveal the Physiology of the Mouse Uterus. Am. J. Reprod. Immunol. 2013, 69, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Nadkarni, S.; Smith, J.; Sferruzzi-Perri, A.N.; Ledwozyw, A.; Kishore, M.; Haas, R.; Mauro, C.; Williams, D.J.; Farsky, S.H.P.; Marelli-Berg, F.M.; et al. Neutrophils Induce Proangiogenic T Cells with a Regulatory Phenotype in Pregnancy. Proc. Natl. Acad. Sci. USA 2016, 113, E8415–E8424. [Google Scholar] [CrossRef] [PubMed]
- King, A.E.; Critchley, H.O.D.; Kelly, R.W. Innate Immune Defences in the Human Endometrium. Reprod. Biol. Endocrinol. 2003, 1, 116. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, G.M.; Maybin, J.A.; Murray, A.A.; Nicol, M.; Walker, C.; Saunders, P.T.K.; Rossi, A.G.; Critchley, H.O.D. Endometrial Apoptosis and Neutrophil Infiltration During Menstruation Exhibits Spatial and Temporal Dynamics that are Recapitulated in a Mouse Model. Sci. Rep. 2017, 12, 17416. [Google Scholar] [CrossRef] [PubMed]
- Salamonsen, L.A.; Lathbury, L.J. Endometrial Leukocytes and Menstruation. Hum. Reprod. Update. 2000, 6, 16–27. [Google Scholar] [CrossRef]
- Harrity, C.; Bereir, M.M.; Walsh, D.J.; Marron, K.D. Moving from Peripheral Blood to Local Uterine Immunophenotype Analysis in Patients with Poor Reproductive History: Pilot Study of a Novel Technique. Ir. J. Med. Sci. 2019, 188, 893–901. [Google Scholar] [CrossRef]
- Flynn, L.; Byrne, B.; Carton, J.; Kelehan, P.; O’Herlihy, C.; O’Farrelly, C. Menstrual Cycle Dependent Fluctuations in NK and T-Lymphocyte Subsets from Non-Pregnant Human Endometrium. Am. J. Reprod. Immunol. 2000, 43, 209–217. [Google Scholar] [CrossRef]
- Chang, H.F.; Bzeih, H.; Chitirala, P.; Ravichandran, K.; Sleiman, M.; Krause, E.; Hahn, U.; Pattu, V.; Rettig, J. Preparing the Lethal Hit: Interplay Between Exo-And Endocytic Pathways in Cytotoxic T Lymphocytes. Cell Mol. Life Sci. 2017, 74, 399–408. [Google Scholar] [CrossRef]
- Vanderstraeten, A.; Tuyaerts, S.; Amant, F. The Immune System in the Normal Endometrium and Implications for Endometrial Cancer Development. J. Reprod. Immunol. 2015, 109, 7–16. [Google Scholar] [CrossRef]
- Zhao, X.; Jiang, Y.; Wang, L.; Li, Z.; Li, Q.; Feng, X. Advances in Understanding the Immune Imbalance between T-Lymphocyte Subsets and NK Cells in Recurrent Spontaneous Abortion. Geburtshilfe Frauenheilkd 2018, 78, 677–683. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, Y.; Sakai, M.; Miyazaki, S.; Higuma, S.; Shiozaki, A.; Saito, S. Decidual and Peripheral Blood CD4+CD25+ Regulatory T Cells in Early Pregnancy Subjects and Spontaneous Abortion Cases. Mol. Hum. Reprod. 2004, 10, 347–353. [Google Scholar] [CrossRef] [PubMed]
- Roberton, S.A.; Care, A.S.; Moldenhauer, L.M. Regulatory T Cells in Embryo Implantation and the Immune Response to Pregnancy. J. Clin. Invest. 2018, 128, 4224–4235. [Google Scholar] [CrossRef] [PubMed]
- Alijotas-Reig, J.; Llurba, E.; Gris, J.M. Potentiating Maternal Immune Tolerance in Pregnancy: A New Challenging Role for Regulatory T Cells. Placenta 2014, 35, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Arruvito, L.; Sanz, M.; Banham, A.H.; Fainboim, L. Expansion of CD4+CD25+and FOXP3+Regulatory T Cells During the Follicular Phase of the Menstrual Cycle: Implications for Human Reproduction. J. Immunol. 2007, 178, 2572–2578. [Google Scholar] [CrossRef]
- Kofod, L.; Lindhard, A.; Hviid, T.V.F. Implications of Uterine NK Cells and Regulatory T Cells in the Endometrium of Infertile Women. Hum. Immunol. 2018, 79, 693–701. [Google Scholar] [CrossRef]
- Jasper, M.J.; Care, A.S.; Sulivan, B.; Ingman, W.V.; Aplin, J.D.; Robertson, S.A. Macrophage-Derived LIF and IL1B Regulate Alpha(1,2)Fucosyltransferase 2 (Fut2) Expression in Mouse Uterine Epithelial Cells During Early Pregnancy. Biol. Reprod. 2011, 84, 179–188. [Google Scholar] [CrossRef]
- Co, E.C.; Gormley, M.; Kapidzic, M.; Rosen, D.B.; Scott, M.A.; Stolp, H.A.; McMaster, M.; Lanier, L.L.; Barcena, A.; Fisher, S.J. Maternal Decidual Macrophages Inhibit NK Cell Killing of Invasive Cytotrophoblasts During Human Pregnancy. Biol. Reprod. 2013, 88, 155. [Google Scholar] [CrossRef]
- Manaster, I.; Mandelboim, O. The Unique Properties of Uterine NK Cells. Am. J. Reprod. Immunol. 2010, 63, 434–444. [Google Scholar] [CrossRef]
- Fu, B.; Li, X.; Sun, R.; Tong, X.; Ling, B.; Tian, Z.; Wei, H. Natural Killer Cells Promote Immune Tolerance by Regulating Inflammatory TH17 Cells at the Human Maternal-Fetal Interface. Proc. Natl. Acad. Sci. USA 2013, 110, E231–E240. [Google Scholar] [CrossRef]
- Kopcow, H.D.; Rosetti, F.; Leung, Y.; Allan, D.S.; Kutok, J.L.; Strominger, J.L. T Cell Apoptosis at the Maternal-Fetal Interface in Early Human Pregnancy, Involvement of Galectin-1. Proc. Natl. Acad. Sci. USA 2008, 105, 18472–18477. [Google Scholar] [CrossRef]
- Vacca, P.; Cantoni, C.; Vitale, M.; Prato, C.; Canegallo, F.; Fenoglio, D.; Ragni, N.; Moretta, L.; Mingari, M.C. Crosstalk Between Decidual NK and CD14+Myelomonocytic Cells Results in Induction of Tregs and Immunosuppression. Proc. Natl. Acad. Sci. USA 2010, 107, 11918–11923. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.H.; Tang, M.X.; Mor, G.; Liao, A.H. Tim-3: Expression on Immune Cells and Roles at the Maternal-Fetal Interface. J. Reprod. Immunol. 2016, 118, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Teles, A.; Schumacher, A.; Kuhnle, M.C.; Linzke, N.; Thuere, C.; Reichardt, P.; Tadokoro, C.E.; Hammerling, G.J.; Zenclussen, A.C. Control of Uterine Microenvironment by foxp3(+) Cells Facilitates Embryo Implantation. Front. Immunol. 2013, 20, 158. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, H.; Araki, Y.; Imakawa, K.; Saito, S.; Daikoku, T.; Shigeta, M.; Kanzaki, H.; Mori, T. Dual Positive Regulation of Embryo Implantation by Endocrine and Immune Systems-Step-by-Step Maternal Recognition of the Developing Embryo. Am. J. Reprod. Immunol. 2016, 75, 281–289. [Google Scholar] [CrossRef]
- Schjenken, J.E.; Robertson, S.A. Seminal Fluid and Immune Adaptation for Pregnancy-Comparative Biology in Mammalian Species. Reprod. Domest. Anim. 2014, 49, 27–36. [Google Scholar] [CrossRef]
- Hsu, P.; Santner-Nanan, B.; Dahlstrom, J.E.; Fadia, M.; Chandra, A.; Peek, M.; Nanan, R. Altered Decidual DC-SIGN+Antigen-Presenting Cells and Impaired Regulatory T-Cell Induction in Preeclampsia. Am. J. Pathol. 2012, 181, 2149–2160. [Google Scholar] [CrossRef]
- Blois, S.M.; Kammerer, U.; Alba Soto, C.; Tometten, M.C.; Shaikly, V.; Barrientos, G.; Jurd, R.; Rukavina, D.; Thomson, A.W.; Klapp, B.F.; et al. Dendritic Cells: Key to Fetal Tolerance? Biol. Reprod. 2007, 77, 590–598. [Google Scholar] [CrossRef]
- Liu, S.; Diao, L.; Huang, C.; Li, Y.; Zeng, Y.; Kwak-Kim, J.Y.H. The Role of Decidual Immune Cells on Human Pregnancy. J. Reprod. Immunol. 2017, 124, 44–53. [Google Scholar] [CrossRef]
- Miyazaki, S.; Tsuda, H.; Sakai, M.; Hori, S.; Sasaki, Y.; Futatani, T.; Miyawaki, T.; Saito, S. Predominance of Th2-Promoting Dendritic Cells in Early Human Pregnancy Decidua. J. Leukoc. Biol. 2003, 74, 514–522. [Google Scholar] [CrossRef]
- Kennedy, P.R.; Chazara, O.; Gardner, L.; Ivarsson, M.A.; Farrell, L.E.; Xiong, S.; Hiby, S.E.; Colucci, F.; Sharkey, A.M.; Moffett, A. Activating KIR2DS4 Is Expressed by Uterine NK Cells and Contributes to Successful Pregnancy. J. Immunol. 2016, 197, 4292–4300. [Google Scholar] [CrossRef] [Green Version]
- Sharkey, A.M.; Xiong, S.; Kennedy, P.R.; Gardner, L.; Farrell, L.E.; Chazara, O.; Ivarsson, M.A.; Hiby, S.E.; Colucci, F.; Moffett, A. Tissue-Specific Education of Decidual NK Cells. J. Immunol. 2015, 195, 3026–3032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajagopalan, S.; Long, E.O. HLA-G-Mediated NK Cell Senescence Promotes Vascular Remodeling: Implications for Reproduction. Cell. Mol. Immunol. 2014, 11, 460–466. [Google Scholar] [CrossRef] [PubMed]
- Fournel, S.; Aguerre-Girr, M.; Huc, X.; Lenfant, F.; Alam, A.; Toubert, A.; Bensussan, A.; Le Bouteiller, P. Cutting Edge: Soluble HLA-G1 Triggers CD95/CD95 Ligand-Mediated Apoptosis in Activated CD8+Cells by Interacting with CD8. J. Immunol. 2000, 164, 6100–6104. [Google Scholar] [CrossRef] [PubMed]
- Mincheva-Nilsson, L.; Baranov, V. Placenta-Derived Exosomes and Syncytiotrophoblast Microparticles and Their Role in Human Reproduction: Immune Modulation for Pregnancy Success. Am. J. Reprod. Immunol. 2014, 72, 440–457. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, G.; Gentile, T.; Miranda, S.; Margni, R.A. Asymmetric antibodies: A Protective Arm in Pregnancy. Chem. Immunol. Allergy 2005, 89, 158–168. [Google Scholar] [PubMed]
- Mao, G.; Wang, J.; Kang, Y.; Tai, P.; Wen, J.; Zou, Q.; Li, G.; Ouyang, H.; Xia, G.; Wang, B. Progesterone Increases Systemic and Local Uterine Proportions of CD4+CD25+Treg Cells During Midterm Pregnancy in Mice. Endocrinology 2010, 151, 5477–5488. [Google Scholar] [CrossRef]
- Szekeres-Bartho, J.; Sucurovic, S.; Mulac-Jericevic, B. The Role of Extracellular Vesicles and PIBF in Embryo-Maternal Immune-Interactions. Front. Immunol. 2018, 13, 2890. [Google Scholar] [CrossRef]
- Schumacher, A.; Brachwitz, N.; Sohr, S.; Engeland, K.; Langwisch, S.; Dolaptchieva, M.; Alexander, T.; Taran, A.; Malfertheiner, S.F.; Costa, S.D.; et al. Human Chorionic Gonadotropin Attracts Regulatory T Cells into the Fetal-Maternal Interface During Early Human Pregnancy. J. Immunol. 2009, 182, 5488–5497. [Google Scholar] [CrossRef]
- Faas, M.M.; De Vos, P. Uterine NK Cells and Macrophages in Pregnancy. Placenta 2017, 56, 44–52. [Google Scholar] [CrossRef]
- Houser, B.L.; Tilburgs, T.; Hill, J.; Nicotra, M.L.; Strominger, J.L. Two Unique Human Decidual Macrophage Populations. J. Immunol. 2011, 186, 2633–2642. [Google Scholar] [CrossRef] [Green Version]
- Bulmer, J.N.; Williams, P.J.; Lash, G.E. Immune Cells in the Placental Bed. Int. J. Dev. Biol. 2010, 54, 281–294. [Google Scholar] [CrossRef] [PubMed]
- Djurisic, S.; Hviid, T.V. HLA Class Ib Molecules and Immune Cells in Pregnancy and Preeclampsia. Front. Immunol. 2014, 5, 652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ning, F.; Liu, H.; Lash, G.E. The Role of Decidual Macrophages During Normal and Pathological Pregnancy. Am. J. Reprod. Immunol. 2016, 75, 298–309. [Google Scholar] [CrossRef] [PubMed]
- Craven, C.M.; Morgan, T.; Ward, K. Decidual Spiral Artery Remodelling Begins Before Cellular Interaction with Cytotrophoblasts. Placenta 1998, 19, 241–252. [Google Scholar] [CrossRef]
- Choudhury, R.H.; Dunk, C.E.; Lye, S.J.; Harris, L.K.; Aplin, J.D.; Jones, R.L. Decidual Leucocytes Infiltrating Human Spiral Arterioles are Rich Source of Matrix Metalloproteinases and Degrade Extracellular Matrix in Vitro and in Situ. Am. J. Reprod. Immunol. 2019, 81, e13054. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.D.; Dunk, C.E.; Aplin, J.D.; Harris, L.K.; Jones, R.L. Evidence for Immune Cell Involvement in Decidual Spiral Arteriole Remodeling in Early Human Pregnancy. Am. J. Pathol. 2009, 174, 1959–1971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vivier, E.; Tomasello, E.; Baratin, E.M.; Walzer, T.; Ugolini, S. Functions of Natural Killer Cells. Nat. Immunol. 2008, 9, 503–510. [Google Scholar] [CrossRef]
- Moffett-King, A.; Entrican, G.; Ellis, S.; Hutchinson, J.; Bainbridge, D. Natural Killer Cells and Reproduction. TRENDS Immunol. 2002, 23, 332–333. [Google Scholar] [CrossRef]
- Koopman, L.A.; Kopcow, H.D.; Rybalov, B.; Boyson, J.E.; Orange, J.S.; Schatz, F.; Masch, R.; Lockwood, C.J.; Schachter, A.D.; Park, P.J.; et al. Human Decidual Natural Killer Cells are a Unique NK Cell Subset with Immunomodulatory Potential. J. Exp. Med. 2003, 198, 1201–1212. [Google Scholar] [CrossRef]
- Giuliani, E.; Parkin, K.L.; Lessey, B.A.; Young, S.L.; Fazleabas, A.T. Characterization of Uterine NK Cells in Women with Infertility or Recurrent Pregnancy Loss and Associated Endometriosis. Am. J. Reprod. Immunol. 2014, 72, 262–269. [Google Scholar] [CrossRef] [Green Version]
- Lash, G.E.; Bulmer, J.N.; Li, T.C.; Innes, B.A.; Mariee, N.; Patel, G.; Sanderson, J.; Quenby, S.; Laird, S.M. Standardisation of Uterine Natural Killer (uNK) Cell Measurements in the Endometrium of Women with Recurrent Reproductive Failure. J. Reprod. Immunol. 2016, 116, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Helige, C.; Ahammer, H.; Moser, G.; Hammer, A.; Dohr, G.; Huppertz, B.; Sedlmayr, P. Distribution of Decidual Natural Killer Cells and Macrophages in the Neighbourhood of the Trophoblast Invasion Front: A Quantitative Evaluation. Hum. Reprod. 2014, 29, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chen, Z.; Smith, G.N.; Croy, B.A. Natural Killer Cell-Triggered Vascular Transformation: Maternal Care Before Birth? Cell. Mol. Immunol. 2011, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Moffett, A.; Chazara, O.; Colucci, F. Maternal Allo-Recognition of the Fetus. Fertil. Steril. 2017, 107, 1269–1272. [Google Scholar] [CrossRef] [PubMed]
- Moffett, A. NK Cell Allorecognition. Nat. Rev. Immunol. 2017, 17, 466. [Google Scholar] [CrossRef]
- Penman, B.S.; Moffett, A.; Chazara, O.; Gupta, S.; Parham, P. Reproduction, Infection and Killer-Cell Immunoglobulin-Like Receptor Haplotype Evolution. Immunogenetics 2016, 68, 755–764. [Google Scholar] [CrossRef]
- Arck, P.C.; Hecher, K. Fetomaternal Immune Cross-Talk and its Consequences for Maternal and Offspring’s Health. Nat. Med. 2013, 19, 548–556. [Google Scholar] [CrossRef]
- Robertson, S.A.; Guerin, L.R.; Bromfield, J.J.; Branson, K.M.; Ahlstrom, A.C.; Care, A.S. Seminal Fluid Drives Expansion of the CD4+CD25+T Regulatory Cell Pool and Induces Tolerance to Paternal Alloantigens in Mice. Biol. Reprod. 2009, 80, 1036–1045. [Google Scholar] [CrossRef]
- Zenclussen, M.L.; Thuere, C.; Ahmad, N.; Wafula, P.O.; Fest, S.; Teles, A.; Leber, A.; Casalis, P.A.; Bechmann, I.; Priller, J.; et al. The Persistence of Paternal Antigens in the Maternal Body is Involved in Regulatory T-Cell Expansion and Fetal-Maternal Tolerance in Murine Pregnancy. Am. J. Reprod. Immunol. 2010, 63, 200–208. [Google Scholar] [CrossRef]
- Tagliani, E.; Erlebacher, A. Dendritic Cell Function at the Maternal-Fetal Interface. Exp. Rev. Clin. Immunol. 2011, 7, 593–602. [Google Scholar] [CrossRef]
- Sharma, A.; Rudra, D. Emerging Functions of Regulatory T Cells in Tissue Homeostasis. Front. Immunol. 2018, 9, 883. [Google Scholar] [CrossRef] [PubMed]
- Pereira, L.M.S.; Gomes, S.T.M.; Ishak, R.; Vallinoto, A.C.R. Regulatory T Cell and Forkhead Box Protein 3 as Modulators of Immune Homeostasis. Front. Immunol. 2017, 8, 605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schumacher, A.; Zenclussen, A.C. Regulatory T Cells: Regulators of Life. Am. J. Reprod. Immunol. 2014, 72, 158–170. [Google Scholar] [CrossRef] [PubMed]
- Guerin, L.R.; Prins, J.R.; Robertson, S.A. Regulatory T-Cells and Immune Tolerance in Pregnancy: A New Target for Infertility Treatment? Hum. Reprod. Update. 2009, 15, 517–535. [Google Scholar] [CrossRef] [PubMed]
- Care, A.S.; Bourque, S.L.; Morton, J.S.; Hjartarson, E.P.; Robertson, S.A.; Davidge, S.T. Reduction in Regulatory T Cells in Early Pregnancy Causes Uterine Artery Dysfunction in Mice. Hypertension 2018, 72, 177–187. [Google Scholar] [CrossRef]
- Shima, T.; Sasaky, Y.; Itoh, M.; Nakashima, A.; Ishii, N.; Sugamura, K.; Saito, S. Regulatory T Cells are Necessary for Implantation and Maintenance of Early Pregnancy but not Late Pregnancy in Allogeneic Mice. J. Reprod. Immunol. 2010, 85, 121–129. [Google Scholar] [CrossRef]
- Guzman-Genuino, R.M.; Diener, K.R. Regulatory B Cells in Pregnancy: Lessons from Autoimmunity, Graft Tolerance, and Cancer. Front. Immunol. 2017, 8, 172. [Google Scholar] [CrossRef] [Green Version]
- Rolle, L.; Memarzadeh Tehran, M.; Morell-Garcia, A.; Raeva, Y.; Schumacher, A.; Hartig, R.; Costa, S.D.; Jensen, F.; Zenclussen, A.C. Cutting Edge: il-10-Producing Regulatory B Cells in Early Human Pregnancy. Am. J. Reprod. Immunol. 2013, 70, 448–453. [Google Scholar] [CrossRef]
- Schumacher, A. Human Chorionic Gonadotropin as a Pivotal Endocrine Immune Regulator Initiating and Preserving Fetal Tolerance. Int. J. Mol. Sci. 2017, 18, 2166. [Google Scholar] [CrossRef]
- Geisert, R.; Fazleabas, A.; Lucy, M.; Mathew, D. Interaction of the Conceptus and Endometrium to Establish Pregnancy in Mammals: Role of Interleukin 1β. Cell Tissue Res. 2012, 349, 825–838. [Google Scholar] [CrossRef]
- Rosario, G.X.; Stewart, C.L. The Multifaceted Actions of Leukaemia Inhibitory Factor in Mediating Uterine Receptivity and Embryo Implantation. Am. J. Reprod. Immunol. 2016, 75, 246–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lash, G.E.; Ernerudh, J. Decidual Cytokines and Pregnancy Complications: Focus on Spontaneous Miscarriage. J. Reprod. Immunol. 2015, 108, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Lindau, R.; Mehta, R.B.; Lash, G.E.; Papapavlou, G.; Boij, R.; Berg, G.; Jenmalm, M.C.; Ernerudh, J.; Svensson-Arvelund, J. Interleukin-34 is Present at the Fetal-Maternal Interface and Induces Immunoregulatory Macrophages of a Decidual Phenotype in Vitro. Hum. Reprod. 2018, 33, 588–599. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.Y.; Li, M.J. Interleukin 23 Regulates the Functions of Human Decidual Immune Cells During Early Pregnancy. Biochem. Biophys. Res. Commun. 2016, 469, 340–344. [Google Scholar] [CrossRef] [PubMed]
- Salamonsen, L.A.; Evans, J.; Nguyen, H.P.; Edgell, T.A. The Microenvironment of Human Implantation: Determinant of Reproductive Success. Am. J. Reprod. Immunol. 2016, 75, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Lash, G.E. Molecular Cross-Talk at the Feto-Maternal Interface. Cold Spring Harb. Perspect. Med. 2015, 5, a023010. [Google Scholar] [CrossRef]
- Du, M.R.; Wang, S.C.; Li, D.J. The Integrative Roles of Chemokines at the Maternal-Fetal Interface in Early Pregnancy. Cell. Mol. Immunol. 2014, 11, 438–448. [Google Scholar] [CrossRef]
- Cheng, J.; Rosario, G.; Cohen, T.V.; Hu, J.; Stewart, C.L. Tissue-Specific Ablation of the LIF Receptor in the Murine Uterine Epithelium Results in Implantation Failure. Endocrinology 2017, 158, 1916–1928. [Google Scholar] [CrossRef] [Green Version]
- Hess, A.P.; Hamilton, A.E.; Talbi Dosiou, C.; Nyegaard, M.; Nayak, N.; Genbecev-Krtolica, O.; Mavrogianis, P.; et al. Decidual Stromal Cell Response to Paracrine Signals from the Trophoblast: Amplification of Immune and Angiogenic Modulators. Biol Reprod. 2007, 76, 102–117. [Google Scholar] [CrossRef]
- Salker, M.; Teklenburg, G.; Molokhia, M.; Lavery, S.; Trew, G.; Aojanepong, T.; Mardon, H.J.; Lokugamage, A.U.; Rai, R.; Landles, C.; et al. Natural Selection of Human Embryos: Impaired Decidualization of Endometrium Disables Embryo-Maternal Interactions and Causes Recurrent Pregnancy loss. PLoS ONE 2010, 5, e10287. [Google Scholar] [CrossRef]
- Salker, M.S.; Nautiyal, J.; Steel, J.H.; Webster, Z.; Sucurovic, S.; Nicou, M.; Singh, Y.; Lucas, E.S.; Murakami, K.; Chan, Y.W.; et al. Disordered IL-33/ST2 Activation in Decidualizing Stromal Cells Prolongs Uterine Receptivity in Women with Recurrent Pregnancy Loss. PLoS ONE 2012, 7, e52252. [Google Scholar] [CrossRef]
- Ticconi, C.; Pietropolli, A.; D’Ippolito, S.; Chiaramonte, C.; Piccione, E.; Scambia, G.; Di Simone, N. Time-To-Pregnancy in Women with Unexplained Recurrent Pregnancy Loss: A Controlled Study. Reprod. Sci. (in press).
- Wang, W.J.; Hao, C.F.; Lin, Q.D. Dysregulation of Macrophage Activation by Decidual Regulatory T Cells in Unexplained Recurrent Miscarriage Patients. J. Reprod. Immunol. 2011, 92, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Clifford, K.; Flanagan, A.M.; Regan, L. Endometrial CD56+Natural Killer Cells in Women with Recurrent Miscarriage: A Histomorphometric Study. Hum. Reprod. 1999, 14, 2727–2730. [Google Scholar] [CrossRef] [PubMed]
- Tuckerman, E.; Laird, S.M.; Prakash, A.; Li, T.C. Prognostic Value of the Measurement of Uterine Natural Killer Cells in the Endometrium of Women with Recurrent Miscarriage. Hum. Reprod. 2007, 22, 2208–2213. [Google Scholar] [CrossRef] [PubMed]
- Kuon, R.J.; Weber, M.; Heger, J.; Santillan, I.; Vomstein, K.; Bar, C.; Strowitzki, T.; Markert, U.R.; Toth, B. Uterine Natural Killer Cells in Patients with Idiopathic Recurrent Miscarriage. Am. J. Reprod. Immunol. 2017, 78, e12721. [Google Scholar] [CrossRef] [PubMed]
- El-Azzamy, H.; Dambaeva, S.V.; Katukurundage, D.; Salazar Garcia, M.D.; Skariah, A.; Hussein, Y.; Germain, A.; Fernandez, E.; Gilman-Sachs, A.; Beaman, K.D.; et al. Dysregulated Uterine Natural Killer Cells and Vascular Remodeling in Women with Recurrent Pregnancy Losses. Am. J. Reprod. Immunol. 2018, 80, e13024. [Google Scholar] [CrossRef]
- Seshadri, S.; Sunkara, S.K. Natural Killer Cells in Female Infertility and Recurrent Miscarriage: A Systematic Review and Meta-Analysis. Hum. Reprod. Update 2014, 20, 429–438. [Google Scholar] [CrossRef]
- Kuon, R.J.; Vomstein, K.; Weber, M.; Muller, F.; Seitz, C.; Wallwiener, S.; Strowitzki, T.; Schleussner, E.; Markert, U.R.; Daniel, V.; et al. The “Killer Cell Story” in Recurrent Miscarriage: Association Between Activated Peripheral Lymphocytes and Uterine Natural Killer Cells. J. Reprod. Immunol. 2017, 119, 9–14. [Google Scholar] [CrossRef]
- Michimata, T.; Ogasawara, M.S.; Tsuda, H.; Suzumori, K.; Aoki, K.; Sakai, M.; Fujimura, M.; Nagata, K.; Nakamura, M.; Saito, S. Distributions of Endometrial NK Cells, B Cells, T Cells, and Th2/Tc2 Cells Fail to Predict Pregnancy Outcome Following Recurrent Abortion. Am. J. Reprod. Immunol. 2002, 47, 196–202. [Google Scholar] [CrossRef]
- LaChapelle, M.H.; Miron, P.; Hemmings, R.; Roy, D.C. Endometrial T, B, and NK Cells in Patients with Recurrent Spontaneous Abortion. Altered Profile and Pregnancy Outcome. J. Immunol. 1996, 156, 4027–4034. [Google Scholar] [PubMed]
- Fuchinoue, K.; Fukui, A.; Chiba, H.; Kamoi, M.; Funamizu, A.; Taima, A.; Fukuhara, R.; Mizunuma, H. Expression of Retinoid-Related Orphan Receptor (ROR)γt on NK22 Cells in the Peripheral Blood and Uterine Endometrium of Women with Unexplained Recurrent Pregnancy Loss and Unexplained Infertility. J. Obstet. Gynaecol. Res. 2016, 42, 1541–1552. [Google Scholar] [CrossRef] [PubMed]
- Kwak-Kim, J.; Yang, K.M.; Gilman-Sachs, A. Recurrent Pregnancy Loss: A Disease of Inflammation and Coagulation. J. Obstet. Gynaecol. Res. 2009, 35, 609–622. [Google Scholar] [CrossRef] [PubMed]
- Kamoi, M.; Fukui, A.; Kwak-Kim, J.; Fuchinoue, K.; Funamizu, A.; Chiba, H.; Yokota, M.; Fukuhara, R.; Mizunuma, H. NK22 Cells in the Uterine Mid-Secretory Endometrium and Peripheral Blood of Women with Recurrent Pregnancy Loss and Unexplained Infertility. Am. J. Reprod. Immunol. 2015, 73, 557–567. [Google Scholar] [CrossRef]
- Fukui, A.; Kamoi, M.; Funamizu, A.; Fuchinoue, K.; Chiba, H.; Yokota, M.; Fukuhara, R.; Mizunuma, H. NK Cell Abnormality and its Treatment in Women with Reproductive Failures Such as Recurrent Pregnancy Loss, Implantation Failures, Preeclampsia, and Pelvic Endometriosis. Reprod Med. Biol. 2015, 14, 151–157. [Google Scholar] [CrossRef]
- O’Hern Perfetto, C.; Fan, X.; Dahl, S.; Krieg, S.; Westphal, L.M.; Bunker Lathi, R.; Nayak, N.R. Expression of Interleukin-22 in Decidua of Patients with Early pRegnancy and Unexplained Recurrent Pregnancy Loss. J. Assist. Reprod. Genet. 2015, 32, 977–984. [Google Scholar] [CrossRef]
- Peng, D.; Xi, W.; Jia, L.; Cui-Yan, Y.; Jing, Y.; Lan-Rong, L.; Qiao-Fei, H.; Jian, L. Simultaneous Detection of Decidual Th1/Th2 and NK1/NK2 Immunophenotyping in Unknown Recurrent Miscarriage Using 8-Color Flow Cytometry with FSC/Vt Extended Strategy. Biosci. Rep. 2017, 37, BSR20170150. [Google Scholar]
- Tang, A.W.; Alfirevic, Z.; Quenby, S. Natural Killer Cells and Pregnancy Outcomes in Women with Recurrent Miscarriage and Infertility: A Systematic Review. Hum. Reprod. 2011, 26, 1971–1980. [Google Scholar] [CrossRef]
- Fukui, A.; Funamizu, A.; Yokota, M.; Yamada, K.; Nakamua, R.; Fukuhara, R.; Kimura, H.; Mizunuma, H. Uterine and Circulating Natural Killer Cells and Their Roles in Women with Recurrent Pregnancy Loss, Implantation Failure and Preeclampsia. J. Reprod. Immunol. 2011, 90, 105–110. [Google Scholar] [CrossRef]
- Ali, S.B.; Jeelall, Y.; Pennell, C.E.; Hart, R.; McLean-Tooke, A.; Lucas, M. The Role of Immunological Testing and Intervention in Reproductive Medicine: A Fertile Collaboration? Am. J. Reprod. Immunol. 2018, 79, e12784. [Google Scholar] [CrossRef]
- Lee, S.K.; Na, B.J.; Kim, J.Y.; Hur, S.E.; Lee, M.; Gilman-Sachs, A.; Kwak-Kim, J. Determination of Clinical Cellular Immune Markers in Women with Recurrent Pregnancy Loss. Am. J. Reprod. Immunol. 2013, 70, 398–411. [Google Scholar] [CrossRef] [PubMed]
- Azargoon, A.; Mirrasouli, Y.; Shokrollahi Barough, M.; Barati, M.; Kokhaei, P. The State of Peripheral Blood Natural Killer Cells and Cytotoxicity in Women with Recurrent Pregnancy Loss and Unexplained Infertility. Int. J. Fertil. Steril. 2019, 13, 12–17. [Google Scholar] [PubMed]
- Davies, M.L.; Dambaeva, S.V.; Katukurundage, D.; Repak, M.; Gilman-Sachs, A.; Kwak-Kim, J.; Beaman, K.D. Predicting NK Cell Subsets Using Gene Expression Levels in Peripheral Blood and Endometrial Biopsy Specimens. Am. J. Reprod. Immunol. 2017, 78, e12730. [Google Scholar] [CrossRef] [PubMed]
- Moffett, A.; Colucci, F. Uterine NK Cells: Active Regulators at the Maternal-Fetal Interface. J. Clin. Invest. 2014, 124, 1872–1879. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Wang, X.; Lu, P.; Song, Y.; Lin, Q. Killer Immunoglobulin-Like Receptor Repertoire on Uterine Natural Killer Cell Subsets in Women with Recurrent Spontaneous Abortions. Eur. J. Obstet. Gynecol. Reprod. Biol. 2008, 140, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Li, Y.P.; Ding, B.; Zhao, Y.R.; Chen, Z.J.; Xu, C.Y. Recurrent Miscarriage is Associated with a Decline of Decidual Natural Killer Cells Expressing Killer Cell Immunoglobulin-Like Receptors Specific for Human Leukocyte Antigen C. J. Obstet. Gynaecol. Res. 2014, 40, 1288–1295. [Google Scholar] [CrossRef] [PubMed]
- Quenby, S.; Nik, H.; Innes, B.; Lash, G.; Turner, M.; Drury, J.; Bulmer, J. Uterine Natural Killer Cells and Angiogenesis in Recurrent Reproductive Failure. Hum. Reprod. 2009, 24, 45–54. [Google Scholar] [CrossRef]
- Fukui, A.; Ntrivalas, E.; Fukuhara, R.; Fujii, S.; Mizunuma, H.; Gilman-Sachs, A.; Beaman, K.; Kwak-Kim, J. Correlation Between Natural Cytotoxicity Receptors and Intracellular Cytokine Expression of Peripheral Blood NK Cells in Women with Recurrent Pregnancy Losses and Implantation Failures. Am. J. Reprod. Immunol. 2009, 62, 371–380. [Google Scholar] [CrossRef]
- Fukui, A.; Kwak-Kim, J.; Ntrivalas, E.; Gilman-Sachs, A.; Lee, S.K.; Beaman, K. Intracellular Cytokine Expression of Peripheral Blood Natural Killer Cell Subsets in Women with Recurrent Spontaneous Abortions and Implantation Failures. Fertil. Steril. 2008, 89, 157–165. [Google Scholar] [CrossRef]
- Plaks, V.; Birnberg, T.; Berkutzki, T.; Sela, S.; BenYashar, A.; Kalchenko, V.; Mor, G.; Keshet, E.; Dekel, N.; Neeman, M.; et al. Uterine DCs are Crucial for Decidua Formation During Embryo Implantation in Mice. J. Clin. Invest. 2008, 118, 3954–3965. [Google Scholar] [CrossRef]
- Huang, C.; Zhang, H.; Chen, X.; Diao, L.; Lian, R.; Zhang, X.; Hu, L.; Zeng, Y. Association of Peripheral Blood Dendritic Cells with Recurrent Pregnancy loss: A Case-Controlled Study. Am. J. Reprod. Immunol. 2016, 76, 326–332. [Google Scholar] [CrossRef] [PubMed]
- Askelund, K.; Liddell, H.S.; Zanderigo, A.M.; Fernando, N.S.; Khong, T.Y.; Stone, P.R.; Chamley, L.W. CD83(+)Dendritic Cells in the Decidua of Women with Recurrent Miscarriage and Normal Pregnancy. Placenta 2004, 25, 140–145. [Google Scholar] [CrossRef]
- Qian, Z.D.; Huang, L.L.; Zhu, X.M. An Immunohistochemical Study of CD83-And CD1a-Positive Dendritic Cells in the Decidua of Women with Recurrent Spontaneous Abortion. Eur. J. Med. Res. 2015, 20, 2. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Wei, H.; Li, Y.; Huang, C.; Lian, R.; Xu, J.; Chen, L.; Zeng, Y. Downregulation of ILT4+ Dendritic Cells in Recurrent Miscarriage and Recurrent Implantation Failure. Am. J. Reprod. Immunol. 2018, 80, e12998. [Google Scholar] [CrossRef]
- Aluvihare, V.R.; Kallikourdis, M.; Betz, A.G. Regulatory T Cells Mediate Maternal Tolerance to the Fetus. Nat. Immunol. 2004, 5, 266–271. [Google Scholar] [CrossRef]
- Darrasse-Jeze, G.; Darasse-Jeze, G.; Klatzmann, D.; Charlotte, F.; Salomon, B.L.; Cohen, J.L. CD4+CD25+Regulatory/Suppressor T Cells Prevent Allogeneic Fetus Rejection in Mice. Immunol. Lett. 2006, 102, 106–109. [Google Scholar] [CrossRef]
- Zenclussen, A.C.; Gerlof, K.; Zenclussen, M.L.; Sollwedel, A.; Bertoja, A.Z.; Ritter, T.; Kotsch, K.; Leber, J.; Volk, H.D. Abnormal T-Cell Reactivity Against Paternal Antigens in Spontaneous Abortion: Adoptive Transfer of Pregnancy-Induced CD4+CD25+T Regulatory Cells Prevents fetal Rejection in a Murine Abortion Model. Am. J. Pathol. 2005, 166, 811–822. [Google Scholar] [CrossRef]
- Wang, W.J.; Liu, F.J.; Liu, X.; Hao, C.F.; Bao, H.C.; Qu, Q.L.; Liu, X.M. Adoptive Transfer of Pregnancy-Induced CD4+CD25+Regulatory T Cells Reverses the Increase in Abortion Rate Caused by Interleukin 17 in the CBA/JxBALB/c Mouse Model. Hum. Reprod. 2014, 29, 946–952. [Google Scholar] [CrossRef]
- Samstein, R.M.; Josefowicz, S.Z.; Arvey, A.; Treuting, P.M.; Rudensky, A.Y. Extrathymic Generation of Regulatory T Cells in Placental Mammals Mitigates Maternal-Fetal Conflict. Cell 2012, 150, 29–38. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.S.; Wu, L.; Tong, X.H.; Wu, L.M.; He, G.P.; Zhou, G.X.; Luo, L.H.; Luan, H.B. Study on the Relationship Between Th17 Cells and Unexplained Recurrent Spontaneous Abortion. Am. J. Reprod. Immunol. 2011, 65, 503–511. [Google Scholar] [CrossRef]
- Zhang, X.X.; Kang, X.M.; Zhao, A.M. Regulation of CD4+FOXP3+T Cells by CCL20/CCR6 Axis in Early Unexplained Recurrent Miscarriage Patients. Genet. Mol. Res. 2015, 14, 9145–9154. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Liu, P.; Cheng, L. Galectin-1 Reduction and Changes in T Regulatory Cells May Play Crucial Roles in Patients with Unexplained Recurrent Spontaneous Abortion. Int. J. Clin. Exp. Pathol. 2015, 8, 1973–1978. [Google Scholar] [PubMed]
- Inada, K.; Shima, T.; Ito, M.; Ushijima, A.; Saito, S. Helios-Positive Functional Regulatory T Cells are Decreased in Decidua of Miscarriage Cases with Normal Fetal Chromosomal Content. J. Reprod. Immunol. 2015, 107, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Qian, J.; Zhang, N.; Lin, J.; Wang, C.; Pan, X.; Chen, L.; Li, D.; Wang, L. Distinct Pattern of Th17/Treg Cells in Pregnant Women with a History of Unexplained Recurrent Spontaneous Abortion. Biosci. Trends 2018, 12, 157–167. [Google Scholar] [CrossRef]
- Wang, W.J.; Hao, C.F.; Lin, Y.; Yin, G.J.; Bao, S.H.; Qui, L.H.; Lin, Q.D. Increased Prevalence of T Helper 17 (Th17) Cells in Peripheral Blood and Decidua in Unexplained Recurrent Spontaneous Abortion Patients. J. Reprod. Immunol. 2010, 84, 164–170. [Google Scholar] [CrossRef]
- Saifi, B.; Rezaee, S.A.; Tajik, N.; Ahmadpour, M.E.; Ashrafi, M.; Vakili, R.; SoleimaniAsl, S.; Aflatoonian, R.; Mehdizadeh, M. Th17 Cells and Related Cytokines in Unexplained Recurrent Spontaneous Miscarriage at the Implantation Window. Reprod. Biomed. Online 2014, 29, 481–489. [Google Scholar] [CrossRef]
- Lee, S.K.; Kim, J.Y.; Hur, S.E.; Kim, C.J.; Na, B.J.; Lee, M.; Gilman-Sachs, A.; Kwak-Kim, J. An Imbalance in Interleukin-17-Producing T and Foxp3⁺ Regulatory T Cells in Women with Idiopathic Recurrent Pregnancy Loss. Hum. Reprod. 2011, 26, 2964–2971. [Google Scholar] [CrossRef]
- Wu, L.; Li, J.; Xu, H.L.; Xu, B.; Tong, X.H.; Kwak-Kim, J.; Liu, Y.S. IL-7/IL-7R Signaling Pathway Might Play a Role in Recurrent Pregnancy Losses by Increasing Inflammatory Th17 Cells and Decreasing Treg Cells. Am. J. Reprod. Immunol. 2016, 76, 454–464. [Google Scholar] [CrossRef]
- Figueiredo, A.S.; Schumacher, A. The T Helper Type 17/Regulatory T Cell Paradigm in Pregnancy. Immunology 2016, 148, 13–21. [Google Scholar] [CrossRef]
- Saxena, D.; Misra, M.K.; Parveen, F.; Phadke, S.R.; Agrawal, S. The Transcription Factor Forkhead Box P3 Gene Variants Affect Idiopathic Recurrent Pregnancy Loss. Placenta 2015, 36, 226–231. [Google Scholar] [CrossRef]
- Zidan, H.E.; Abdul-Maksoud, R.S.; Mowafy, H.E.; Elsayed, W.S.H. The Association of IL-33 and Foxp3 Gene Polymorphisms with Recurrent Pregnancy Loss in Egyptian Women. Cytokine 2018, 108, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Elkord, E.; Abd Al Samid, M.; Chaudhary, B. Helios, and not FoxP3, is the Marker of Activated Tregs Expressing GARP/LAP. Oncotarget 2015, 6, 20026–20036. [Google Scholar] [CrossRef] [PubMed]
- Sadlon, T.; Brown, C.Y.; Bandara, V.; Hope, C.M.; Schjenken, J.E.; Pederson, S.M.; Breen, J.; Forrest, A.; Beyer, M.; Robertson, S.; et al. Unravelling the Molecular Basis for Regulatory T-Cell Plasticity and Loss of Function in Disease. Clin. Transl. Immunol. 2018, 7, e1011. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Qiu, T.; Wang, Y.; Chen, Y.; Cheng, W. Expression of C-Type Lectin Receptors and Toll-Like Receptors in Decidua of Patients with Unexplained Recurrent Spontaneous Abortion. Reprod. Fertil. Dev. 2017, 29, 1613–1624. [Google Scholar] [CrossRef] [PubMed]
- Bao, S.H.; Wang, X.P.; De Lin, Q.; Wang, W.J.; Yin, G.J.; Qiu, L.H. Decidual CD4+CD25+CD127dim/- Regulatory T Cells in Patients with Unexplained Recurrentspontaneous Miscarriage. Eur. J. Obstet. Gynecol. Reprod. Biol. 2011, 155, 94–98. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Li, J.; Huang, S.Y.; Sun, X. Characterization of the Subsets of Human NKT-Like Cells and the Expression of Th1/Th2 Cytokines in Patients with Unexplained Recurrent Spontaneous Abortion. J. Reprod. Immunol. 2015, 110, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Piccinni, M.P.; Beloni, L.; Livi, C.; Maggi, E.; Scarselli, G.; Romagnani, S. Defective Production of Both Leukemia Inhibitory Factor and Type 2 T-Helper Cytokines by Decidual T Cells in Unexplained Recurrent Abortions. Nat. Med. 1998, 4, 1020–1024. [Google Scholar] [CrossRef]
- Vives, A.; Balasch, J.; Yague, J.; Quinto, L.; Ordi, J.; Vanrell, J.A. Type-1 and type-2 Cytokines in Human Decidual Tissue and Trophoblasts from Normal and Abnormal Pregnancies Detected by Reverse Transcriptase Polymerase Chain Reaction (RT-PCR). Am. J. Reprod. Immunol. 1999, 42, 361–368. [Google Scholar] [CrossRef]
- Comba, C.; Bastu, E.; Dural, O.; Yasa, C.; Keskin, G.; Ozsurmeli, M.; Buyru, F.; Serdaroglu, H. Role of Inflammatory Mediators in Patients with Recurrent Pregnancy Loss. Fertil. Steril. 2015, 104, 1467–1474. [Google Scholar] [CrossRef]
- Bates, M.D.; Quenby, S.; Takakuwa, K.; Johnson, P.M.; Vince, G.S. Aberrant Cytokine Production by Peripheral Blood Mononuclear Cells in Recurrent Pregnancy Loss? Hum. Reprod. 2002, 17, 2439–2444. [Google Scholar] [CrossRef]
- Daher, S.; De Arruda Geraldes Denardi, K.; Blotta, M.H.; Mamoni, R.L.; Reck, A.P.; Camano, L.; Mattar, R. Cytokines in Recurrent Pregnancy Loss. J. Reprod. Immunol. 2004, 62, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Talukdar, A.; Rai, R.; Aparna Sharma, K.; Rao, D.N.; Sharma, A. Peripheral Gamma Delta T Cells Secrete Inflammatory Cytokines in Women with Idiopathic Recurrent Pregnancy Loss. Cytokine 2018, 102, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Bombell, S.; McGuire, W. Cytokine Polymorphisms in Women with Recurrent Pregnancy Loss: Meta-Analysis. Aust. N. Z. J. Obstet. Gynaecol. 2008, 48, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Xu, J.; Bao, X.; Niu, W.; Wang, L.; Du, L.; Zhang, N.; Sun, Y. Association Between Genetic Polymorphisms in Interleukin Genes and Recurrent Pregnancy Loss-A Systematic Review and Meta-Analysis. PLoS ONE 2017, 12, e0169891. [Google Scholar] [CrossRef]
- Grimstad, F.; Krieg, S. Immunogenetic Contributions to Recurrent Pregnancy Loss. J. Assist. Reprod. Genet. 2016, 33, 833–847. [Google Scholar] [CrossRef]
- Krieg, S.A.; Fan, X.; Hong, Y.; Sang, Q.X.; Giaccia, A.; Westphal, L.M.; Lathi, R.B.; Krieg, A.J.; Nayak, N.R. Global Alteration in Gene Expression Profiles of Deciduas from Women with Idiopathic Recurrent Pregnancy Loss. Mol. Hum. Reprod. 2012, 189, 442–450. [Google Scholar] [CrossRef]
- Jasper, M.J.; Tremellen, K.P.; Robertson, S.A. Reduced Expression of IL-6 and IL-1alpha mRNAs in Secretory Phase Endometrium of Women with Recurrent Miscarriage. J. Reprod. Immunol. 2007, 73, 74–84. [Google Scholar] [CrossRef]
- D’Ippolito, S.; Tersigni, C.; Marana, R.; Di Nicuolo, F.; Gaglione, R.; Rossi, E.D.; Castellani, R.; Scambia, G.; Di Simone, N. Inflammosome in the Human Endometrium: Further Step in the Evaluation of the “Maternal Side”. Fertil. Steril. 2016, 105, 111–118. [Google Scholar] [CrossRef]
- Von Wolff, M.; Thaler, C.J.; Strowitzki, T.; Broome, J.; Stolz, W.; Tabibzadeh, S. Regulated Expression of Cytokines in Human Endometrium Throughout the Menstrual Cycle: Dysregulation in Habitual Abortion. Mol. Hum. Reprod. 2000, 6, 627–634. [Google Scholar] [CrossRef]
- Wang, W.J.; Liu, F.J.; Qu, H.M.; Hao, C.F.; Qu, Q.L.; Xiong, W.; Bao, H.C.; Wang, X.R. Regulation of the Expression of Th17 Cells and Regulatory T Cells by IL-27 in Patients with Unexplained Early Recurrent Miscarriage. J. Reprod. Immunol. 2013, 99, 39–45. [Google Scholar] [CrossRef]
- Karaer, A.; Cigremis, Y.; Celik, E.; Urhan Gonullu, R. Prokineticin 1 and Leukemia Inhibitory Factor mRNA Expression in the Endometrium of Women with Idiopathic Recurrent Pregnancy Loss. Fertil. Steril. 2014, 102, 1091–1095. [Google Scholar] [CrossRef] [PubMed]
- Brezina, P.R.; Kutteh, W.H. Classic and Cutting Edge Strategies for the Management of Early Pregnancy Loss. Obstet. Gynecol. Clin. North. Am. 2014, 41, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Christiansen, O.B.; Kolte, A.M.; Krog, M.C.; Nielsen, H.S.; Egerup, P. Treatment with Intravenous Immunoglobulin in Patients with Recurrent Pregnancy Loss: An Update. J. Reprod. Immunol. 2019, 133, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Crawford, G.; Ray, A.; Gudi, A.; Shah, A.; Homburg, R. The Role of Seminal Plasma for Improved Outcomes During in Vitro Fertilization Treatment: Review of the Literature and Meta-Analysis. Hum. Reprod. Update 2015, 21, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.J.; Liu, F.J.; Zhang, X.; Liu, X.M.; Qu, Q.L.; Li, F.H.; Zhuang, L.L.; Li, X.X.; Hao, C.F. Periodic Elevation of Regulatory T Cells on the Day of Embryo Transfer is Associated with Better in Vitro Fertilization Outcome. J. Reprod. Immunol. 2017, 119, 49–53. [Google Scholar] [CrossRef] [PubMed]
- Ata, B.; Abou-Setta, A.M.; Seyhan, A.; Buckett, W. Application of Seminal Plasma to Female Genital Tract Prior to Embryo Transfer in Assisted Reproductive Technology Cycles (IVF, ICSI and Frozen Embryo Transfer. Cochrane Database Syst. Rev. 2018, 28, CD011809. [Google Scholar] [CrossRef] [PubMed]
- Bouet, P.E.; El Hachem, H.; Monceau, E.; Gariepy, G.; Kadoch, I.J.; Sylvestre, C. Chronic Endometritis in Women with Recurrent Pregnancy Loss and Recurrent Implantation Failure: Prevalence and Role of Office Hysteroscopy and Immunohistochemistry in Diagnosis. Fertil. Steril. 2016, 105, 106–110. [Google Scholar] [CrossRef]
- Kitaya, K. Prevalence of Chronic Endometritis in Recurrent Miscarriages. Fertil. Steril. 2011, 95, 1156–1158. [Google Scholar] [CrossRef]
- McQueen, D.B.; Perfetto, C.O.; Hazard, F.K.; Lathi, R.B. Pregnancy Outcomes in Women with Chronic Endometritis and Recurrent Pregnancy Loss. Fertil. Steril. 2015, 104, 927–931. [Google Scholar] [CrossRef]
- Wu, D.; Kimura, F.; Zheng, L.; Ishida, M.; Niwa, Y.; Hirata, K.; Takebayashi, A.; Takashima, A.; Zhang, G.; Murakami, T. Chronic Endometritis Modifies Decidualization in Human Endometrial Stromal Cells. Reprod. Biol. Endocrinol. 2017, 15, 16. [Google Scholar] [CrossRef]
- Cicinelli, E.; Matteo, M.; Tinelli, R.; Pinto, V.; Marinaccio, M.; Indraccolo, U.; De Ziegler, D.; Resta, L. Chronic Endometritis Due to Common Bacteria is Prevalent in Women with Recurrent Miscarriage as Confirmed by Improved Pregnancy Outcome after Antibiotic Treatment. Reprod. Sci. 2014, 21, 640–647. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Morphological (Tissutal and Cell) Changes |
| Secretory transformation of endometrial glands, compaction of surface epithelial cells, stromal edema, stromal cell proliferation, differentiation of fibroblast-like stromal cells into epithelioid-like cells, massive leukocyte infiltration (mainly uterine natural killer cells and mast cells), spiral arteries remodeling |
| Extracellular Matrix Changes |
| Increased production by decidualized stromal cells of collagen IV, fibronectin, laminin, decorin, heparan-sulphate proteoglycans, IGFBP-1, PRL, LEFTY-2, αvβ3 integrin, osteopontin |
| Changes in the Expression of Genes Involved in the Following Cellular Functions |
| Cell cycle regulation, cytoskeleton remodeling, oxidative stress response, ion and water transport, response to steroid hormone, deposition of extracellular matrix (ECM), modulation of transcription, epigenetic patterning, post-translation modification, growth factor, angiogenesis, cytokine and chemokine signaling, immune modulation |
| Cell Signaling and Pathways Involved in the Decidualization |
| cAMP/PKA pathway, progesterone signaling pathway, Nodal pathway, Notch signaling pathway, ERK1/2 pathway, WNT/β-catenin pathway, cSRC pathway, JAK-STAT pathway, lipid signaling (endocannabinoid system), TGFβ signaling pathway, BMP2-WNT4 signaling cascade, phosphatidylinositol 3-kinase/AKT pathway, Ras/Raf1/MAPK pathway, EPAC1 and EPAC2 signaling |
| Cell Type | Phenotype | Density (%) of Leukocyte Population in the Endometrium | Changes during the Menstrual Cycle and in Early Pregnancy | Preferential Location in the Endometrium | Relevant Molecules Secreted | Suggested Functions | References |
|---|---|---|---|---|---|---|---|
| Uterine Natural killer Cells (uNK) | CD3−CD56brightCD16− (predominant phenotype) | 30%–40% of stromal cells 70% of endometrial leukocytes in the late luteal phase (LP) and in early pregnancy | Progressively increase from the follicular phase (FP) to the LP. Maximal density in late LP and in gestational decidua | Surround the arteries and the glands | IFN-γ, VEGF, PlGF, TGF-β, TNF-α, IL-10, GM-CSF, IL-1β, LIF, CSF-1, AP-2 | Tissue (spiral arteries) remodeling, enhancement of angiogenesis, control of trophoblast invasion | [10,20,24,25,26,27,28,29] |
| Macrophages (Mφ) | CD68+ | 20%–25% of total leukocytes in the decidua | Progressively increase from the FP to the LP. Maximal density before menstruation and in pregnancy. Acquire tolerogenic phenotype | Scattered throughout the endometrium; preferentially found around the glands and at implantation site | TGF-β, IL-10, IDO, PGE2 | Involved in corpus luteum maintenance, blastocyst implantation, spiral arteries remodeling, control of trophoblast invasion, protection of the fetus against intrauterine infection | [20,29,30,31,32,33,34,35] |
| Mast Cells (MCs) | MCT; MCTC; MCC (endometrial MCs) | 3%–5% of total endometrial cells; | Unchanged throughout menstrual cycle; changes in phenotype during the menstrual cycle; activated in the early and midluteal phase | More prominent in the basal endometrial compartment | VEGF | Initiation of menstruation Enhance tissue and spiral artery remodeling, support implantation and angiogenesis | [20,36,37] |
| Uterine Dendritic Cells (DCs) | CD1a+ (immature, tolerogenic DCs); CD83+ (mature DCs) | Density of immature DCs in the endometrium is higher than that of mature DCs; DC 1%–2% of the immune cells in the decidua | Immature DCs increase from FP to LP; peak in the menstrual phase (controversial finding); No changes in mature DCs with the menstrual cycle | Both mature and immature DCs are found mainly in the basal layer of the endometrium in the LP; Scattered through the gestational decidua In mice grouped in cluster-like structures | TGF-β, IL-10, IDO | Involved in the maternal acceptance of the embryo, trophoblast invasion and differentiation; Uterine remodeling, angiogenesis; Determine the differentiation of T cell progenitors into Tregs and expansion and activation of Tregs | [29,33,38,39,40,41,42,43] |
| Neutrophil (N) Granulocytes | CD11b+; CD16b+; CD66c+ | 1% to 6%–15% of endometrial cells | Considerably increase in the late (premenstrual) LP | Endometrial Stroma | CCL2, CXCL8, TNF-α, IL-6, VEGF | Menstruation, tissue breakdown and repair, Proangiogenic and tolerogenic in the pregnant decidua | [44,45,46,47] |
| Cell Type | Phenotype | Density (%) of Leukocyte (CD45+) Population in the Endometrium | Changes during the Menstrual Cycle and in Early Pregnancy | Preferential Location in the Endometrium | Relevant Molecules Secreted | Suggested Functions | References |
|---|---|---|---|---|---|---|---|
| BLymphocytes | CD45+ CD19+ | 0.2%–4.5% | Very slight increase in late LP | Clusters among stromal cells in the perimenstrual period | IL-10 | Still undetermined; Potentially implicated in early pregnancy | [20,47,48] |
| T Lymphocytes | CD45+ CD3+ | 1%–2% to 28% | Reportedly decreased or unchanged from follicular phase (FP) to luteal phase (LP) | Lymphoid aggregates; scattered throughout epithelium and stroma | Variable according to the specific cell subset | Protective or harmful for the embryo according to the specific cell subset | [24,29,48,49] |
| T Helper Lymphocytes (Th1, Th2, Th17, Tregs) | CD45+ CD3+ CD4+ | 3.8%–21.4% | No clear variations reported | Present in uterine mucosa as unique aggregates surrounding a B cell core | TNF-α and IFN-γ (by Th1); IL-4 (by Th2); IL-8 (by Th17) | Th1 produce inflammatory cytokines; Th2 produce anti-inflammatory cytokines; Th17 has pro-inflammatory effects (for Tregs see the dedicated section in the table) | [33,48,50] |
| T Cytotoxic Lymphocytes | CD45+ CD3+ CD8+ | 4.4%–34.5% to 66% | Significantly decrease from FP to LP | Lymphoid aggregates | Release cytotoxic substances (granzymes, perforin) | Potentially harmful to the embryo. Blocked in successful pregnancy | [47,48,49,51] |
| Tregs | CD4+ CD25+ FOXP3+ | Not clearly defined in humans | Expansion in preimplantation endometrium; Increase in the decidua at implantation site and in early pregnancy until midgestation | Insufficient data in human preimplantation endometrium | Galectin-1, TGF-β, IL-10, HO-1 | Essential in the control of an excessive maternal inflammatory response at the implantation site; Involved in maternal immune tolerance to fetal allograft particularly in early pregnancy; Block maternal effector T cells Involved in the regulation of maternal vascular remodeling | [20,33,49,50,52,53,54,55,56,57] |
| Mechanism | Effect | References |
|---|---|---|
| Increased secretion of LIF and IL-1β by uterine macrophages | Enhancement of embryo attachment to endometrial epithelium | [58] |
| TGFβ production by maternal decidual macrophages | Suppression of EVT rejection mediated by NK cells | [59] |
| Production by macrophages of TGFβ, IL-10, IDO, and PGE2 | Immunoinhibitory and pro-tolerance actions | [33] |
| Specific characteristics of uNK cells in the endometrium and of dNK in pregnancy | Acquisition of a regulatory role rather than classic cell killing ability | [21,27,60] |
| dNK cells dampen Th17 cells through the production of IFN | Suppress Th17-induced inflammatory response | [61] |
| Galectin-1 production by dNK cells | Induces apoptosis of activated CD8+ T cells | [62,63,64] |
| Endometrial recruitment of innate immune cells (Mφ, DCs, and granulocytes) triggered by seminal fluid before implantation | Activation and expansion of Tregs which, in turn, creates a uterine microenvironment favorable for embryo implantation and enhances maternal tolerance towards paternal MHC antigens | [21,54,65,66,67] |
| Induction and expansion of tolerogenic DCs phenotype | Involved in the control and activation of Tregs | [33,54,68,69] |
| Reduced production of IL-12 by DCs | Priming of decidual CD4+ cells into a Th2 phenotype | [70,71] |
| EVT cells express HLA-C, HLA-E, and HLA-G but not HLA-A and HLA-B Lack of expression of MHC molecules by syncytiotrophoblasts | Inhibition of cytolytic activity of dNK cells against the trophoblasts Induction of NK senscence-Enhanced apoptosis of activated CD8+ cells | [70,72,73,74,75] |
| Trophoblast production of exosomes | Downregulation of maternal immunity towards the trophoblast | [76] |
| Development of a specific cytokine and chemokine network in the endometrium and decidua | Achievement of a correct immune cell recruitment and dialogue favoring embryo implantation and proper pregnancy evolution | [77] |
| Presence of asymmetric maternal Abs against paternal antigens | Bind trophoblast but are unable to trigger destructive immune response | [78] |
| Increased production of PIBF | Upregulation of Th2 cytokines production. Downregulation of dNK activity. Increase the production of Glycodelin A which induces apoptosis in T cells. These effects can contribute to trophoblast immune protection | [66,79] |
| Increased progesterone production in LP and in pregnancy | Expansion of Tregs and enhancement of their immunosuppressive actions | [33,78] |
| hCG production by syncytiotrophoblast | Recruitment of Tregs at the maternal–fetal interface | [80] |
| Cytokine | Tissue/Cells | Type of Variation | Expression/Production | Methods of Detection | References |
|---|---|---|---|---|---|
| IL-1 (α and β) | Decidua | Decrease | IL-1 pathway gene expression | Microarray | [188] |
| IL-1α | Endometrium | Decrease | mRNA expression | RT-PCR | [189] |
| IL-1β | Endometrium Endometrium | Increase Decrease | Tissue secretion mRNA expression | ELISA RNase protection assay | [190] [191] |
| IL-2 | Decidua | Increase | Cytokine production | ELISA | [176] |
| IL-4 | Decidua Decidual T cells | Decrease Decrease | mRNA and protein expression Cytokine production | ELISA + RT-PCR ELISA | [178] [179] |
| IL-6 | Decidua Endometrium Endometrium | Increase Decrease Decrease | mRNA and protein expression mRNA expression mRNA expression | RT-PCR RT-PCR RNase protection assay | [166] [189] [191] |
| IL-7 | Decidual stromal cells | Increase | Cytokine expression | IHC | [170] |
| IL-8 | Decidua | Increase | IL-8 pathway gene expression | Microarray | [188] |
| IL-10 | Decidua Decidua Decidua Decidual Tregs Decidual T cells | Decrease Decrease Decrease Decrease Decrease | mRNA and protein expression Cytokine production Protein expression Cytokine expression Cytokine production | ELISA + RT-PCR ELISA RT-PCT Flow cytometry ELISA | [178] [176] [180] [177] [179] |
| IL-12 | Endometrium | Increase | Tissue levels | ELISA | [181] |
| IL-17 | Decidua Decidua | Unchanged Increase | mRNA and protein expression mRNA and protein expression | qRT-PCR + IHC +WB RT-PCR + WB | [138] [167] |
| IL-18 | Endometrium Endometrium | Increase Increase | Tissue levels Tissue secretion | ELISA ELISA | [181] [190] |
| IL-22 | Decidua | Decrease | mRNA and protein expression | qRT-PCR + IHC +WB | [138] |
| IL-23 | Decidua Decidua | Unchanged Increase | mRNA and protein expression mRNA and protein expression | qRT-PCR + IHC +WB RT-PCR + WB | [138] [167] |
| IL-27 | Decidua | Decrease | mRNA and protein expression | qRT-PCR + WB | [192] |
| TGF-β | Decidual Tregs | Decrease | Cytokine expression | Flow cytometry | [177] |
| TGF-β1 | Decidua Decidua | Decrease Decrease | mRNA and protein expression Cytokine production | RT-PCR + ELISA ELISA | [166] [176] |
| IFN-γ | Endometrium Decidua Decidua | Increase Increase Increase | Tissue levels Cytokine production mRNA and protein expression | ELISA ELISA RT-PCR + ELISA | [181] [176] [178] |
| TNF-α | Decidua | Increase | mRNA and protein expression | RT-PCR + ELISA | [166] |
| LIF | Endometrium Endometrium Decidual T cells | Decrease Increase Decrease | Tissue levels mRNA expression Cytokine production | ELISA RT-PCR ELISA | [181] [193] [179] |
| MIF | Endometrium Decidual T cells | Decrease Decrease | Tissue levels Cytokine production | ELISA ELISA | [181] [179] |
| Suggested Alteration | Potential Pathogenetic Mechanism(s) |
|---|---|
| Abnormal decidualization of the ESC | Altered control of the local trafficking of immune cells by ESC; Altered modulation of the inflammatory response to the implanting embryo; Dysregulation in cytokine production |
| Abnormal killer immunoglobulin receptor (KIR)–uNK interaction | Abolished or reduced prevention of the maternal immune rejection of the conceptus |
| Increased toxicity of uNK | Impaired capacity to limit T cell cytotoxicity; Potential toxicity toward the cytotrophoblast; Abnormal cytokine production; Defective remodeling of the spiral arteries |
| Abnormal number/function of endometrial/decidual Tregs | Lack of inhibition of Th1 and Th17 cells; Defective/absent resolution of the inflammatory reaction consequent to initial embryo implantation; Reduction or abolition of the maternal immunologic tolerance toward the fetus; Defective remodeling of the spiral arteries |
| Limited differentiation of macrophages toward the M2 immunophenotype | Reduced production of tolerogenic cytokines (IL-10, TGF-β) |
| Lack of immature dendritic cells | Lack of the expansion of Treg population |
| Activation of lymphocytes T effector | Immune attack to the trophoblast |
| Aberrant local cytokine network | Development of a local immunomodulatory environment unfavorable to maternal tolerance toward the conceptus |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ticconi, C.; Pietropolli, A.; Di Simone, N.; Piccione, E.; Fazleabas, A. Endometrial Immune Dysfunction in Recurrent Pregnancy Loss. Int. J. Mol. Sci. 2019, 20, 5332. https://doi.org/10.3390/ijms20215332
Ticconi C, Pietropolli A, Di Simone N, Piccione E, Fazleabas A. Endometrial Immune Dysfunction in Recurrent Pregnancy Loss. International Journal of Molecular Sciences. 2019; 20(21):5332. https://doi.org/10.3390/ijms20215332
Chicago/Turabian StyleTicconi, Carlo, Adalgisa Pietropolli, Nicoletta Di Simone, Emilio Piccione, and Asgerally Fazleabas. 2019. "Endometrial Immune Dysfunction in Recurrent Pregnancy Loss" International Journal of Molecular Sciences 20, no. 21: 5332. https://doi.org/10.3390/ijms20215332
APA StyleTicconi, C., Pietropolli, A., Di Simone, N., Piccione, E., & Fazleabas, A. (2019). Endometrial Immune Dysfunction in Recurrent Pregnancy Loss. International Journal of Molecular Sciences, 20(21), 5332. https://doi.org/10.3390/ijms20215332