Global Transcriptional Insights of Pollen-Pistil Interactions Commencing Self-Incompatibility and Fertilization in Tea [Camellia sinensis (L.) O. Kuntze]

 ,
,  , ,
, ,
Abstract
:
1. Introduction
2. Results
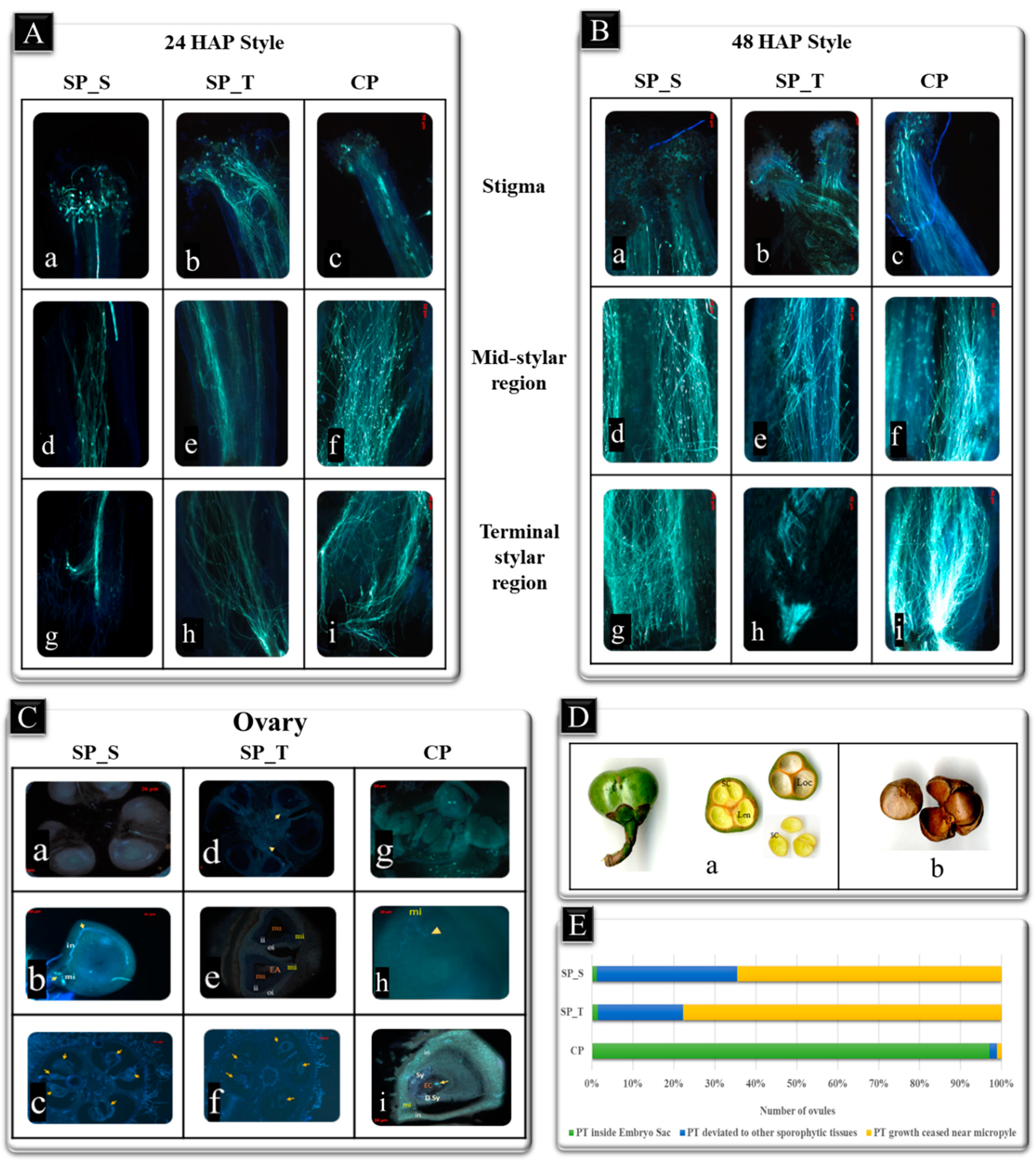
2.1. Field Study and Microscopy Analysis
2.2. Illumina Sequencing, Sequence Assembly and Functional Annotation
2.3. Global Transcripts Expression Dynamics and Gene Ontology Enrichment Analysis
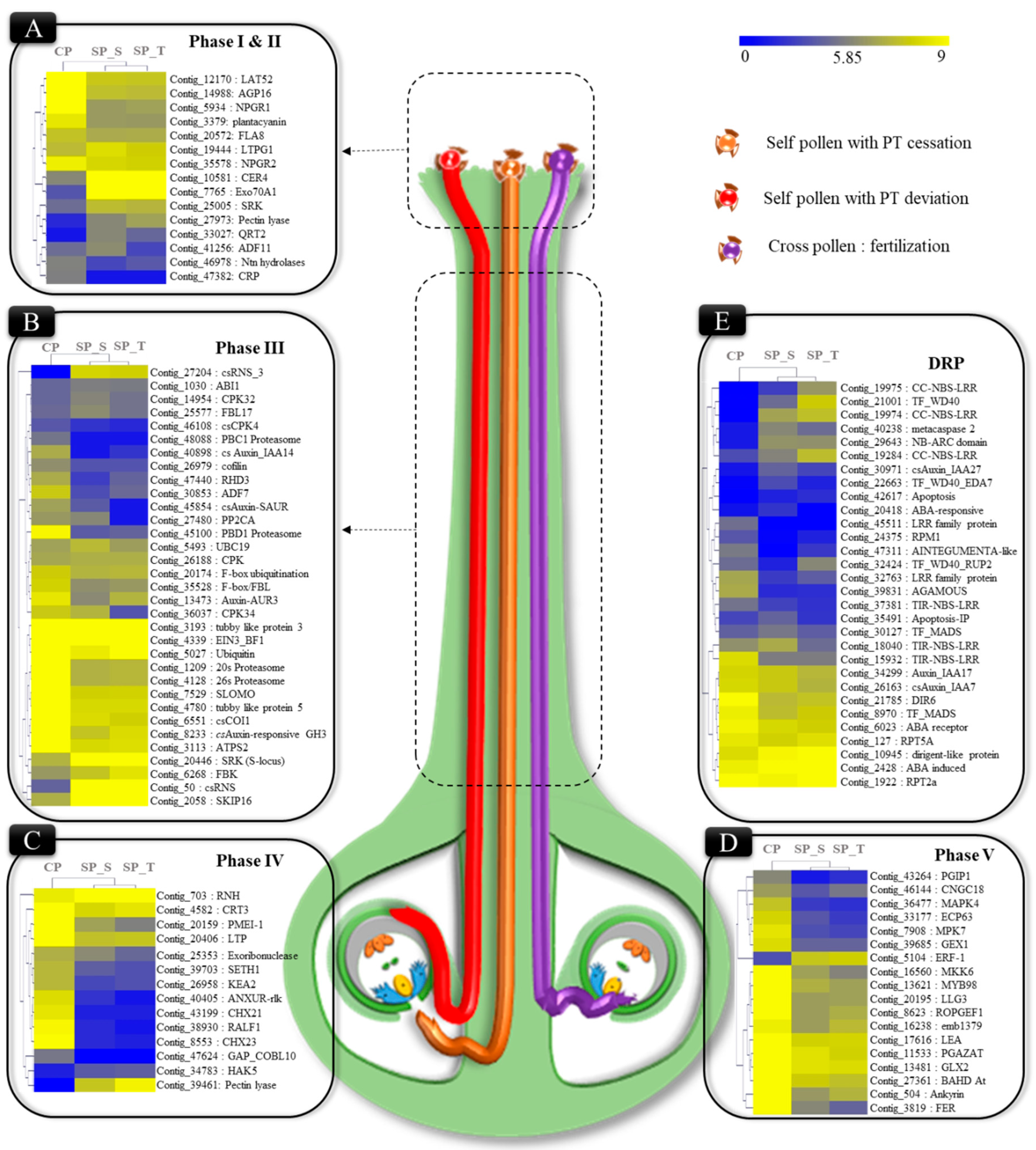
2.4. Phase Specific Differentially Expressed Transcripts Involved in Pollen-Pistil Interaction
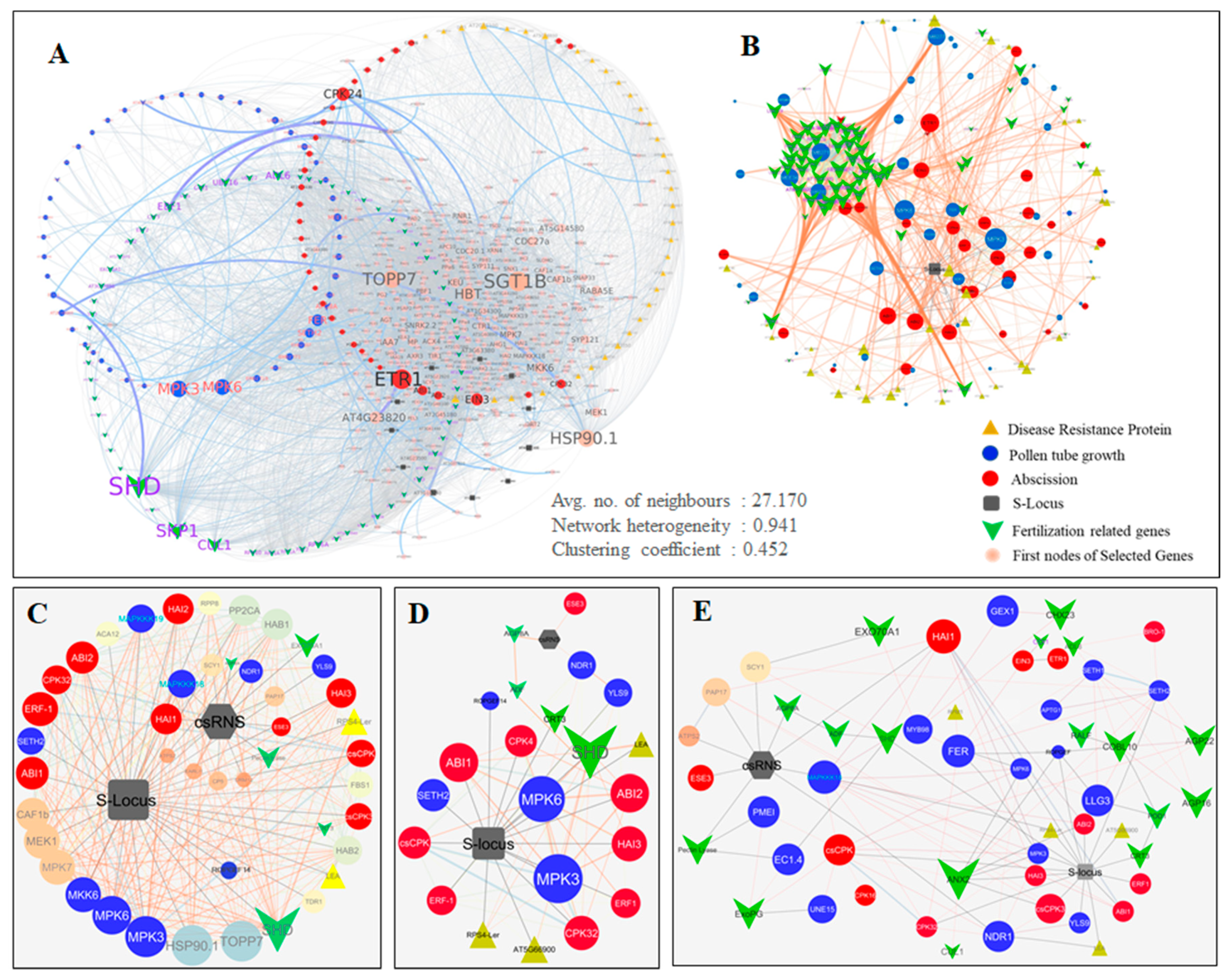
2.5. Protein-Protein Interactome Network Analysis
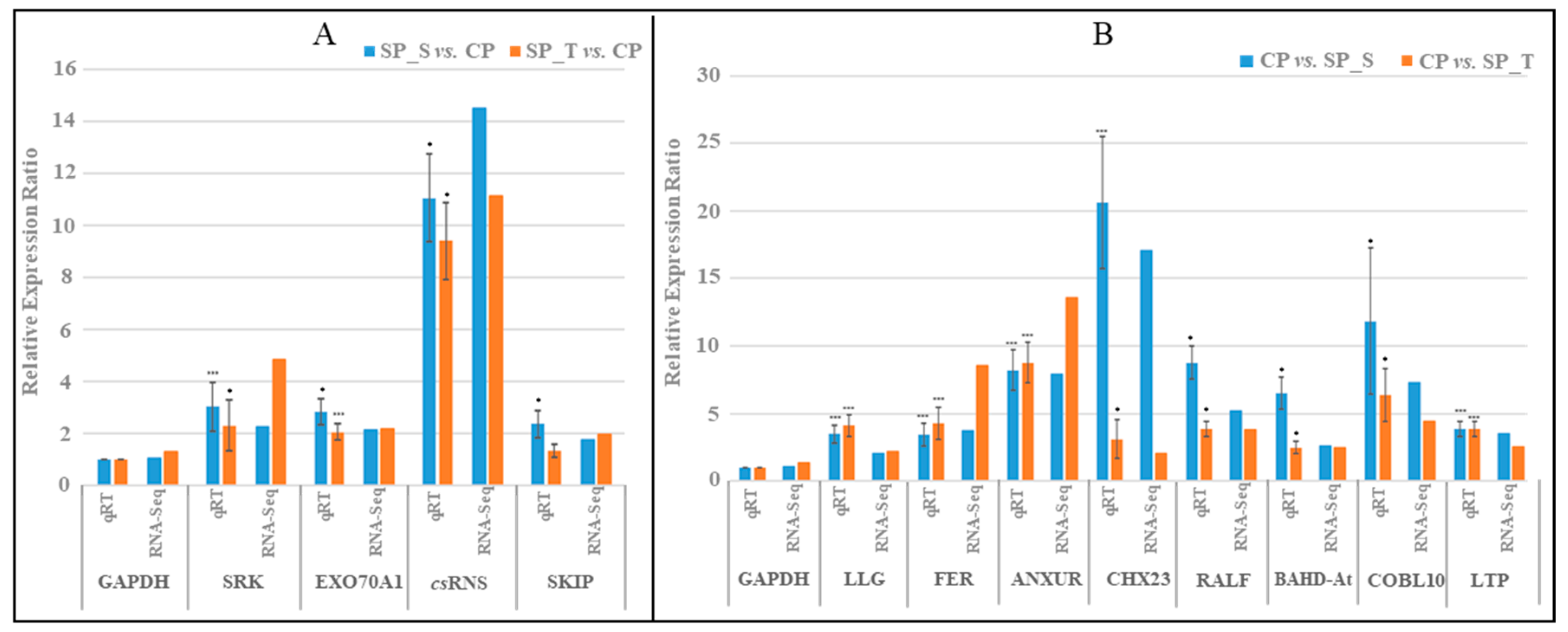
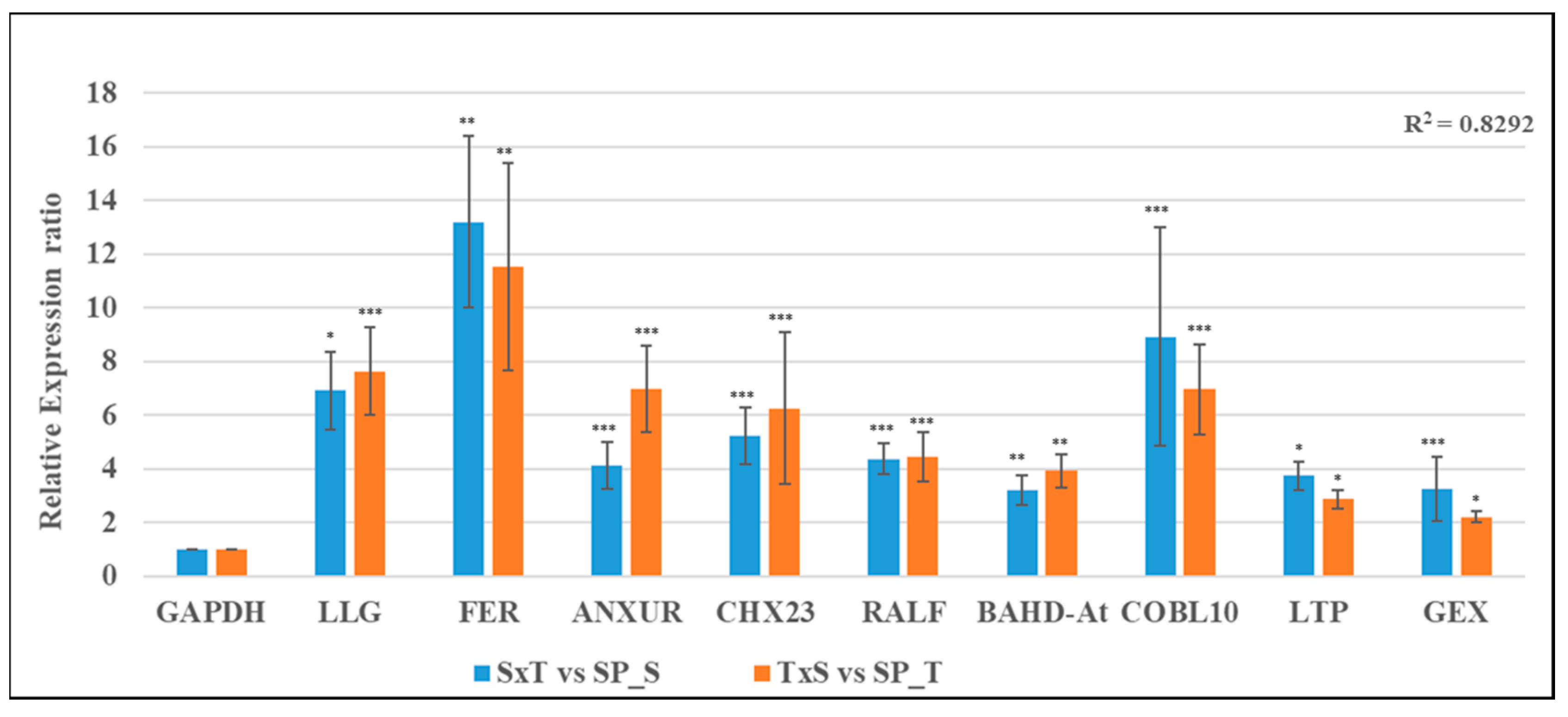
2.6. RNA-Seq Data Validation by qRT-PCR
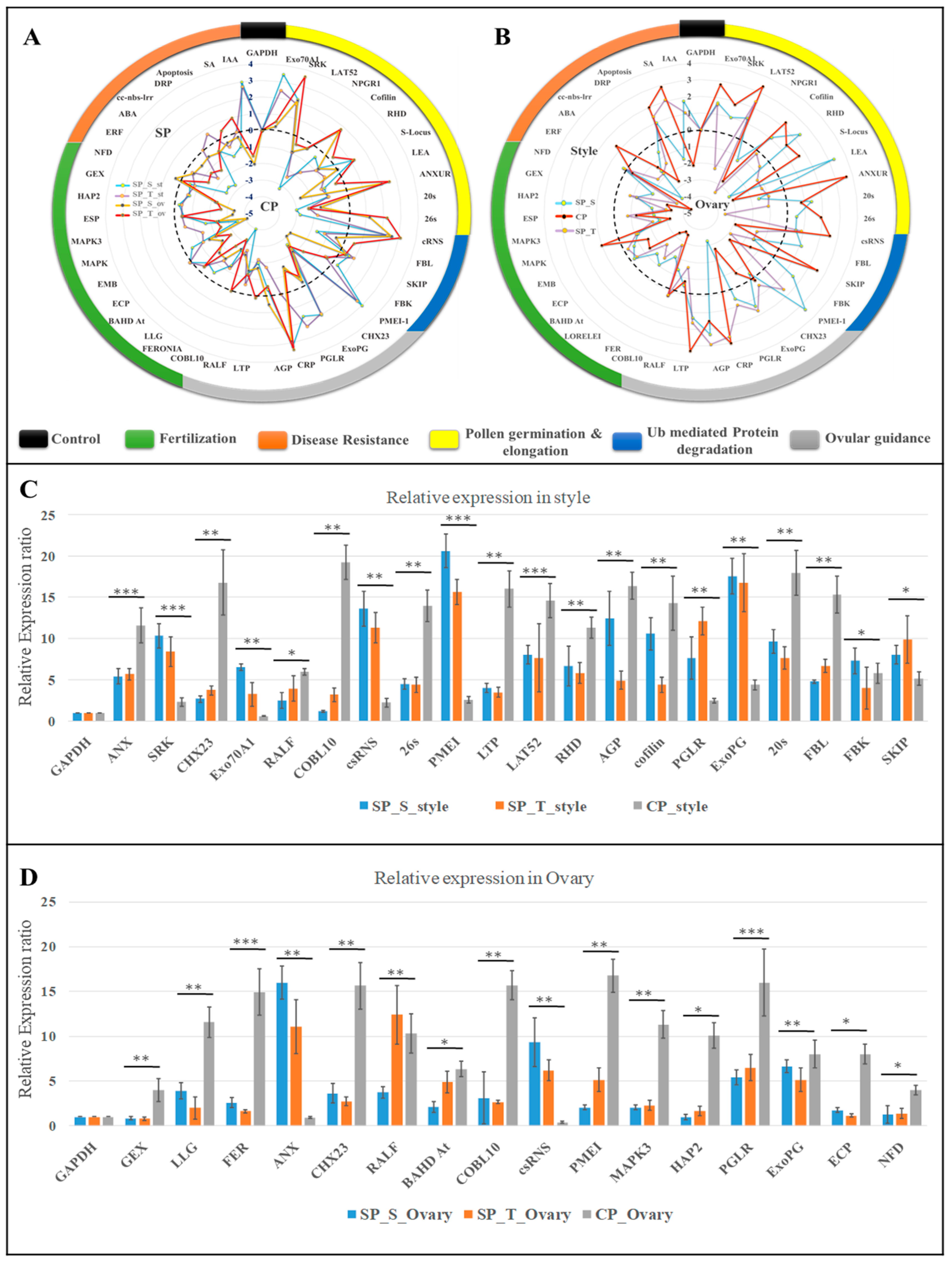
2.7. Tissue-Specific qRT-PCR Expression Dynamics during Pollen-Pistil Interaction
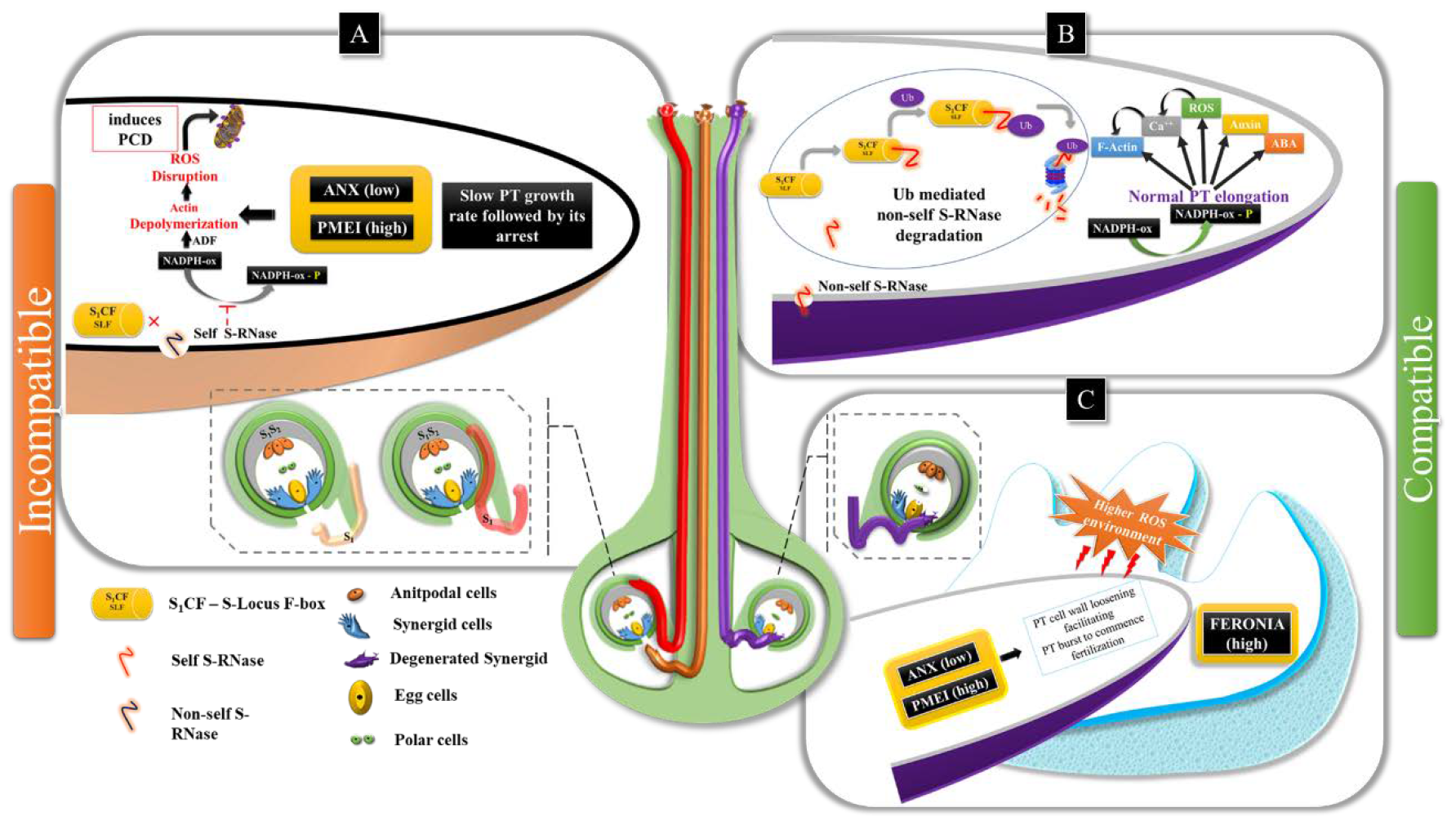
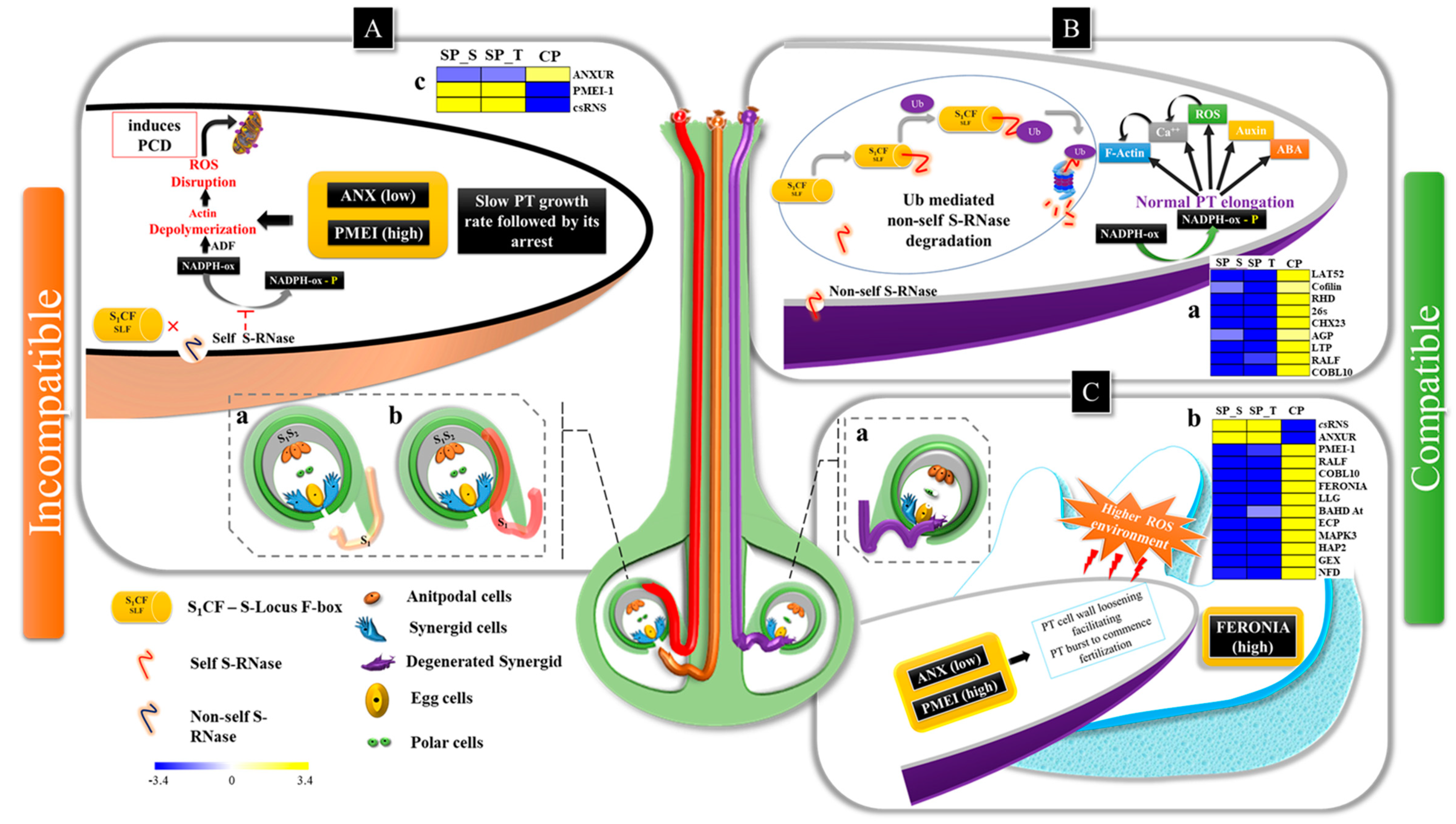
3. Discussion
3.1. Pollen Germination & PT Elongation (Phase I-III)
3.2. PT-Ovular Guidance (PHASE IV & V)
4. Materials and Methods
4.1. Plant Material
4.2. Microscopic Analysis
4.3. RNA Extraction, cDNA Library Preparation And Illumina Sequencing
4.4. Quality Filtering, Sequence Assembly and Differential Expression Dynamics
4.5. Transcripts Homology, Functional Classification and GO Enrichment Analysis
4.6. Protein-Protein Interactome Network Analysis
4.7. RNA-Seq Expression Pattern Validation Using Real-Time PCR
4.8. Tissue-Specific Transcript Expression Analysis Using qRT-PCR
5. Conclusions
Supplementary Materials
Data availability and material
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| SI | Self-incompatibility |
| LSI | Late-acting gametophytic self-incompatibility |
| CC | Cross-compatibility (Fertilization) |
| SP | Self-Pollinated |
| CP | Cross-pollinated |
| PT | Pollen tube |
| HAP | Hours after pollination |
| DAP | Days after pollination |
| KEGG | Kyoto encyclopedia of genes and genomes |
| GO | Gene ontology |
| DGE | Differential gene expression |
| NGS | next generation sequencing |
| SLF/SCF | S-locus F-box protein |
| SRK | S-receptor kinase |
| CPK | Calcium-dependent protein kinases |
| TLP | Tubby like proteins |
| RHD | Root hair defective |
| PMEI | Pectin methylesterase inhibitor |
| GEX | Gamete expressed |
| ARF | auxin response factors |
| DRP | Disease resistance proteins |
| RALF | Rapid alkalization factors |
| LLG | LORELLEI like glycoprotein |
| GPI-Ap | Glycosylphosphatidylinositol anchored protein |
| CRP | Cystein rich protein |
| qRT-PCR | Quantitative Real-Time PCR |
References
- Darwin, C. The Effects of Cross and Self Fertilisation in the Vegetable Kingdom; Cambridge University Press: Cambridge, UK, 1876. [Google Scholar]
- De Nettancourt, D. Incompatibility in angiosperms. Sex. Plant Reprod. 1997, 10, 185–199. [Google Scholar] [CrossRef]
- Chapman, L.A.; Goring, D.R. Pollen-pistil interactions regulating successful fertilization in the Brassicaceae. J. Exp. Bot. 2010, 61, 1987–1999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibbs, P.E. Late-acting self-incompatibility-the pariah breeding system in flowering plants. New Phytol. 2014, 203, 717–734. [Google Scholar] [CrossRef] [PubMed]
- Lewis, D. Comparative Incompatibility in Angiosperms and Fungi. Adv. Genet. 1954, 6, 235–285. [Google Scholar] [CrossRef]
- Bateman, A.J. Self-incompatibility systems in angiosperms II. Iberis amara. Heredity 1954, 8, 305–332. [Google Scholar] [CrossRef] [Green Version]
- Haasen, K.E.; Goring, D.R. The recognition and rejection of self-incompatible pollen in the Brassicaceae. Bot. Stud. 2010, 51, 1–6. [Google Scholar]
- Samuel, M.A.; Chong, Y.T.; Haasen, K.E.; Aldea-Brydges, M.G.; Stone, S.L.; Goring, D.R. Cellular pathways regulating responses to compatible and self-incompatible pollen in Brassica and Arabidopsis stigmas intersect at Exo70A1, a putative component of the exocyst complex. Plant Cell 2009, 21, 2655–2671. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Williams, J.S.; Li, S.; Wu, L.; Khatri, W.A.; Stone, P.G.; Keebaugh, M.D.; Kao, T. S-Locus F-Box Proteins Are Solely Responsible for Pollen Function in S-RNase-Based Self-Incompatibility of Petunia. Plant Cell 2018, tpc.00615. [Google Scholar] [CrossRef]
- McClure, B.A.; Franklin-Tong, V. Gametophytic self-incompatibility: Understanding the cellular mechanisms involved in “self” pollen tube inhibition. Planta 2006, 224, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Pease, J.B.; Guerrero, R.F.; Sherman, N.A.; Hahn, M.W. Molecular mechanisms of postmating prezygotic reproductive isolation uncovered by transcriptome analysis. Mol. Ecol. 2016, 25, 2592–2608. [Google Scholar] [CrossRef] [Green Version]
- Wilkins, K.A.; Bosch, M.; Haque, T.; Teng, N.; Poulter, N.S.; Franklin-Tong, V.E. Self-incompatibility-induced programmed cell death in field poppy pollen involves dramatic acidification of the incompatible pollen tube cytosol. Plant Physiol. 2015, 167, 766–779. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Meng, D.; Gu, Z.; Li, W.; Yuan, H.; Duan, X.; Yang, Q.; Li, Y.; Li, T. SLFL Genes Participate in the Ubiquitination and Degradation Reaction of S-RNase in Self-compatible Peach. Front. Plant Sci. 2018, 9, 227. [Google Scholar] [CrossRef] [PubMed]
- Seavey, S.R.; Bawa, K.S. Late-acting self-incompatibility in angiosperms. Bot. Rev. 1986, 52, 195–219. [Google Scholar] [CrossRef]
- Liao, T.; Yuan, D.-Y.; Zou, F.; Gao, C.; Yang, Y.; Zhang, L.; Tan, X.-F. Self-sterility in Camellia oleifera may be due to the prezygotic late-acting self-incompatibility. PLoS ONE 2014, 9, e99639. [Google Scholar] [CrossRef]
- Allen, A.M.; Hiscock, S.J. Evolution and phylogeny of self-incompatibility systems in angiosperms. In Self-Incompatibility in Flowering Plants; Springer Verlag: Berlin, Germany, 2008; pp. 73–101. [Google Scholar]
- Preedy, V.R. Tea in Health and Disease Prevention; Academic Press: Cambrage, MA, USA, 2012; ISBN 0123849381. [Google Scholar]
- Singh, S.; Sud, R.K.; Gulati, A.; Joshi, R.; Yadav, A.K.; Sharma, R.K. Germplasm appraisal of western Himalayan tea: A breeding strategy for yield and quality improvement. Genet. Resour. Crop Evol. 2013, 60, 1501–1513. [Google Scholar] [CrossRef]
- Sharma, R.K.; Negi, M.S.; Sharma, S.; Bhardwaj, P.; Kumar, R.; Bhattachrya, E.; Tripathi, S.B.; Vijayan, D.; Baruah, A.R.; Das, S.C. AFLP-based genetic diversity assessment of commercially important tea germplasm in India. Biochem. Genet. 2010, 48, 549–564. [Google Scholar] [CrossRef] [PubMed]
- Raina, S.N.; Ahuja, P.S.; Sharma, R.K.; Das, S.C.; Bhardwaj, P.; Negi, R.; Sharma, V.; Singh, S.S.; Sud, R.K.; Kalia, R.K. Genetic structure and diversity of India hybrid tea. Genet. Resour. Crop Evol. 2012, 59, 1527–1541. [Google Scholar] [CrossRef]
- Sharma, H.; Kumar, R.; Sharma, V.; Kumar, V.; Bhardwaj, P.; Ahuja, P.S.; Kumar, R. Identification and cross-species transferability of 112 novel unigene-derived microsatellite markers in tea (Camellia sinensis). Am. J. Bot. 2011, 98, 133–138. [Google Scholar] [CrossRef]
- Unamba, C.I.N.; Nag, A.; Sharma, R.K. Next Generation Sequencing technologies: The doorway to the unexplored genomics of non-model plants. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef]
- Jayaswall, K.; Mahajan, P.; Singh, G.; Parmar, R.; Seth, R.; Raina, A.; Swarnkar, M.K.; Singh, A.K.; Shankar, R.; Sharma, R.K. Transcriptome Analysis Reveals Candidate Genes involved in Blister Blight defense in Tea (Camellia sinensis (L.) Kuntze). Sci. Rep. 2016, 6. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, L.; Wei, K.; Wu, L.; Li, H.; Zhang, F.; Cheng, H. Transcriptome analysis reveals self-incompatibility in the tea plant (Camellia sinensis) might be under gametophytic control. BMC Genom. 2016, 17, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Chen, C.; Zeng, Z.; Zou, Z.; Li, H.; Zhou, Q.; Chen, X.; Sun, K.; Li, X. Transcriptomic analysis between self-and cross-pollinated pistils of tea plants (Camellia sinensis). BMC Genom. 2018, 19, 289. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Hao, S.; Wang, L.; Fang, W.; Wang, Y.; Li, X. Late-acting self-incompatibility in tea plant (Camellia sinensis). Biol. Sect. Bot. 2012, 672, 347–351. [Google Scholar] [CrossRef]
- Heslop-Harrison, Y.; Shivanna, K.R. The receptive surface of the angiosperm stigma. Ann. Bot. 1977, 41, 1233–1258. [Google Scholar] [CrossRef]
- Xia, E.-H.; Zhang, H.-B.; Sheng, J.; Li, K.; Zhang, Q.-J.; Kim, C.; Zhang, Y.; Liu, Y.; Zhu, T.; Li, W. The Tea Tree Genome Provides Insights into Tea Flavor and Independent Evolution of Caffeine Biosynthesis. Mol. Plant 2017, 10, 866–877. [Google Scholar] [CrossRef] [PubMed]
- Dresselhaus, T.; Franklin-Tong, N. Male-female crosstalk during pollen germination, tube growth and guidance, and double fertilization. Mol. Plant 2013, 6, 1018–1036. [Google Scholar] [CrossRef] [PubMed]
- Bhandawat, A.; Singh, G.; Seth, R.; Singh, P.; Sharma, R.K.R.K. Genome-wide transcriptional profiling to elucidate key candidates involved in bud burst and rattling growth in a subtropical bamboo (Dendrocalamus hamiltonii). Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Kusonmano, K. Gene Expression Analysis Through Network Biology: Bioinformatics Approaches. In Network Biology; Springer Nature: Basingstoke, UK, 2016; pp. 15–32. [Google Scholar]
- Dresselhaus, T.; Sprunck, S.; Wessel, G.M. Fertilization mechanisms in flowering plants. Curr. Biol. 2016, 26, R125–R139. [Google Scholar] [CrossRef] [PubMed]
- Muschietti, J.; Dircks, L.; Vancanneyt, G.; McCormick, S. LAT52 protein is essential for tomato pollen development: Pollen expressing antisense LAT52 RNA hydrates and germinates abnormally and cannot achieve fertilization. Plant J. 1994, 6, 321–338. [Google Scholar] [CrossRef] [PubMed]
- Boisson-Dernier, A.; Roy, S.; Kritsas, K.; Grobei, M.A.; Jaciubek, M.; Schroeder, J.I.; Grossniklaus, U. Disruption of the pollen-expressed FERONIA homologs ANXUR1 and ANXUR2 triggers pollen tube discharge. Development 2009, 136, 3279–3288. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Ge, F.R.; Xu, M.; Zhao, X.Y.; Huang, G.Q.; Zhou, L.Z.; Wang, J.G.; Kombrink, A.; McCormick, S.; Zhang, X.S.; Zhang, Y. Arabidopsis COBRA-LIKE 10, a GPI-anchored protein, mediates directional growth of pollen tubes. Plant J. 2013, 74, 486–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Y.-Y.; Lin, X.-J.; Liang, H.-M.; Wang, F.-F.; Chen, L.-Y. The Long Journey of Pollen Tube in the Pistil. Int. J. Mol. Sci. 2018, 19, 3529. [Google Scholar] [CrossRef] [PubMed]
- Cerveau, N.; Jackson, D.J. Combining independent de novo assemblies optimizes the coding transcriptome for nonconventional model eukaryotic organisms. BMC Bioinform. 2016, 17, 525. [Google Scholar] [CrossRef] [PubMed]
- Kovi, M.R.; Amdahl, H.; Alsheikh, M.; Rognli, O.A. De novo and reference transcriptome assembly of transcripts expressed during flowering provide insight into seed setting in tetraploid red clover. Sci. Rep. 2017, 7, 44383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, X.; Li, W.; Liu, Y.; Tan, M.; Ganal, M.; Chetelat, R.T. A farnesyl pyrophosphate synthase gene expressed in pollen functions in S-RNase-independent unilateral incompatibility. Plant J. 2018, 93, 417–430. [Google Scholar] [CrossRef] [PubMed]
- Park, S.-W.; Kang, S.-W.; Goo, T.-W.; Kim, S.-R.; Lee, G.; Paik, S.-Y. Tissue-specific gene expression analysis of silkworm (Bombyx mori) by quantitative real-time RT-PCR. BMB Rep. 2010, 43, 480–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, D.; Tang, C.; Wang, R.; Gu, C.; Wu, X.; Hu, S.; Jiao, J.; Zhang, S. Transcriptome and phytohormone analysis reveals a comprehensive phytohormone and pathogen defence response in pear self-/cross-pollination. Plant Cell Rep. 2017, 36, 1785–1799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higashiyama, T.; Takeuchi, H. The Mechanism and Key Molecules Involved in Pollen Tube Guidance. Annu. Rev. Plant Biol. 2015, 66, 393–413. [Google Scholar] [CrossRef] [PubMed]
- McClure, B. Darwin’s foundation for investigating self-incompatibility and the progress toward a physiological model for S.-RNase-based SI. J. Exp. Bot. 2009, 60, 1069–1081. [Google Scholar] [CrossRef] [PubMed]
- Onelli, E.; Idilli, A.I.; Moscatelli, A. Emerging roles for microtubules in angiosperm pollen tube growth highlight new research cues. Front. Plant Sci. 2015, 6, 51. [Google Scholar] [CrossRef]
- Boisson-Dernier, A.; Lituiev, D.S.; Nestorova, A.; Franck, C.M.; Thirugnanarajah, S.; Grossniklaus, U. ANXUR Receptor-Like Kinases Coordinate Cell Wall Integrity with Growth at the Pollen Tube Tip Via NADPH Oxidases. PLoS Biol. 2013, 11. [Google Scholar] [CrossRef]
- Geitmann, A.; Palanivelu, R. Fertilization Requires Communication: Signal Generation and Perception During Pollen Tube Guidance. Floric. Ornam. Biotechnol. 2007, 1, 77–89. [Google Scholar]
- Woriedh, M.; Wolf, S.; Márton, M.L.; Hinze, A.; Gahrtz, M.; Becker, D.; Dresselhaus, T. External application of gametophyte-specific ZmPMEI1 induces pollen tube burst in maize. Plant Reprod. 2013, 26, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Qu, L.J.; Li, L.; Lan, Z.; Dresselhaus, T. Peptide signalling during the pollen tube journey and double fertilization. J. Exp. Bot. 2015, 66, 5139–5150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhardwaj, P.; Kumar, R.; Sharma, H.; Tewari, R.; Ahuja, P.S.; Sharma, R.K. Development and utilization of genomic and genic microsatellite markers in Assam tea (Camellia assamica ssp. assamica) and related Camellia species. Plant Breed. 2013, 132, 748–763. [Google Scholar] [CrossRef]
- Martin, F.W. Staining and observing pollen tubes in the style by means of fluorescence. Stain Technol. 1959, 34, 125–128. [Google Scholar] [CrossRef]
- Ghawana, S.; Paul, A.; Kumar, H.; Kumar, A.; Singh, H.; Bhardwaj, P.K.; Rani, A.; Singh, R.S.; Raizada, J.; Singh, K.; Kumar, S. An RNA isolation system for plant tissues rich in secondary metabolites. BMC Res. Notes 2011, 4, 85. [Google Scholar] [CrossRef]
- Patel, R.K.; Jain, M. NGS QC Toolkit: A toolkit for quality control of next generation sequencing data. PLoS ONE 2012, 7, e30619. [Google Scholar] [CrossRef]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M. De novo transcript sequence reconstruction from RNA-seq using the Trinity platform for reference generation and analysis. Nat. Protoc. 2013, 8, 1494–1512. [Google Scholar] [CrossRef] [Green Version]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. CD-HIT: Accelerated for clustering the next-generation sequencing data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Lun, A.T.L.; Smyth, G.K. Differential expression analysis of complex RNA-seq experiments using edgeR. In Statistical Analysis of Next Generation Sequencing Data; Datta, S., Nettleton, D., Eds.; Springer International Publishing: New York, NY, USA, 2014; pp. 51–74. [Google Scholar]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, Z.; Zhou, X.; Ling, Y.; Zhang, Z.; Su, Z. agriGO: A GO analysis toolkit for the agricultural community. Nucleic Acids Res. 2010, 45, gkq310. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Franceschini, A.; Wyder, S.; Forslund, K.; Heller, D.; Huerta-Cepas, J.; Simonovic, M.; Roth, A.; Santos, A.; Tsafou, K.P. STRING v10: Protein–protein interaction networks, integrated over the tree of life. Nucleic Acids Res. 2014, 43, gku1003. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative expression software tool (REST©) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002, 30, e36. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Outgoing Interactions | Incoming Interactions | ||||
|---|---|---|---|---|---|
| PT Germination & Elongation | S-Locus Related | Fertilization | DRP | Abscission | |
| PT germination & elongation | 67 | 3 | 26 | 29 | 16 |
| S-locus related | 2 | 11 | 5 | 3 | 8 |
| Fertilization | 57 | 11 | 43 | 5 | 26 |
| DRP | 7 | 2 | 6 | 30 | 4 |
| abscission | 27 | 10 | 20 | 2 | 31 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seth, R.; Bhandawat, A.; Parmar, R.; Singh, P.; Kumar, S.; Sharma, R.K. Global Transcriptional Insights of Pollen-Pistil Interactions Commencing Self-Incompatibility and Fertilization in Tea [Camellia sinensis (L.) O. Kuntze]. Int. J. Mol. Sci. 2019, 20, 539. https://doi.org/10.3390/ijms20030539
Seth R, Bhandawat A, Parmar R, Singh P, Kumar S, Sharma RK. Global Transcriptional Insights of Pollen-Pistil Interactions Commencing Self-Incompatibility and Fertilization in Tea [Camellia sinensis (L.) O. Kuntze]. International Journal of Molecular Sciences. 2019; 20(3):539. https://doi.org/10.3390/ijms20030539
Chicago/Turabian StyleSeth, Romit, Abhishek Bhandawat, Rajni Parmar, Pradeep Singh, Sanjay Kumar, and Ram Kumar Sharma. 2019. "Global Transcriptional Insights of Pollen-Pistil Interactions Commencing Self-Incompatibility and Fertilization in Tea [Camellia sinensis (L.) O. Kuntze]" International Journal of Molecular Sciences 20, no. 3: 539. https://doi.org/10.3390/ijms20030539
APA StyleSeth, R., Bhandawat, A., Parmar, R., Singh, P., Kumar, S., & Sharma, R. K. (2019). Global Transcriptional Insights of Pollen-Pistil Interactions Commencing Self-Incompatibility and Fertilization in Tea [Camellia sinensis (L.) O. Kuntze]. International Journal of Molecular Sciences, 20(3), 539. https://doi.org/10.3390/ijms20030539