Post-Genomic Methodologies and Preclinical Animal Models: Chances for the Translation of Cardioprotection to the Clinic





Abstract
:1. Introduction
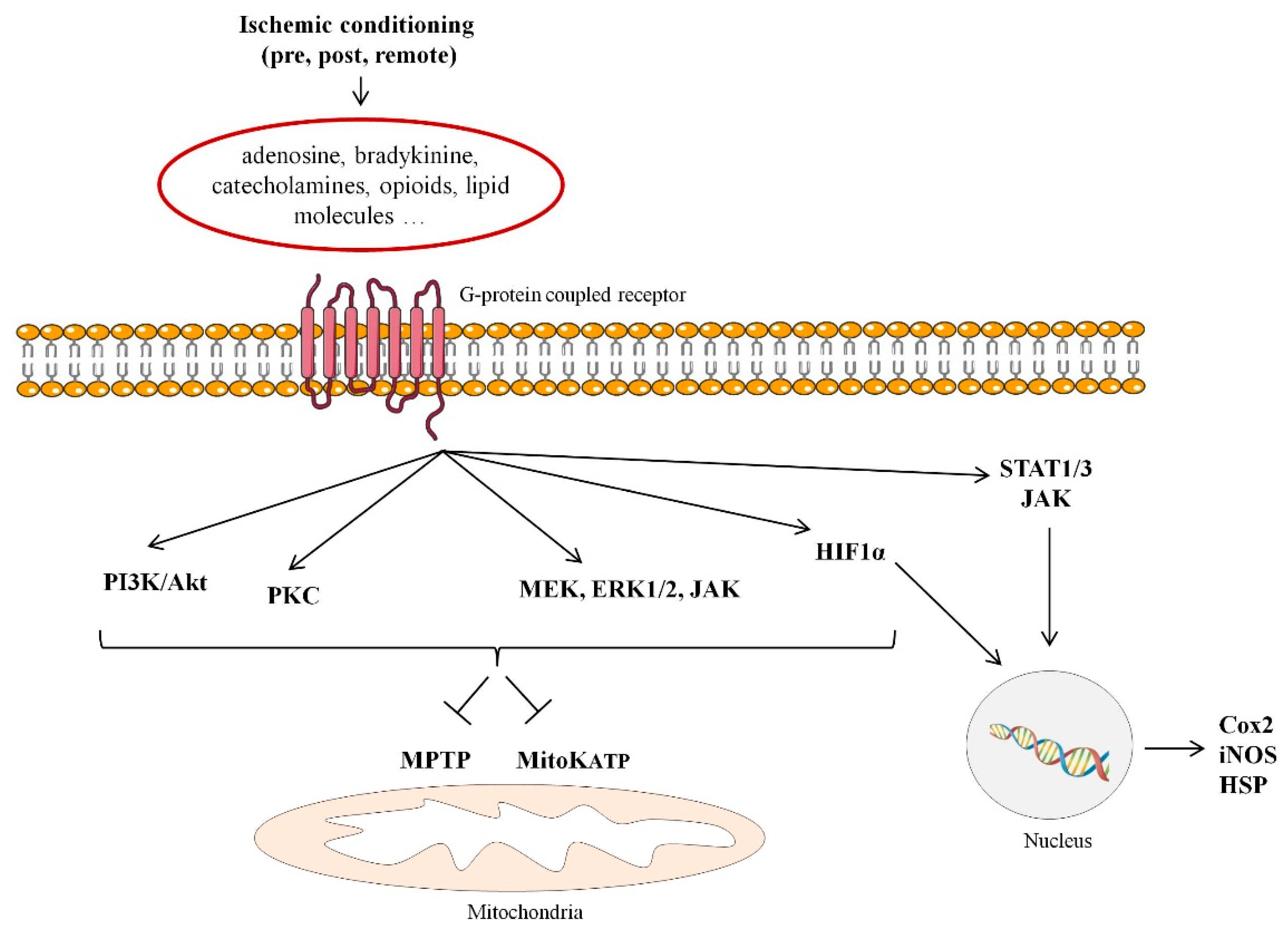
2. Cardioprotection
3. Animal Models of Cardioprotection: Why Pre-Clinical Animal Models?
3.1. Rodents vs. Swine
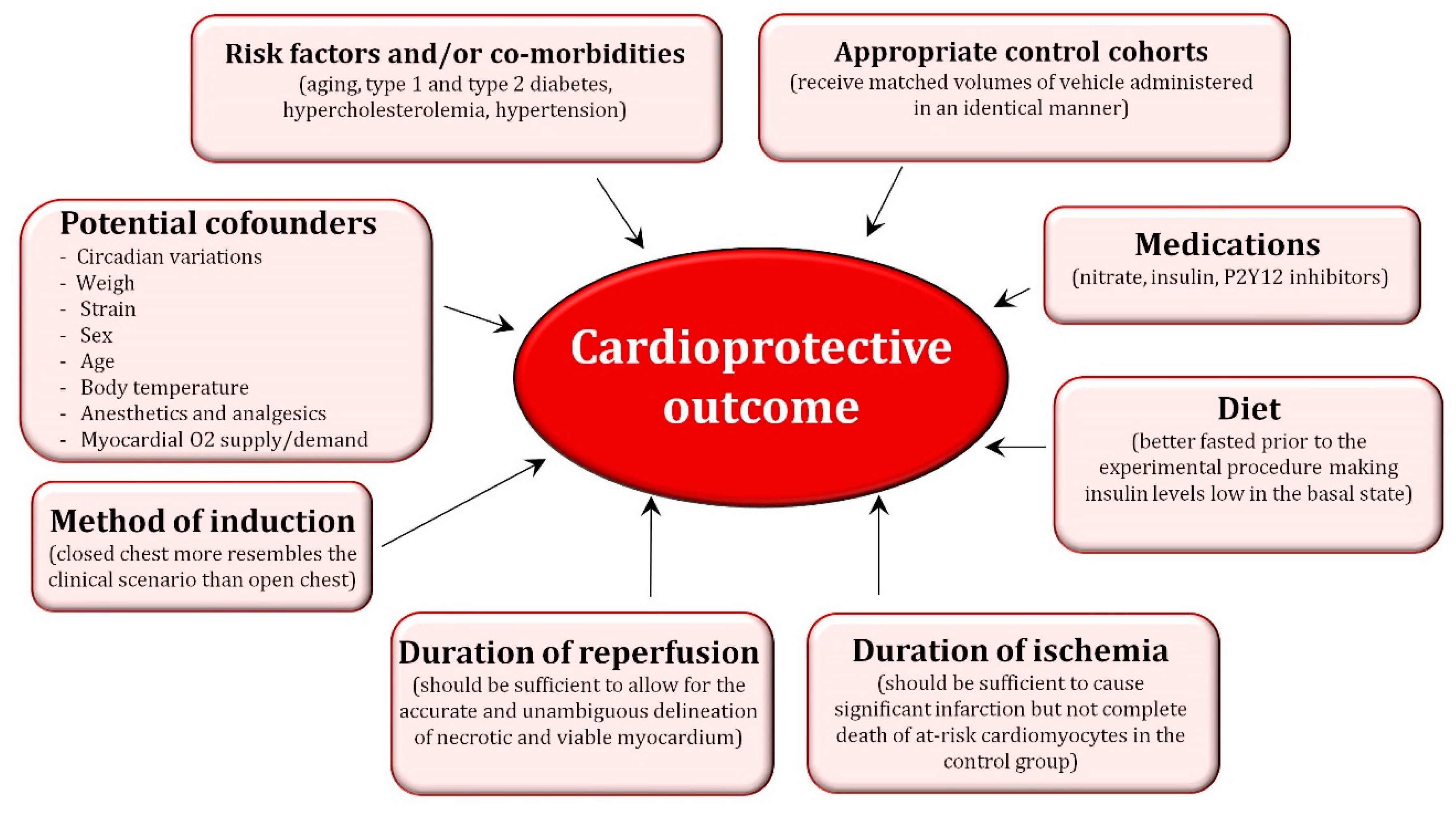
3.2. Important Factors to Consider When Performing Cardioprotective Studies in Pre-Clinical Animal Models
4. Implementation of Post-Genomic Technologies in Preclinical Animal Models to Identify New Targets/Strategies for Cardioprotection
4.1. Transcriptomics
4.2. Proteomics and Post-Translational Modifications
4.2.1. Proteomics
4.2.2. Post-Translational Modifications
Phosphoproteomics
O-GlcNAcylation
4.3. Metabolomics and Lipidomics
4.4. Epigenetic Modifications: Methylation and Acetylation
5. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Human Genome Project Information. Available online: https://www.genome.gov/10001772/all-about-the--human-genome-project-hgp/ (accessed on 24 January 2019).
- Venter, J.C.; Adams, M.D.; Myers, E.W.; Li, P.W.; Mural, R.J.; Sutton, G.G.; Smith, H.O.; Yandell, M.; Evans, C.A.; Holt, R.A.; et al. The sequence of the human genome. Science 2001, 291, 1304–1351. [Google Scholar] [CrossRef] [PubMed]
- Roger, V.L.; Go, A.S.; Lloyd-Jones, D.M.; Benjamin, E.J.; Berry, J.D.; Borden, W.B.; Bravata, D.M.; Dai, S.; Ford, E.S.; Fox, C.S.; et al. Heart disease and stroke statistics—2012 update: A report from the American Heart Association. Circulation 2012, 125, e2–e220. [Google Scholar] [PubMed]
- Bernink, F.J.; Timmers, L.; Beek, A.M.; Diamant, M.; Roos, S.T.; Van Rossum, A.C.; Appelman, Y. Progression in attenuating myocardial reperfusion injury: An overview. Int. J. Cardiol. 2014, 170, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Vilahur, G.; Juan-Babot, O.; Pena, E.; Onate, B.; Casani, L.; Badimon, L. Molecular and cellular mechanisms involved in cardiac remodeling after acute myocardial infarction. J. Mol. Cell. Cardiol. 2011, 50, 522–533. [Google Scholar] [CrossRef] [PubMed]
- Ibanez, B.; Heusch, G.; Ovize, M.; Van de Werf, F. Evolving therapies for myocardial ischemia/reperfusion injury. J. Am. Coll. Cardiol. 2015, 65, 1454–1471. [Google Scholar] [CrossRef] [PubMed]
- Heusch, G. Cardioprotection: Chances and challenges of its translation to the clinic. Lancet 2013, 381, 166–175. [Google Scholar] [CrossRef]
- Vilahur, G.; Casani, L.; Pena, E.; Juan-Babot, O.; Mendieta, G.; Crespo, J.; Badimon, L. HMG-CoA reductase inhibition prior reperfusion improves reparative fibrosis post-myocardial infarction in a preclinical experimental model. Int. J. Cardiol. 2014, 175, 528–538. [Google Scholar] [CrossRef]
- Vilahur, G.; Casani, L.; Pena, E.; Duran, X.; Juan-Babot, O.; Badimon, L. Induction of RISK by HMG-CoA reductase inhibition affords cardioprotection after myocardial infarction. Atherosclerosis 2009, 206, 95–101. [Google Scholar] [CrossRef]
- Ibanez, B.; Prat-Gonzalez, S.; Speidl, W.S.; Vilahur, G.; Pinero, A.; Cimmino, G.; Garcia, M.J.; Fuster, V.; Sanz, J.; Badimon, J.J. Early metoprolol administration before coronary reperfusion results in increased myocardial salvage: Analysis of ischemic myocardium at risk using cardiac magnetic resonance. Circulation 2007, 115, 2909–2916. [Google Scholar] [CrossRef]
- Bulluck, H.; Yellon, D.M.; Hausenloy, D.J. Reducing myocardial infarct size: Challenges and future opportunities. Heart 2016, 102, 341–348. [Google Scholar] [CrossRef]
- Hausenloy, D.J.; Barrabes, J.A.; Botker, H.E.; Davidson, S.M.; Di Lisa, F.; Downey, J.; Engstrom, T.; Ferdinandy, P.; Carbrera-Fuentes, H.A.; Heusch, G.; et al. Ischaemic conditioning and targeting reperfusion injury: A 30 year voyage of discovery. Basic Res. Cardiol. 2016, 111, 70. [Google Scholar] [CrossRef] [PubMed]
- Murry, C.E.; Jennings, R.B.; Reimer, K.A. Preconditioning with ischemia: A delay of lethal cell injury in ischemic myocardium. Circulation 1986, 74, 1124–1136. [Google Scholar] [CrossRef] [PubMed]
- Heusch, G. Molecular basis of cardioprotection: Signal transduction in ischemic pre-, post-, and remote conditioning. Circ. Res. 2015, 116, 674–699. [Google Scholar] [CrossRef] [PubMed]
- Hausenloy, D.J. Cardioprotection techniques: Preconditioning, postconditioning and remote conditioning (basic science). Curr. Pharm. Des. 2013, 19, 4544–4563. [Google Scholar] [CrossRef] [PubMed]
- Hausenloy, D.J.; Yellon, D.M. Preconditioning and postconditioning: Underlying mechanisms and clinical application. Atherosclerosis 2009, 204, 334–341. [Google Scholar] [CrossRef]
- Yang, Q.; He, G.W.; Underwood, M.J.; Yu, C.M. Cellular and molecular mechanisms of endothelial ischemia/reperfusion injury: Perspectives and implications for postischemic myocardial protection. Am. J. Transl. Res. 2016, 8, 765–777. [Google Scholar]
- Ibanez, B.; Heusch, G.; García-Dorado, D.; Fuster, V. Molecular and Cellular Mechanisms of Myocardial Ischemia/Reperfusion Injury, 14th ed.; McGraw-Hill: New York, NY, USA, 2017. [Google Scholar]
- Meerson, F.Z.; Gomzakov, O.A.; Shimkovich, M.V. Adaptation to high altitude hypoxia as a factor preventing development of myocardial ischemic necrosis. Am. J. Cardiol. 1973, 31, 30–34. [Google Scholar] [CrossRef]
- Mallet, R.T.; Manukhina, E.B.; Ruelas, S.S.; Caffrey, J.L.; Downey, H.F. Cardioprotection by intermittent hypoxia conditioning: Evidence, mechanisms, and therapeutic potential. Am. J. Physiol.-Heart Circ. Physiol. 2018, 315, H216–H232. [Google Scholar] [CrossRef]
- Wojcik, B.; Knapp, M.; Gorski, J. Non-ischemic heart preconditioning. J. Physiol. Pharmacol. 2018, 69. [Google Scholar] [CrossRef]
- Lindsey, M.L.; Kassiri, Z.; Virag, J.A.I.; de Castro Bras, L.E.; Scherrer-Crosbie, M. Guidelines for measuring cardiac physiology in mice. Am. J. Physiol.-Heart Circ. Physiol. 2018, 314, H733–H752. [Google Scholar] [CrossRef]
- Lindsey, M.L.; Bolli, R.; Canty, J.M., Jr.; Du, X.J.; Frangogiannis, N.G.; Frantz, S.; Gourdie, R.G.; Holmes, J.W.; Jones, S.P.; Kloner, R.A.; et al. Guidelines for experimental models of myocardial ischemia and infarction. Am. J. Physiol.-Heart Circ. Physiol. 2018, 314, H812–H838. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Chen, X.; Zhao, X.; Zhao, B.; Qian, K.; Shi, Y.; Baruscotti, M.; Wang, Y. Screening and Identification of Cardioprotective Compounds from Wenxin Keli by Activity Index Approach and in vivo Zebrafish Model. Front. Pharmacol. 2018, 9, 1288. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.M.; Liu, Y.; Liu, Y.; Tandon, N.; Kambayashi, J.; Downey, J.M.; Cohen, M.V. Attenuation of infarction in cynomolgus monkeys: Preconditioning and postconditioning. Basic Res. Cardiol. 2010, 105, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Hughes, G.C.; Post, M.J.; Simons, M.; Annex, B.H. Translational physiology: Porcine models of human coronary artery disease: Implications for preclinical trials of therapeutic angiogenesis. J. Appl. Physiol. 2003, 94, 1689–1701. [Google Scholar] [CrossRef] [PubMed]
- Bertho, E.; Gagnon, G. A Comparative Study in Three Dimension of the Blood Supply of the Normal Interventricular Septum in Human, Canine, Bovine, Procine, Ovine and Equine Heart. Dis. Chest 1964, 46, 251–262. [Google Scholar] [CrossRef] [PubMed]
- Tsang, H.G.; Rashdan, N.A.; Whitelaw, C.B.; Corcoran, B.M.; Summers, K.M.; MacRae, V.E. Large animal models of cardiovascular disease. Cell Biochem. Funct. 2016, 34, 113–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lecour, S.; Botker, H.E.; Condorelli, G.; Davidson, S.M.; Garcia-Dorado, D.; Engel, F.B.; Ferdinandy, P.; Heusch, G.; Madonna, R.; Ovize, M.; et al. ESC working group cellular biology of the heart: Position paper: Improving the preclinical assessment of novel cardioprotective therapies. Cardiovasc. Res. 2014, 104, 399–411. [Google Scholar] [CrossRef]
- Lelovas, P.P.; Kostomitsopoulos, N.G.; Xanthos, T.T. A comparative anatomic and physiologic overview of the porcine heart. J. Am. Assoc. Lab. Anim. Sci. 2014, 53, 432–438. [Google Scholar]
- Varga, Z.V.; Giricz, Z.; Bencsik, P.; Madonna, R.; Gyongyosi, M.; Schulz, R.; Mayr, M.; Thum, T.; Puskas, L.G.; Ferdinandy, P. Functional Genomics of Cardioprotection by Ischemic Conditioning and the Influence of Comorbid Conditions: Implications in Target Identification. Curr. Drug Targets 2015, 16, 904–911. [Google Scholar] [CrossRef]
- Ferdinandy, P.; Hausenloy, D.J.; Heusch, G.; Baxter, G.F.; Schulz, R. Interaction of risk factors, comorbidities, and comedications with ischemia/reperfusion injury and cardioprotection by preconditioning, postconditioning, and remote conditioning. Pharmacol. Rev. 2014, 66, 1142–1174. [Google Scholar] [CrossRef]
- Ostadal, B.; Netuka, I.; Maly, J.; Besik, J.; Ostadalova, I. Gender differences in cardiac ischemic injury and protection—Experimental aspects. Exp. Biol. Med. 2009, 234, 1011–1019. [Google Scholar] [CrossRef] [PubMed]
- Hunter, J.C.; Korzick, D.H. Age- and sex-dependent alterations in protein kinase C (PKC) and extracellular regulated kinase 1/2 (ERK1/2) in rat myocardium. Mech. Ageing Dev. 2005, 126, 535–550. [Google Scholar] [CrossRef] [PubMed]
- Marongiu, E.; Crisafulli, A. Cardioprotection acquired through exercise: The role of ischemic preconditioning. Curr. Cardiol. Rev. 2014, 10, 336–348. [Google Scholar] [CrossRef] [PubMed]
- Ferdinandy, P. Myocardial ischaemia/reperfusion injury and preconditioning: Effects of hypercholesterolaemia/hyperlipidaemia. Br. J. Pharmacol. 2003, 138, 283–285. [Google Scholar] [CrossRef]
- Ferdinandy, P.; Schulz, R.; Baxter, G.F. Interaction of cardiovascular risk factors with myocardial ischemia/reperfusion injury, preconditioning, and postconditioning. Pharmacol. Rev. 2007, 59, 418–458. [Google Scholar] [CrossRef]
- Tuggle, C.K.; Wang, Y.; Couture, O. Advances in swine transcriptomics. Int. J. Biol. Sci. 2007, 3, 132–152. [Google Scholar] [CrossRef]
- Schroyen, M.; Tuggle, C.K. Current transcriptomics in pig immunity research. Mamm. Genome 2015, 26, 1–20. [Google Scholar] [CrossRef]
- Pena, R.N.; Quintanilla, R.; Manunza, A.; Gallardo, D.; Casellas, J.; Amills, M. Application of the microarray technology to the transcriptional analysis of muscle phenotypes in pigs. Anim. Genet. 2014, 45, 311–321. [Google Scholar] [CrossRef] [Green Version]
- Hausenloy, D.J.; Garcia-Dorado, D.; Botker, H.E.; Davidson, S.M.; Downey, J.; Engel, F.B.; Jennings, R.; Lecour, S.; Leor, J.; Madonna, R.; et al. Novel targets and future strategies for acute cardioprotection: Position Paper of the European Society of Cardiology Working Group on Cellular Biology of the Heart. Cardiovasc. Res. 2017, 113, 564–585. [Google Scholar] [CrossRef] [Green Version]
- Ong, S.B.; Katwadi, K.; Kwek, X.Y.; Ismail, N.I.; Chinda, K.; Ong, S.G.; Hausenloy, D.J. Non-coding RNAs as therapeutic targets for preventing myocardial ischemia-reperfusion injury. Expert Opin. Ther. Targets 2018, 22, 247–261. [Google Scholar] [CrossRef]
- Cheng, Y.; Zhu, P.; Yang, J.; Liu, X.; Dong, S.; Wang, X.; Chun, B.; Zhuang, J.; Zhang, C. Ischaemic preconditioning-regulated miR-21 protects heart against ischaemia/reperfusion injury via anti-apoptosis through its target PDCD4. Cardiovasc. Res. 2010, 87, 431–439. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhu, H.; Zhang, X.; Liu, Y.; Chen, J.; Medvedovic, M.; Li, H.; Weiss, M.J.; Ren, X.; Fan, G.C. Loss of the miR-144/451 cluster impairs ischaemic preconditioning-mediated cardioprotection by targeting Rac-1. Cardiovasc. Res. 2012, 94, 379–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tu, Y.; Wan, L.; Fan, Y.; Wang, K.; Bu, L.; Huang, T.; Cheng, Z.; Shen, B. Ischemic postconditioning-mediated miRNA-21 protects against cardiac ischemia/reperfusion injury via PTEN/Akt pathway. PLoS ONE 2013, 8, e75872. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Yao, K.; Wang, Q.; Guo, J.; Shi, H.; Ma, L.; Liu, H.; Gao, W.; Zou, Y.; Ge, J. Ischemic Postconditioning-Regulated miR-499 Protects the Rat Heart Against Ischemia/Reperfusion Injury by Inhibiting Apoptosis through PDCD4. Cell. Physiol. Biochem. 2016, 39, 2364–2380. [Google Scholar] [CrossRef] [PubMed]
- Baars, T.; Skyschally, A.; Klein-Hitpass, L.; Cario, E.; Erbel, R.; Heusch, G.; Kleinbongard, P. microRNA expression and its potential role in cardioprotection by ischemic postconditioning in pigs. Pflug. Arch. Eur. J. Physiol. 2014, 466, 1953–1961. [Google Scholar] [CrossRef] [PubMed]
- Hinkel, R.; Penzkofer, D.; Zuhlke, S.; Fischer, A.; Husada, W.; Xu, Q.F.; Baloch, E.; van Rooij, E.; Zeiher, A.M.; Kupatt, C.; et al. Inhibition of microRNA-92a protects against ischemia/reperfusion injury in a large-animal model. Circulation 2013, 128, 1066–1075. [Google Scholar] [CrossRef]
- Bellera, N.; Barba, I.; Rodriguez-Sinovas, A.; Ferret, E.; Asin, M.A.; Gonzalez-Alujas, M.T.; Perez-Rodon, J.; Esteves, M.; Fonseca, C.; Toran, N.; et al. Single intracoronary injection of encapsulated antagomir-92a promotes angiogenesis and prevents adverse infarct remodeling. J. Am. Heart Assoc. 2014, 3, e000946. [Google Scholar] [CrossRef]
- Holdt, L.M.; Kohlmaier, A.; Teupser, D. Molecular functions and specific roles of circRNAs in the cardiovascular system. Non-Coding RNA Res. 2018, 3, 75–98. [Google Scholar] [CrossRef]
- Wang, K.; Gan, T.Y.; Li, N.; Liu, C.Y.; Zhou, L.Y.; Gao, J.N.; Chen, C.; Yan, K.W.; Ponnusamy, M.; Zhang, Y.H.; et al. Circular RNA mediates cardiomyocyte death via miRNA-dependent upregulation of MTP18 expression. Cell Death Differ. 2017, 24, 1111–1120. [Google Scholar] [CrossRef]
- Elmadhun, N.Y.; Sadek, A.A.; Sabe, A.A.; Lassaletta, A.D.; Sellke, F.W. Alcohol and the Heart: A Proteomics Analysis of Pericardium and Myocardium in a Swine Model of Myocardial Ischemia. Ann. Thorac. Surg. 2015, 100, 1627–1635. [Google Scholar] [CrossRef]
- Hartl, F.U.; Bracher, A.; Hayer-Hartl, M. Molecular chaperones in protein folding and proteostasis. Nature 2011, 475, 324–332. [Google Scholar] [CrossRef] [PubMed]
- Vilahur, G.; Cubedo, J.; Casani, L.; Padro, T.; Sabate-Tenas, M.; Badimon, J.J.; Badimon, L. Reperfusion-triggered stress protein response in the myocardium is blocked by post-conditioning. Systems biology pathway analysis highlights the key role of the canonical aryl-hydrocarbon receptor pathway. Eur. Heart J. 2013, 34, 2082–2093. [Google Scholar] [CrossRef] [PubMed]
- Cubedo, J.; Vilahur, G.; Casani, L.; Mendieta, G.; Gomez-Jabalera, E.; Juan-Babot, O.; Padro, T.; Badimon, L. Targeting the molecular mechanisms of ischemic damage: Protective effects of alpha-crystallin-B. Int. J. Cardiol. 2016, 215, 406–416. [Google Scholar] [CrossRef] [PubMed]
- Theilmeier, G.; Schmidt, C.; Herrmann, J.; Keul, P.; Schafers, M.; Herrgott, I.; Mersmann, J.; Larmann, J.; Hermann, S.; Stypmann, J.; et al. High-density lipoproteins and their constituent, sphingosine-1-phosphate, directly protect the heart against ischemia/reperfusion injury in vivo via the S1P3 lysophospholipid receptor. Circulation 2006, 114, 1403–1409. [Google Scholar] [CrossRef] [PubMed]
- Vilahur, G.; Gutierrez, M.; Casani, L.; Cubedo, J.; Capdevila, A.; Pons-Llado, G.; Carreras, F.; Hidalgo, A.; Badimon, L. Hypercholesterolemia Abolishes High-Density Lipoprotein-Related Cardioprotective Effects in the Setting of Myocardial Infarction. J. Am. Coll. Cardiol. 2015, 66, 2469–2470. [Google Scholar] [CrossRef] [PubMed]
- Padro, T.; Cubedo, J.; Camino, S.; Bejar, M.T.; Ben-Aicha, S.; Mendieta, G.; Escola-Gil, J.C.; Escate, R.; Gutierrez, M.; Casani, L.; et al. Detrimental Effect of Hypercholesterolemia on High-Density Lipoprotein Particle Remodeling in Pigs. J. Am. Coll. Cardiol. 2017, 70, 165–178. [Google Scholar] [CrossRef] [PubMed]
- Porter, K.; Medford, H.M.; McIntosh, C.M.; Marsh, S.A. Cardioprotection requires flipping the ‘posttranslational modification’ switch. Life Sci. 2012, 90, 89–98. [Google Scholar] [CrossRef]
- Rubin, C.S.; Rosen, O.M. Protein phosphorylation. Annu. Rev. Biochem. 1975, 44, 831–887. [Google Scholar] [CrossRef]
- Datta, S.R.; Dudek, H.; Tao, X.; Masters, S.; Fu, H.; Gotoh, Y.; Greenberg, M.E. Akt phosphorylation of BAD couples survival signals to the cell-intrinsic death machinery. Cell 1997, 91, 231–241. [Google Scholar] [CrossRef]
- Barry, S.P.; Townsend, P.A.; Latchman, D.S.; Stephanou, A. Role of the JAK-STAT pathway in myocardial injury. Trends Mol. Med. 2007, 13, 82–89. [Google Scholar] [CrossRef]
- Gedik, N.; Kruger, M.; Thielmann, M.; Kottenberg, E.; Skyschally, A.; Frey, U.H.; Cario, E.; Peters, J.; Jakob, H.; Heusch, G.; et al. Proteomics/phosphoproteomics of left ventricular biopsies from patients with surgical coronary revascularization and pigs with coronary occlusion/reperfusion: Remote ischemic preconditioning. Sci. Rep. 2017, 7, 7629. [Google Scholar] [CrossRef] [PubMed]
- Marsh, S.A.; Chatham, J.C. The paradoxical world of protein O-GlcNAcylation: A novel effector of cardiovascular (dys)function. Cardiovasc. Res. 2011, 89, 487–488. [Google Scholar] [CrossRef] [PubMed]
- Laczy, B.; Marsh, S.A.; Brocks, C.A.; Wittmann, I.; Chatham, J.C. Inhibition of O-GlcNAcase in perfused rat hearts by NAG-thiazolines at the time of reperfusion is cardioprotective in an O-GlcNAc-dependent manner. Am. J. Physiol.-Heart Circ. Physiol. 2010, 299, H1715–H1727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ngoh, G.A.; Watson, L.J.; Facundo, H.T.; Jones, S.P. Augmented O-GlcNAc signaling attenuates oxidative stress and calcium overload in cardiomyocytes. Amino Acids 2011, 40, 895–911. [Google Scholar] [CrossRef] [PubMed]
- Mayr, M.; Liem, D.; Zhang, J.; Li, X.; Avliyakulov, N.K.; Yang, J.I.; Young, G.; Vondriska, T.M.; Ladroue, C.; Madhu, B.; et al. Proteomic and metabolomic analysis of cardioprotection: Interplay between protein kinase C epsilon and delta in regulating glucose metabolism of murine hearts. J. Mol. Cell. Cardiol. 2009, 46, 268–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bodi, V.; Sanchis, J.; Morales, J.M.; Marrachelli, V.G.; Nunez, J.; Forteza, M.J.; Chaustre, F.; Gomez, C.; Mainar, L.; Minana, G.; et al. Metabolomic profile of human myocardial ischemia by nuclear magnetic resonance spectroscopy of peripheral blood serum: A translational study based on transient coronary occlusion models. J. Am. Coll. Cardiol. 2012, 59, 1629–1641. [Google Scholar] [CrossRef] [PubMed]
- Kouassi Nzoughet, J.; Bocca, C.; Simard, G.; Prunier-Mirebeau, D.; Chao de la Barca, J.M.; Bonneau, D.; Procaccio, V.; Prunier, F.; Lenaers, G.; Reynier, P. A Nontargeted UHPLC-HRMS Metabolomics Pipeline for Metabolite Identification: Application to Cardiac Remote Ischemic Preconditioning. Anal. Chem. 2017, 89, 2138–2146. [Google Scholar] [CrossRef] [PubMed]
- Nadtochiy, S.M.; Urciuoli, W.; Zhang, J.; Schafer, X.; Munger, J.; Brookes, P.S. Metabolomic profiling of the heart during acute ischemic preconditioning reveals a role for SIRT1 in rapid cardioprotective metabolic adaptation. J. Mol. Cell. Cardiol. 2015, 88, 64–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, Y.T.; Depre, C.; Yan, L.; Park, J.Y.; Tian, B.; Jain, K.; Chen, L.; Zhang, Y.; Kudej, R.K.; Zhao, X.; et al. Repetitive ischemia by coronary stenosis induces a novel window of ischemic preconditioning. Circulation 2008, 118, 1961–1969. [Google Scholar] [CrossRef] [PubMed]
- Bravo, C.; Kudej, R.K.; Yuan, C.; Yoon, S.; Ge, H.; Park, J.Y.; Tian, B.; Stanley, W.C.; Vatner, S.F.; Vatner, D.E.; et al. Metabolomic analysis of two different models of delayed preconditioning. J. Mol. Cell. Cardiol. 2013, 55, 19–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gregorich, Z.R.; Cai, W.; Lin, Z.; Chen, A.J.; Peng, Y.; Kohmoto, T.; Ge, Y. Distinct sequences and post-translational modifications in cardiac atrial and ventricular myosin light chains revealed by top-down mass spectrometry. J. Mol. Cell. Cardiol. 2017, 107, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Ronnebaum, S.M.; Patterson, C. The FoxO family in cardiac function and dysfunction. Annu. Rev. Physiol. 2010, 72, 81–94. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Tan, C.Y.; Oresic, M.; Vidal-Puig, A. Integrating post-genomic approaches as a strategy to advance our understanding of health and disease. Genome Med. 2009, 1, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Animal | Strengths | Weaknesses |
|---|---|---|
| Rodents |
|
|
| Swine |
|
|
| Mice | microRNA | Associated Function | Reference |
|---|---|---|---|
| Pre-conditioning | miR21 | Related with apoptosis | [43] |
| miR451 | Related with oxidative stress | [44] | |
| Post-conditioning | miR21 | Related with apoptosis | [45] |
| miR499 | Related with apoptosis | [46] | |
| Swine | |||
| Post-conditioning | miR29b | Muscle specific | [47] |
| miR133a | Related with fibrosis | [47] | |
| miR146b | Related with inflammation | [47] | |
| miR92a | Related with angiogenesis | [48,49] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Badimon, L.; Mendieta, G.; Ben-Aicha, S.; Vilahur, G. Post-Genomic Methodologies and Preclinical Animal Models: Chances for the Translation of Cardioprotection to the Clinic. Int. J. Mol. Sci. 2019, 20, 514. https://doi.org/10.3390/ijms20030514
Badimon L, Mendieta G, Ben-Aicha S, Vilahur G. Post-Genomic Methodologies and Preclinical Animal Models: Chances for the Translation of Cardioprotection to the Clinic. International Journal of Molecular Sciences. 2019; 20(3):514. https://doi.org/10.3390/ijms20030514
Chicago/Turabian StyleBadimon, Lina, Guiomar Mendieta, Soumaya Ben-Aicha, and Gemma Vilahur. 2019. "Post-Genomic Methodologies and Preclinical Animal Models: Chances for the Translation of Cardioprotection to the Clinic" International Journal of Molecular Sciences 20, no. 3: 514. https://doi.org/10.3390/ijms20030514
APA StyleBadimon, L., Mendieta, G., Ben-Aicha, S., & Vilahur, G. (2019). Post-Genomic Methodologies and Preclinical Animal Models: Chances for the Translation of Cardioprotection to the Clinic. International Journal of Molecular Sciences, 20(3), 514. https://doi.org/10.3390/ijms20030514