Enhanced Expression of QTL qLL9/DEP1 Facilitates the Improvement of Leaf Morphology and Grain Yield in Rice





 ,
, 
Abstract
:1. Introduction
2. Results
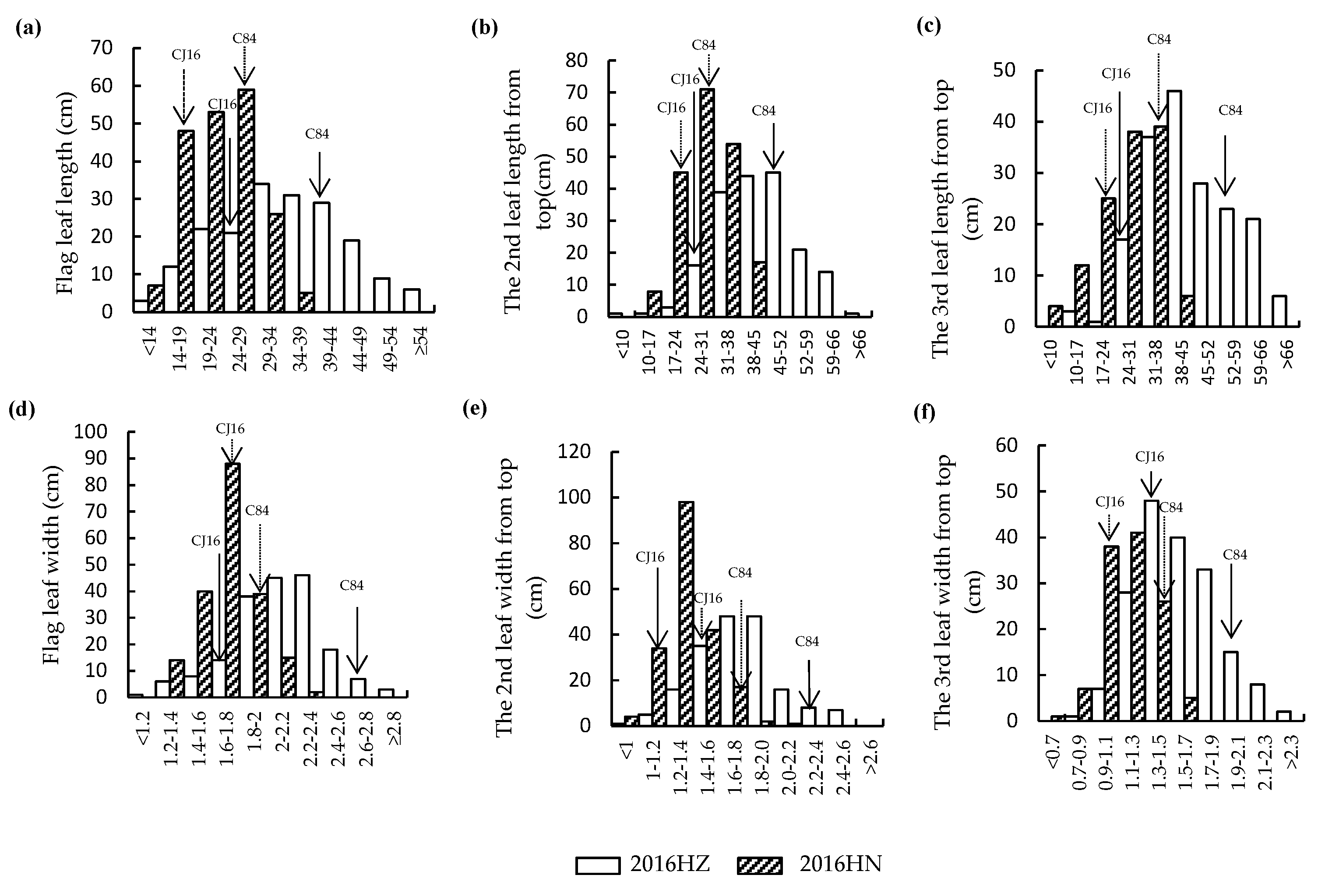
2.1. Analysis of Leaf Morphology in the RIL Population and Their Parental Cultivars
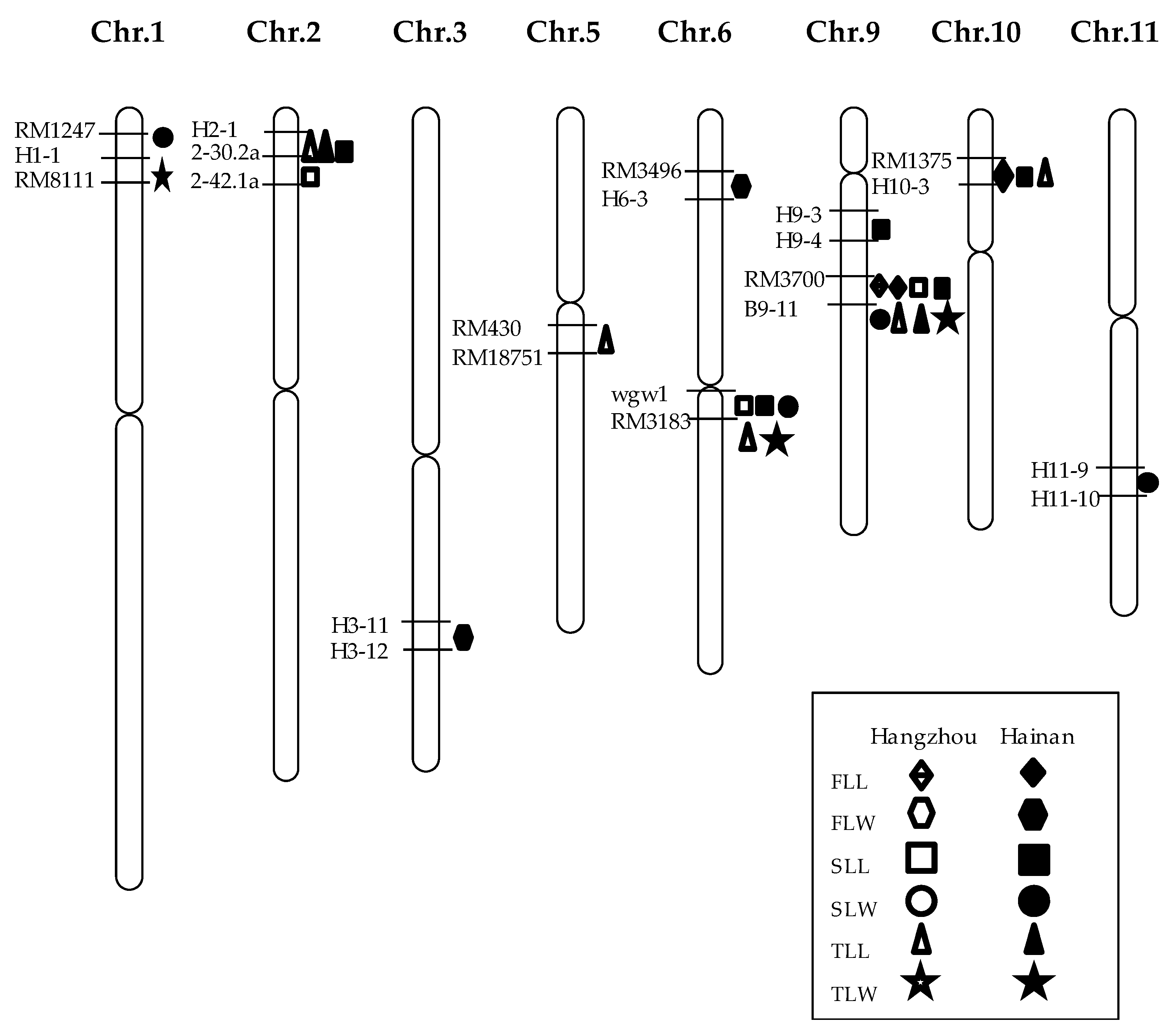
2.2. Correlation Analysis and QTL Mapping
2.3. Fine Mapping and Leaf Shape Characterization of qLL9
2.4. Determination of the Candidate Gene
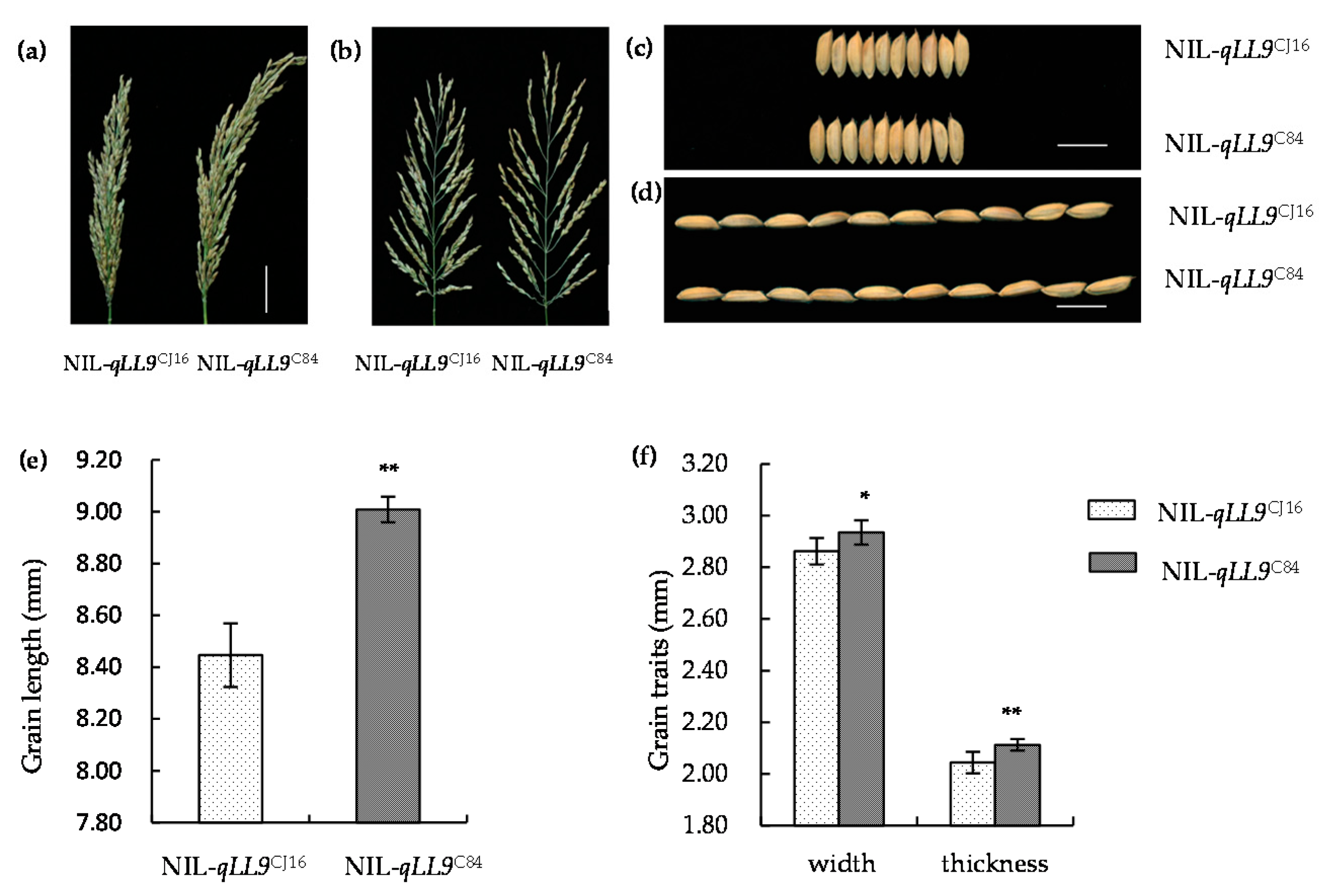
2.5. qLL9 Affecting the Yield Traits
3. Discussion
4. Materials and Methods
4.1. RIL Population and Field Trial
4.2. Statistical and Genetic Analysis
4.3. Map-Based Cloning and Candidate-Gene Promoter Activity for qLL9
4.4. RNA Extraction and qRT-PCR
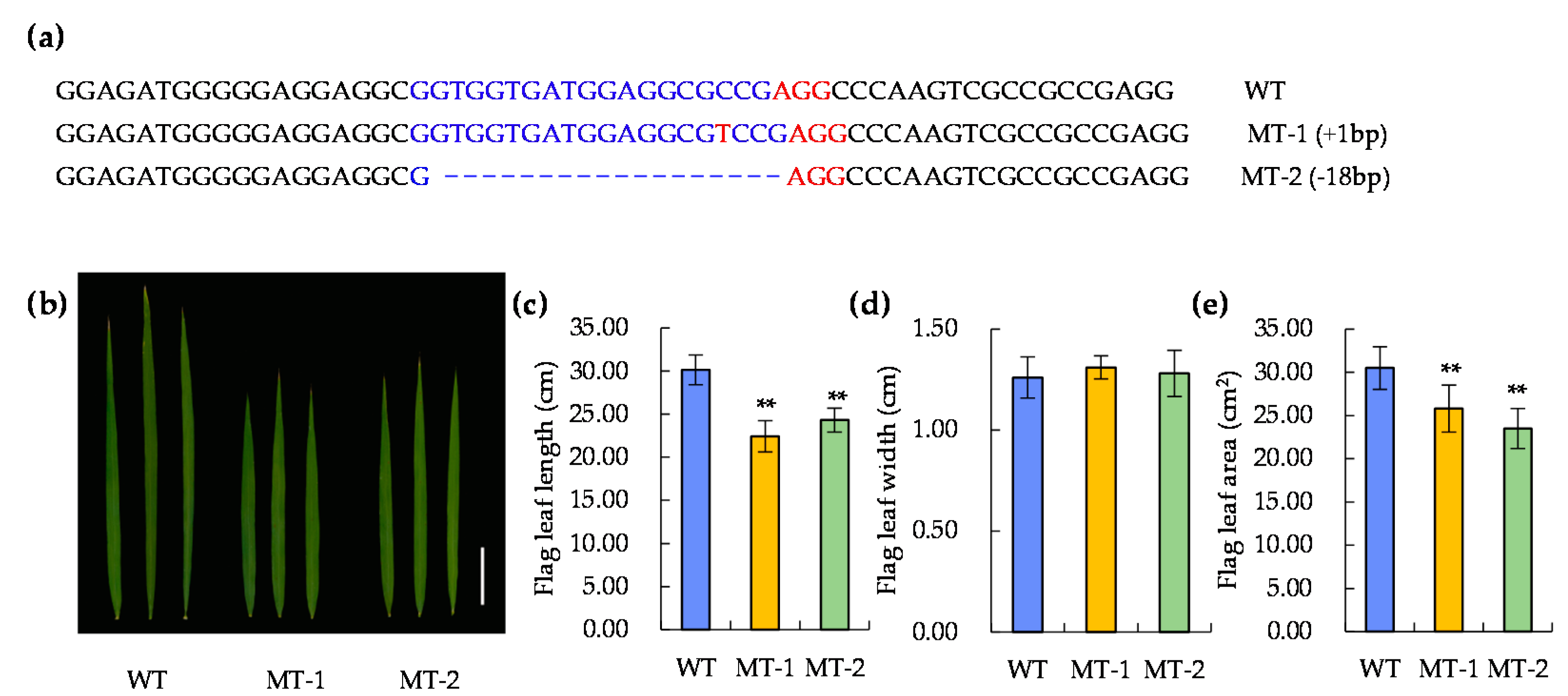
4.5. CRISPR/Cas9 Transgene Analysis
4.6. Construction of Near Isogenic Lines and Chromosome Segment Substitution Lines and Trait Measurement
4.7. Morphological Observation on Leaf Epidermal Cytology
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Heath, D.V.; Gregory, F.G. The constancy of the mean net assimilation rate and its ecological importance. Ann. Bot. Lond. 1938, 2, 811–818. [Google Scholar] [CrossRef]
- Donald, C.M. The breeding of crop ideotypes. Euphytica 1968, 17, 385–403. [Google Scholar] [CrossRef]
- Mason, T.G.; Maskell, E.J. Studies on the transport of carbohydrates in the cotton plant. I. A study of diurnal variation in the carbohydrates of leaf, bark, and wood, and of the effects of ringing. Ann. Bot.-Lond. 1928, 42, 189–253. [Google Scholar] [CrossRef]
- Hasson, A.; Blein, T.; Laufs, P. Leaving the meristem behind: The genetic and molecular control of leaf patterning and morphogenesis. C. R. Biol. 2010, 333, 350–360. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Li, P.C.; Li, X.; Liu, C.Y.; Cao, S.Y.; Chu, C.C.; Cao, X.F. Loss of function of OsDCL1 affects micro RNA accumulation and causes developmental defects in rice. Plant Physiol. 2005, 139, 296–305. [Google Scholar] [CrossRef] [PubMed]
- Wang, E.; Xu, X.; Zhang, L.; Zhang, H.; Lin, L.; Wang, Q.; Li, Q.; Ge, S.; Lu, B.R.; Wang, W.; et al. Duplication and independent selection of cell-wall invertase genes GIF1 and OsCIN1 during rice evolution and domestication. BMC Evol. Biol. 2010, 10, 108–120. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.F.; Li, M.; Liu, K.; Tang, D.; Sun, M.F.; Li, Y.F.; Shen, Y.; Du, G.J.; Cheng, Z.K. Semi-Rolled Leaf2 modulates rice leaf rolling by regulating abaxial side cell differentiation. J. Exp. Bot. 2016, 67, 2139–2150. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.H.; Xu, Q.; Zhu, X.D.; Qian, Q.; Xue, H.W. SHALLOT-LIKE1 is a KANADI transcription factor that modulates rice leaf rolling by regulating leaf abaxial cell development. Plant Cell 2009, 21, 719–735. [Google Scholar] [CrossRef]
- Yan, C.J.; Yan, S.; Zhang, Z.q.; Liang, G.H.; Lu, J.F.; Gu, M.H. Genetic analysis and gene fine mapping for a rice novel mutant (rl9(t)), with rolling leaf character. Chin. Sci. Bull. 2006, 51, 63–69. [Google Scholar] [CrossRef]
- Kong, X.Q.; Gao, X.H.; Sun, W.; An, J.; Zhao, Y.X.; Zhang, H. Cloning and functional characterization of a cation–chloride cotransporter gene OsCCC1. Plant Mol. Biol. 2011, 75, 567–578. [Google Scholar] [CrossRef]
- Tian, X.; Zhang, D.B. cDNA cloning and expression analysis of OsWOX4 in rice. J. Shanghai Jiaotong Univ. Agric. Sci. 2011, 29, 8–14. [Google Scholar]
- Cho, S.H.; Yoo, S.C.; Zhang, H.T.; Pandeya, D.; Koh, H.J.; Hwang, J.Y.; Kim, G.T.; Paek, N.C. The rice narrow leaf2 and narrow leaf3 loci encode WUSCHEL-related homeobox 3A OsWOX3A, and function in leaf, spikelet, tiller and lateral root development. New Phytol. 2013, 198, 1071–1084. [Google Scholar] [CrossRef] [PubMed]
- Luan, W.J.; Liu, Y.Q.; Zhang, F.X.; Song, Y.L.; Wang, Z.Y.; Peng, Y.K.; Sun, Z.X. OsCD1 encodes a putative member of the cellulose synthase-like D sub-family and is essential for rice plant architecture and growth. Plant Biotechnol. J. 2010, 9, 513–524. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Zhu, Li.; Zeng, D.L.; Gao, Z.Y.; Guo, L.B.; Fang, Y.X.; Zhang, G.H.; Dong, G.J.; Yan, M.X.; Liu, J.; Qian, Q. Identification and characterization of NARROW AND ROLLED LEAF 1, a novel gene regulating leaf morphology and plant architecture in rice. Plant Mol. Biol. 2010, 73, 283–292. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.H.; Li, S.Y.; Wang, L.; Ye, W.J.; Zeng, D.L.; Rao, Y.C.; Peng, Y.L.; Hu, J.; Yang, Y.L.; Xu, J.; et al. LSCHL4 from japonica cultivar, which is allelic to NAL1, increases yield of indica super rice 93-11. Mol. Plant 2014, 7, 1350–1364. [Google Scholar] [CrossRef] [PubMed]
- Fujino, K.; Matsuda, Y.; Ozawa, K.; Nishimura, T.; Koshiba, T.; Fraaije, M.W.; Sekiguchi, H. NARROW LEAF 7 controls leaf shape mediated by auxin in rice. Mol. Genet. Genom. 2008, 279, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Sazuka, T.; Kamiya, N.; Nishimura, T.; Ohmae, K.; Sato, Y.; Imamura, K.; Nagato, Y.; Koshiba, T.; Nagamura, Y.; Ashikari, M.; et al. A rice tryptophan deficient dwarf mutant, tdd1, contains a reduced level of indole acetic acid and develops abnormal flowers and organless embryos. Plant J. 2009, 60, 227–241. [Google Scholar] [CrossRef]
- Woo, Y.M.; Park, H.J.; Su’udi, M.; Yang, J.I.; Park, J.J.; Back, K.; Park, Y.M.; An, G. Constitutively wilted 1, a member of the rice YUCCA gene family, is required for maintaining water homeostasis and an appropriate root to shoot ratio. Plant Mol. Biol. 2007, 65, 125–136. [Google Scholar] [CrossRef]
- Zhang, S.Z.; Wu, T.; Liu, S.J.; Liu, X.; Jiang, L.; Wan, J.M. Disruption of OsARF19 is Critical for Floral Organ Development and Plant Architecture in Rice (Oryza sativa L.). Plant Mol. Biol. Rep. 2016, 34, 748–760. [Google Scholar] [CrossRef]
- Li, W.; Wu, C.; Hu, G.C.; Xing, Li.; Qian, W.J.; Si, H.M.; Sun, Z.X.; Wang, X.C.; Fu, Y.P.; Liu, W.Z. Characterization and fine mapping of a novel rice narrow leaf mutant nal9. J. Integr. Plant Biol. 2013, 55, 1016–1025. [Google Scholar] [CrossRef]
- Hibara, K.; Obara, M.; Hayashida, E.; Abe, M.; Ishimaru, T.; Satoh, H.; Itoh, J.; Nagato, Y. The ADAXIALIZED LEAF1 gene functions in leaf and embryonic pattern formation in rice. Dev. Biol. 2009, 334, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.Y.; Wang, J.; Wan, X.S.; Shen, G.Z.; Wang, X.; Zhang, J.L. Over-expression of rice OsAGO7 gene induces upward curling of the leaf blade that enhanced erect-leaf habit. Planta 2007, 226, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Xue, X.; Zou, S.M.; Chen, Z.X.; Zhang, Y.F.; Li, Q.Q.; Zhu, J.K.; Ma, Y.Y.; Pan, X.B.; Pan, C.H. Suppressed expressed of AGO1a leads to adaxial leaf rolling in rice. Chin. J. Rice Sci. 2013, 273, 223–230. [Google Scholar]
- Li, L.; Shi, Z.Y.; Li, L.; Shen, G.Z.; Wang, X.Q.; An, L.S.; Zhang, J.L. Overexpression of ACL1 abaxially curled leaf 1, increased bulliform cells and induced abaxial curling of leaf blades in rice. Mol. Plant 2010, 35, 807–817. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Hu, J.; Qian, Q.; Xue, H.W. LC2 and OsVIL2 promote rice flowering by photoperoid-induced epigenetic silencing of OsLF. Mol. Plant 2013, 62, 514–527. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.Q.; Hu, J.; Guo, L.B.; Qian, Q.; Xue, H.W. Rice leaf inclination 2, a VIN3-like protein, regulates leaf angle through modulating cell division of the collar. Cell Res. 2010, 20, 935–947. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Xiong, G.Y.; Li, R.; Cui, J.J.; Tang, D.; Zhang, B.C.; Pauly, M.; Cheng, Z.K.; Zhou, Y.H. Rice cellulose synthase-like D4 is essential for normal cell-wall biosynthesis and plant growth. Plant J. 2009, 60, 1055–1069. [Google Scholar] [CrossRef] [Green Version]
- Yoshikaw, T.; Eiguchi, M.; Hibara, K.I.; Ito, J.I.; Nagato, Y. Rice SLENDER LEAF 1 gene encodes cellulose synthase-like D4 and is specifically expressed in M-phase cells to regulate cell proliferation. J. Exp. Bot. 2013, 647, 2049–2061. [Google Scholar] [CrossRef]
- Ding, Z.Q.; Lin, Z.F.; Li, Q.; Wu, H.; Xiang, C.Y.; Wang, J.F. DNL1, encodes cellulose synthase-like D4, is a major QTL for plant height and leaf width in rice (Oryza sativa L.). Biochem. Biophys. Res. Commun. 2014, 457, 133–140. [Google Scholar] [CrossRef]
- Li, Y.Y.; Shen, A.; Xiong, W.; Sun, Q.L.; Luo, Q.; Song, T.; Li, Z.L.; Luan, W.J. Overexpression of OsHox32 results in pleiotropic effects on plant type architecture and leaf development in rice. Rice 2016, 9, 46. [Google Scholar] [CrossRef]
- Yang, C.H.; Li, D.Y.; Liu, X.; Ji, C.J.; Hao, L.L.; Zhao, X.F.; Li, X.B.; Chen, C.Y.; Cheng, Z.K.; Zhu, L.H. OsMYB103L, an R2R3-MYB transcription factor, influences leaf rolling and mechanical strength in rice (Oryza sativa L.). BMC Plant Biol. 2014, 14, 158. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Wang, Y.H.; Long, Q.Z.; Huang, J.X.; Wang, Y.L.; Zhou, K.N.; Zheng, M.; Sun, J.; Chen, H.; Chen, S.H.; et al. Overexpression of OsZHD1, a zinc finger homeodomain class homeobox transcription factor, induces abaxially curled and drooping leaf in rice. Planta 2009, 239, 803–816. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.L.; Xie, Q.J.; Gao, J.; Wang, W.Y.; Sun, B.; Liu, B.H.; Zhu, H.T.; Peng, H.F.; Zhao, H.B.; Liu, C.H.; et al. Characterization of Rolled and Erect Leaf 1 in regulating leave morphology in rice. J. Exp. Bot. 2015, 66, 6047–6058. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.Q.; Li, W.Q.; Miao, H.; Gan, P.F.; Qiao, L.; Chang, Y.L.; Shi, C.H.; Chen, K.M. REL2, A Gene Encoding An Unknown Function Protein which Contains DUF630 and DUF632 Domains Controls Leaf Rolling in Rice. Rice 2016, 9, 37. [Google Scholar] [CrossRef] [PubMed]
- Fang, L.K.; Zhao, F.M.; Cong, Y.F.; Sang, X.C.; Du, Q.; Wang, D.Z.; Li, Y.F.; Ling, Y.H.; Yang, Z.L.; He, G.H. Rolling-leaf14 is a 2OG-Fe II, oxygenase family protein that modulates rice leaf rolling by affecting secondary cellwall formation in leaves. Plant Biotechnol. J. 2012, 10, 524–532. [Google Scholar] [CrossRef] [PubMed]
- Zou, L.P.; Sun, X.H.; Zhang, Z.G.; Liu, P.; Wu, J.X.; Tian, C.J.; Qiu, J.L.; Lu, T.G. Leaf Rolling Controlled by the Homeodomain Leucine Zipper Class IV Gene Roc5 in Rice. Plant Physiol. 2011, 156, 1589–1602. [Google Scholar] [CrossRef] [PubMed]
- Li, W.Q.; Zhang, M.J.; Gan, P.F.; Qiao, L.; Yang, S.Q.; Miao, H.; Wang, G.F.; Zhang, M.M.; Liu, W.T.; Li, H.F.; et al. CLD1/SRL1 modulates leaf rolling by affecting cell wall formation, epidermis integrity and water homeostasis in rice. Plant J. 2017, 92, 904–923. [Google Scholar] [CrossRef]
- Xiang, J.J.; Zhang, G.H.; Qian, Q.; Xue, H.W. Semi-rolled leaf1 encodes a putative glycosylphosphatidylinositol-anchored protein and modulates rice leaf rolling by regulating the formation of bulliform cells. Plant Physiol. 2012, 159, 1488–1500. [Google Scholar] [CrossRef]
- Alamin, M.; Zeng, D.D.; Qin, R.; Sultana, H.; Jin, X.L.; Shi, C.H. Characterization and fine mapping of SFL1, a gene controlling Screw Flag Leaf in Rice. Plant Mol. Biol. Rep. 2017, 35, 491–503. [Google Scholar] [CrossRef]
- Zhang, J.J.; Wu, S.Y.; Jiang, L.; Wang, J.L.; Zhang, X.; Guo, X.P.; Wu, C.Y.; Wan, J.M. A detailed analysis of the leaf rolling mutant sll2 reveals complex nature in regulation of bulliform cell development in rice (Oryza sativa L.). Plant Biol. 2015, 17, 437–448. [Google Scholar] [CrossRef]
- Dai, M.Q.; Zhao, Y.; Ma, Q.; Hu, Y.F.; Hedden, P.; Zhang, Q.F.; Zhou, D.X. The rice YABBY1 gene is involved in the feedback regulation of gibberellin metabolism. Plant Physiol. 2007, 144, 121–133. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Wang, D.; Wu, T.; Yang, Y.; Liu, C.; Yan, L.; Tang, D.; Zhao, X.; Zhu, Y.; Lin, J.; et al. LRRK1, a receptor-like cytoplasmic kinase, regulates leaf rolling through modulating bulliform cell development in rice. Mol. Breed. 2018, 38, 48. [Google Scholar] [CrossRef]
- Ma, L.; Sang, X.C.; Zhang, T.; Yu, Z.Y.; Li, Y.F.; Zhao, F.M.; Wang, Z.W.; Wang, Y.T.; Yu, P.; Wang, N.; et al. ABNORMAL VASCULAR BUNDLES regulates cell proliferation and procambium cell establishment during aerial organ development in rice. New Phytol. 2017, 213, 275–286. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.F.; Wu, K.; Chen, J.F.; Liu, Q.; Wu, Y.J.; Liu, B.M.; Fu, X.D. OsSND2, a NAC family transcription factor, is involved in secondary cell wall biosynthesis through regulating MYBs expression in rice. Rice 2018, 11, 36. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.H.; Li, S.B.; He, S.; Waßmann, F.; Yu, C.; Qin, G.; Schreiber, L.; Qu, L.J.; Gu, H. CFL1, a WW domain protein, regulates cuticle development by modulating the function of HDG1, a class IV homeodomain transcription factor, in rice and Arabidopsis. Plant Cell 2011, 23, 3392–3411. [Google Scholar] [CrossRef] [PubMed]
- Jing, W.; Cao, C.J.; Shen, L.K.; Zhang, H.S.; Jing, G.Q.; Zhang, W.H. Characterization and fine mapping of a rice leaf-rolling mutant deficient in commissural veins. Crop Sci. 2017, 57, 2595–2604. [Google Scholar] [CrossRef]
- Li, S.G.; He, P.; Wang, Y.P.; Li, H.Y.; Chen, Y.; Zhou, K.D.; Zhu, L.H. Genetic analysis and gene mapping of the leaf traits in rice Oryza sativa L. Acta Agron. Sin. 2000, 26, 261–265. [Google Scholar]
- Yan, J.Q.; Zhu, J.; He, C.X.; Benmoussa, M.; Wu, P. Molecular marker-assisted dissection of genotype × environment interaction for plant type traits in rice (Oryza sativa L.). Crop Sci. 1999, 39, 538–544. [Google Scholar] [CrossRef]
- Farooq, M.; Tagle, A.G.; Santos, R.E.; Ebron, L.A.; Fujita, D.; Kobayashi, N. Quantitative trait loci mapping for leaf length and leaf width in rice cv. IR64 derived lines. J. Integr. Plant Biol. 2010, 52, 578–584. [Google Scholar] [CrossRef]
- Li, Z.K.; Pinson, S.R.M.; Stansel, J.W.; Paterson, A. Genetic dissection of the source-sink relationship affecting fecundity and yield in rice (Oryza sativa L.). Mol. Breed. 1998, 4, 419–426. [Google Scholar] [CrossRef]
- Mei, H.W.; Luo, L.J.; Ying, C.S.; Wang, Y.P.; Yu, X.Q.; Guo, L.B.; Paterson, A.H.; Li, Z.K. Gene actions of QTLs affecting several agronomic traits resolved in a recombinant inbred rice population and two testcross populations. Theor. Appl. Genet. 2005, 110, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Cui, K.H.; Peng, S.; Xing, Y.; Yu, S.; Xu, C.; Zhang, Q. Molecular dissection of the genetic relationships of source, sink and transport tissue with yield traits in rice. Theor. Appl. Genet. 2003, 106, 649–658. [Google Scholar] [CrossRef] [PubMed]
- Yue, B.; Xue, W.Y.; Luo, L.J.; Xing, Y.Z. QTL Analysis for flag leaf characteristics and their relationships with yield and yield traits in rice. Acta Agron. Sin. 2006, 33, 824–832. [Google Scholar] [CrossRef]
- Jiang, S.k.; Zhang, X.J.; Wang, J.Y.; Chen, W.F.; Xu, Z.J. Fine mapping of the quantitative trait locus qFLL9 controlling flag leaf length in rice. Euphytica 2010, 176, 341–347. [Google Scholar] [CrossRef]
- Wang, P.; Zhou, G.L.; Cui, K.H.; Li, Z.K.; Yu, S.B. Clustered QTL for source leaf size and yield traits in rice (Oryza sativa L.). Mol. Breed. 2012, 29, 99–113. [Google Scholar] [CrossRef]
- Wang, P.; Zhou, G.L.; Yu, H.H.; Yu, S.B. Fine mapping a major QTL for flag leaf size and yield-related traits in rice. Theor. Appl. Genet. 2011, 123, 1319–1330. [Google Scholar] [CrossRef]
- Ding, X.P.; Li, X.K.; Xiong, L.Z. Evaluation of near-isogenic lines for drought resistance QTL and fine mapping of a locus affecting flag leaf width, spikelet number, and root volume in rice. Theor. Appl. Genet. 2011, 123, 815–826. [Google Scholar] [CrossRef]
- Chen, M.L.; Luo, J.; Shao, G.N.; Wei, X.J.; Tang, S.Q.; Sheng, Z.H.; Song, J.; Hu, P.S. Fine mapping of a major QTL for flag leaf width in rice, qFLW4, which might be caused by alternative splicing of NAL1. Plant Cell Rep. 2012, 31, 863–872. [Google Scholar] [CrossRef]
- Fujita, D.; Trijatmiko, K.R.; Tagle, A.G.; Sapasap, M.V.; Koide, Y.; Sasaki, K.; Tsakirpaloglou, N.; Gannaban, R.B.; Nishimura, T.; Yanagihara, S.; et al. NAL1 allele from a rice landrace greatly increases yield in modern indica cultivars. Proc. Natl. Acad. Sci. USA 2013, 110, 20431–20436. [Google Scholar] [CrossRef]
- Hu, J.; Wang, Y.X.; Fang, Y.X.; Zeng, L.J.; Xu, J.; Yu, H.P.; Shi, Z.Y.; Pan, J.J.; Zhang, D.; Kang, S.J.; et al. A Rare allele of GS2 enhances grain size and grain yield in rice. Mol. Plant 2015, 8, 1455–1465. [Google Scholar] [CrossRef]
- Duan, P.; Ni, S.; Wang, J.M.; Zhang, B.L.; Xu, R.; Wang, Y.X.; Chen, H.Q.; Zhu, X.D.; Li, Y.H. Regulation of OsGRF4 by OsmiR396 controls grainsize and yield in rice. Nat. Plants 2015, 2, 15203. [Google Scholar] [CrossRef] [PubMed]
- Quarrie, S.A.; Pekic, S.; Radosevic, Q.; Rancic, R.D.; Kaminska, A.; Barnes, J.D.; Leverington, M.; Ceoloni, C.; Dodig, D. Dissecting a wheat QTL for yield present in a range of environments: From the QTL to candidate genes. J. Exp. Bot. 2006, 57, 2627–2637. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.Z.; Qian, Q.; Liu, Z.B.; Sun, H.Y.; He, S.Y.; Luo, D.; Xia, G.M.; Chu, C.C.; Li, J.Y.; Fu, X.D. Natural variation at the DEP1 locus enhances grain yield in rice. Nat. Genet. 2009, 41, 494–497. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.Y.; Nakazaki, T.; Chen, S.Q.; Chen, W.F.; Saito, H.; Tsukiyama, T.; Okumoto, Y.; Xu, Z.J.; Tanisaka, T. Identification and characterization of the erect-pose panicle gene EP conferring high grain yield in rice (Oryza sativa L.). Theor. Appl. Genet. 2009, 119, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhu, J.Y.; Li, Z.Y.; Yi, C.D.; Liu, J.; Zhang, H.G.; Tang, S.Z.; Gu, M.H.; Liang, G.H. Deletion in a quantitative trait gene qpe9-1 associated with panicle erectness improves plant architecture during rice domestication. Genetics 2009, 183, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Mao, S.; Lu, G.H.; Yu, K.H.; Bo, Z.; Chen, J.H. Specific protein detection using thermally reduced graphene oxide sheet decorated with gold nanoparticle-antibody conjugates. Adv. Mater 2010, 22, 3521–3526. [Google Scholar] [CrossRef] [PubMed]
- Kosugi, S.; Suzuka, I.; Ohashi, Y. Two of three promoter elements identified in a rice gene for proliferating cell nuclear antigen are essential for meristematic tissue-specific expression. Plant J. 1995, 7, 877–886. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.Z.; Sun, J.; Xiao, Z.Q.; Cheng, F.; Xu, H.; Tang, L.; Chen, W.F.; Xu, Z.J.; Xu, Q. Variations in DENSE AND ERECT PANICLE 1 (DEP1) contribute to the diversity of the panicle trait in high-yielding japonica rice varieties in northern China. Breed. Sci. 2016, 66, 599–605. [Google Scholar] [CrossRef]
- Narita, N.N.; Moore, S.; Horiguchi, G.; Kubo, M.; Demura, T.; Fukuda, H.; Goodrich, J.; Tsukaya, H. Overexpression of a novel small peptide ROTUNDIFOLIA4 decreases cell proliferation and alters leaf shape in Arabidopsis thaliana. Plant J. 2004, 38, 399–713. [Google Scholar] [CrossRef]
- Tsuge, T.; Tsukaya, H.; Uchimiya, H. Two independent and polarized processes of cell elongation regulate leaf blade expansion in Arabidopsis thaliana (L.) Heynh. Development 1996, 122, 1589–1600. [Google Scholar]
- Xu, J.; Wang, L.; Wang, Y.X.; Zeng, D.L.; Zhou, M.Y.; Fu, X.; Ye, W.J.; Hu, J.; Zhu, L.; Ren, D.Y.; et al. Reduction of OsFLW7 expression enhanced leaf area and grain production in rice. Sci. Bull. 2017, 62, 1631–1633. [Google Scholar] [CrossRef]
- Snyder, M.; He, W.; Zhang, J.J. The DNA replication factor MCM5 is essential for stat1-mediated transcriptional activation. Proc. Natl. Acad. Sci. USA 2005, 102, 14539–14544. [Google Scholar] [CrossRef] [PubMed]
- Imai, K.K.; Ohashi, Y.H.; Tsuge, T.; Yoshizumi, T.; Matsui, M.; Oka, A.; Aoyama, T. The a-type cyclin CYCA2;3 is a key regulator of ploidy levels in Arabidopsis endoreduplication. Plant Cell 2006, 18, 382–396. [Google Scholar] [CrossRef] [PubMed]
- Boudolf, V.; Lammens, T.; Boruc, J.; Leene, J.V.; Daele, H.V.D.; Maes, S.; Isterdael, G.V.; Russinova, E.; Kondorosi, E.; Witters, E.; et al. CDKB1;1 Forms a functional complex with CYCA2;3 to suppress endocycle onset. Plant Physiol. 2009, 150, 1482–1493. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.Y.; Song, X.W.; Xu, J.; Fu, X.; Li, T.; Zhu, Y.C.; Xiao, X.Y.; Mao, Y.J.; Zeng, D.L.; Hu, J.; et al. Construction of genetic map and mapping and verification of grain traits QTLs using recombinant inbred lines derived from a cross between indica C84 and japonica CJ16B. Chin. J. Rice Sci. 2018, 32, 207–218. [Google Scholar]
- Shen, L.; Hua, Y.F.; Fu, Y.P.; Li, J.; Liu, Q.; Jiao, X.Z.; Xin, G.W.; Wang, J.J.; Wang, X.C.; Yan, C.J.; et al. Rapid generation of genetic diversity by multiplex CRISPR/Cas9 genome editing in rice. Sci. China Life Sci. 2017, 60, 506–515. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.L.; Zhang, Q.Y.; Zhu, Q.L.; Liu, W.; Chen, Y.; Qiu, R.; Wang, B.; Yang, Z.F.; Li, H.Y.; Lin, Y.R.; et al. A robust CRISPR/Cas9 system for convenient, high-efficiency multiplex genome editing in monocot and dicot plants. Mol. Plant 2015, 8, 1274–1284. [Google Scholar] [CrossRef]
- Hellens, R.P.; Allan, A.C.; Friel, E.N.; Bolitho, K.; Grafton, K.; Templeton, M.D.; Karunairetnam, S.; Gleave, A.P.; Laing, W.A. Transient expression vectors for functional genomics, quantification of promoter activity and RNA silencing in plants. Plant Methods 2005, 1, 13. [Google Scholar] [CrossRef]
- You, M.K.; Lim, S.H.; Kim, M.J.; Jeong, Y.S.; Lee, M.G.; Ha, S.H. Improvement of the fluorescence intensity during a flow cytometric analysis for rice protoplasts by localization of a green fluorescent protein into chloroplasts. Int. J. Mol. Sci. 2015, 16, 788–804. [Google Scholar] [CrossRef]
- Ishimaru, K.; Shirota, K.; Higa, M.; Kawamitsu, Y. Identification of quantitative trait loci for adaxial and abaxial stomatal frequencies in Oryza sativa. Plant Physiol. Biochem. 2001, 39, 173–177. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HZ | HN | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| FLL | FLW | SLL | SLW | TLL | TLW | FLL | FLW | SLL | SLW | TLL | |
| HZ-FLL | |||||||||||
| HZ-FLW | −0.078 | ||||||||||
| HZ-SLL | 0.833 ** | −0.013 | |||||||||
| HZ-SLW | −0.178 * | 0.627 ** | −0.072 | ||||||||
| HZ-TLL | 0.525 ** | 0.000 | 0.746 ** | −0.023 | |||||||
| HZ-TLW | −0.203 ** | 0.597 ** | −0.073 | 0.699 ** | 0.086 | ||||||
| HN-FLL | 0.341 ** | −0.317 ** | 0.271 ** | −0.202 * | 0.336 ** | −0.252 ** | |||||
| HN-FLW | −0.250 ** | 0.389 ** | −0.298 ** | 0.382 ** | −0.287 ** | 0.246 ** | 0.076 | ||||
| HN-SLL | 0.225 ** | −0.268 ** | 0.135 | −0.137 | 0.195 * | −0.226 ** | 0.873 ** | 0.229 ** | |||
| HN-SLW | −0.418 ** | 0.390 ** | −0.510 ** | 0.441 ** | −0.395 ** | 0.361 ** | −0.021 | 0.754 ** | 0.160 | ||
| HN-TLL | 0.000 | −0.182 * | -0.098 | −0.087 | 0.007 | −0.132 | 0.642 ** | 0.254 ** | 0.813 ** | 0.313 ** | |
| HN-TLW | −0.405 ** | 0.246 ** | −0.477 ** | 0.337 ** | −0.355 ** | 0.300 ** | 0.088 | 0.624 ** | 0.206 * | 0.760 ** | 0.358 ** |
| Trait | QTL | Interval | Peak Position | Additive Effect | Explained Phenotypic Variance (%) | ||||
|---|---|---|---|---|---|---|---|---|---|
| Marker 1 | Marker 2 | HZ | HN | HZ | HN | HZ | HN | ||
| FLL | qFLL9 | RM3700 | B9-11 | 61.1 | 56.87 | −7.1707 | −6.8682 | 32.41 | 30.63 |
| qFLL10 | RM1375 | H10-3 | 20.09 | 3.3926 | 5.09 | ||||
| FLW | qFLW3 | H3-11 | H3-12 | 152.3 | 0.0648 | 6.6 | |||
| qFLW6 | RM3496 | H6-3 | 27.14 | −0.0839 | 16.07 | ||||
| SLL | qSLL2-1 | 2-30.2-a | 2-42.1-a | 36.2 | −2.8643 | 8.98 | |||
| qSLL2-2 | H2-1 | 2-30.2-a | 23.9 | −1.3914 | 5.29 | ||||
| qSLL6 | wgw1 | RM3183 | 59.4 | 60.1 | 3.5234 | −4.2842 | 14.97 | 17.29 | |
| qSLL9 | RM3700 | B9-11 | 60.1 | 57.6 | −6.4659 | −6.5431 | 29.62 | 20.85 | |
| qSLL9-2 | H9-3 | H9-4 | 12.18 | −0.7557 | 8.8 | ||||
| qSLL10 | RM1375 | H10-3 | 27.11 | 3.9058 | 5.91 | ||||
| SLW | qSLW1 | RM1247 | H1-1 | 24.2 | −0.0759 | 10.69 | |||
| qSLW6 | wgw1 | RM3183 | 59.35 | −0.0727 | 10.31 | ||||
| qSLW9 | RM3700 | B9-11 | 59.2 | 0.0839 | 18.39 | ||||
| qSLW11 | H11-9 | H11-10 | 71.4 | −0.0329 | 1.85 | ||||
| TLL | qTLL2 | H2-1 | 2-30.2-a | 13.0 | 19.44 | −3.6555 | −0.9935 | 13.10 | 8.99 |
| qTLL5 | RM430 | RM18751 | 70.3 | −2.5759 | 8.43 | ||||
| qTLL6 | wgw1 | RM3183 | 35.2 | −4.8389 | 15.65 | ||||
| qTLL9 | RM3700 | B9-11 | 59.1 | 61.2 | −6.0826 | −4.8326 | 21.87 | 19.19 | |
| qTLL10 | RM5689 | RM1375 | 28.6 | 2.3523 | 7.76 | ||||
| TLW | qTLW1 | H1-1 | RM8111 | 22.41 | −0.0669 | 10.79 | |||
| qTLW6 | wgw1 | RM3183 | 59.35 | −0.0684 | 13.84 | ||||
| qTLW9 | RM3700 | B9-11 | 61.31 | 0.0834 | 16.28 | ||||
| Primer | Forward (5′-3′) | Reverse (5′-3′) | Experiment |
|---|---|---|---|
| RM3700 | AAATGCCCCATGCACAAC | TTGTCAGATTGTCACCAGGG | Fine mapping |
| C9-1594 | CCTGTACACTGTAGGCCTGT | GGTGTCAAAGTACATAGGCCC | |
| C9-1635 | GGTGGAAAGGAAGGAGAGCT | CTAGCCCTGCCTCGTTGTAA | |
| C9-1638 | GTGTGTGTGTGTGTGTGTGT | TCATAGTACATGCCCTCCGT | |
| C9-1640 | ATAAGTCCATATTGCCCACCTC | AAGCTTCTGGATCGTTAACAGG | |
| C9-1642 | GTACCCTCCTCCGATGACAC | TTGTGGAGGACGAGAAGGTG | |
| C9-1715 | GGTGGCGAGAAGAATTTGCA | TTTCGCCTCTCACTGACCTT | |
| B9-11 | TCTTACGAATAGGCCCTTGG | AGAGCCCACAACACTTGTGC | |
| Actin | ATCCATCTTGGCATCTCTCAGC | CACAATGGATGGGCCAGACT | qRT-PCR |
| LOC_Os09g26960 | CTGAGCCTCGCCAATCTG | CGAAGATCTCCTCCATGCTC | |
| LOC_Os09g26970 | CAAACATCTGGGCTTGGTCT | TCTAAGCAACCTGCCCAATC | |
| LOC_Os09g26980 | ATTGATGTGAAAGGGCAAGACT | CACCTTAAGCCCAAGGTTGTAG | |
| LOC_Os09g26999 | GTAGCTGCAAGCCAAGCTG | TTGAAGCAGCTGGAGCAAC | |
| POs26999 | GGCCAGTGCCAAGCTTAAGGGAAGTTGGCCGCCTGCC | AGGGTCTTGCAGATCTCTCCACACGCAGCACGCCAAC | Vector construction |
| Os09g26999-g++/g-- | GGCAGGTGGTGATGGAGGCGCCG | AAACCGGCGCCTCCATCACCACC | |
| Os09g26999-JC | CGGCGATTTATACCCACCAC | CGCTCACCTTGAGGAACGT | Detection of target mutations |
| Hyg-F1 | GCTGTTATGCGGCCATTGTC | GACGTCTGTCGAGAAGTTTC | |
| Cas9-F2/pC1300-R2 | ACCAGACACGAGACGACTAA | ATCGGTGCGGGCCTCTTC | |
| T3 | ATCGGTGCGGGCCTCTTC |
| Gene ID | Annotation from the Rice Genome Annotation Project |
|---|---|
| LOC_Os09g26970 | Retrotransposon protein, Putative, Unclassified, Expressed |
| LOC_Os09g26980 | Cytochrome P450, Putative, Expressed |
| LOC_Os09g26999 | Gγ subunit; Dense and Erect Panicle1; DENSE PANICLE 1 |
| Locus Name | Position on chr.9 | Position on gene | CJ16 | C84 | CJ16 | C84 |
|---|---|---|---|---|---|---|
| LOC_Os09g26970 | 16393266 | 397 | G | C | Val | Leu |
| LOC_Os09g26970 | 16393717 | 848 | T | C | Val | Ala |
| LOC_Os09g26970 | 16395923 | 3054 | T | C | Val | Ala |
| LOC_Os09g26980 | 16403850 | 4146 | G | A | Ala | Val |
| LOC_Os09g26980 | 16404094 | 3902 | T | C | Met | Cys |
| LOC_Os09g26999 | 16414735 | 3182 | A | G | Tyr | Cys |
| Position | CJ16 | C84 |
|---|---|---|
| −1341 | A | C |
| −1255 | G | C |
| −951 | C | G |
| −906 | T | G |
| −629 | G | T |
| −503 | A | C |
| −493 | G | C |
| −486 | T | G |
| −31 | TG | T |
| Trait | NIL-qLL9CJ16 | NIL-qLL9C84 |
|---|---|---|
| Panicle length (cm) | 20.44 ± 0.84 | 26.7 ± 0.83 ** |
| Panicles per plant | 9.25 ± 1.42 | 9.42 ± 1.26 |
| Number of primary branches | 22.11 ± 2.18 | 21.4 ± 2.42 |
| Number of secondry branches | 80.00 ± 15.51 | 77.60 ± 7.84 |
| Grains per panicle | 404.11 ± 71.70 | 355.17 ± 34.09 |
| 1000-grain weight (g) | 20.79 ± 0.74 | 22.99 ± 0.45 ** |
| Seed setting rate (%) | 75.80 ± 0.02 | 69.71 ± 0.02 ** |
| Yield per plant (g) | 27.93 ± 4.41 | 32.56 ± 8.19 |
| Actual yield per plot (kg/48 m2) | 31.52 ± 3.20 | 36.28 ± 3.27 * |
| Actual yield change (%) | – | 15.08 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, X.; Xu, J.; Zhou, M.; Chen, M.; Shen, L.; Li, T.; Zhu, Y.; Wang, J.; Hu, J.; Zhu, L.; et al. Enhanced Expression of QTL qLL9/DEP1 Facilitates the Improvement of Leaf Morphology and Grain Yield in Rice. Int. J. Mol. Sci. 2019, 20, 866. https://doi.org/10.3390/ijms20040866
Fu X, Xu J, Zhou M, Chen M, Shen L, Li T, Zhu Y, Wang J, Hu J, Zhu L, et al. Enhanced Expression of QTL qLL9/DEP1 Facilitates the Improvement of Leaf Morphology and Grain Yield in Rice. International Journal of Molecular Sciences. 2019; 20(4):866. https://doi.org/10.3390/ijms20040866
Chicago/Turabian StyleFu, Xue, Jing Xu, Mengyu Zhou, Minmin Chen, Lan Shen, Ting Li, Yuchen Zhu, Jiajia Wang, Jiang Hu, Li Zhu, and et al. 2019. "Enhanced Expression of QTL qLL9/DEP1 Facilitates the Improvement of Leaf Morphology and Grain Yield in Rice" International Journal of Molecular Sciences 20, no. 4: 866. https://doi.org/10.3390/ijms20040866
APA StyleFu, X., Xu, J., Zhou, M., Chen, M., Shen, L., Li, T., Zhu, Y., Wang, J., Hu, J., Zhu, L., Gao, Z., Dong, G., Guo, L., Ren, D., Chen, G., Lin, J., Qian, Q., & Zhang, G. (2019). Enhanced Expression of QTL qLL9/DEP1 Facilitates the Improvement of Leaf Morphology and Grain Yield in Rice. International Journal of Molecular Sciences, 20(4), 866. https://doi.org/10.3390/ijms20040866