Provitamin A Biofortification of Durum Wheat through a TILLING Approach

 ,
, 

 and
and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
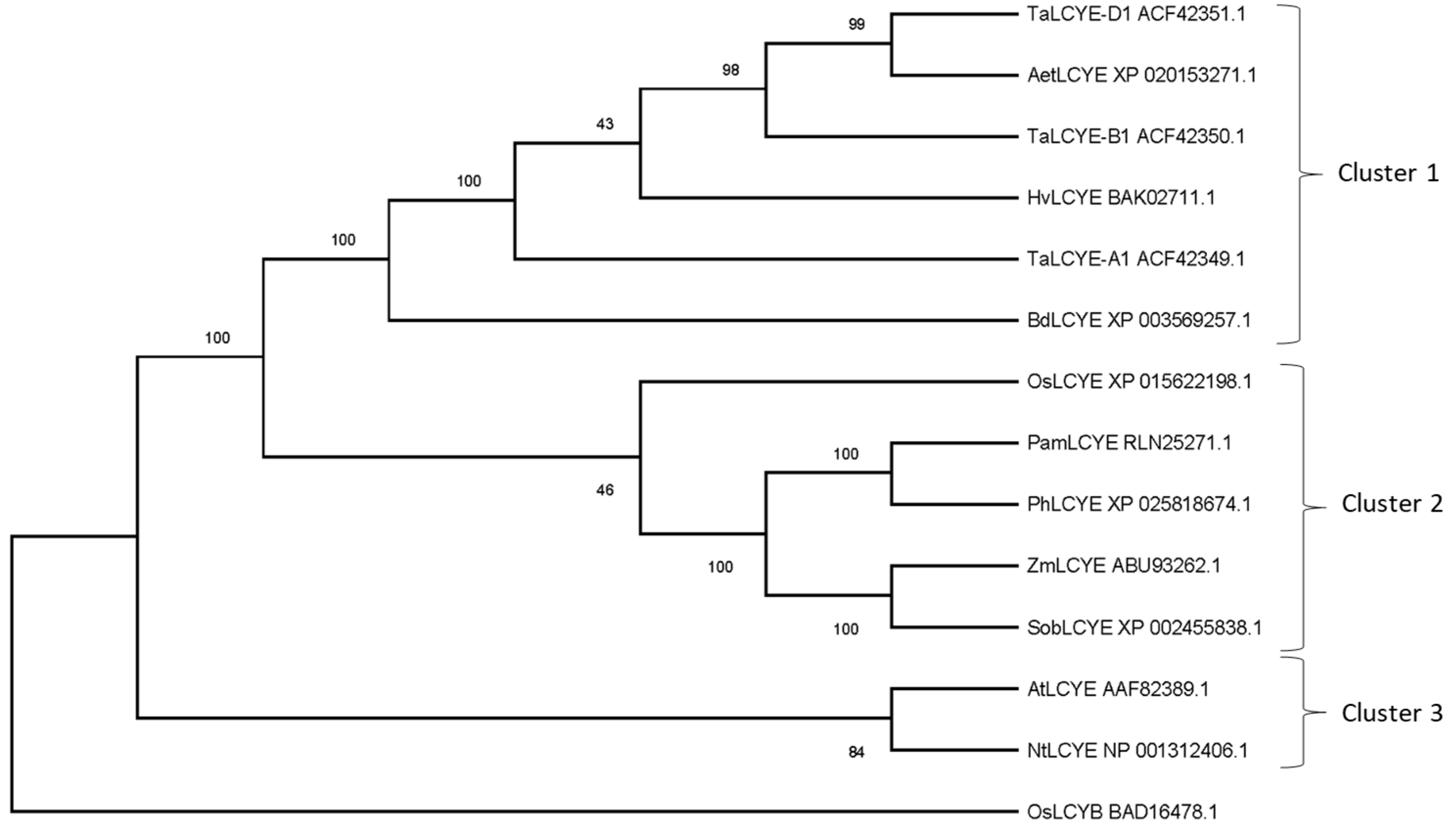
2.1. Comparison of LCYE Amino Acid Sequences between Wheat and Other Species
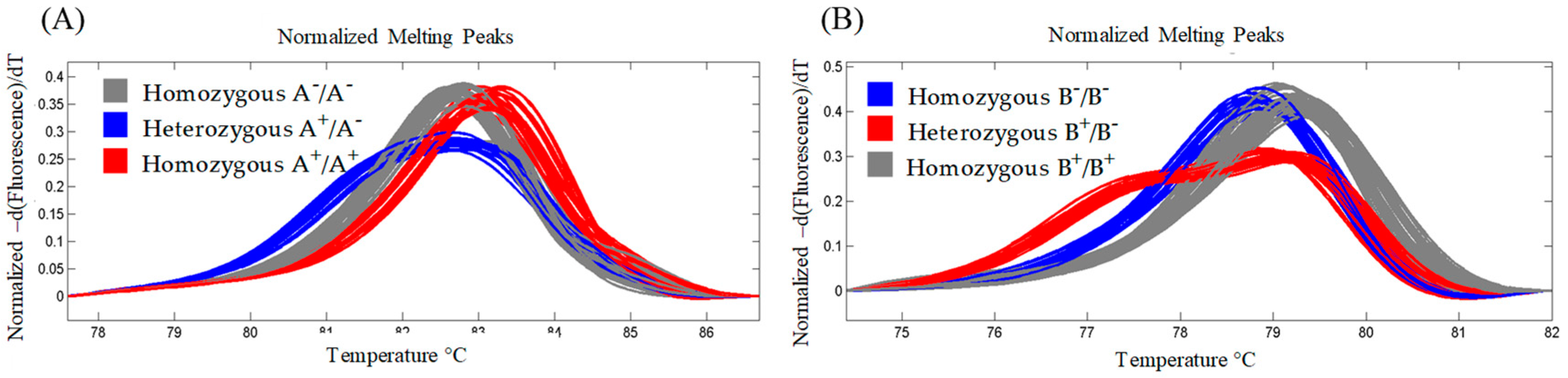
2.2. Identification of TILLING Durum Wheat Lines with Knockout Mutation on the lcyE Homeoalleles
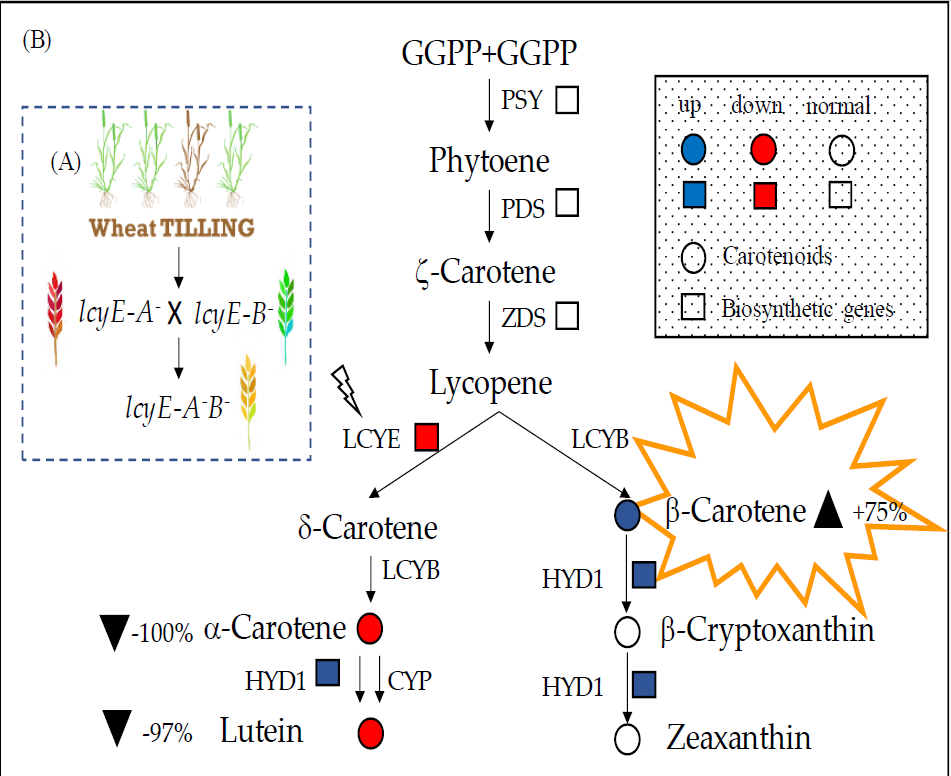
2.3. Pyramiding the LCYE Null Mutations in the Durum Wheat cv. Kronos
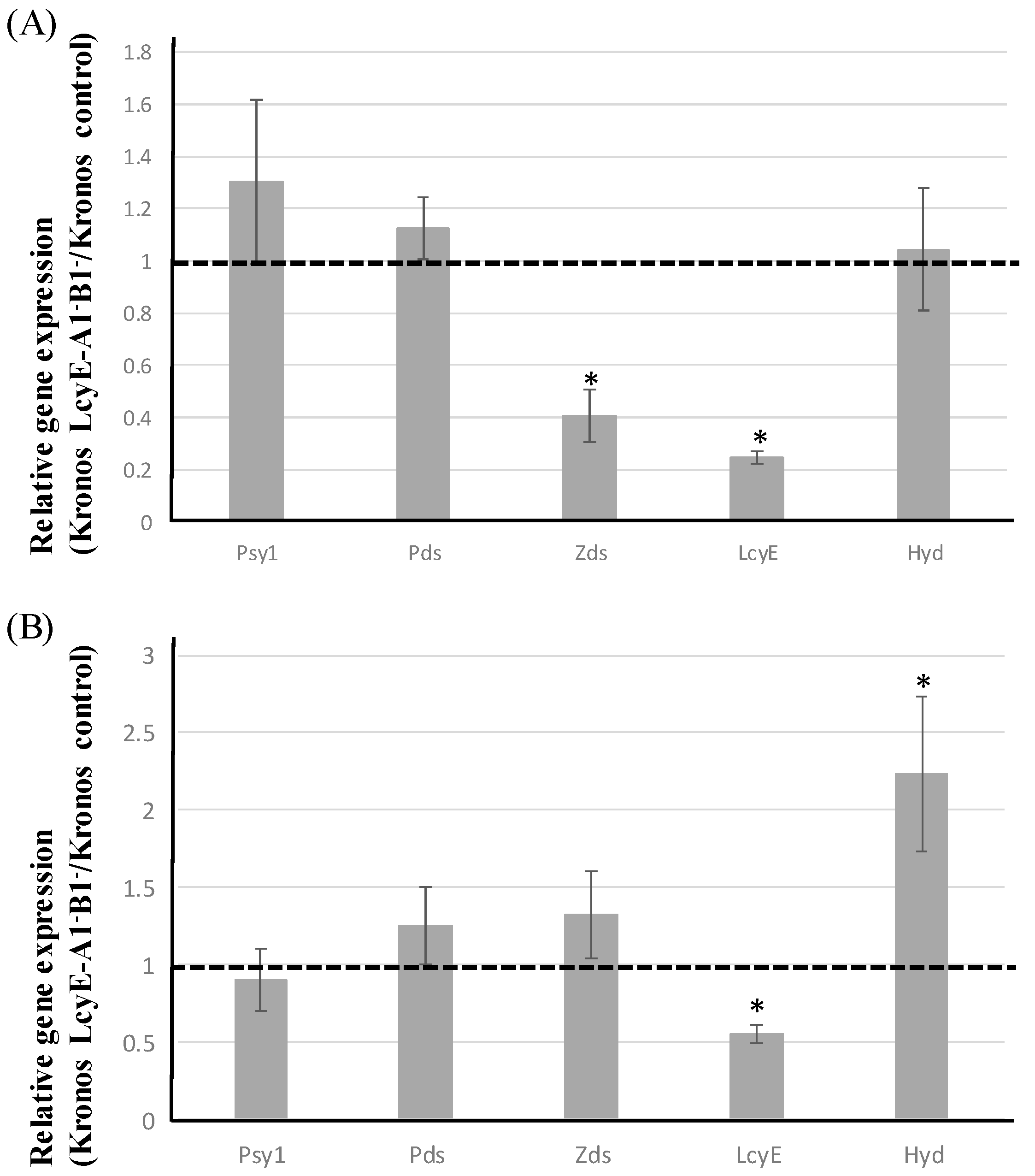
2.4. Expression Analysis of Major Genes Involved in Carotenoid Biosynthesis
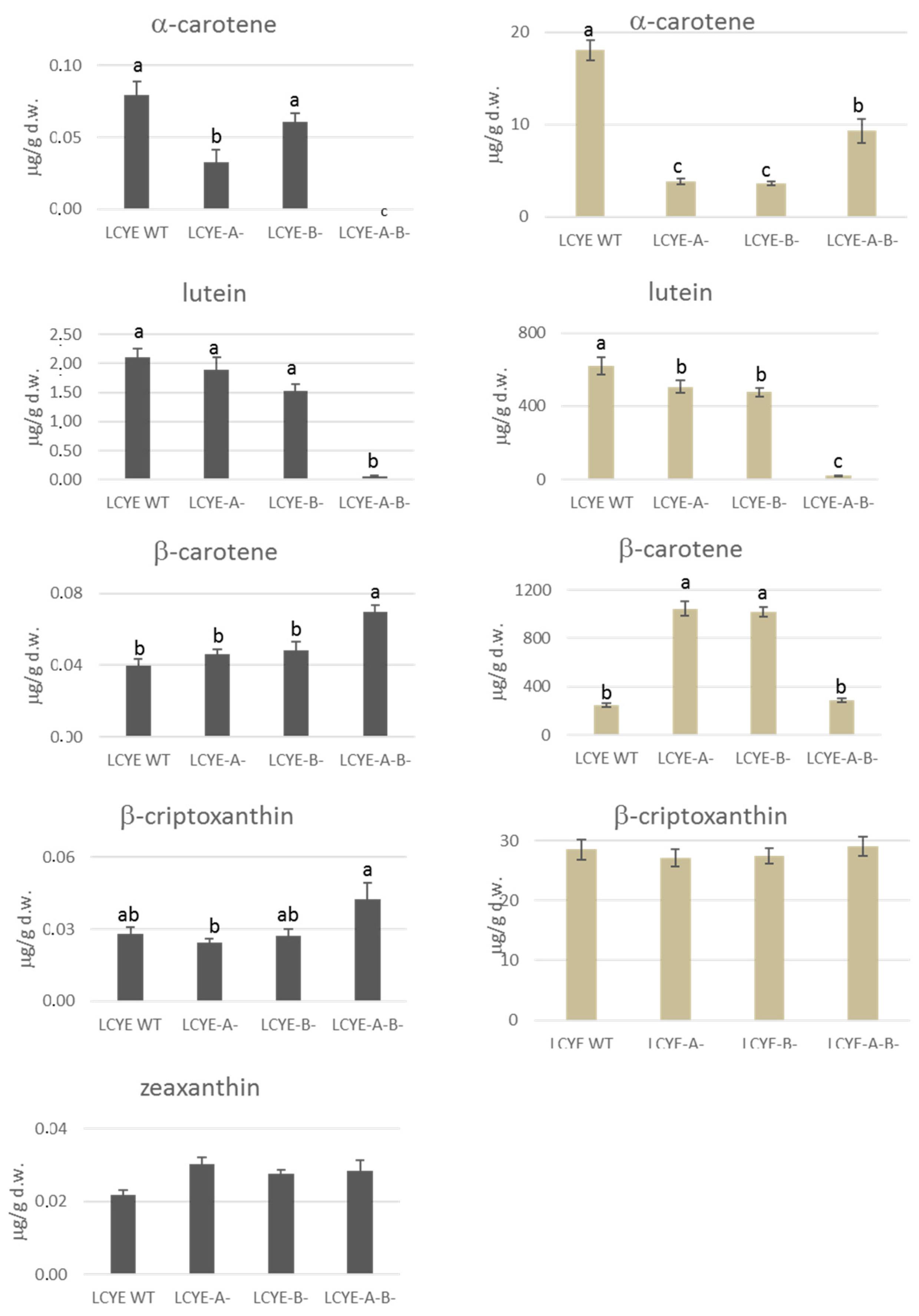
2.5. Carotenoids Evaluation in Grain and Leaves
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Isolation of LCYE Sequence and Their Phylogenesis
4.3. High-Resolution Melting Genotyping
4.4. Quantitative Real Time-PCR (qRT-PCR)
4.5. DNA Sequencing
4.6. Carotenoid Extraction from Seeds and Leaves
4.7. Carotenoid Analysis by HPLC- Diode Array Detector (DAD)
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| WHO | World Health Organization |
| VAD | Vitamin A deficiency |
| MEP | Methyl-erythrose 4-phosphate |
| GGPP | Geranylgeranyl pyrophosphate |
| PSY | Phytoene synthase |
| PDS | Phytoene desaturase |
| ZDS | ζ-carotene desaturase |
| ZISO | ζ-carotene isomerase |
| CRTISO | Carotene isomerase |
| LCYB | Lycopene β-cyclase |
| LCYE | Lycopene ε-cyclase |
| HYD | β-ring hydroxylases |
| TILLING | Targeting Induced Local Lesions in Genomes |
| qRT-PCR | Quantitative real-time PCR |
References
- World Health Organization. The State of Food Security and Nutrition in the World 2018: Building Climate Resilience for Food Security and Nutrition. Available online: https://www.wfp.org/publications/2018-state-food-security-and-nutrition-world-sofi-report (accessed on 13 November 2019).
- Mayer, J.E.; Pfeiffer, W.H.; Beyer, P. Biofortified crops to alleviate micronutrient malnutrition. Curr. Opin. Plant Biol. 2008, 11, 166–170. [Google Scholar] [CrossRef]
- Ahmad, A.; Ahmed, Z. Fortification in Beverages. In Production and Management of Beverages, 1st ed.; Grumezescu, A., Holban, A.M., Eds.; Woodhead Publishing: Oxford, UK, 2019; pp. 85–122. [Google Scholar]
- Song, P.; Wang, J.; Wei, W.; Chang, X.; Wang, M.; An, L. The prevalence of vitamin A deficiency in Chinese children: A systematic review and Bayesian meta-analysis. Nutrients 2017, 9, 1285. [Google Scholar] [CrossRef]
- Imdad, A.; Yakoob, M.Y.; Sudfeld, C.; Haider, B.A.; Black, R.E.; Bhutta, Z.A. Impact of vitamin A supplementation on infant and childhood mortality. BMC Public Health 2011, 11, S20. [Google Scholar] [CrossRef]
- Underwood, B.A. Hypovitaminosis A and its control. Bull. World Health Organ. 1978, 56, 525. [Google Scholar]
- Fairfield, K.M.; Fletcher, R.H. Vitamins for chronic disease prevention in adults: Scientific review. J. Am. Med. Assoc. 2002, 287, 3116–3126. [Google Scholar] [CrossRef]
- Zhai, S.; Xia, X.; He, Z. Carotenoids in staple cereals: Metabolism, regulation, and genetic manipulation. Front. Plant Sci. 2016, 7, 1197. [Google Scholar] [CrossRef]
- Nisar, N.; Li, L.; Lu, S.; Khin, N.C.; Pogson, B.J. Carotenoid metabolism in plants. Mol. Plant 2015, 8, 68–82. [Google Scholar] [CrossRef]
- Farré, G.; Sanahuja, G.; Naqvi, S.; Bai, C.; Capell, T.; Zhu, C.; Christou, P. Travel advice on the road to carotenoids in plants. Plant Sci. 2010, 179, 28–48. [Google Scholar] [CrossRef]
- Giuliano, G. Provitamin A biofortification of crop plants: A gold rush with many miners. Curr. Opin. Biotechnol. 2017, 44, 169–180. [Google Scholar] [CrossRef]
- Giuliano, G.; Tavazza, R.; Diretto, G.; Beyer, P.; Taylor, M.A. Metabolic engineering of carotenoid biosynthesis in plants. Trends Biotechnol. 2008, 26, 139–145. [Google Scholar] [CrossRef]
- Bai, C.; Rivera, S.M.; Medina, V.; Alves, R.; Vilaprinyo, E.; Sorribas, A.; Canela, R.; Capell, T.; Sandmann, G.; Christou, P.; et al. An in vitro system for the rapid functional characterization of genes involved in carotenoid biosynthesis and accumulation. Plant J. 2014, 77, 464–475. [Google Scholar] [CrossRef]
- Römer, S.; Lübeck, J.; Kauder, F.; Steiger, S.; Adomat, C.; Sandmann, G. Genetic engineering of a zeaxanthin-rich potato by antisense inactivation and co-suppression of carotenoid epoxidation. Metab. Eng. 2002, 4, 263–272. [Google Scholar] [CrossRef]
- Zeng, J.; Wang, X.; Miao, Y.; Wang, C.; Zang, M.; Chen, X.; Li, M.; Li, X.; Wang, Q.; Li, K.; et al. Metabolic engineering of wheat provitamin A by simultaneously overexpressing CrtB and silencing carotenoid hydroxylase (TaHYD). J. Agric. Food Chem. 2015, 63, 9083–9092. [Google Scholar] [CrossRef]
- Galpaz, N.; Wang, Q.; Menda, N.; Zamir, D.; Hirschberg, J. Abscisic acid deficiency in the tomato mutant high-pigment 3 leading to increased plastid number and higher fruit lycopene content. Plant J. 2008, 53, 717–730. [Google Scholar] [CrossRef]
- Carrera, A.; Echenique, V.; Zhang, W.; Carrera, A.; Echenique, V.; Zhang, W.; Helguera, M.; Manthey, F.; Schrager, A.; Picca, A.; et al. A deletion at the Lpx-B1 locus is associated with low lipoxygenase activity and improved pasta color in durum wheat (Triticum turgidum ssp. durum). J. Cereal Sci. 2007, 45, 67–77. [Google Scholar] [CrossRef]
- Che, P.; Zhao, Z.Y.; Glassman, K.; Dolde, D.; Hu, T.X.; Jones, T.J.; Gruis, D.F.; Obukosia, S.; Wambugu, F.; Albertsen, M.C. Elevated vitamin E content improves all-trans β-carotene accumulation and stability in biofortified sorghum. Proc. Natl. Acad. Sci. USA 2016, 113, 11040–11045. [Google Scholar] [CrossRef]
- McCallum, C.M.; Comai, L.; Greene, E.A.; Henikoff, S. Targeting induced local lesions in genomes (TILLING) for plant functional genomics. Plant Physiol. 2000, 123, 439–442. [Google Scholar] [CrossRef]
- Krasileva, K.V.; Vasquez-Gross, H.A.; Howell, T.; Bailey, P.; Paraiso, F.; Clissold, L.; Simmonds, J.; Ramirez-Gonzalez, R.H.; Wang, X.; Borrill, P.; et al. Uncovering hidden variation in polyploid wheat. Proc. Natl. Acad. Sci. USA 2017, 114, E913–E921. [Google Scholar] [CrossRef]
- Alquézar, B.; Zacarías, L.; Rodrigo, M.J. Molecular and functional characterization of a novel chromoplast-specific lycopene β-cyclase from Citrus and its relation to lycopene accumulation. J. Exp. Bot. 2009, 60, 1783–1797. [Google Scholar] [CrossRef]
- Cunningham, F.X.; Pogson, B.; Sun, Z.; McDonald, K.A.; DellaPenna, D.; Gantt, E. Functional analysis of the beta and epsilon lycopene cyclase enzymes of Arabidopsis reveals a mechanism for control of cyclic carotenoid formation. Plant Cell 1996, 8, 1613–1626. [Google Scholar] [CrossRef]
- Krubasik, P.; Sandmann, G. Molecular evolution of lycopene cyclases involved in the formation of carotenoids with ionone end groups. Biochem. Soc. Trans. 2000, 28, 806–810. [Google Scholar] [CrossRef]
- Richaud, D.; Stange, C.; Gadaleta, A.; Colasuonno, P.; Parada, R.; Schwember, A.R. Identification of Lycopene epsilon cyclase (LCYE) gene mutants to potentially increase β-carotene content in durum wheat (Triticum turgidum L. ssp. durum) through TILLING. PLoS ONE 2018, 13, e0208948. [Google Scholar] [CrossRef]
- Talsma, E.F.; Melse-Boonstra, A.; Brouwer, I.D. Acceptance and adoption of biofortified crops in low-and middle-income countries: A systematic review. Nutr. Rev. 2017, 75, 798–829. [Google Scholar] [CrossRef]
- Palmer, A.C.; Siamusantu, W.; Chileshe, J.; Schulze, K.J.; Barffour, M.; Craft, N.E.; Molobeka, N.; Kalungwana, N.; Arguello, M.A.; Mitra, M.; et al. Provitamin A–biofortified maize increases serum β-carotene, but not retinol, in marginally nourished children: A cluster-randomized trial in rural Zambia. Am. J. Clin. Nutr. 2016, 104, 181–190. [Google Scholar] [CrossRef]
- Ye, X.; Al-Babili, S.; Klöti, A.; Zhang, J.; Lucca, P.; Beyer, P.; Potrykus, I. Engineering the provitamin A (β-carotene) biosynthetic pathway into (carotenoid-free) rice endosperm. Science 2000, 287, 303–305. [Google Scholar] [CrossRef]
- Diretto, G.; Al-Babili, S.; Tavazza, R.; Papacchioli, V.; Beyer, P.; Giuliano, G. Metabolic engineering of potato carotenoid content through tuber-specific overexpression of a bacterial mini-pathway. PLoS ONE 2007, 2, e350. [Google Scholar] [CrossRef]
- Wang, C.; Zeng, J.; Li, Y.; Hu, W.; Chen, L.; Miao, Y.J.; Deng, P.Y.; Yuan, C.H.; Ma, C.; Chen, X.; et al. Enrichment of provitamin A content in wheat (Triticum aestivum L.) by introduction of the bacterial carotenoid biosynthetic genes CrtB and CrtI. J. Exp. Bot. 2014, 65, 2545–2556. [Google Scholar] [CrossRef]
- Zhu, C.; Naqvi, S.; Breitenbach, J.; Sandmann, G.; Christou, P.; Capell, T. Combinatorial genetic transformation generates a library of metabolic phenotypes for the carotenoid pathway in maize. Proc. Natl. Acad. Sci. USA 2008, 105, 18232–18237. [Google Scholar] [CrossRef]
- Sestili, F.; Botticella, E.; Lafiandra, D. TILLING for improved starch composition in wheat. In Genomics of Plant Genetic Resources; Tuberosa, R., Graner, A., Frison, E., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 467–487. [Google Scholar]
- Slade, A.J.; Fuerstenberg, S.I.; Loeffler, D.; Steine, M.N.; Facciotti, D. A reverse genetic, nontransgenic approach to wheat crop improvement by TILLING. Nat. Biotechnol. 2005, 23, 75. [Google Scholar] [CrossRef]
- Parry, M.A.; Madgwick, P.J.; Bayon, C.; Tearall, K.; Hernandez-Lopez, A.; Baudo, M.; Rakszegi, M.; Hamada, W.; l-Yassin, A.; Ouabbou, H.; et al. Mutation discovery for crop improvement. J. Exp. Bot. 2009, 60, 2817–2825. [Google Scholar] [CrossRef]
- Uauy, C.; Paraiso, F.; Colasuonno, P.; Tran, R.K.; Tsai, H.; Berardi, S.; Comai, L.; Dubcovsky, J. A modified TILLING approach to detect induced mutations in tetraploid and hexaploid wheat. BMC Plant Biol. 2009, 9, 115. [Google Scholar] [CrossRef]
- Dong, C.; Dalton-Morgan, J.; Vincent, K.; Sharp, P. A modified TILLING method for wheat breeding. Plant Gen. 2009, 2, 39–47. [Google Scholar] [CrossRef] [Green Version]
- Sestili, F.; Botticella, E.; Bedo, Z.; Phillips, A.; Lafiandra, D. Production of novel allelic variation for genes involved in starch biosynthesis through mutagenesis. Mol. Breed. 2010, 25, 145. [Google Scholar] [CrossRef]
- Bovina, R.; Brunazzi, A.; Gasparini, G.; Sestili, F.; Palombieri, S.; Botticella, E.; Lafiandra, D.; Mantovani, P.; Massi, A. Development of a TILLING resource in durum wheat for reverse-and forward-genetic analyses. Crop Pasture Sci. 2014, 65, 112–124. [Google Scholar] [CrossRef]
- Uauy, C.; Wulff, B.B.; Dubcovsky, J. Combining traditional mutagenesis with new high-throughput sequencing and genome editing to reveal hidden variation in polyploid wheat. Ann. Rev. Genet. 2017, 51, 435–454. [Google Scholar] [CrossRef]
- Wicker, T.; Taudien, S.; Houben, A.; Keller, B.; Graner, A.; Platzer, M.; Stein, N. A whole-genome snapshot of 454 sequences exposes the composition of the barley genome and provides evidence for parallel evolution of genome size in wheat and barley. Plant J. 2009, 59, 712–722. [Google Scholar] [CrossRef] [Green Version]
- Harjes, C.E.; Rocheford, T.R.; Bai, L.; Brutnell, T.P.; Kandianis, C.B.; Sowinski, S.G.; Stapleton, A.E.; Vallabhaneni, R.; Williams, M.; Wurtzel, E.T.; et al. Natural genetic variation in lycopene epsilon cyclase tapped for maize biofortification. Science 2008, 319, 330–333. [Google Scholar] [CrossRef] [Green Version]
- Diretto, G.; Tavazza, R.; Welsch, R.; Pizzichini, D.; Mourgues, F.; Papacchioli, V.; Beyer, P.; Giuliano, G. Metabolic engineering of potato tuber carotenoids through tuber-specific silencing of lycopene epsilon cyclase. BMC Plant Biol. 2006, 6, 13. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.H.; Kim, Y.H.; Ahn, Y.O.; Ahn, M.J.; Jeong, J.C.; Lee, H.S.; Kwak, S.S. Downregulation of the lycopene ϵ-cyclase gene increases carotenoid synthesis via the β-branch-specific pathway and enhances salt-stress tolerance in sweetpotato transgenic calli. Physiol. Plant. 2013, 147, 432–442. [Google Scholar] [CrossRef]
- Yu, B.; Lydiate, D.J.; Young, L.W.; Schäfer, U.A.; Hannoufa, A. Enhancing the carotenoid content of Brassica napus seeds by downregulating lycopene epsilon cyclase. Transgenic Res. 2008, 17, 573–585. [Google Scholar] [CrossRef]
- Digesù, A.M.; Platani, C.; Cattivelli, L.; Mangini, G.; Blanco, A. Genetic variability in yellow pigment components in cultivated and wild tetraploid wheats. J. Cereal Sci. 2009, 50, 210–218. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Botticella, E.; Sestili, F.; Hernandez-Lopez, A.; Phillips, A.; Lafiandra, D. High resolution melting analysis for the detection of EMS induced mutations in wheat Sbella genes. BMC Plant Biol. 2011, 11, 156. [Google Scholar] [CrossRef] [Green Version]
- Sestili, F.; Palombieri, S.; Botticella, E.; Mantovani, P.; Bovina, R.; Lafiandra, D. TILLING mutants of durum wheat result in a high amylose phenotype and provide information on alternative splicing mechanisms. Plant Sci. 2015, 233, 127–133. [Google Scholar] [CrossRef]
- Camerlengo, F.; Sestili, F.; Silvestri, M.; Colaprico, G.; Margiotta, B.; Ruggeri, R.; Lupi, R.; Masci, S.; Lafiandra, D. Production and molecular characterization of bread wheat lines with reduced amount of α-type gliadins. BMC Plant Biol. 2017, 17, 248. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Rodriguez-Amaya, D.B.; Kimura, M. HarvestPlus handbook for carotenoid analysis. In HarvestPlus Technical Monograph 2; International Food Policy Research Institute (IFPRI) and International Center for Tropical Agriculture (CIAT): Washington, DC, USA, 2004. [Google Scholar]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sestili, F.; Garcia-Molina, M.D.; Gambacorta, G.; Beleggia, R.; Botticella, E.; De Vita, P.; Savatin, D.V.; Masci, S.; Lafiandra, D. Provitamin A Biofortification of Durum Wheat through a TILLING Approach. Int. J. Mol. Sci. 2019, 20, 5703. https://doi.org/10.3390/ijms20225703
Sestili F, Garcia-Molina MD, Gambacorta G, Beleggia R, Botticella E, De Vita P, Savatin DV, Masci S, Lafiandra D. Provitamin A Biofortification of Durum Wheat through a TILLING Approach. International Journal of Molecular Sciences. 2019; 20(22):5703. https://doi.org/10.3390/ijms20225703
Chicago/Turabian StyleSestili, Francesco, Maria Dolores Garcia-Molina, Gianluca Gambacorta, Romina Beleggia, Ermelinda Botticella, Pasquale De Vita, Daniel Valentin Savatin, Stefania Masci, and Domenico Lafiandra. 2019. "Provitamin A Biofortification of Durum Wheat through a TILLING Approach" International Journal of Molecular Sciences 20, no. 22: 5703. https://doi.org/10.3390/ijms20225703
APA StyleSestili, F., Garcia-Molina, M. D., Gambacorta, G., Beleggia, R., Botticella, E., De Vita, P., Savatin, D. V., Masci, S., & Lafiandra, D. (2019). Provitamin A Biofortification of Durum Wheat through a TILLING Approach. International Journal of Molecular Sciences, 20(22), 5703. https://doi.org/10.3390/ijms20225703