Genome-Wide Analysis of the YABBY Transcription Factor Family in Pineapple and Functional Identification of AcYABBY4 Involvement in Salt Stress

 , ,
, , 
Abstract
:1. Introduction
2. Results
2.1. Identification and Characterization of the Pineapple YABBY Transcription Factors
2.2. Phylogenetic Analysis of YABBY Family Genes
2.3. Gene Structure Analysis and Identification of Conserved Motifs
2.4. AcYABBY Protein Homology Modeling and Sequence Alignment
2.5. Expression Profiling and Subcellular Localization of AcYABBYs
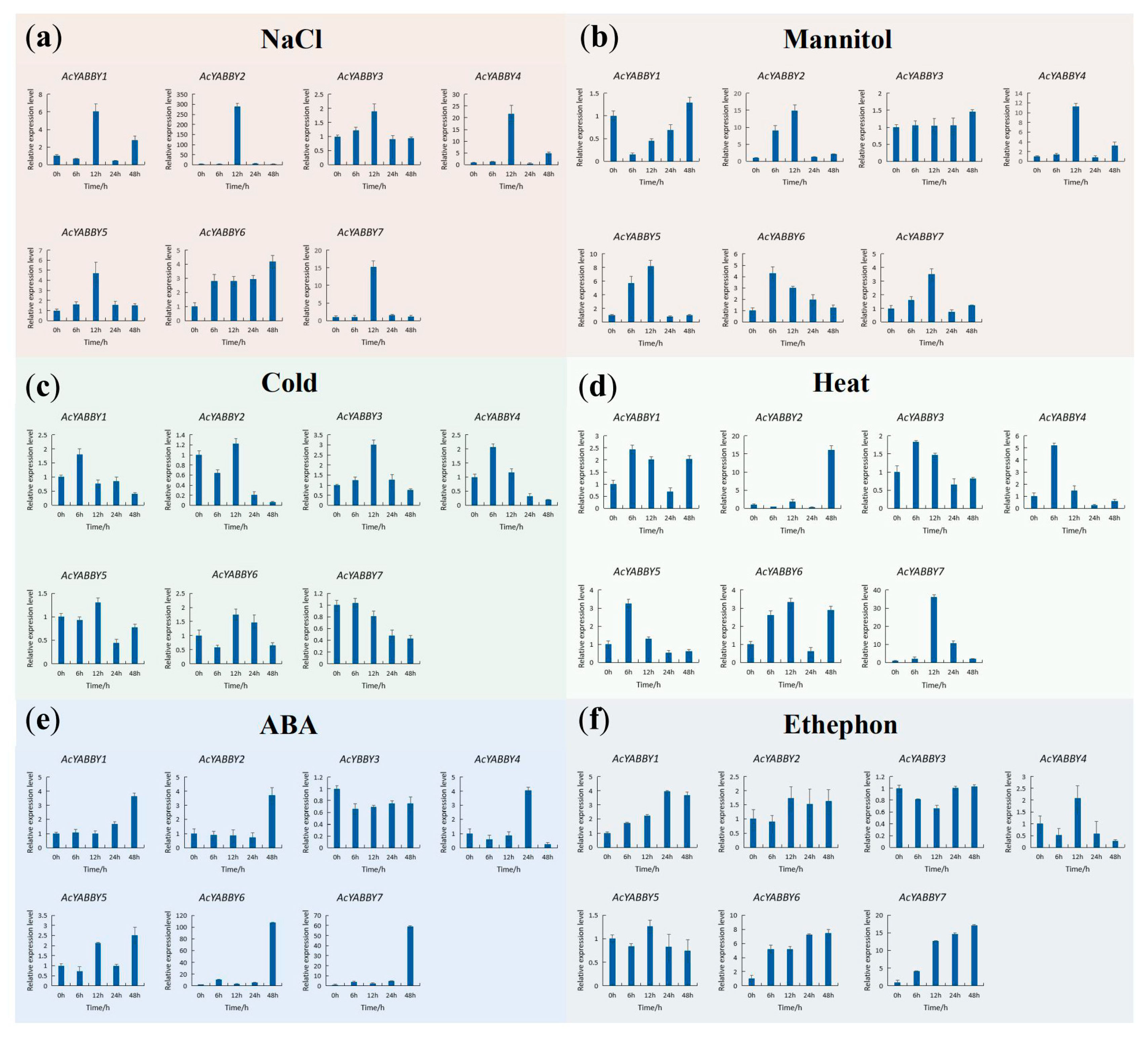
2.6. Cis-Acting Elements and RT-qPCR Analysis of AcYABBY Ggenes
2.7. AcYABBY4 Negatively Regulates the High Salinity Tolerance in Arabidopsis
3. Discussion
3.1. Diversity of YABBY Transcription Factors in Plants
3.2. AcYABBY Gene Expression Profiles Analysis
3.3. AcYABBY4 Inhibits Root Growth of Seedlings under Salt Stress
4. Materials and Methods
4.1. Identification of YABBY Transcription Factors in Pineapple
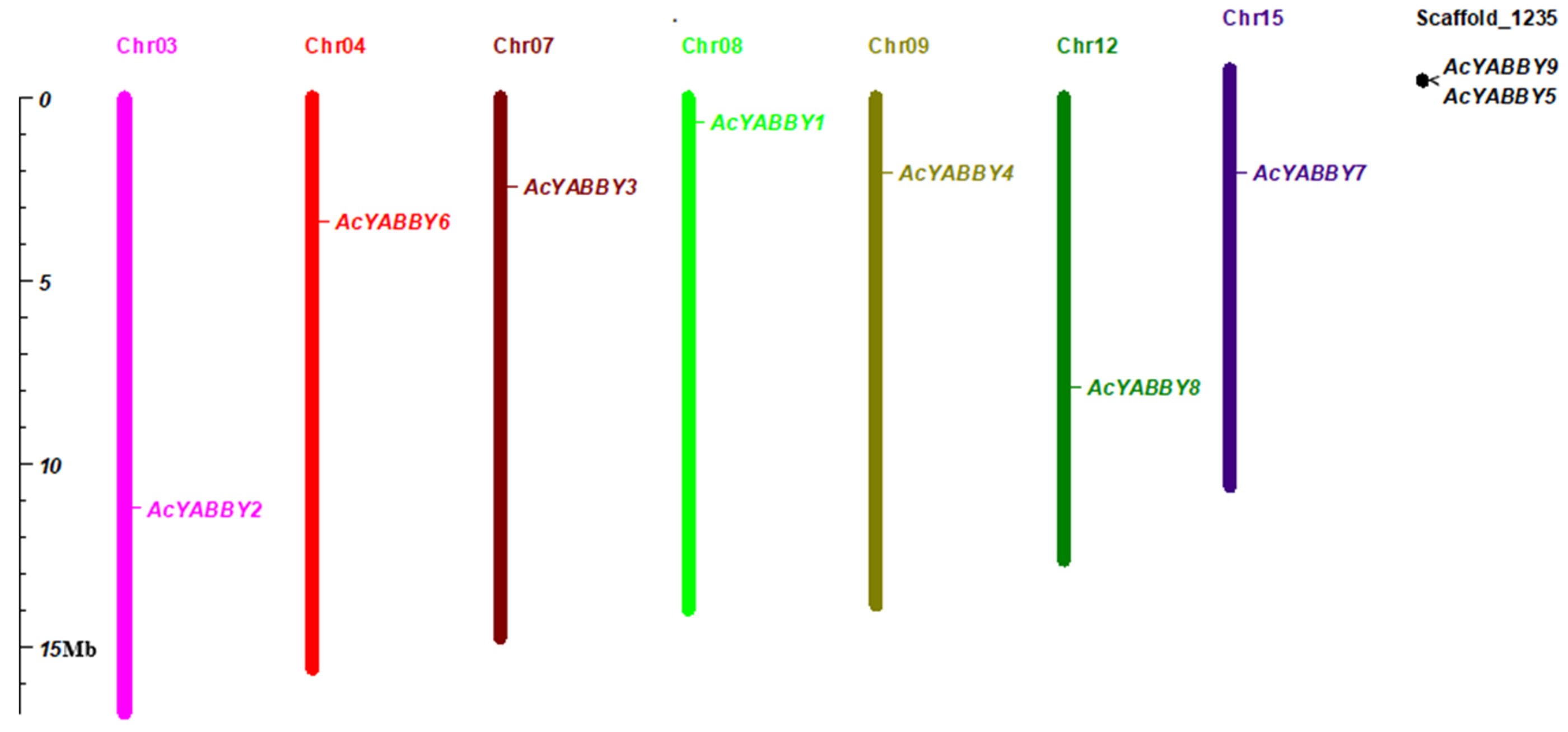
4.2. Chromosome Locations of Pineapple YABBY Genes
4.3. Construction of Phylogenetic Tree
4.4. Gene Structure Analysis and Identification of Conserved Motifs
4.5. Modeling of Protein 3D Structures and Sequence Alignment of AcYABBY Proteins
4.6. RNA-Seq and RT-qPCR Data Analysis
4.7. Vector Construction and Subcellular Localization
4.8. Cis-Acting Elements in the Pineapple YABBY Genes Promoter region
4.9. Stress Treatment
4.10. Tolerance Assays under Stress Conditions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ABA | Abscisic acid; |
| GSDS | Gene Structure Display Server |
| HMM | Hidden Markov Models |
| MEGE | Molecular Evolutionary Genetics Analysis |
| MEME | Multiple Em for Motif Elicitation |
| RT-qPCR | Real-time Quantitative PCR |
| RNA-Seq | RNA sequencing |
| SMART | Simple Modular Architecture Research Tool |
| MS | Murashige and Skoog |
| PEG | Polyethylene glycol |
| WT | Wild type |
| GFP | Green fluorescent protein |
| AtYABBY | Arabidopsis thaliana YABBY |
| OsYABBY | rice (Oryza sativa) YABBY |
| AcYABBY | pineapple (Ananas comosus) YABBY |
| CDS | Coding sequence |
| ATG | Starting codon |
| FPKM | Fragments Per Kilobase Million |
| DAPI | 4′,6-Diamidino-2-Phenylindole |
| MS | Murashige and Skoog |
| HMG | High mobility group |
| MYB | V-myb Myeloblastosis Viral Oncogene Homolog |
| MYC | Myelocytomatosis oncogenes |
| ERE | Ethylene Responsive Element |
References
- Zhao, S.P.; Lu, D.; Yu, T.F.; Ji, Y.J.; Zheng, W.J.; Zhang, S.X.; Chai, S.C.; Chen, Z.Y.; Cui, X.Y. Genome-wide analysis of the YABBY family in soybean and functional identification of GmYABBY10 involvement in high salt and drought stresses. Plant Physiol. Biochem. 2017, 119, 132–146. [Google Scholar] [CrossRef] [PubMed]
- Sawa, S.; Watanabe, K.; Goto, K.; Liu, Y.G.; Shibata, D.; Kanaya, E.; Morita, E.H.; Okada, K. FILAMENTOUS FLOWER, a meristem and organ identity gene of Arabidopsis, encodes a protein with a zinc finger and HMG-related domains. Genes Dev. 1999, 13, 1079–1088. [Google Scholar] [CrossRef] [PubMed]
- Sarojam, R.; Sappl, P.G.; Goldshmidt, A.; Efroni, I.; Floyd, S.K.; Eshed, Y.; Bowman, J.L. Differentiating Arabidopsis shoots from leaves by combined YABBY activities. Plant Cell 2010, 22, 2113–2130. [Google Scholar] [CrossRef]
- Yamada, T.; Ito, M.; Kato, M. Expression pattern of INNER NO OUTER homologue in Nymphaea (water lily family, Nymphaeaceae). Dev. Genes Evol. 2003, 213, 510–513. [Google Scholar] [CrossRef] [PubMed]
- Siegfried, K.R.; Eshed, Y.; Baum, S.F.; Otsuga, D.; Drews, G.N.; Bowman, J.L. Members of the YABBY gene family specify abaxial cell fate in Arabidopsis. Development 1999, 126, 4117–4128. [Google Scholar] [PubMed]
- Balasubramanian, S.; Schneitz, K. NOZZLE links proximal-distal and adaxial-abaxial pattern formation during ovule development in Arabidopsis thaliana. Development 2002, 129, 4291–4300. [Google Scholar] [PubMed]
- Eshed, Y.; Baum, S.F.; Bowman, J.L. Distinct mechanisms promote polarity establishment in carpels of Arabidopsis. Cell 1999, 99, 199–209. [Google Scholar] [CrossRef]
- Kerstetter, R.A.; Bollman, K.; Taylor, R.A.; Bomblies, K.; Poethig, R.S. KANADI regulates organ polarity in Arabidopsis. Nature 2001, 411, 706–709. [Google Scholar] [CrossRef]
- Yamada, T.; Yokota, S.; Hirayama, Y.; Imaichi, R.; Kato, M.; Gasser, C.S. Ancestral expression patterns and evolutionary diversification of YABBY genes in angiosperms. Plant J. 2011, 67, 26–36. [Google Scholar] [CrossRef]
- Eckardt, N.A. YABBY genes and the development and origin of seed plant leaves. Plant Cell 2010, 22, 2103. [Google Scholar] [CrossRef]
- Stahle, M.I.; Kuehlich, J.; Staron, L.; von Arnim, A.G.; Golz, J.F. YABBYs and the transcriptional corepressors LEUNIG and LEUNIG_HOMOLOG maintain leaf polarity and meristem activity in Arabidopsis. Plant Cell 2009, 21, 3105–3118. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.; Hur, J.; Kim, S.J.; Han, M.J.; Kim, S.R.; An, G. Ectopic expression of OsYAB1 causes extra stamens and carpels in rice. Plant Mol. Biol. 2004, 56, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, T.; Nagasawa, N.; Kawasaki, S.; Matsuoka, M.; Nagato, Y.; Hirano, H.Y. The YABBY gene DROOPING LEAF regulates carpel specification and midrib development in Oryza sativa. Plant Cell 2004, 16, 500–509. [Google Scholar] [CrossRef] [PubMed]
- Ha, C.M.; Jun, J.H.; Fletcher, J.C. Control of Arabidopsis leaf morphogenesis through regulation of the YABBY and KNOX families of transcription factors. Genetics 2010, 186, 197–206. [Google Scholar] [CrossRef]
- Zhang, X.L.; Yang, Z.P.; Zhang, J.; Zhang, L.G. Ectopic expression of BraYAB1–702, a member of YABBY gene family in Chinese cabbage, causes leaf curling, inhibition of development of shoot apical meristem and flowering stage delaying in Arabidopsis thaliana. Int. J. Mol. Sci. 2013, 14, 14872–14891. [Google Scholar] [CrossRef]
- Toriba, T.; Harada, K.; Takamura, A.; Nakamura, H.; Ichikawa, H.; Suzaki, T.; Hirano, H.Y. Molecular characterization the YABBY gene family in Oryza sativa and expression analysis of OsYABBY1. Mol. Genet Genom. 2007, 277, 457–468. [Google Scholar] [CrossRef]
- Villanueva, J.M.; Broadhvest, J.; Hauser, B.A.; Meister, R.J.; Schneitz, K.; Gasser, C.S. INNER NO OUTER regulates abaxial- adaxial patterning in Arabidopsis ovules. Genes Dev. 1999, 13, 3160–3169. [Google Scholar] [CrossRef]
- Ohmori, Y.; Toriba, T.; Nakamura, H.; Ichikawa, H.; Hirano, H.Y. Temporal and spatial regulation of DROOPING LEAF gene expression that promotes midrib formation in rice. Plant J. 2011, 65, 77–86. [Google Scholar] [CrossRef]
- Lin, Z.; Li, X.; Shannon, L.M.; Yeh, C.T.; Wang, M.L.; Bai, G.; Peng, Z.; Li, J.; Trick, H.N.; Clemente, T.E.; et al. Parallel domestication of the Shattering1 genes in cereals. Nat. Genet 2012, 44, 720–724. [Google Scholar] [CrossRef]
- Ming, R.; VanBuren, R.; Wai, C.M.; Tang, H.; Schatz, M.C.; Bowers, J.E.; Lyons, E.; Wang, M.L.; Chen, J.; Biggers, E.; et al. The pineapple genome and the evolution of CAM photosynthesis. Nat. Genet 2015, 47, 1435–1442. [Google Scholar] [CrossRef]
- Guo, A.Y.; Zhu, Q.H.; Chen, X.; Luo, J.C. GSDS: A gene structure display server. Yi Chuan 2007, 29, 1023–1026. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Elkan, C. Fitting a mixture model by expectation maximization to discover motifs in biopolymers. Proc. Int. Conf. Intell. Syst. Mol. Biol. 1994, 2, 28–36. [Google Scholar] [PubMed]
- Bowman, J.L. The YABBY gene family and abaxial cell fate. Curr Opin Plant Biol 2000, 3, 17–22. [Google Scholar] [CrossRef]
- Finet, C.; Floyd, S.K.; Conway, S.J.; Zhong, B.; Scutt, C.P.; Bowman, J.L. Evolution of the YABBY gene family in seed plants. Evol. Dev. 2016, 18, 116–126. [Google Scholar] [CrossRef]
- Dai, M.; Hu, Y.; Zhao, Y.; Liu, H.; Zhou, D.X. A WUSCHEL-LIKE HOMEOBOX gene represses a YABBY gene expression required for rice leaf development. Plant Physiol. 2007, 144, 380–390. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, J.; Ming, R. Genomic analyses of the CAM plant pineapple. J. Exp. Bot. 2014, 65, 3395–3404. [Google Scholar] [CrossRef]
- Su, Z.X.; Wang, L.L.; Li, W.M.; Zhao, L.H.; Huang, X.Y.; Azam, S.M.; Qin, Y. Genome-Wide Identification of Auxin Response Factor (ARF) Genes Family and its Tissue-Specific Prominent Expression in Pineapple (Ananas comosus). Trop Plant Biol. 2017, 10, 86–96. [Google Scholar] [CrossRef]
- He, Y.A.; Li, W.; Lv, J.; Jia, Y.B.; Wang, M.C.; Xia, G.M. Ectopic expression of a wheat MYB transcription factor gene, TaMYB73, improves salinity stress tolerance in Arabidopsis thaliana. J. Exp. Bot. 2012, 63, 1511–1522. [Google Scholar] [CrossRef]
- Yang, A.; Dai, X.Y.; Zhang, W.H. A R2R3-type MYB gene, OsMYB2, is involved in salt, cold and dehydration tolerance in rice. J. Exp. Bot. 2012, 63, 2541–2556. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, D.; Yang, C.; Kong, N.; Shi, Z.; Zhao, P.; Nan, Y.; Nie, T.; Wang, R.; Ma, H.; et al. Genome-wide identification of the potato WRKY transcription factor family. PLoS ONE 2017, 12, e0181573. [Google Scholar] [CrossRef]
- Li, W.; Yan, M.; Hu, B.; Priyadarshani, S.V.G.N.; Hou, Z.; Ojolo, S.P.; Xiong, J.; Zhao, H.; Qin, Y. Characterization and the Expression Analysis of Nitrate Transporter (NRT) Gene Family in Pineapple. Trop. Plant Biol. 2018, 11, 177–191. [Google Scholar] [CrossRef]
- Zhang, M.; Liu, Y.; Shi, H.; Guo, M.; Chai, M.; He, Q.; Yan, M.; Cao, D.; Zhao, L.; Cai, H.; et al. Evolutionary and expression analyses of soybean basic Leucine zipper transcription factor family. BMC Genomics 2018, 19, 159. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Hua, X.; Xu, J.; Chen, Z.; Fan, T.; Zeng, Z.; Wang, H.; Hour, A.L.; Yu, Q.; Ming, R.; et al. Comparative genomics revealed the gene evolution and functional divergence of magnesium transporter families in Saccharum. BMC Genomics 2019, 20, 1471–2164. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clough, S.J.; Bent, A.F. Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Priyadarshani, S.; Hu, B.; Li, W.; Ali, H.; Jia, H.; Zhao, L.; Ojolo, S.P.; Azam, S.M.; Xiong, J.; Yan, M.; et al. Simple protoplast isolation system for gene expression and protein interaction studies in pineapple (Ananas comosus L.). Plant Methods 2018, 14, 95. [Google Scholar] [CrossRef] [Green Version]
- Xie, T.; Chen, C.; Li, C.; Liu, J.; Liu, C.; He, Y. Genome-wide investigation of WRKY gene family in pineapple: evolution and expression profiles during development and stress. BMC Genomics 2018, 19, 490. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Gene Locus | Chromosome Location | PI | MW (Da) | Protein Length(aa) | CDS Length(bp) | Exon |
|---|---|---|---|---|---|---|---|
| AcYABBY1 | Aco007606 | 8 | 9.45 | 20,042.95 | 182 | 549 | 5 |
| AcYABBY2 | Aco016279 | 3 | 6.88 | 24,706.13 | 226 | 681 | 7 |
| AcYABBY3 | Aco005138 | 7 | 9.25 | 19,962.60 | 178 | 537 | 6 |
| AcYABBY4 | Aco008751 | 9 | 8.64 | 20,161.95 | 178 | 537 | 6 |
| AcYABBY5 | Aco028479 | scaffold1235 | 9.05 | 15,073.06 | 136 | 411 | 5 |
| AcYABBY6 | Aco002202 | 4 | 7.71 | 20,872.43 | 188 | 567 | 6 |
| AcYABBY7 | Aco003917 | 15 | 9.05 | 21,453.75 | 190 | 573 | 7 |
| AcYABBY8 | Aco026269 | 12 | 7.01 | 5383.98 | 49 | 150 | 2 |
| AcYABBY9 | Aco028478 | scaffold1235 | 5.82 | 5988.82 | 53 | 162 | 2 |
| Gene | Motif Sequence | AcYABBY1 | AcYABBY2 | AcYABBY3 | AcYABBY4 | AcYABBY5 | AcYABBY6 | AcYABBY7 |
|---|---|---|---|---|---|---|---|---|
| MYB | TAACCA | 6 | 5 | 6 | 9 | 2 | 4 | 2 |
| CAACAG | ||||||||
| CAACCA | ||||||||
| MYC | CATTTG | 4 | 3 | 4 | 7 | 8 | 5 | 5 |
| CATGTG | ||||||||
| CAATTG | ||||||||
| ERE | ATTTCATA | 3 | 2 | 0 | 0 | 1 | 3 | 0 |
| ATTTTAAA |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Li, G.; Cai, M.; Priyadarshani, S.V.G.N.; Aslam, M.; Zhou, Q.; Huang, X.; Wang, X.; Liu, Y.; Qin, Y. Genome-Wide Analysis of the YABBY Transcription Factor Family in Pineapple and Functional Identification of AcYABBY4 Involvement in Salt Stress. Int. J. Mol. Sci. 2019, 20, 5863. https://doi.org/10.3390/ijms20235863
Li Z, Li G, Cai M, Priyadarshani SVGN, Aslam M, Zhou Q, Huang X, Wang X, Liu Y, Qin Y. Genome-Wide Analysis of the YABBY Transcription Factor Family in Pineapple and Functional Identification of AcYABBY4 Involvement in Salt Stress. International Journal of Molecular Sciences. 2019; 20(23):5863. https://doi.org/10.3390/ijms20235863
Chicago/Turabian StyleLi, Zeyun, Gang Li, Mingxing Cai, Samaranayaka V.G.N. Priyadarshani, Mohammad Aslam, Qiao Zhou, Xiaoyi Huang, Xiaomei Wang, Yeqiang Liu, and Yuan Qin. 2019. "Genome-Wide Analysis of the YABBY Transcription Factor Family in Pineapple and Functional Identification of AcYABBY4 Involvement in Salt Stress" International Journal of Molecular Sciences 20, no. 23: 5863. https://doi.org/10.3390/ijms20235863
APA StyleLi, Z., Li, G., Cai, M., Priyadarshani, S. V. G. N., Aslam, M., Zhou, Q., Huang, X., Wang, X., Liu, Y., & Qin, Y. (2019). Genome-Wide Analysis of the YABBY Transcription Factor Family in Pineapple and Functional Identification of AcYABBY4 Involvement in Salt Stress. International Journal of Molecular Sciences, 20(23), 5863. https://doi.org/10.3390/ijms20235863