Discovery of a Highly Selective MC1R Agonists Pentapeptide to Be Used as a Skin Pigmentation Enhancer and with Potential Anti-Aging Properties



Abstract
:1. Introduction
2. Results
2.1. MC1R-Agonist Peptides Activate MC1R
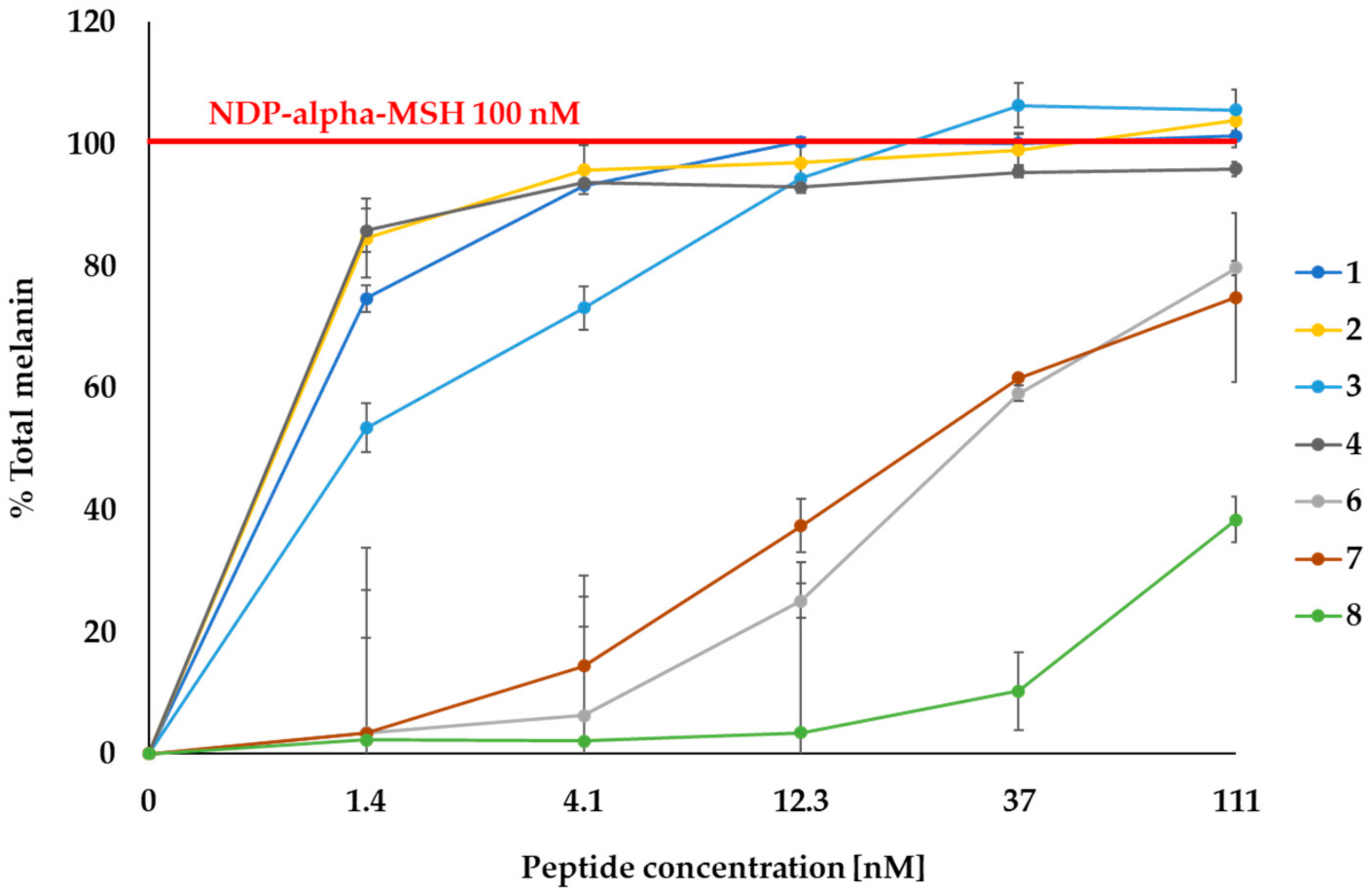
2.2. MC1R-Agonist Peptides Stimulated Melanin Synthesis in Mouse Melanoma Cells
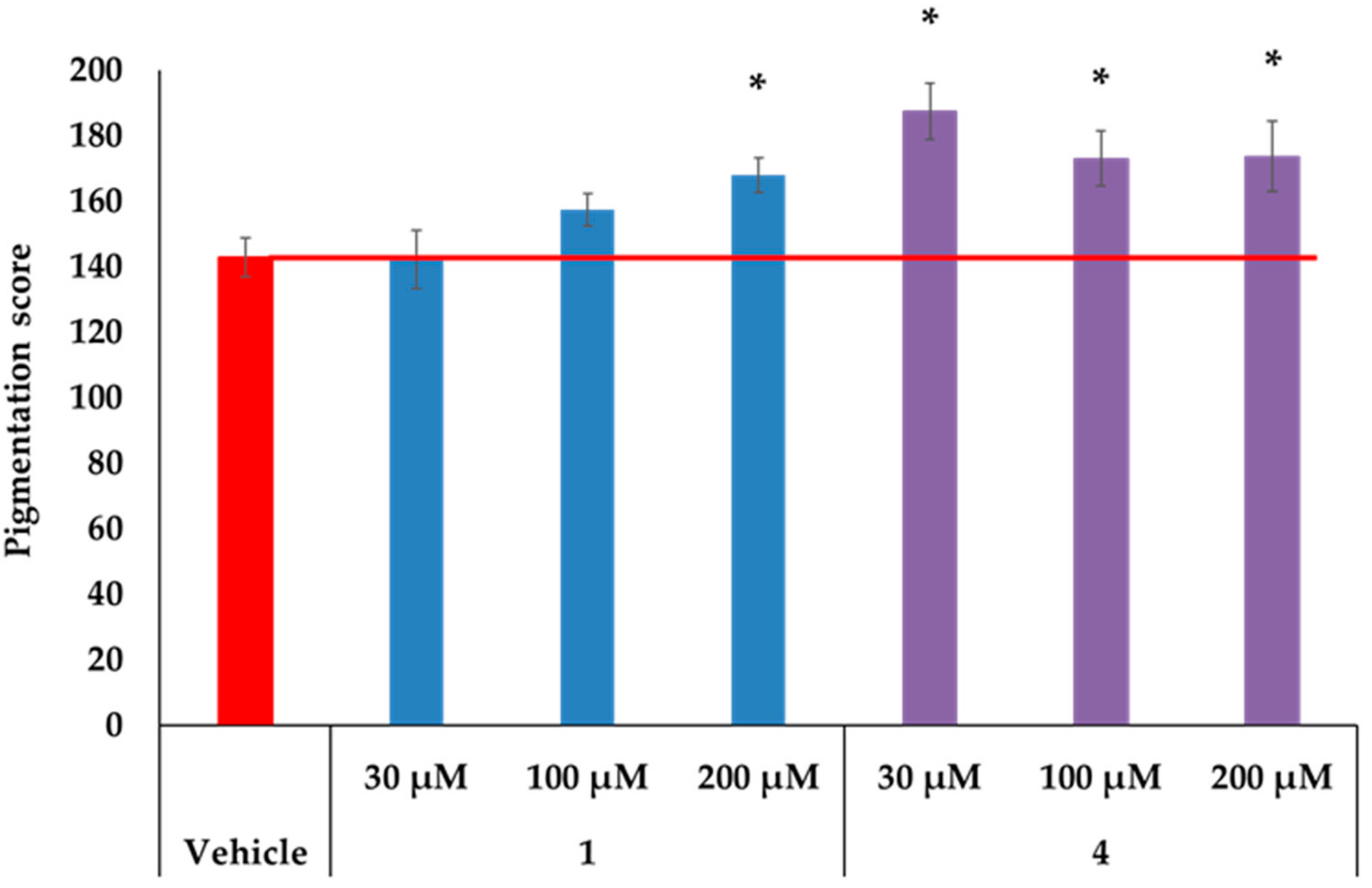
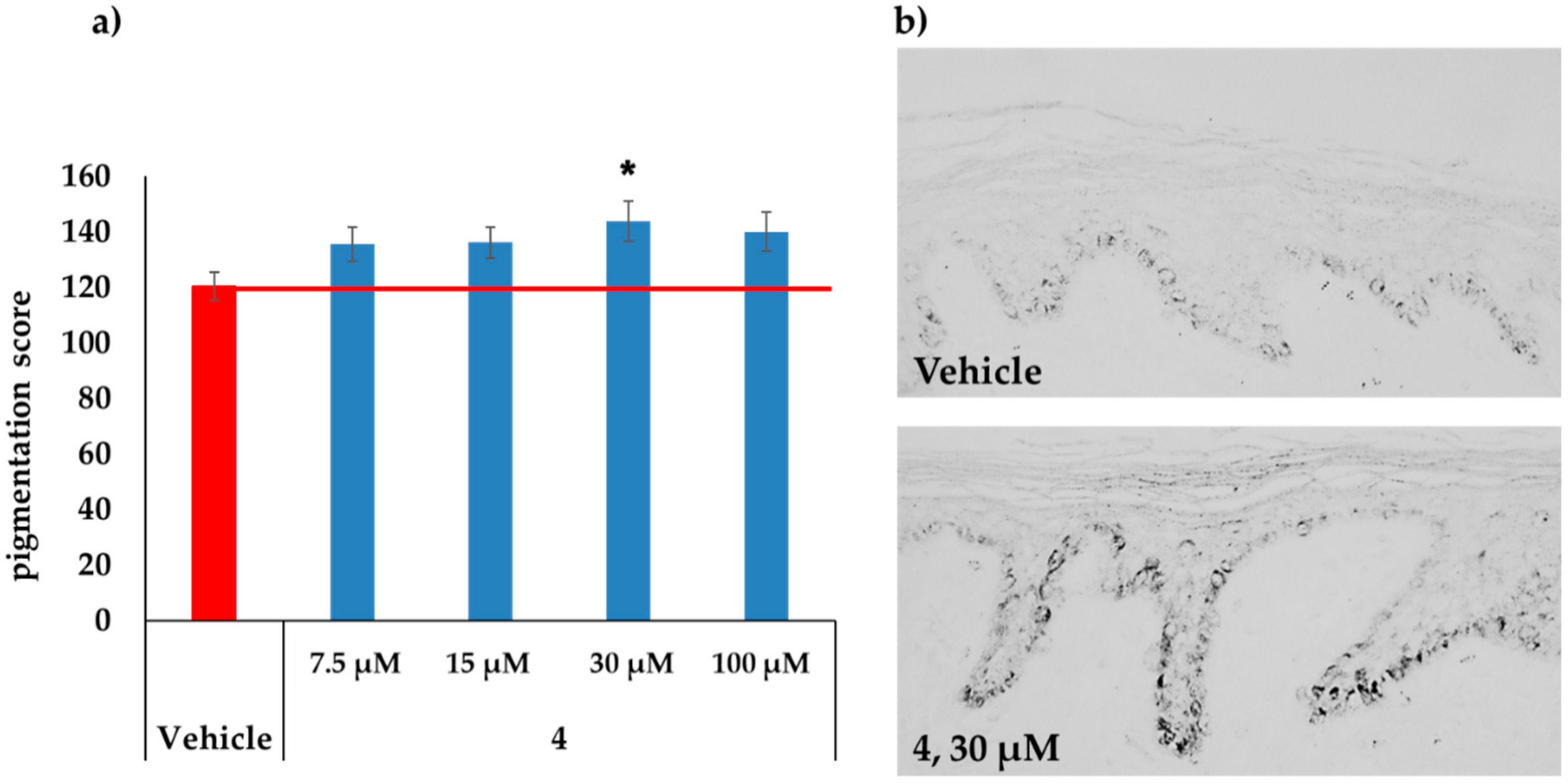
2.3. MC1R-Agonist Peptides Stimulated Pigmentation in Human Skin Ex Vivo
2.4. MC1R-Agonist Peptides Activated MC1R Selectively
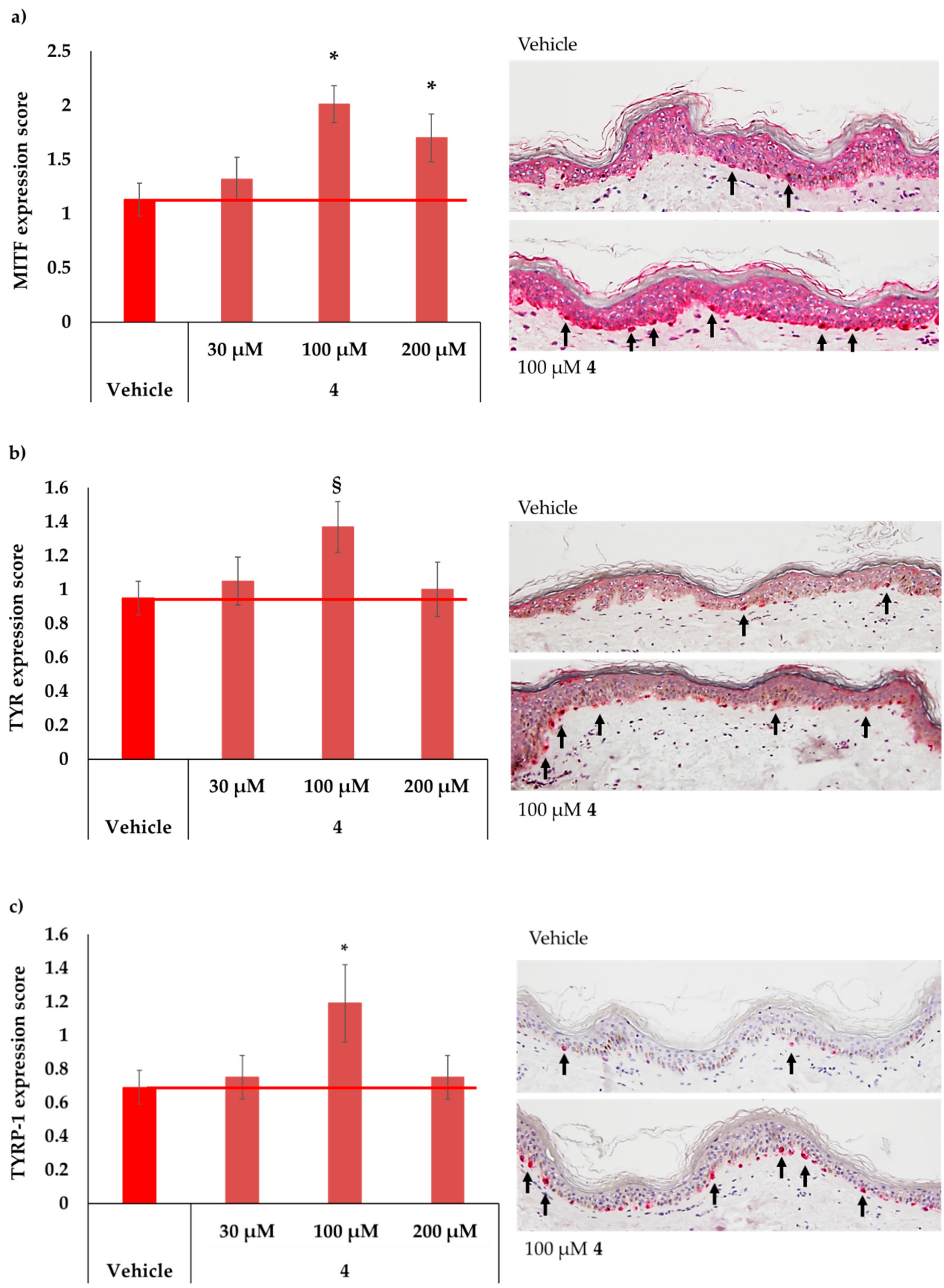
2.5. MC1R-Agonist Peptide 4 Induced Key Melanogenesis Markers in Human Skin Ex Vivo
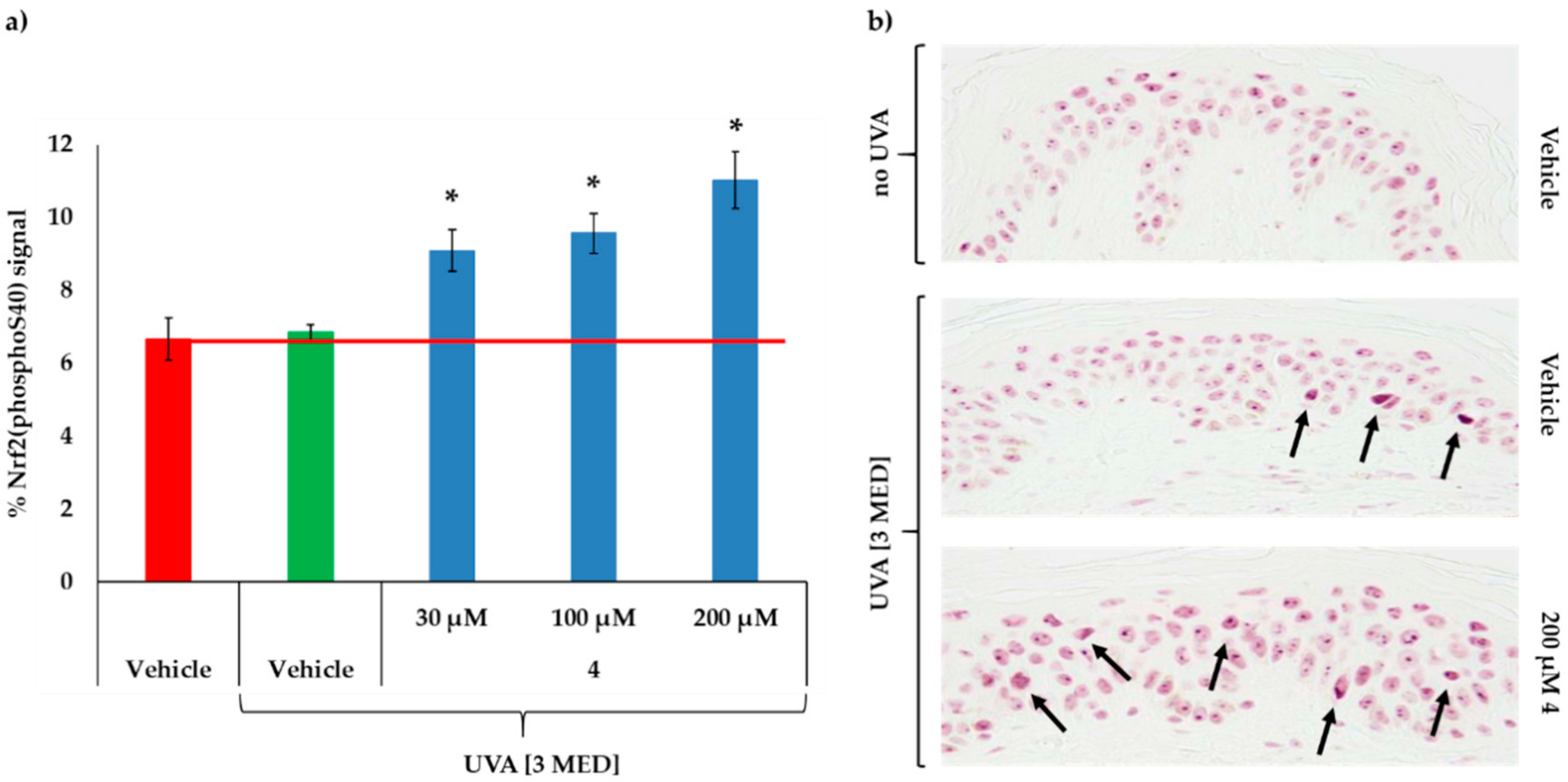
2.6. MC1R-Agonist Peptide 4 Induced Nrf2 Serine 40 Phosphorylation Ex Vivo
3. Discussion
4. Material and Methods
4.1. Synthesis
4.2. MCR-Activation Assays
4.3. MT- and OP-Receptor Activation Assays
4.4. MCH-, ETB-, and 5-HT2A-Receptor Activation Assays
4.5. Cell Culture for Melanogenesis Stimulation
4.6. Tissue Culture to Assess Induction of Melanogenesis
4.7. Immunohistochemistry of Melanogenesis Markers
4.8. Nrf2 Activation in Skin Tissue after UVA-Irradiation
5. Patents
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Abdel-Malek, Z.; Suzuki, I.; Tada, A.; Im, S.; Akcali, C. The Melanocortin-1 Receptor and Human Pigmentation. Ann. N. Y. Acad. Sci. 1999, 885, 117–133. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Malek, Z.; Scott, M.C.; Suzuki, I.; Tada, A.; Im, S.; Lamoreux, L.; Ito, S.; Barsh, G.; Hearing, V.J. The Melanocortin-1 Receptor is a Key Regulator of Human Cutaneous Pigmentation. Pigment Cell Res. 2000, 13, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Mountjoy, K.G.; Robbins, L.S.; Mortrud, M.T.; Cone, R.D. The cloning of a family of genes that encode the melanocortin receptors. Science 1992, 257, 1248–1251. [Google Scholar] [CrossRef] [PubMed]
- Chhajlani, V.; Wikberg, J.E. Molecular cloning and expression of the human melanocyte stimulating hormone receptor cDNA. FEBS Lett. 1992, 309, 417–420. [Google Scholar] [CrossRef] [Green Version]
- Chhajlani, V.; Muceniece, R.; Wikberg, J.E.S. Molecular cloning of a novel human melanocortin receptor. Biochem. Biophys. Res. Com. 1993, 195, 866–873. [Google Scholar] [CrossRef] [PubMed]
- Gantz, I.; Konda, Y.; Tashiro, T.; Shimoto, Y.; Miwa, H.; Munzert, G.; Watson, S.J.; DelValle, J.; Yamada, T. Molecular Cloning of a Novel Melanocortin Receptor. J. Biol. Chem. 1993, 268, 8246–8250. [Google Scholar] [PubMed]
- Gantz, I.; Miwa, H.; Konda, Y.; Shimoto, Y.; Tashiro, T.; Watson, S.J.; DelValle, J.; Yamada, T. Molecular Cloning, Expression, and Gene Localization of a Fourth Melanocortin Receptor. J. Biol. Chem. 1993, 268, 15174–15179. [Google Scholar]
- Gantz, I.; Shimoto, Y.; Konda, Y.; Miwa, H.; Dickinson, C.J.; Yamada, T. Molecular cloning, expression, and characterization of a fifth melanocortin receptor. Biochem. Biophys. Res. Com. 1994, 200, 1214–1220. [Google Scholar] [CrossRef] [Green Version]
- Cai, M.; Hruby, V.J. The Melanocortin Receptor System: A Target for Multiple Degenerative Diseases. Curr. Protein Pept. Sci. 2016, 17, 488–496. [Google Scholar] [CrossRef]
- Bohm, M.; Luger, T.A.; Tobin, D.J.; Garcia-Borron, J.C. Melanocortin receptor ligands: New horizons for skin biology and clinical dermatology. J. Investig. Dermatol. 2006, 126, 1966–1975. [Google Scholar] [CrossRef] [Green Version]
- Kadekaro, A.L.; Kavanagh, R.; Kanto, H.; Terzieva, S.; Hauser, J.; Kobayashi, N.; Schwemberger, S.; Cornelius, J.; Babcock, G.; Shertzer, H.G.; et al. A-Melanocortin and Endothelin-1 Activate Antiapoptotic Pathways and Reduce DNA Damage in Human Melanocytes. Cancer Res. 2005, 65, 4292–4299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, X.; Mosby, N.; Yang, J.; Xu, A.; Abdel-Malek, Z.; Kadekaro, A.L. alpha-MSH activates immediate defense responses to UV-induced oxidative stress in human melanocytes. Pigment Cell Melanoma Res. 2009, 22, 809–818. [Google Scholar] [CrossRef] [PubMed]
- Wintzen, M.; Gilchrest, B.A. Proopiomelanocortin, its derived peptides, and the skin. J. Investig. Dermatol. 1996, 106, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Borron, J.C.; Abdel-Malek, Z.; Jimenez-Cervantes, C. MC1R, the cAMP pathway, and the response to solar UV: Extending the horizon beyond pigmentation. Pigment Cell Melanoma Res. 2014, 27, 699–720. [Google Scholar] [CrossRef]
- Ou-Yang, H.; Stamatas, G.; Kollias, N. Spectral Responses of Melanin to Ultraviolet A Irradiation. J. Investig. Dermatol. 2004, 122, 492–496. [Google Scholar] [CrossRef] [Green Version]
- Brenner, M.; Hearing, V.J. The Protective Role of Melanin Against UV Damage in Human Skin. Photochem. Photobiol. 2008, 84, 539–549. [Google Scholar] [CrossRef] [Green Version]
- D’Orazio, J.; Jarrett, S.; Amaro-Ortiz, A.; Scott, T. UV radiation and the skin. Int. J. Mol. Sci. 2013, 14, 12222–12248. [Google Scholar] [CrossRef] [Green Version]
- Schiller, M.; Brzoska, T.; Bohm, M.; Metze, D.; Scholzen, T.E.; Rougier, A.; Luger, T.A. Solar-simulated ultraviolet radiation-induced upregulation of the melanocortin-1 receptor, proopiomelanocortin, and alpha-melanocyte-stimulating hormone in human epidermis in vivo. J. Investig. Dermatol. 2004, 122, 468–476. [Google Scholar]
- Bolognia, J.; Murray, M.; Pawelek, J. UVB-induced melanogenesis may be mediated through the MSH-receptor system. J. Investig. Dermatol. 1989, 92, 651–656. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, A.K.; Orlow, S.J.; Bolognia, J.L.; Pawelek, J.M. Structural/functional relationships between internal and external MSH receptors: Modulation of expression in Cloudman melanoma cells by UVB radiation. J. Cell. Physiol. 1991, 147, 1–6. [Google Scholar] [CrossRef]
- Tobin, D.J.; Swanson, N.N.; Pittelkow, M.R.; Peters, E.M.; Schallreuter, K.U. Melanocytes are not absent in lesional skin of long duration vitiligo. J. Pathol. 2000, 191, 407–416. [Google Scholar] [CrossRef]
- Barysch, M.J.; Braun, R.P.; Kolm, I.; Ahlgrimm-Siesz, V.; Hofmann-Wellenhof, R.; Duval, C.; Warrick, E.; Bernerd, F.; Nouveau, S.; Dummer, R. Keratinocytic Malfunction as a Trigger for the Development of Solar Lentigines. Dermatopathology (Basel) 2019, 6, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Choi, W.; Yin, L.; Smuda, C.; Batzer, J.; Hearing, V.J.; Kolbe, L. Molecular and histological characterization of age spots. Exp. Dermatol. 2017, 26, 242–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fink, B.; Grammer, K.; Matts, P. Visible skin color distribution plays a role in the perception of age, attractiveness, and health in female faces. Evol. Hum. Behav. 2006, 27, 433–442. [Google Scholar] [CrossRef]
- Porta, E.A. Pigments in Aging: An Overview. Ann. N. Y. Acad. Sci. 2002, 959, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.R.; Parrish, J.A. The Optics of Human Skin. J. Investig. Dermatol. 1981, 77, 13–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kokot, A.; Metze, D.; Mouchet, N.; Galibert, M.D.; Schiller, M.; Luger, T.A.; Bohm, M. Alpha-melanocyte-stimulating hormone counteracts the suppressive effect of UVB on Nrf2 and Nrf-dependent gene expression in human skin. Endocrinology 2009, 150, 3197–3206. [Google Scholar] [CrossRef] [Green Version]
- Ikehata, H.; Yamamoto, M. Roles of the KEAP1-NRF2 system in mammalian skin exposed to UV radiation. Toxicol. Appl. Pharmacol. 2018, 360, 69–77. [Google Scholar] [CrossRef]
- Huang, H.C.; Nguyen, T.; Pickett, C.B. Phosphorylation of Nrf2 at Ser-40 by protein kinase C regulates antioxidant response element-mediated transcription. J. Biol. Chem. 2002, 277, 42769–42774. [Google Scholar] [CrossRef] [Green Version]
- Alam, J.; Stewart, D.; Touchard, C.; Boinapally, S.; Choi, A.M.; Cook, J.L. Nrf2, a Cap‘n’Collar transcription factor, regulates induction of the heme oxygenase-1 gene. J. Biol. Chem. 1999, 274, 26071–26078. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Malek, Z.A.; Ruwe, A.; Kavanagh-Starner, R.; Kadekaro, A.L.; Swope, V.; Haskell-Luevano, C.; Koikov, L.; Knittel, J.J. alpha-MSH tripeptide analogs activate the melanocortin 1 receptor and reduce UV-induced DNA damage in human melanocytes. Pigment Cell Melanoma Res. 2009, 22, 635–644. [Google Scholar] [CrossRef] [PubMed]
- Scharffetter, K.; Wlaschek, M.; Hogg, A.; Bolsen, K.; Schothorst, A.; Goerz, G.; Krieg, T.; Plewig, G. UVA irradiation induces collagenase in human dermal fibroblasts in vitro and in vivo. Arch. Dermatol. Res. 1991, 283, 506–511. [Google Scholar] [CrossRef]
- Herrmann, G.; Wlaschek, M.; Lange, T.S.; Prenzel, K.; Goerz, G.; Scharffetter-Kochanek, K. UVA irradiation stimulates the synthesis of various matrix-metalloproteinases (MMPs) in cultured human fibroblasts. Exp. Dermatol. 1993, 2, 92–97. [Google Scholar] [CrossRef] [PubMed]
- Naylor, E.C.; Watson, R.E.; Sherratt, M.J. Molecular aspects of skin ageing. Maturitas 2011, 69, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Langton, A.K.; Sherratt, M.J.; Griffiths, C.E.; Watson, R.E. A new wrinkle on old skin: The role of elastic fibres in skin ageing. Int. J. Cosmet. Sci. 2010, 32, 330–339. [Google Scholar] [CrossRef] [PubMed]
- Imokawa, G.; Ishida, K. Biological mechanisms underlying the ultraviolet radiation-induced formation of skin wrinkling and sagging I: Reduced skin elasticity, highly associated with enhanced dermal elastase activity, triggers wrinkling and sagging. Int. J. Mol. Sci. 2015, 16, 7753–7775. [Google Scholar] [CrossRef] [Green Version]
- Imokawa, G.; Nakajima, H.; Ishida, K. Biological mechanisms underlying the ultraviolet radiation-induced formation of skin wrinkling and sagging II: Over-expression of neprilysin plays an essential role. Int. J. Mol. Sci. 2015, 16, 7776–7795. [Google Scholar] [CrossRef] [Green Version]
- Rawlings, A.V. Ethnic skin types: Are there differences in skin structure and function? Int. J. Cosmet. Sci. 2006, 28, 81–95. [Google Scholar] [CrossRef]
- Hillebrand, G.G.; Levine, M.J.; Miyamoto, K. The Age-Dependent Changes in Skin Condition in African Americans, Asian Indians, Caucasians, East Asians, and Latinos. IFSCC Magazine 2001, 4, 259–266. [Google Scholar]
- Campiche, R.; Trevisan, S.; Seroul, P.; Rawlings, A.V.; Adnet, C.; Imfeld, D.; Voegeli, R. Appearance of aging signs in differently pigmented facial skin by a novel imaging system. J. Cosmet. Dermatol. 2019, 18, 614–627. [Google Scholar] [CrossRef]
- Slominski, A. Neuroendocrine activity of the melanocyte. Exp. Dermatol. 2009, 18, 760–763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slominski, A.; Paus, R.; Schadendorf, D. Melanocytes as “sensory” and regulatory cells in the epidermis. J. Theor. Biol. 1993, 164, 103–120. [Google Scholar] [CrossRef] [PubMed]
- Bocheva, G.; Slominski, R.M.; Slominski, A.T. Neuroendocrine Aspects of Skin Aging. Int. J. Mol. Sci. 2019, 20, 2798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slominski, A.T.; Zmijewski, M.A.; Skobowiat, C.; Zbytek, B.; Slominski, R.M.; Steketee, J.D. Sensing the environment: Regulation of local and global homeostasis by the skin’s neuroendocrine system. Adv. Anat. Embryol. Cell Biol. 2012, 212, v, vii, 1–115. [Google Scholar] [PubMed]
- Pain, S.; Dezutter, C.; Reymermier, C.; Vogelgesang, B.; Delay, E.; Andre, V. Age-related changes in pro-opiomelanocortin (POMC) and related receptors in human epidermis. Int. J. Cosmet. Sci. 2010, 32, 266–275. [Google Scholar] [CrossRef]
- Liu, F.; Hamer, M.A.; Deelen, J.; Lall, J.S.; Jacobs, L.; van Heemst, D.; Murray, P.G.; Wollstein, A.; de Craen, A.J.; Uh, H.W.; et al. The MC1R Gene and Youthful Looks. Curr. Biol. 2016, 26, 1213–1220. [Google Scholar] [CrossRef] [Green Version]
- Law, M.H.; Medland, S.E.; Zhu, G.; Yazar, S.; Vinuela, A.; Wallace, L.; Shekar, S.N.; Duffy, D.L.; Bataille, V.; Glass, D.; et al. Genome-Wide Association Shows that Pigmentation Genes Play a Role in Skin Aging. J. Investig. Dermatol. 2017, 137, 1887–1894. [Google Scholar] [CrossRef] [Green Version]
- Sawyer, T.K.; Sanfilippo, P.J.; Hruby, V.J.; Engel, M.H.; Heward, C.B.; Burnett, J.B.; Hadley, M.E. 4-Norleucine, 7-D-phenylalanine-alpha-melanocyte-stimulating hormone: A highly potent alpha-melanotropin with ultralong biological activity. Proc. Natl. Acad. Sci. USA 1980, 77, 5754–5758. [Google Scholar] [CrossRef] [Green Version]
- Langendonk, J.G.; Balwani, M.; Anderson, K.E.; Bonkovsky, H.L.; Anstey, A.V.; Bissell, D.M.; Bloomer, J.; Edwards, C.; Neumann, N.J.; Parker, C.; et al. Afamelanotide for Erythropoietic Protoporphyria. N. Engl. J. Med. 2015, 373, 48–59. [Google Scholar] [CrossRef]
- Mowlazadeh Haghighi, S.; Zhou, Y.; Dai, J.; Sawyer, J.R.; Hruby, V.J.; Cai, M. Replacement of Arg with Nle and modified D-Phe in the core sequence of MSHs, Ac-His-D-Phe-Arg-Trp-NH2, leads to hMC1R selectivity and pigmentation. Eur. J. Med. Chem. 2018, 151, 815–823. [Google Scholar] [CrossRef]
- Wilson, K.R.; Todorovic, A.; Proneth, B.; Haskell-Luevano, C. Overview of endogenous and synthetic melanocortin peptides. Cell Mol. Biol. (Noisy-le-grand) 2006, 52, 3–20. [Google Scholar]
- Mujahid, N.; Liang, Y.; Murakami, R.; Choi, H.G.; Dobry, A.S.; Wang, J.; Suita, Y.; Weng, Q.Y.; Allouche, J.; Kemeny, L.V.; et al. A UV-Independent Topical Small-Molecule Approach for Melanin Production in Human Skin. Cell Reports 2017, 19, 2177–2184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holder, J.R.; Marques, F.F.; Xiang, Z.; Bauzo, R.M.; Haskell-Luevano, C. Characterization of aliphatic, cyclic, and aromatic N-terminally “capped” His-d-Phe-Arg-Trp-NH2 tetrapeptides at the melanocortin receptors. Eur. J. Pharmacol. 2003, 462. [Google Scholar] [CrossRef]
- Holder, J.R.; Xiang, Z.; Bauzo, R.M.; Haskell-Luevano, C. Structure–activity relationships of the melanocortin tetrapeptide Ac-His-DPhe-Arg-Trp-NH2 at the mouse melanocortin receptors. Part 3: modifications at the Arg position. Peptides 2003, 24, 73–82. [Google Scholar] [CrossRef]
- Fischer, T.W.; Kleszczynski, K.; Hardkop, L.H.; Kruse, N.; Zillikens, D. Melatonin enhances antioxidative enzyme gene expression (CAT, GPx, SOD), prevents their UVR-induced depletion, and protects against the formation of DNA damage (8-hydroxy-2’-deoxyguanosine) in ex vivo human skin. J. Pineal Res. 2013, 54, 303–312. [Google Scholar] [CrossRef]
- Kauser, S.; Schallreuter, K.U.; Thody, A.J.; Gummer, C.; Tobin, D.J. Regulation of human epidermal melanocyte biology by beta-endorphin. J. Investig. Dermatol. 2003, 120, 1073–1080. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.J.; Park, M.K.; Kim, S.Y.; Park Choo, H.Y.; Lee, A.Y.; Lee, C.H. Serotonin induces melanogenesis via serotonin receptor 2A. Br. J. Dermatol. 2011, 165, 1344–1348. [Google Scholar] [CrossRef]
- Hoogduijn, M.J.; Ancans, J.; Suzuki, I.; Eastdale, S.; Thody, A.J. Melanin-concentrating hormone and its receptor are expressed and functional in human skin. Biochem. Biophys. Res. Com. 2002, 296, 698–701. [Google Scholar] [CrossRef]
- Kemp, E.H.; Weetman, A.P. Melanin-concentrating hormone and melanin-concentrating hormone receptors in mammalian skin physiopathology. Peptides 2009, 30, 2071–2075. [Google Scholar] [CrossRef]
- Swope, V.B.; Abdel-Malek, Z.A. Significance of the Melanocortin 1 and Endothelin B Receptors in Melanocyte Homeostasis and Prevention of Sun-Induced Genotoxicity. Front. Genet. 2016, 7, 146. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Malek, Z.A.; Kadekaro, A.L.; Kavanagh, R.J.; Todorovic, A.; Koikov, L.N.; McNulty, J.C.; Jackson, P.J.; Millhauser, G.L.; Schwemberger, S.; Babcock, G.; et al. Melanoma prevention strategy based on using tetrapeptide alpha-MSH analogs that protect human melanocytes from UV-induced DNA damage and cytotoxicity. FASEB J. 2006, 20, 1561–1563. [Google Scholar] [CrossRef]
- Chung, S.; Lim, G.J.; Lee, J.Y. Quantitative analysis of melanin content in a three-dimensional melanoma cell culture. Sci. Rep. 2019, 9, 780. [Google Scholar] [CrossRef]
- Bigliardi-Qi, M.; Bigliardi, P.L.; Eberle, A.N.; Buchner, S.; Rufli, T. beta-endorphin stimulates cytokeratin 16 expression and downregulates mu-opiate receptor expression in human epidermis. J. Investig. Dermatol. 2000, 114, 527–532. [Google Scholar] [CrossRef] [Green Version]
- Bohm, M.; Luger, T. Are melanocortin peptides future therapeutics for cutaneous wound healing? Exp. Dermatol. 2019, 28, 219–224. [Google Scholar] [CrossRef] [Green Version]
- Nasti, T.H.; Timares, L. MC1R, eumelanin and pheomelanin: Their role in determining the susceptibility to skin cancer. Photochem. Photobiol. 2015, 91, 188–200. [Google Scholar] [CrossRef] [Green Version]
- Jarrett, S.G.; Wolf Horrell, E.M.; Boulanger, M.C.; D’Orazio, J.A. Defining the Contribution of MC1R Physiological Ligands to ATR Phosphorylation at Ser435, a Predictor of DNA Repair in Melanocytes. J. Investig. Dermatol. 2015, 135, 3086–3095. [Google Scholar] [CrossRef] [Green Version]
- Duffy, D.L.; Lee, K.J.; Jagirdar, K.; Pflugfelder, A.; Stark, M.S.; McMeniman, E.K.; Soyer, H.P.; Sturm, R.A. High naevus count and MC1R red hair alleles contribute synergistically to increased melanoma risk. Br. J. Dermatol. 2019, 181, 1009–1016. [Google Scholar] [CrossRef] [Green Version]
- Del Bino, S.; Sok, J.; Bessac, E.; Bernerd, F. Relationship between skin response to ultraviolet exposure and skin color type. Pigment Cell Res. 2006, 19, 606–614. [Google Scholar] [CrossRef]
- Chardon, A.; Cretois, I.; Hourseau, C. Skin colour typology and suntanning pathways. Int. J. Cosmet. Sci. 1991, 13, 191–208. [Google Scholar] [CrossRef]
- Fitzpatrick, T.B. The Validity and Practicality of Sun-Reactive Skin Types I Through VI. Arch. Dermatol. 1988, 124, 869–871. [Google Scholar] [CrossRef]
- Degorce, F.; Card, A.; Soh, S.; Trinquet, E.; Knapik, G.P.; Xie, B. HTRF: A Technology Tailored for Drug Discovery –A Review of Theoretical Aspects and Recent Applications. Current Chemical. Genomics 2009, 3, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Le Poul, E.; Hisada, S.; Mizuguchi, Y.; Dupriez, V.J.; Burgeon, E.; Detheux, M. Adaptation of aequorin functional assay to high throughput screening. J. Biomol. Screen 2002, 7, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
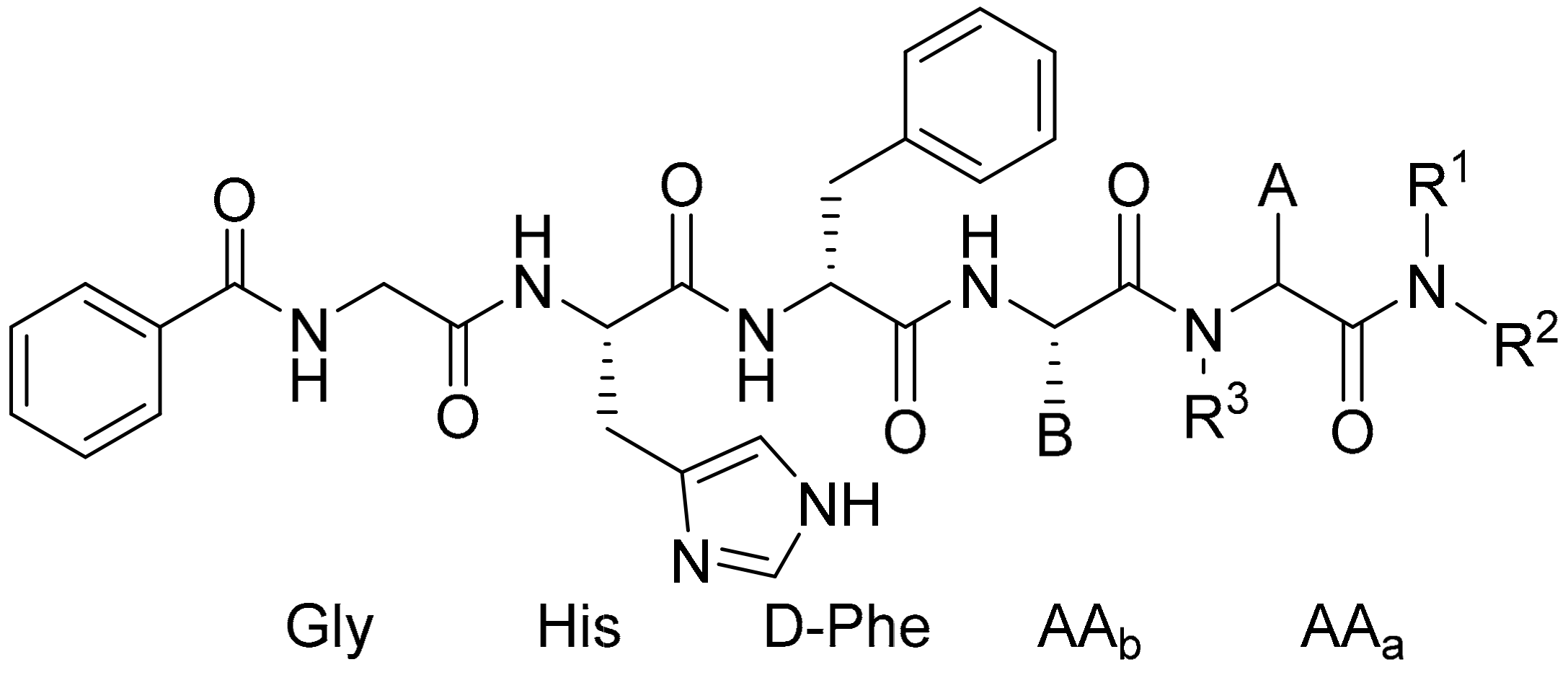
| Entry | AAb | AAa | R1 | R2 | R3 | EC50 (nM) |
|---|---|---|---|---|---|---|
| 1 | Arg | L-Trp | H | H | H | 0.041 |
| 2 | Arg | L-NaphAla | H | H | H | 0.11 |
| 3 | Arg | D-NaphAla | H | H | H | 0.25 |
| 4 | Arg | D-Trp | Propyl | Propyl | H | 0.42 |
| 5 | Arg | L-Trp | Propyl | Propyl | H | 0.88 |
| 6 | Arg | D-Phe | H | H | H | 1.79 |
| 7 | Arg | Gly | H | H | CH2-3-Indolyl | 4.4 |
| 8 | Dab | L-Trp | H | H | H | 7.51 |
| Peptide Entry | Receptor | EC50 [nM] (Activation) |
|---|---|---|
| 4 | MC1R | 0.42 |
| 4 | MC2R | > 100,000 |
| 4 | MC3R | 877 |
| 4 | MC4R | 136 |
| 4 | MC5R | > 100,000 |
| 4 | MT1R | > 100,000 |
| 4 | MT2R | > 100,000 |
| 4 | OP1R | > 100,000 |
| 4 | OP2R | > 100,000 |
| 4 | OP3R | > 100,000 |
| 4 | MCH1R | > 100,000 |
| 4 | MCH2R | > 100,000 |
| 4 | ETBR | > 100,000 |
| 4 | 5-HT2AR | > 100,000 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jackson, E.; Heidl, M.; Imfeld, D.; Meeus, L.; Schuetz, R.; Campiche, R. Discovery of a Highly Selective MC1R Agonists Pentapeptide to Be Used as a Skin Pigmentation Enhancer and with Potential Anti-Aging Properties. Int. J. Mol. Sci. 2019, 20, 6143. https://doi.org/10.3390/ijms20246143
Jackson E, Heidl M, Imfeld D, Meeus L, Schuetz R, Campiche R. Discovery of a Highly Selective MC1R Agonists Pentapeptide to Be Used as a Skin Pigmentation Enhancer and with Potential Anti-Aging Properties. International Journal of Molecular Sciences. 2019; 20(24):6143. https://doi.org/10.3390/ijms20246143
Chicago/Turabian StyleJackson, Eileen, Marc Heidl, Dominik Imfeld, Laurent Meeus, Rolf Schuetz, and Remo Campiche. 2019. "Discovery of a Highly Selective MC1R Agonists Pentapeptide to Be Used as a Skin Pigmentation Enhancer and with Potential Anti-Aging Properties" International Journal of Molecular Sciences 20, no. 24: 6143. https://doi.org/10.3390/ijms20246143
APA StyleJackson, E., Heidl, M., Imfeld, D., Meeus, L., Schuetz, R., & Campiche, R. (2019). Discovery of a Highly Selective MC1R Agonists Pentapeptide to Be Used as a Skin Pigmentation Enhancer and with Potential Anti-Aging Properties. International Journal of Molecular Sciences, 20(24), 6143. https://doi.org/10.3390/ijms20246143