Genome-Wide Identification of Long Non-coding RNA in Trifoliate Orange (Poncirus trifoliata (L.) Raf) Leaves in Response to Boron Deficiency

Abstract
:1. Introduction
2. Results
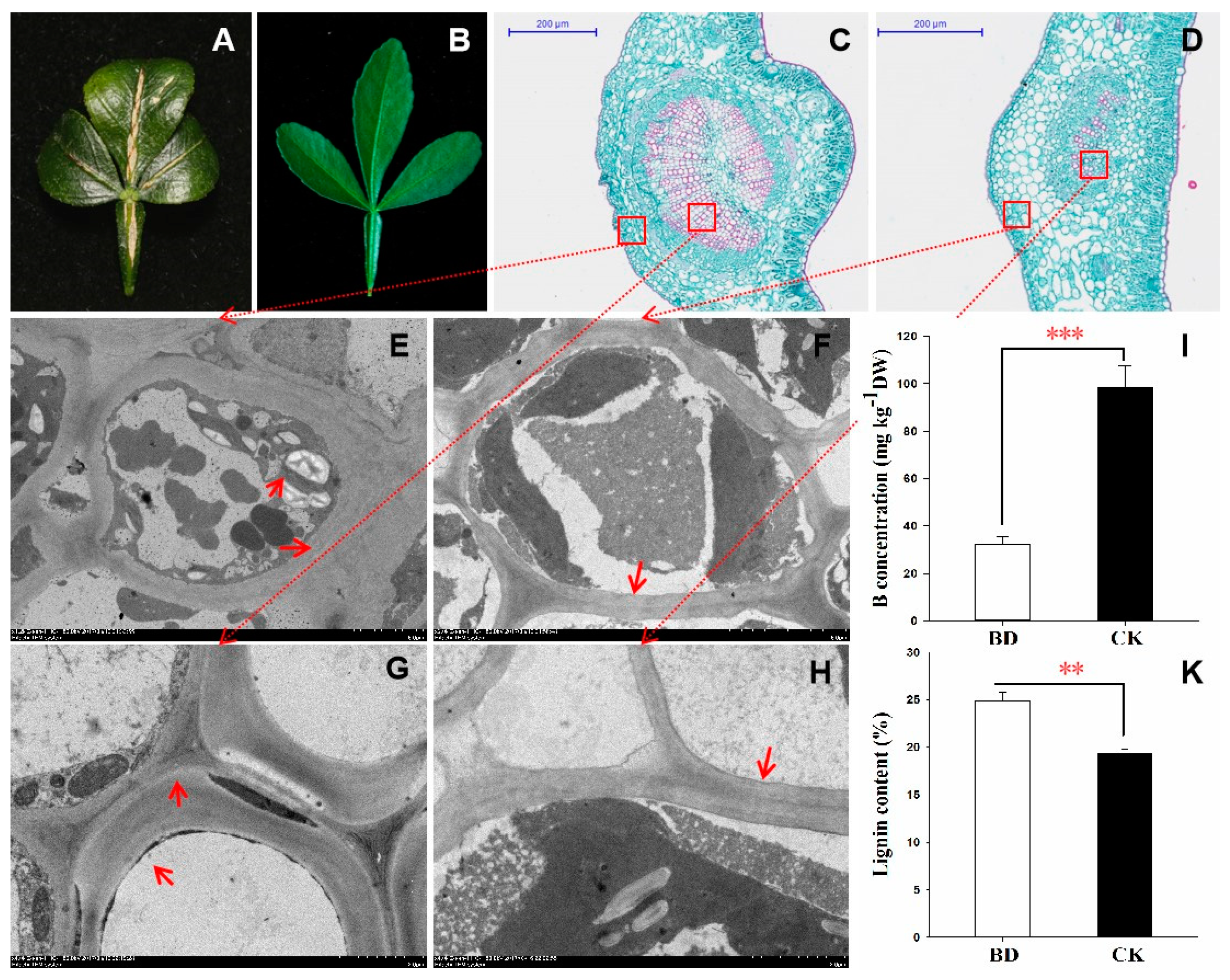
2.1. The Differential Performance of the Leaves under B Deficiency Stress
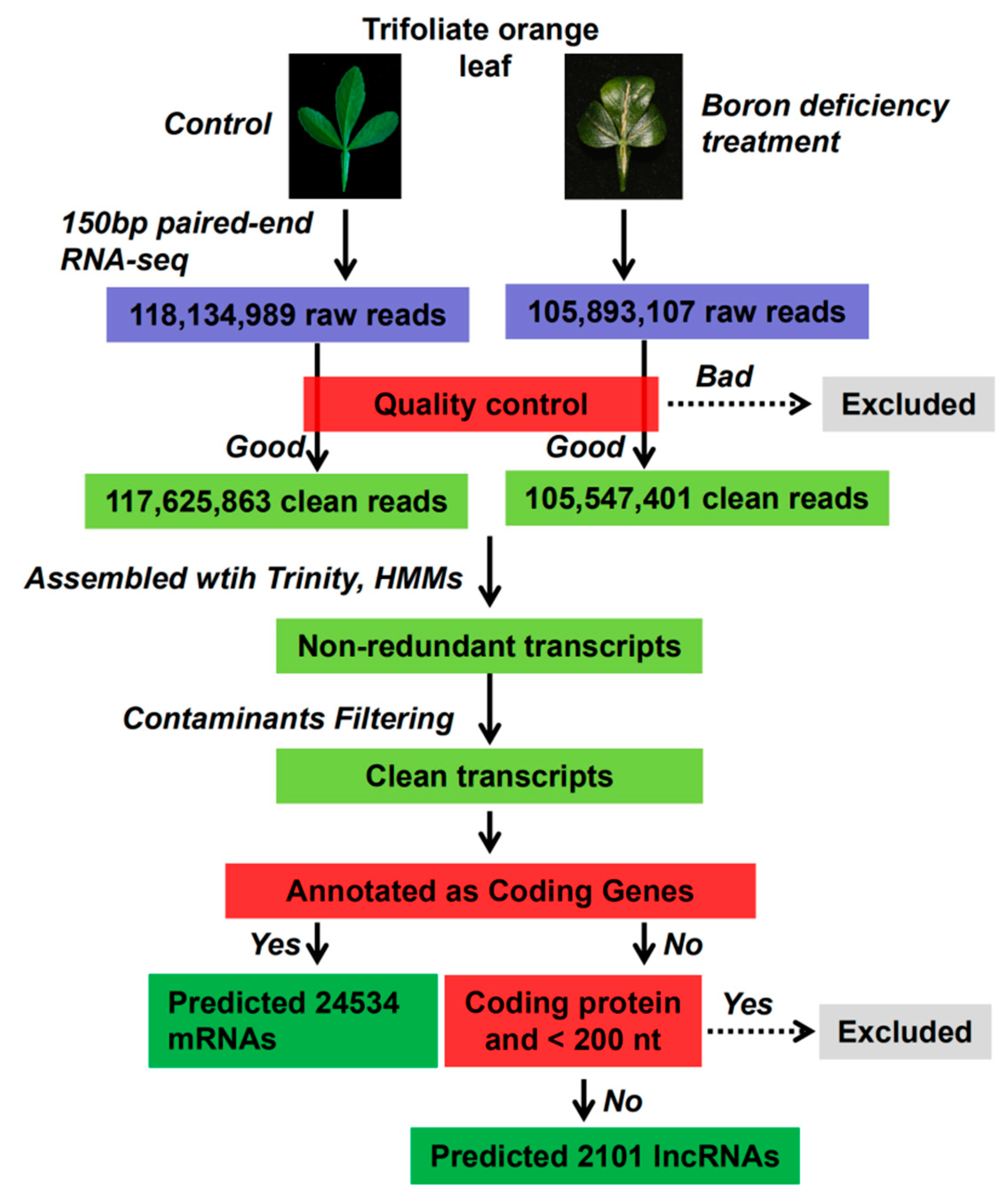
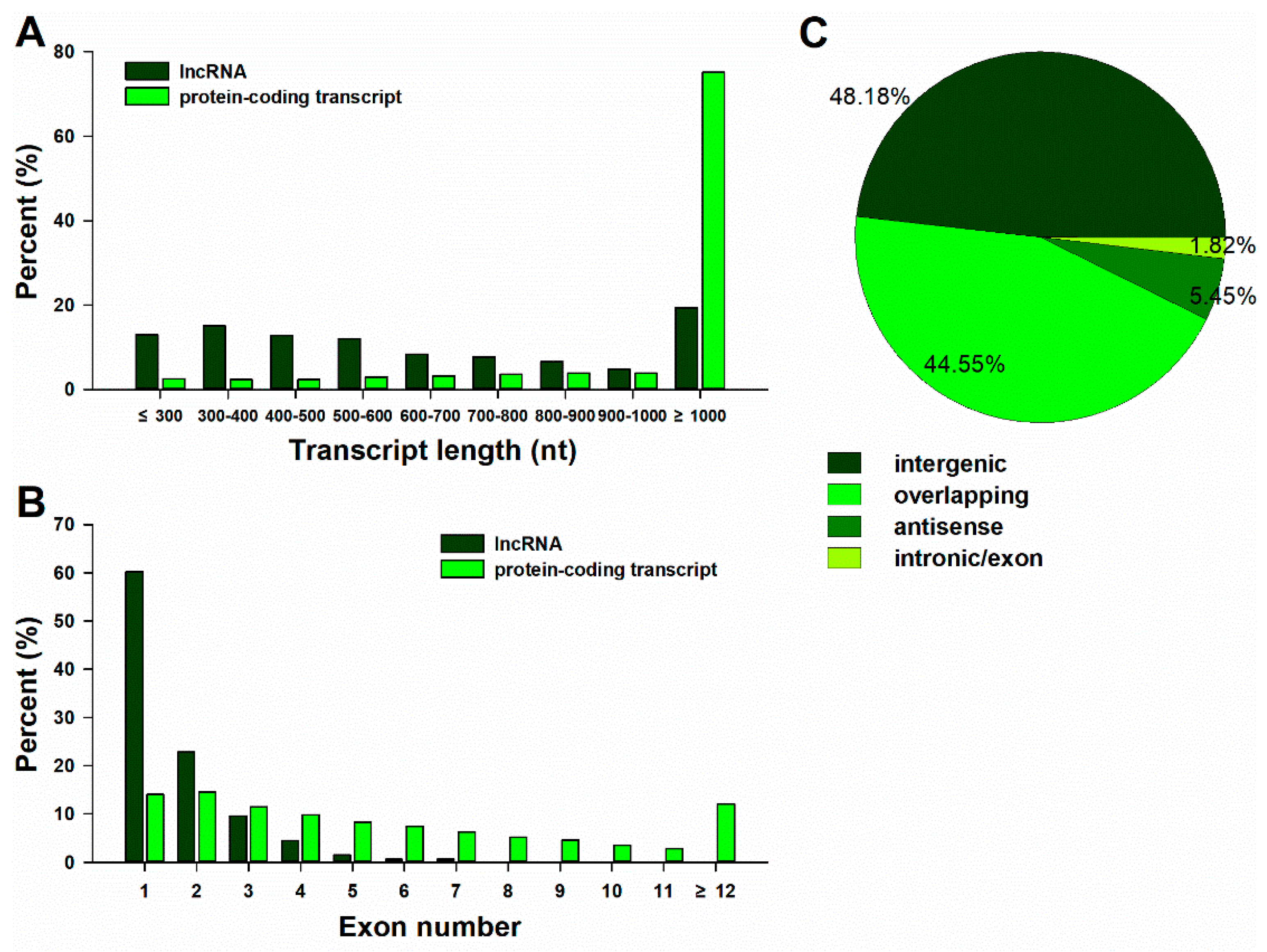
2.2. Identification and Characterization of lncRNAs and mRNAs in the Trifoliate Orange Transcriptome
2.3. Genomic Organization of Trifoliate Orange lncRNAs According to the Clementine Reference Sequence
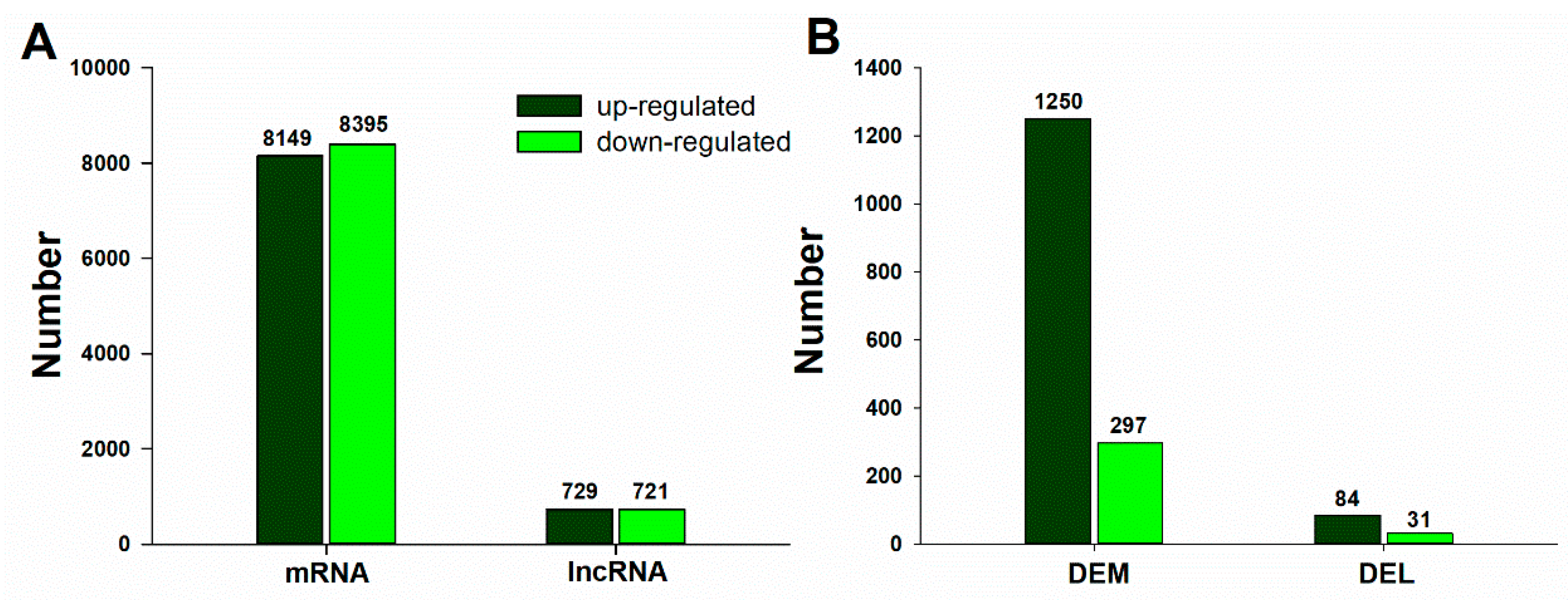
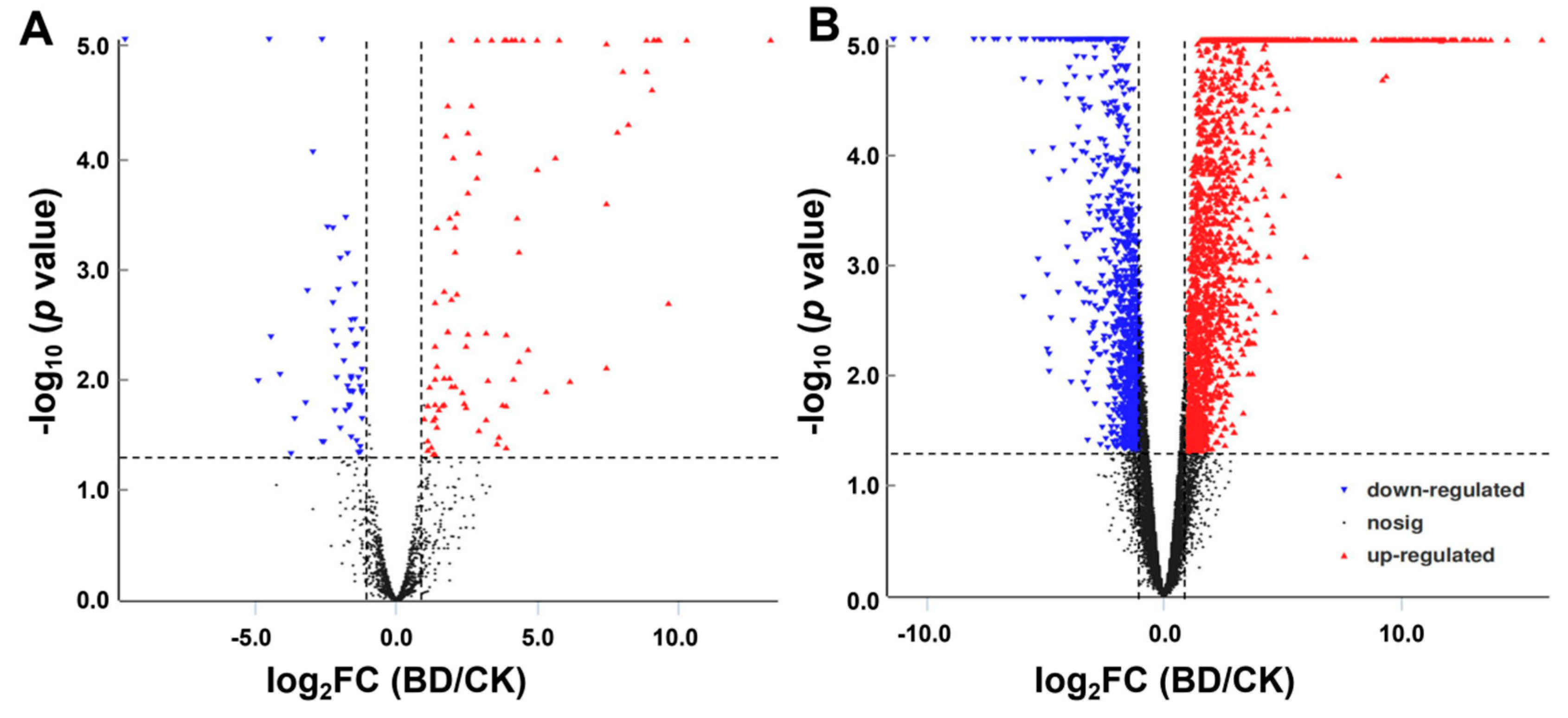
2.4. Differential Expression of mRNAs and lncRNAs in Response to B Deficiency
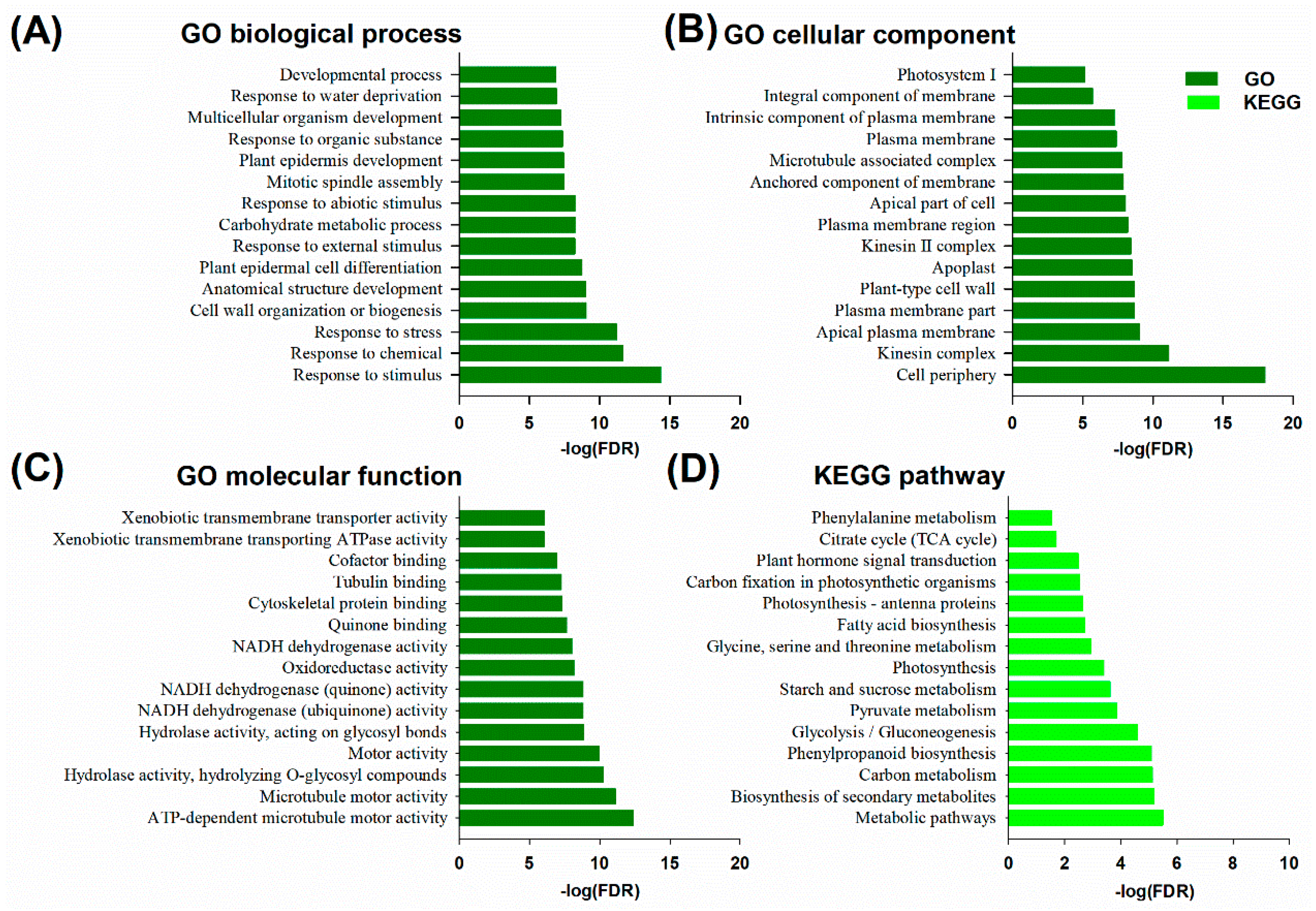
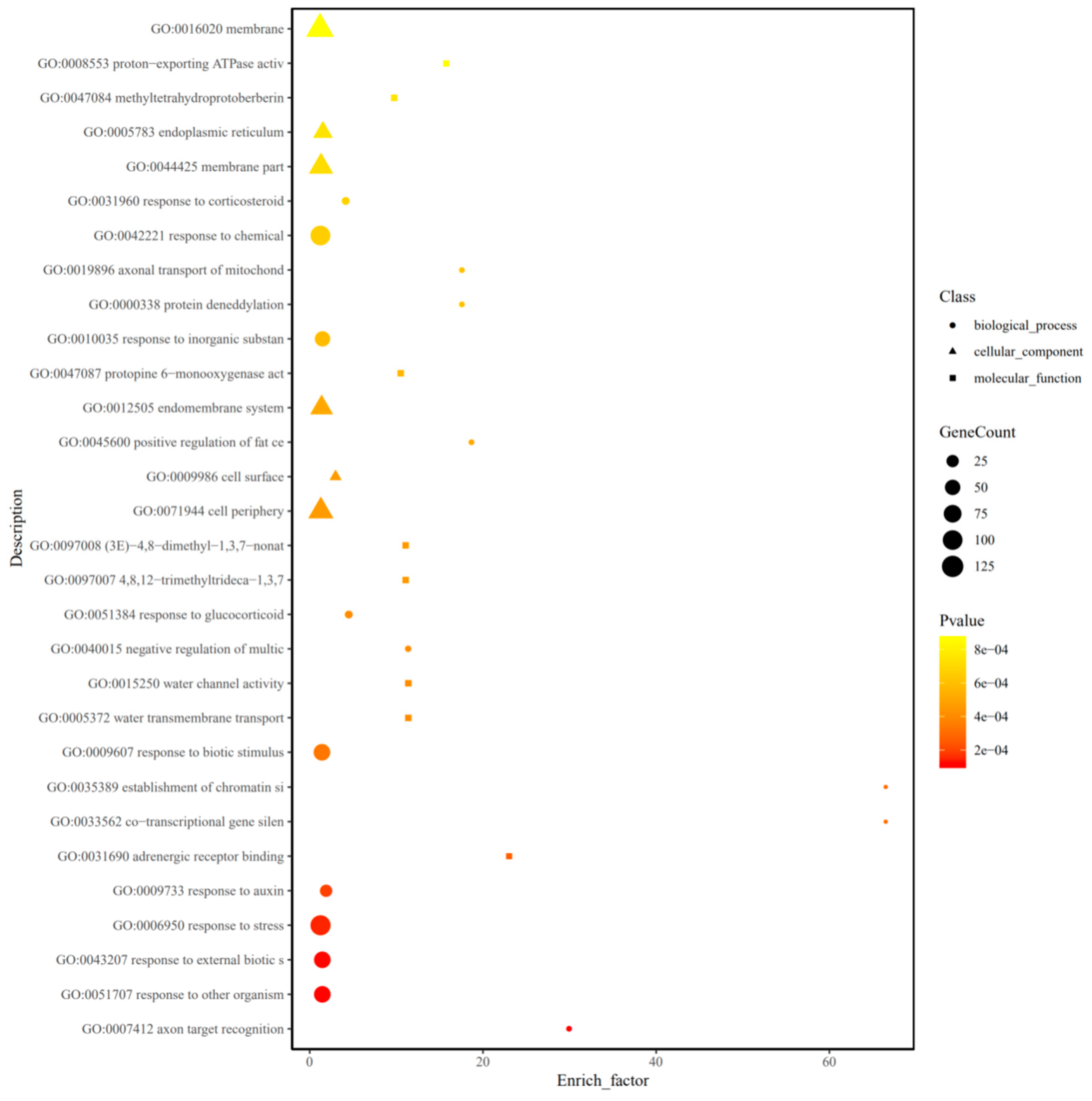
2.5. GO and KEGG Enrichment Analysis of DEM and DEL Target Genes
2.6. LncRNA-mRNA Interaction Network Analysis
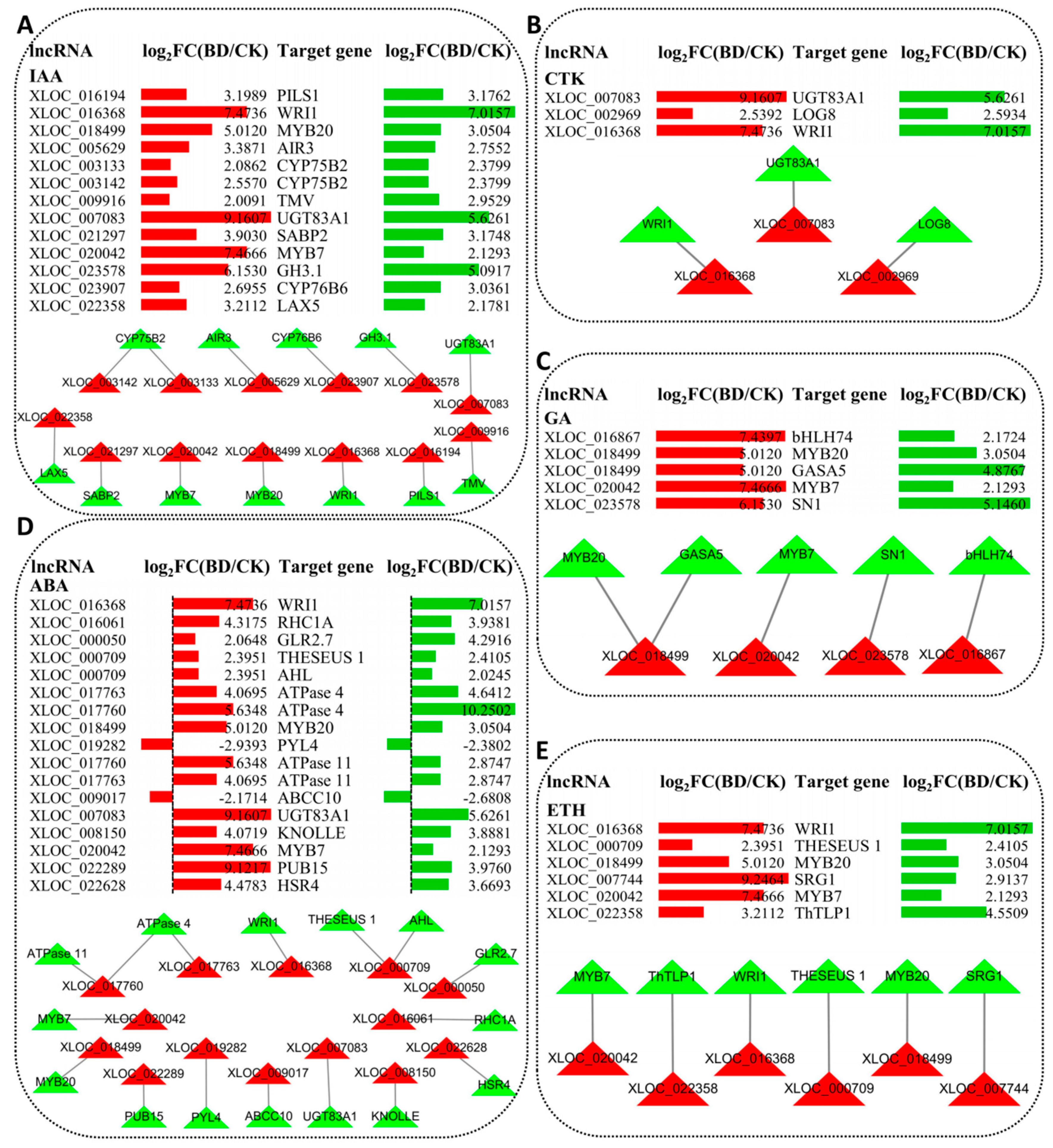
2.7. LncRNA Involved in Plant Hormone Biosynthesis and Signal Transduction
3. Discussion
3.1. The Expression of lncRNAs and mRNAs involved in Several Metabolic Pathway are Altered in Response to B Deficiency Stress
3.2. The role of Plant Hormones in Symptom Formation and Exacerbation
3.3. The Mechanism of lncRNAs Response to B Deficiency Stress
4. Materials and methods
4.1. Plant materials and B Deficiency Treatment
4.2. Determination of B and Lignin Concentration
4.3. Anatomical Analysis
4.4. RNA Isolation, lncRNA and mRNA Library Construction
4.5. Expression Analysis of lncRNAs and mRNA
4.6. GO and KEGG Pathway Analysis
4.7. Construction of lncRNA-mRNA Co-Expression Network
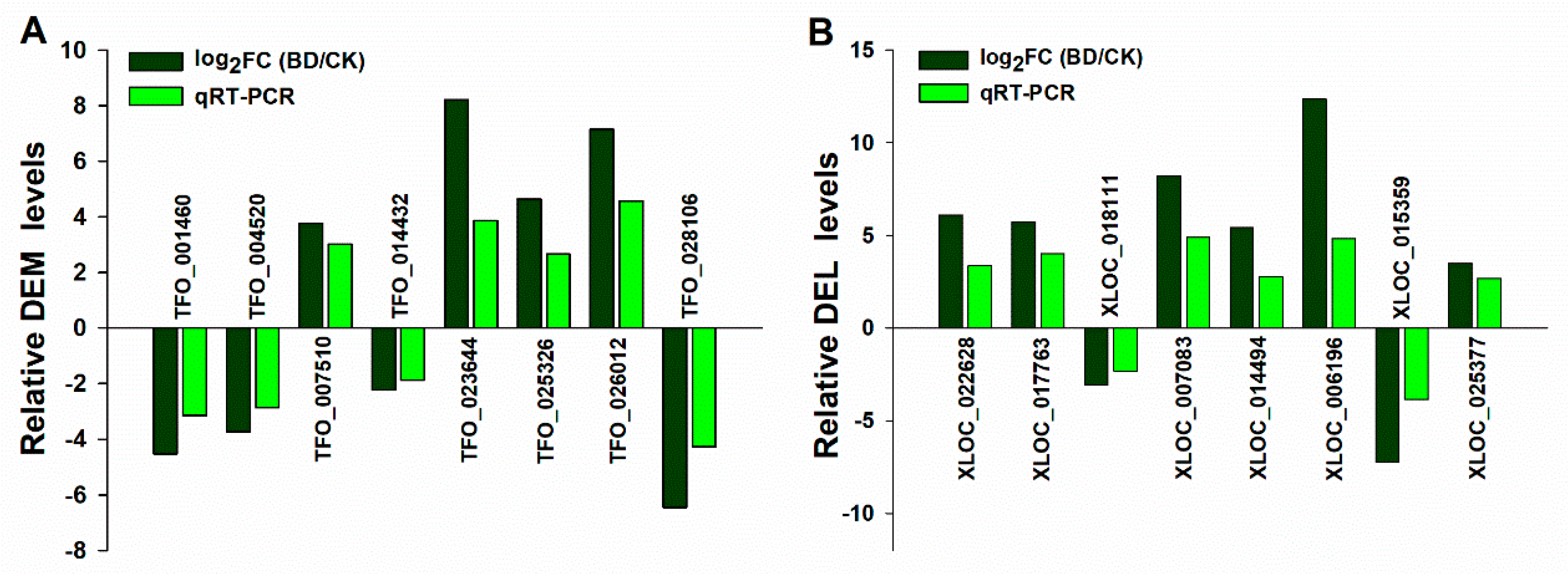
4.8. Quantitative RT-PCR
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 4CL | 4-Coumarate: CoA Ligase |
| ABA | Abscisic Acid |
| B | Boron |
| BD | Boron Deficiency |
| Ca2+ | Calcium |
| CCR | Cinnamoyl-CoA Reductase |
| CK | Control |
| CTK | Cytokinin |
| DEL | Differentially Expressed LncRNA |
| DEM | Differentially Expressed MRNA |
| ETH | Ethylene |
| GA | Gibberellin |
| GO | Gene Ontology |
| IAA | Auxin |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| lncRNA | Long Non-Coding RNA |
| ncRNA | Non-Coding RNA |
| PAL | Phenylalanine-Ammonium Lyase |
| POD | Peroxidase |
| qRT-PCR | Quantitative Real-Time PCR |
References
- Marschner, H. Mineral Nutrition of Higher Plants, 2nd ed.; Academic Press: London, UK, 1995. [Google Scholar]
- Cakmak, I.; Römheld, V. Boron deficiency-induced impairments of cellular functions in plants. Plant Soil 1997, 193, 71–83. [Google Scholar] [CrossRef]
- Camacho-Cristóbal, J.J.; Anzellotti, D.; González-Fontes, A. Changes in phenolic metabolism of tobacco plants during short-term boron deficiency. Plant Physiol. Biochem. 2002, 40, 997–1002. [Google Scholar] [CrossRef]
- Camacho-Cristóbal, J.J.; González-Fontes, A. Boron deficiency decreases plasmalemma H+-ATPase expression and nitrate uptake and promotes ammonium assimilation into asparagine in tobacco roots. Planta 2007, 226, 443–451. [Google Scholar] [CrossRef] [PubMed]
- Shorrocks, V.M. The occurrence and correction of boron deficiency. Plant Soil 1997, 193, 121–148. [Google Scholar] [CrossRef]
- O’Neill, M.A.; Ishii, T.; Albersheim, P.; Darvill, A.G. Rhamnogalacturonan II: Structure and function of a borate cross-linked cell wall pectic polysaccharide. Annu. Rev. Plant Biol. 2004, 55, 109–139. [Google Scholar] [CrossRef] [PubMed]
- Sheng, O.; Song, S.W.; Peng, S.A.; Deng, X.X. The effects of low boron on growth, gas exchange, boron concentration and distribution of ‘Newhall’ navel orange (Citrus sinensis Osb.) plants grafted on two rootstocks. Sci. Hortic. 2009, 121, 278–283. [Google Scholar] [CrossRef]
- Han, S.; Chen, L.S.; Jiang, H.X.; Smith, B.R.; Yang, L.T.; Xie, C.Y. Boron deficiency decreases growth and photosynthesis, and increases starch and hexoses in leaves of citrus seedlings. J. Plant Physiol. 2008, 165, 1331–1341. [Google Scholar] [CrossRef]
- Han, S.; Ning, T.; Jiang, H.X.; Yang, L.T.; Li, Y.; Chen, L.S. CO₂ assimilation, photosystem II photochemistry, carbohydrate metabolism and antioxidant system of citrus leaves in response to boron stress. Plant Sci. 2009, 176, 143–153. [Google Scholar] [CrossRef]
- Liu, G.D.; Jiang, C.C.; Wang, Y.H. Distribution of boron and its forms in young ‘Newhall’ navel orange (Citrus sinensis Osb.) plants grafted on two rootstocks in response to deficient and excessive boron. Soil Sci. Plant Nutr. 2011, 57, 93–104. [Google Scholar]
- Zhou, G.F.; Peng, S.A.; Liu, Y.Z.; Wei, Q.J.; Han, J.; Islam, M.Z. The physiological and nutritional responses of seven different citrus rootstock seedlings to boron deficiency. Trees 2014, 28, 295–307. [Google Scholar] [CrossRef]
- Zhou, G.F.; Liu, Y.Z.; Sheng, O.; Wei, Q.J.; Yang, C.Q.; Peng, S.A. Transcription profiles of boron-deficiency-responsive genes in citrus rootstock root by suppression subtractive hybridization and cDNA microarray. Front. Plant Sci. 2015, 5, 795. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.Q.; Liu, Y.Z.; An, J.C.; Li, S.; Jin, L.F.; Zhou, G.F.; Wei, Q.J.; Yan, H.Q.; Wang, N.N.; Fu, L.N.; et al. Digital gene expression analysis of corky split vein caused by boron deficiency in ‘Newhall’ Navel Orange (Citrus sinensis Osbeck) for selecting differentially expressed genes related to vascular hypertrophy. PLoS ONE 2013, 8, e65737. [Google Scholar] [CrossRef] [PubMed]
- Mei, L.; Sheng, O.; Peng, S.A.; Zhou, G.F.; Wei, Q.J. Growth, root morphology and boron uptake by citrus rootstock seedlings differing in boron-deficiency responses. Sci. Hortic. 2011, 129, 426–432. [Google Scholar] [CrossRef]
- Mei, L.; Li, Q.H.; Wang, H.; Sheng, O.; Peng, S.A. Boron deficiency affects root vessel anatomy and mineral nutrient allocation of poncirus trifoliata (L.) raf. Acta Physiol. Plant. 2016, 38, 1–8. [Google Scholar] [CrossRef]
- Amor, B.B.; Wirth, S.; Merchan, F.; Laporte, P.; d’Aubenton-Carafa, Y.; Hirsch, J.; Maizel, A.; Mallory, A.; Lucas, A.; Deragon, J.M.; et al. Novel long non-protein coding RNAs involved in Arabidopsis differentiation and stress responses. Genome Res. 2009, 19, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Bajic, V.B.; Zhang, Z. On the classifification of long noncoding RNAs. RNA Biol. 2013, 10, 925–933. [Google Scholar] [CrossRef]
- Rohrig, H.; Schmidt, J.; Miklashevichs, E.; Schell, J.; John, M. Soybean ENOD40 encodes two peptides that bind to sucrose synthase. Proc. Natl. Acad. Sci. USA 2002, 99, 1915–1920. [Google Scholar] [CrossRef]
- Mattick, J.S.; Rinn, J.L. Discovery and annotation of long noncoding RNAs. Nat. Struct. Mol. Biol. 2015, 22, 5–7. [Google Scholar] [CrossRef]
- Liu, X.; Hao, L.; Li, D.; Zhu, L.; Hu, S. Long non-coding RNAs and their biological roles in plants. Genom. Proteom. Bioinf. 2015, 13, 137–147. [Google Scholar] [CrossRef]
- Liu, J.; Wang, H.; Chua, N.H. Long non-coding RNA transcriptome of plants. Plant Biotechnol. J. 2015, 13, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Meng, X.; Dobrovolskaya, O.B.; Orlov, Y.L.; Chen, M. Non-coding RNAs and their roles in stress response in plants. Genom. Proteom. Bioinf. 2017, 15, 301. [Google Scholar] [CrossRef] [PubMed]
- Xuan, H.D.; Zhang, L.Z.; Liu, X.S.; Han, G.M.; Li, J.; Li, X.; Liu, A.G.; Liao, M.Z.; Zhang, S.H. PLNlncRbase: A resource for experimentally identified lncRNAs in plants. Gene 2015, 573, 328–332. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.W.; Zhou, X.H.; Wang, R.R.; Peng, W.L.; An, Y.; Chen, L.L. Functional analysis of long intergenic non-coding RNAs in phosphate-starved rice using competing endogenous RNA network. Sci. Rep. 2016, 6, 20715. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Wang, C.L.; Bao, H.; Chen, H.; Wang, Y.W. Genome-wide identification and characterization of novel lncRNAs in Populus under nitrogen deficiency. Mol. Gen. Genom. 2016, 291, 1663–1680. [Google Scholar] [CrossRef]
- Shin, S.Y.; Jeong, J.S.; Lim, J.Y.; Kim, T.; Park, J.H.; Kim, J.K.; Shin, C. Transcriptomic analyses of rice (oryza sativa) genes and non-coding RNAs under nitrogen starvation using multiple omics technologies. BMC Genom. 2018, 19, 532. [Google Scholar] [CrossRef]
- Yuan, J.; Zhang, Y.; Dong, J.; Sun, Y.; Lim, B.L.; Liu, D.; Lu, Z.J. Systematic characterization of novel lncRNAs responding to phosphate starvation in Arabidopsis thaliana. BMC Genom. 2016, 17, 655. [Google Scholar] [CrossRef]
- Huanca-Mamani, W.; Arias-Carrasco, R.; Cárdenas-Ninasivincha, S.; Rojas-Herrera, M.; Sepúlveda-Hermosilla, G.; Caris-Maldonado, J.C.; Bastías, E.; Maracaja-Coutinho, V. Long non-coding RNAs responsive to salt and boron stress in the hyper-arid lluteño maize from Atacama Desert. Genes 2018, 9, 170. [Google Scholar] [CrossRef]
- Franco-Zorrilla, J.M.; Valli, A.; Todesco, M.; Mateos, I.; Puga, M.I.; Rubio-Somoza, I.; Leyva, A.; Weigel, D.; Garcia, J.A.; Paz-Ares, J. Target mimicry provides a new mechanism for regulation of microRNA activity. Nat. Genet. 2007, 39, 1033–1037. [Google Scholar] [CrossRef]
- Bellaloui, N. Soybean seed phenol, lignin, and isoflavones and sugars composition altered by foliar boron application in soybean under water stress. Food Nutr. Sci. 2012, 3, 579–590. [Google Scholar] [CrossRef]
- Dell, B.; Huang, L.B. Physiological response of plants to low boron. Plant Soil 1997, 193, 103–120. [Google Scholar] [CrossRef]
- Lu, Y.B.; Yang, L.T.; Li, Y.; Xu, J.; Liao, T.T.; Chen, Y.B.; Chen, L.S. Effects of boron deficiency on major metabolites, key enzymes and gas exchange in leaves and roots of Citrus sinensis seedlings. Tree Physiol. 2014, 34, 608–618. [Google Scholar] [CrossRef]
- Lu, X.P.; Jiang, C.C.; Dong, X.C.; Wu, X.W.; Lei, Y. Leaf photosynthetic characteristics of citrus rootstocks with different boron efficiency. J. Plant Nutr. Fertil. 2017, 23, 476–483. [Google Scholar]
- Zhang, C.; Tang, G.; Peng, X.; Sun, F.; Liu, S.; Xi, Y. Long non-coding RNAs of switchgrass (panicum virgatum L.) in multiple dehydration stresses. BMC Plant Biol. 2018, 18, 79. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Chen, X.; Mu, M.; Wang, J.; Ye, W. Genome-wide analysis of long noncoding RNAs and their responses to drought stress in cotton (gossypium hirsutum L.). PLoS ONE 2016, 11, e0156723. [Google Scholar] [CrossRef] [PubMed]
- Eggert, K.; von Wirén, N. Response of the plant hormone network to boron deficiency. New Phytol. 2017, 216, 868–881. [Google Scholar] [CrossRef] [Green Version]
- Parr, A.J.; Loughman, B.C. Boron and membrane function in plants. In Metals and Micronutrients: Uptake and Utilization by Plants; Robb, D.A., Pierpoint, W.S., Eds.; Academic Press: New York, NY, USA, 1983; pp. 87–107. [Google Scholar]
- Li, Q.H.; Liu, Y.Z.; Pan, Z.Y.; Xie, S.; Peng, S.A. Boron deficiency alters root growth and development and interacts with auxin metabolism by influencing the expression of auxin synthesis and transport genes. Biotechnol. Biotec. Eq. 2016, 30, 661–668. [Google Scholar] [CrossRef]
- Scarpella, E.; Barkoulas, M.; Tsiantis, M. Control of Leaf and Vein Development by Auxin. CSH Perspect. Biol. 2010, 2, a001511. [Google Scholar] [CrossRef]
- Fukuda, H. Tracheary element differentiation. Plant Cell 1997, 9, 1147. [Google Scholar] [CrossRef]
- Matsumoto-Kitano, M.; Kusumoto, T.; Tarkowski, P.; Kinoshita-Tsujimura, K.; Václavíková, K.; Miyawaki, K.; Kakimoto, T. Cytokinins are central regulators of cambial activity. Proc. Natl. Acad. Sci. USA 2008, 105, 20027–20031. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, F.J.; Kieber, J.J. Cytokinin signaling. Curr. Opin. Plant Biol. 2005, 8, 518–525. [Google Scholar] [CrossRef]
- Muller, B.; Sheen, J. Advances in cytokinin signaling. Sci. Signal. 2007, 318, 68. [Google Scholar] [CrossRef]
- Koshiba, T.; Kobayashi, M.; Ishihara, A.; Matoh, T. Boron nutrition of cultured tobacco BY-2 cells. VI. Calcium is involved in early responses to boron deprivation. Plant Cell Physiol. 2010, 51, 323–327. [Google Scholar] [CrossRef]
- Quiles-Pando, C.; Rexach, J.; Navarro-Gochicoa, M.T.; Camacho-Cristóbal, J.J.; Herrera-Rodríguez, M.B.; González-Fontes, A. Boron deficiency increases the levels of cytosolic Ca2+ and expression of Ca2+-related genes in Arabidopsis thaliana roots. Plant Physiol. Biochem. 2013, 8, 55–60. [Google Scholar] [CrossRef]
- Quiles-Pando, C.; Navarro-Gochicoa, M.T.; Herrera-Rodríguez, M.B.; Camacho-Cristóbal, J.J.; González-Fontes, A.; Rexach, J. Boron Deficiency Increases Cytosolic Ca2+ Levels Mainly via Ca2+ Influx from the Apoplast in Arabidopsis thaliana Roots. Int. J. Mol. Sci. 2019, 20, 2297. [Google Scholar] [CrossRef]
- González-Fontes, A.; Navarro-Gochicoa, M.T.; Camacho-Cristóbal, J.J.; Herrera-Rodríguez, M.B.; Quiles-Pando, C.; Rexach, J. Is Ca2+ involved in the signal transduction pathway of boron deficiency? new hypotheses for sensing boron deprivation. Plant Sci. 2014, 217–218, 135–139. [Google Scholar] [CrossRef]
- Wang, X.T.; Arai, S.; Song, X.Y.; Reichart, D.; Du, K.; Pascual, G.; Tempst, P.; Rosenfeld, M.G.; Glass, C.K.; Kurokawa, R. Induced ncRNAs allosterically modify RNA-binding proteins in cis to inhibit transcription. Nature 2008, 454, 126–130. [Google Scholar] [CrossRef]
- Shamovsky, I.; Ivannikov, M.; Kandel, E.S.; Gershon, D.; Nudler, E. RNA-mediated response to heat shock in mammalian cells. Nature 2006, 440, 556–560. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.S. The water culture method for growing plants without soil. Cal. Agric. Exp. St. Circ. 1950, 347, 305–311. [Google Scholar]
- Bruce, R.J.; West, C.A. Elicitation of lignin biosynthesis and isoperoxidase activity by pectic fragments in suspension cultures of castor bean. Plant Physiol. 1989, 91, 889–897. [Google Scholar] [CrossRef]
- Wang, C.Y.; Liu, S.R.; Zhang, X.Y.; Ma, Y.J.; Hu, C.G.; Zhang, J.Z. Genome-wide screening and characterization of long non-coding rnas involved in flowering development of trifoliate orange (poncirus trifoliata l. raf.). Sci. Rep. 2017, 7, 43226. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Summary | Control | Boron Deficiency | ||||
|---|---|---|---|---|---|---|
| CK_1 | CK_2 | CK_3 | BD_1 | BD_2 | BD_3 | |
| Raw reads | 106,350,380 | 128,309,312 | 119,745,274 | 113,804,668 | 100,898,426 | 102,976,228 |
| Clean reads | 105,992,604 | 127,931,954 | 118,953,032 | 113,440,572 | 100,542,180 | 102,659,450 |
| Clean reads rate (%) | 99.66 | 99.71 | 99.34 | 99.68 | 99.65 | 99.69 |
| Q20% | 97.81 | 97.83 | 97.62 | 97.71 | 97.70 | 97.82 |
| Q30% | 94.43 | 94.44 | 93.98 | 94.16 | 92.20 | 94.43 |
| GC% | 42.24 | 42.57 | 42.66 | 42.75 | 43.08 | 43.03 |
| Mapped reads | 30,810,136 | 30,324,690 | ||||
| Unique mapped reads | 22,371,249 | 25,678,257 | ||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, G.-F.; Zhang, L.-P.; Li, B.-X.; Sheng, O.; Wei, Q.-J.; Yao, F.-X.; Guan, G.; Liu, G.-D. Genome-Wide Identification of Long Non-coding RNA in Trifoliate Orange (Poncirus trifoliata (L.) Raf) Leaves in Response to Boron Deficiency. Int. J. Mol. Sci. 2019, 20, 5419. https://doi.org/10.3390/ijms20215419
Zhou G-F, Zhang L-P, Li B-X, Sheng O, Wei Q-J, Yao F-X, Guan G, Liu G-D. Genome-Wide Identification of Long Non-coding RNA in Trifoliate Orange (Poncirus trifoliata (L.) Raf) Leaves in Response to Boron Deficiency. International Journal of Molecular Sciences. 2019; 20(21):5419. https://doi.org/10.3390/ijms20215419
Chicago/Turabian StyleZhou, Gao-Feng, Li-Ping Zhang, Bi-Xian Li, Ou Sheng, Qing-Jiang Wei, Feng-Xian Yao, Guan Guan, and Gui-Dong Liu. 2019. "Genome-Wide Identification of Long Non-coding RNA in Trifoliate Orange (Poncirus trifoliata (L.) Raf) Leaves in Response to Boron Deficiency" International Journal of Molecular Sciences 20, no. 21: 5419. https://doi.org/10.3390/ijms20215419
APA StyleZhou, G. -F., Zhang, L. -P., Li, B. -X., Sheng, O., Wei, Q. -J., Yao, F. -X., Guan, G., & Liu, G. -D. (2019). Genome-Wide Identification of Long Non-coding RNA in Trifoliate Orange (Poncirus trifoliata (L.) Raf) Leaves in Response to Boron Deficiency. International Journal of Molecular Sciences, 20(21), 5419. https://doi.org/10.3390/ijms20215419