Comprehensive Comparative Analysis of Cholesterol Catabolic Genes/Proteins in Mycobacterial Species

 , and
, and 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Deducing Cholesterol Catabolic Pathway in M. Tuberculosis H37Rv
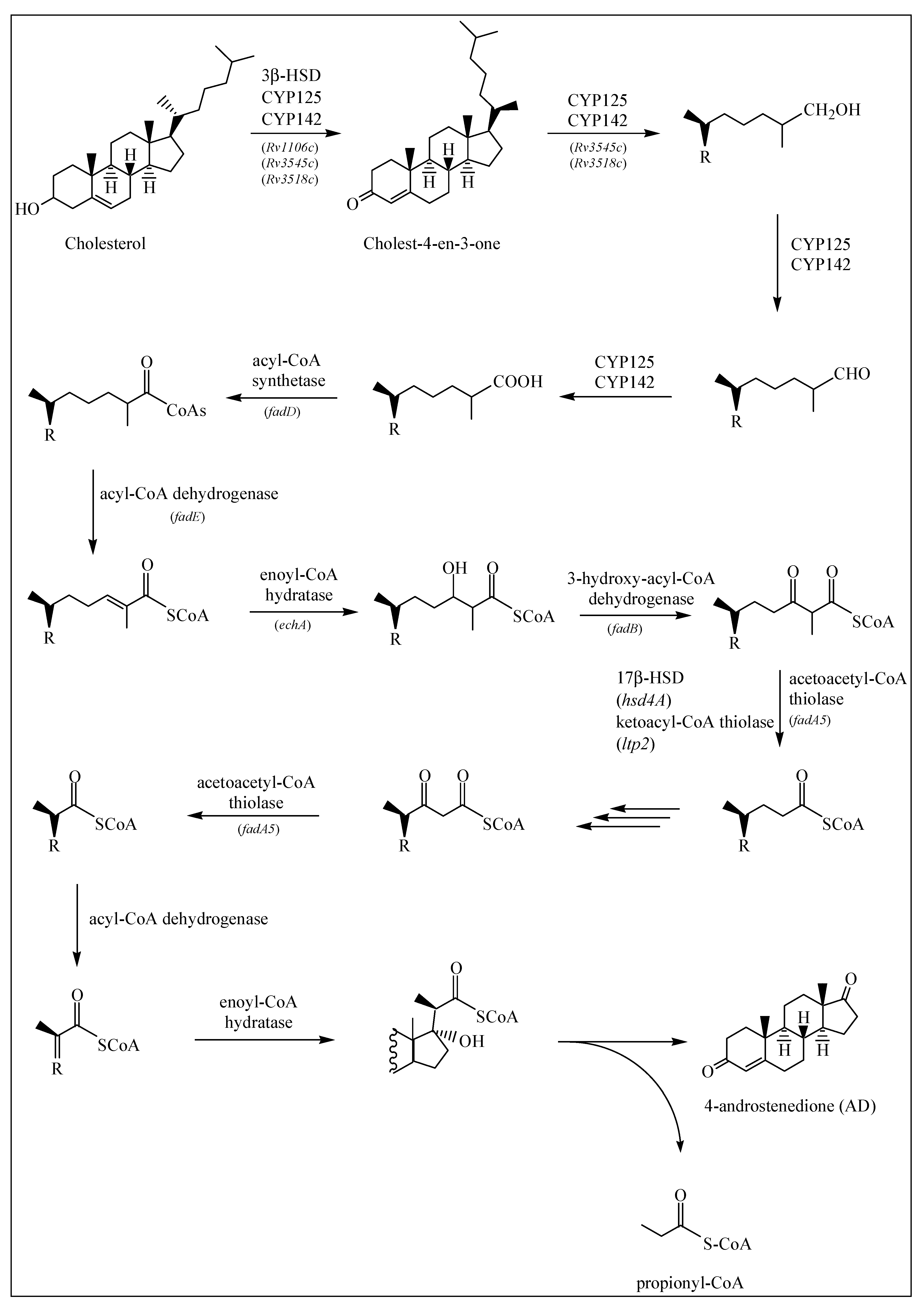
2.1.1. Degradation of Cholesterol: Side Chain Degradation
2.1.2. Degradation of Cholesterol: Sterol Ring Degradation
2.2. Genes/Proteins Involved in Cholesterol Catabolism in M. Tuberculosis H37Rv
2.2.1. Genes Predicted to be Specifically Required for Growth on Cholesterol
2.2.2. Cholesterol Catabolic Genes Proven to be or Predicted to be Essential for Survival of M. Tuberculosis in Macrophage Cells and in Murine Infection
2.2.3. Genes/Proteins that are Up-Regulated during Growth on Cholesterol
2.2.4. Genes Involved in Cholesterol Catabolism by M. Tuberculosis H37Rv, but Not Confirmed or Predicted to Be Essential
2.3. Key Cholesterol Catabolic Genes/Proteins are Not Found in a Large Number of Mycobacterial Species
2.3.1. Most of the M. Tuberculosis Complex Species Have the Ability to Catabolize Cholesterol
2.3.2. M. Chelonae-Abscessus Complex Species Lack Key Cholesterol Catabolic Genes/Proteins
2.3.3. Most of the M. Avium Complex Species Have the Ability to Catabolize Cholesterol
2.3.4. Mycobacterium Causing Leprosy Species Does Not Have the Ability to Catabolize Cholesterol
2.3.5. Uncertainty about Non-Tuberculosis Mycobacterium and Saprophyte Species’ Ability to Utilize Cholesterol
3. Materials and Methods
3.1. Species and Database
3.2. Cholesterol Catabolism
3.3. Cholesterol Catabolic Genes/Proteins Analysis in Mycobacterial Species
3.3.1. BLAST Analysis
3.3.2. Excel Program for Extracting KEGG BLAST Data
3.3.3. Data Collection and Protein Domain/Function Analysis
3.3.4. Assessing the Presence or Absence of Cholesterol Catabolic Gene/Protein Homologs in Mycobacterial Species
3.4. Generation of Gene/Protein Heatmaps
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- World Health Organization (WHO). Available online: https://www.who.int/tb/publications/global_report/en/ (accessed on 15 January 2019).
- Sotgiu, G.; Sulis, G.; Matteelli, A. Tuberculosis—A World Health Organization Perspective. Microbiol. Spectr. 2017, 5, 1–2. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Available online: http://www.who.int/tb/publications/global_report/gtbr15_main_text.pdf. (accessed on 15 January 2019).
- Glickman, M.S.; Jacobs, W.R. Microbial pathogenesis of Mycobacterium tuberculosis: Dawn of a discipline. Cell 2001, 104, 477–485. [Google Scholar] [CrossRef]
- Flynn, J.L.; Chan, J. Tuberculosis: Latency and reactivation. Infect. Immun. 2001, 69, 4195–4201. [Google Scholar] [CrossRef] [PubMed]
- Migliori, G.B.; Centis, R.; D’Ambrosio, L.; Spanevello, A.; Borroni, E.; Cirillo, D.M.; Sotgiu, G. Totally drug-resistant and extremely drug-resistant tuberculosis: The same disease? Clin. Infect. Dis. 2012, 54, 379–1380. [Google Scholar] [CrossRef] [PubMed]
- Clark-Curtiss, J.E.; Haydel, S.E. Molecular genetics of Mycobacterium tuberculosis pathogenesis. Annu. Rev. Microbiol. 2003, 57, 517–549. [Google Scholar] [CrossRef] [PubMed]
- Koul, A.; Herget, T.; Klebl, B.; Ullrich, A. Interplay between mycobacteria and host signalling pathways. Nat. Rev. Microbiol. 2004, 2, 189–202. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Wainwright, H.C.; Locketz, M.; Bekker, L.G.; Walther, G.B.; Dittrich, C.; Visser, A.; Wang, W.; Hsu, F.F.; Wiehart, U.; et al. Caseation of human tuberculosis granulomas correlates with elevated host lipid metabolism. EMBO Mol. Med. 2010, 2, 258–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schäfer, G.; Guler, R.; Murray, G.; Brombacher, F.; Brown, G.D. The role of scavenger receptor B1 in infection with Mycobacterium tuberculosis in a murine model. PLoS ONE 2009, 4, e8448. [Google Scholar] [CrossRef] [PubMed]
- Martens, G.W.; Arikan, M.C.; Lee, J.; Ren, F.; Vallerskog, T.; Kornfeld, H. Hypercholesterolemia impairs immunity to tuberculosis. Infect. Immun. 2008, 76, 3464–3472. [Google Scholar] [CrossRef] [PubMed]
- Gatfield, J.; Pieters, J. Essential role for cholesterol in entry of mycobacteria into macrophages. Science 2000, 288, 1647–1651. [Google Scholar] [CrossRef] [PubMed]
- Peyron, P.; Bordier, C.; Elsa-Noah, N.; Maridonneau-Parini, I. Nonopsonic phagocytosis of Mycobacterium kansasii by human neutrophils depends on cholesterol and is mediated by CR3 associated with glycosylphosphatidylinositol-anchored proteins. J. Immunol. 2000, 165, 5186–5191. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, S.; Rivas-Santiago, B.; Enciso, J.A. Mycobacterium tuberculosis entry into mast cells through cholesterol-rich membrane microdomains. Scand. J. Immunol. 2009, 70, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Pieters, J.; Gatfield, J. Hijacking the host; survival of pathogenic mycobacteria inside macrophages. Trends Microbiol. 2002, 10, 142–146. [Google Scholar] [CrossRef]
- Nguyen, L.; Pieters, J. The Trojan horse: Survival tactics of pathogenic mycobacteria in macrophages. Trends Cell Biol. 2005, 15, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.K.; Sassetti, C.M. Mycobacterial persistence requires the utilization of host cholesterol. Proc. Natl. Acad. Sci. USA 2008, 105, 4376–4380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Chastellier, C.; Thilo, L. Cholesterol depletion in Mycobacterium avium-infected macrophages overcomes the block in phagosome maturation and leads to the reversible sequestration of viable mycobacteria in phagolysosome-derived autophagic vacuoles. Cell. Microbiol. 2006, 8, 242–256. [Google Scholar] [CrossRef] [PubMed]
- Ouellet, H.; Johnston, J.B.; de Montellano, P.R.O. Cholesterol catabolism as a therapeutic target in Mycobacterium tuberculosis. Trends Microbiol. 2011, 19, 530–539. [Google Scholar] [CrossRef] [PubMed]
- García, J.L.; Uhía, I.; Galán, B. Catabolism and biotechnological applications of cholesterol degrading bacteria. Microb. Biotechnol. 2012, 5, 679–699. [Google Scholar] [CrossRef] [PubMed]
- Dubnau, E.; Chan, J.; Mohan, V.P.; Smith, I. Responses of Mycobacterium tuberculosis to growth in the mouse lung. Infect. Immun. 2005, 73, 3754–3757. [Google Scholar] [CrossRef] [PubMed]
- Nesbitt, N.M.; Yang, X.; Fontán, P.; Kolesnikova, I.; Smith, I.; Sampson, N.S.; Dubnau, E. A thiolase of Mycobacterium tuberculosis is required for virulence and production of androstenedione and androstadienedione from cholesterol. Infect. Immun. 2010, 78, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.T.; VanderVen, B.C.; Sherman, D.R.; Russell, D.G.; Sampson, N.S. Pathway profiling in Mycobacterium tuberculosis elucidation of cholesterol-derived catabolite and enzymes that catalyze its metabolism. J. Biol. Chem. 2011, 286, 43668–43678. [Google Scholar] [CrossRef] [PubMed]
- Świzdor, A.; Panek, A.; Milecka-Tronina, N.; Kołek, T. Biotransformations utilizing β-oxidation cycle reactions in the synthesis of natural compounds and medicines. Int. J. Mol. Sci. 2012, 13, 16514–16543. [Google Scholar] [CrossRef] [PubMed]
- Van der Geize, R.; Yam, K.; Heuser, T.; Wilbrink, M.H.; Hara, H.; Anderton, M.C.; Sim, E.; Dijkhuizen, L.; Davies, J.E.; Mohn, W.W.; et al. A gene cluster encoding cholesterol catabolism in a soil actinomycete provides insight into Mycobacterium tuberculosis survival in macrophages. Proc. Natl. Acad. Sci. USA 2007, 104, 1947–1952. [Google Scholar] [CrossRef] [PubMed]
- Griffin, J.E.; Gawronski, J.D.; DeJesus, M.A.; Loerger, T.R.; Akerley, B.J.; Sassetti, C.M. High-resolution phenotypic profiling defines genes essential for mycobacterial growth and cholesterol catabolism. PLoS Pathol. 2011, 7, e1002251. [Google Scholar] [CrossRef] [PubMed]
- Crowe, A.M.; Casabon, I.; Brown, K.L.; Liu, J.; Lian, J.; Rogalski, J.C.; Hurst, T.E.; Snieckus, V.; Foster, L.J.; Eltis, L.D. Catabolism of the last two steroid rings in Mycobacterium tuberculosis and other bacteria. mBio 2017, 8, e00321-17. [Google Scholar] [CrossRef] [PubMed]
- Rosłoniec, K.Z.; Wilbrink, M.H.; Capyk, J.K.; Mohn, W.W.; Ostendorf, M.; Van Der Geize, R.; Dijkhuizen, L.; Eltis, L.D. Cytochrome P450 125 (CYP125) catalyses C26-hydroxylation to initiate sterol side-chain degradation in Rhodococcus jostii RHA1. Mol. Microbiol. 2009, 74, 1031–1043. [Google Scholar] [CrossRef] [PubMed]
- Johnston, J.B.; Ouellet, H.; de Montellano, P.R.O. Functional redundancy of steroid C26-monooxygenase activity in Mycobacterium tuberculosis revealed by biochemical and genetic analyses. J. Biol. Chem. 2010, 285, 36352–36360. [Google Scholar] [CrossRef] [PubMed]
- García-Fernández, E.; Frank, D.J.; Galán, B.; Kells, P.M.; Podust, L.M.; García, J.L.; Ortiz de Montellano, P.R. A highly conserved mycobacterial cholesterol catabolic pathway. Environ. Microbiol. 2013, 15, 2342–2359. [Google Scholar] [CrossRef] [PubMed]
- Wipperman, M.F.; Yang, M.; Thomas, S.T.; Sampson, N.S. Shrinking the FadE proteome of Mycobacterium tuberculosis: Insights into cholesterol metabolism through identification of an α2β2 heterotetrameric acyl coenzyme A dehydrogenase family. J. Bacteriol. 2013, 195, 4331–4341. [Google Scholar] [CrossRef] [PubMed]
- Schnappinger, D.; Ehrt, S.; Voskuil, M.I.; Liu, Y.; Mangan, J.A.; Monahan, I.M.; Dolganov, G.; Efron, B.; Butcher, P.D.; Nathan, C.; et al. Transcriptional adaptation of Mycobacterium tuberculosis within macrophages. J. Exp. Med. 2003, 198, 693–704. [Google Scholar] [CrossRef] [PubMed]
- Brzostek, A.; Dziadek, B.; Rumijowska-Galewicz, A.; Pawelczyk, J.; Dziadek, J. Cholesterol oxidase is required for virulence of Mycobacterium tuberculosis. FEMS Microbiol. Lett. 2007, 275, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Gao, J.; Smith, I.; Dubnau, E.; Sampson, N.S. Cholesterol is not an essential source of nutrition for Mycobacterium tuberculosis during infection. J. Bacteriol. 2011, 193, 1473–1476. [Google Scholar] [CrossRef] [PubMed]
- Brzostek, A.; Pawelczyk, J.; Rumijowska-Galewicz, A.; Dziadek, B.; Dziadek, J. Mycobacterium tuberculosis is able to accumulate and utilize cholesterol. J. Bacteriol. 2009, 191, 6584–6591. [Google Scholar] [CrossRef] [PubMed]
- Petrusma, M.; van der Geize, R.; Dijkhuizen, L. 3-Ketosteroid 9α-hydroxylase enzymes: Rieske non-heme monooxygenases essential for bacterial steroid degradation. Antonie Leeuwenhoek 2014, 106, 157–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.; Van Der Geize, R.; Besra, G.S.; Gurcha, S.S.; Liu, A.; Rohde, M.; Singh, M.; Coates, A. 3-ketosteroid 9α-hydroxylase is an essential factor in the pathogenesis of Mycobacterium tuberculosis. Mol. Microbiol. 2010, 75, 107–121. [Google Scholar] [CrossRef] [PubMed]
- Rengarajan, J.; Bloom, B.R.; Rubin, E.J. Genome-wide requirements for Mycobacterium tuberculosis adaptation and survival in macrophages. Proc. Natl. Acad. Sci. USA 2005, 102, 8327–8332. [Google Scholar] [CrossRef] [PubMed]
- Dresen, C.; Lin, L.Y.C.; D’Angelo, I.; Tocheva, E.I.; Strynadka, N.; Eltis, L.D. A flavin-dependent monooxygenase from Mycobacterium tuberculosis involved in cholesterol catabolism. J. Biol. Chem. 2010, 285, 22264–22275. [Google Scholar] [CrossRef] [PubMed]
- Lack, N.A.; Yam, K.C.; Lowe, E.D.; Horsman, G.P.; Owen, R.L.; Sim, E.; Eltis, L.D. Characterization of a carbon-carbon hydrolase from Mycobacterium tuberculosis involved in cholesterol metabolism. J. Biol. Chem. 2010, 285, 434–443. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, S.; Nair, S.; Ghosh, S. Pathogenesis in tuberculosis: Transcriptomic approaches to unraveling virulence mechanisms and finding new drug targets. FEMS Microbiol. Rev. 2012, 36, 463–485. [Google Scholar] [CrossRef] [PubMed]
- Van der Geize, R.; Grommen, A.W.F.; Hessels, G.I.; Jacobs, A.A.C.; Dijkhuizen, L. The steroid catabolic pathway of the intracellular pathogen Rhodococcus equi is important for pathogenesis and a target for vaccine development. PLoS Pathol. 2011, 7, e1002181. [Google Scholar] [CrossRef] [PubMed]
- Driscoll, M.D.; McLean, K.J.; Levy, C.; Mast, N.; Pikuleva, I.A.; Lafite, P.; Rigby, S.E.; Leys, D.; Munro, A.W. Structural and biochemical characterization of Mycobacterium tuberculosis CYP142 evidence for multiple cholesterol 27-hydroxylase activities in a human pathogen. J. Biol. Chem. 2010, 285, 38270–38282. [Google Scholar] [CrossRef] [PubMed]
- Marques, M.A.M.; Berrêdo-Pinho, M.; Rosa, T.L.; Pujari, V.; Lemes, R.M.; Lery, L.M.; Silva, C.A.M.; Guimarães, A.C.R.; Atella, G.C.; Wheat, W.H.; et al. The essential role of cholesterol metabolism in the intracellular survival of Mycobacterium leprae is not coupled to central carbon metabolism and energy production. J. Bacteriol. 2015, 197, 3698–3707. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.; Zhang, T.; Li, B.; Wang, Z.; Ju, F.; Liang, Y.-T. Mycobacterial species and their contribution to cholesterol degradation in wastewater treatment plants. Sci. Rep 2019, 9, 836. [Google Scholar] [CrossRef] [PubMed]
- Ventura, M.; Canchaya, C.; Tauch, A.; Chandra, G.; Fitzgerald, G.F.; Chater, K.F.; van Sinderen, D. Genomics of Actinobacteria: Tracing the evolutionary history of an ancient phylum. Microbiol. Mol. Biol. Rev. 2007, 71, 495–548. [Google Scholar] [CrossRef] [PubMed]
- Parvez, M.; Qhanya, L.B.; Mthakathi, N.T.; Kgosiemang, I.K.R.; Bamal, H.D.; Pagadala, N.S.; Xie, T.; Yang, H.; Chen, H.; Theron, C.W.; et al. Molecular evolutionary dynamics of cytochrome P450 monooxygenases across kingdoms: Special focus on mycobacterial P450s. Sci. Rep. 2016, 6, 33099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tortoli, E. Phylogeny of the genus Mycobacterium: Many doubts, few certainties. Infect. Genet. Evol. 2012, 12, 827–831. [Google Scholar] [CrossRef] [PubMed]
- Mills, N. ChemDraw Ultra 10.0. J. Am. Chem. Soc. 2006, 128, 13649–13650. [Google Scholar] [CrossRef]
- Marchler-Bauer, A.; Bo, Y.; Han, L.; He, J.; Lanczycki, C.J.; Lu, S.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; et al. CDD/SPARCLE: Functional classification of proteins via subfamily domain architectures. Nucleic Acids Res. 2017, 45, D200–D203. [Google Scholar] [CrossRef] [PubMed]
- Mthethwa, B.; Chen, W.; Ngwenya, M.; Kappo, A.; Syed, P.; Karpoormath, R.; Yu, J.H.; Nelson, D.; Syed, K. Comparative analyses of cytochrome P450s and those associated with secondary metabolism in Bacillus Species. Int. J. Mol. Sci. 2018, 19, 3623. [Google Scholar] [CrossRef] [PubMed]
- Saeed, A.I.; Sharov, V.; White, J.; Li, J.; Liang, W.; Bhagabati, N.; Braisted, J.; Klapa, M.; Currier, T.; Thiagarajan, M.; et al. TM4: A free, open-source system for microarray data management and analysis. Biotechniques 2003, 34, 374–378. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Furumichi, M.; Tanabe, M.; Sato, Y.; Morishima, K. KEGG: New perspectives on genomes, pathways, diseases and drugs. Nucleic Acids Res. 2017, 45, D353–D361. [Google Scholar] [CrossRef] [PubMed]
- Cole, S.; Brosch, R.; Parkhill, J.; Garnier, T.; Churcher, C.; Harris, D.; Gordon, S.V.; Eiglmeier, K.; Gas, S.; Barry Iii, C.E.; et al. Deciphering the biology of Mycobacterium tuberculosis from the complete genome sequence. Nature 1998, 393, 537–544. [Google Scholar] [CrossRef] [PubMed]
- Fleischmann, R.D.; Alland, D.; Eisen, J.A.; Carpenter, L.; White, O.; Peterson, J.; DeBoy, R.; Dodson, R.; Gwinn, M.; Haft, D.; et al. Whole-genome comparison of Mycobacterium tuberculosis clinical and laboratory strains. J. Bacteriol. 2002, 184, 5479–5490. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Lu, L.; Wang, B.; Pu, S.; Zhang, X.; Zhu, G.; Shi, W.; Zhang, L.; Wang, H.; Wang, S.; et al. Genetic basis of virulence attenuation revealed by comparative genomic analysis of Mycobacterium tuberculosis strain H37Ra versus H37Rv. PLoS ONE 2008, 3, e2375. [Google Scholar] [CrossRef] [PubMed]
- Madhavilatha, G.K.; Joseph, B.V.; Paul, L.K.; Kumar, R.A.; Hariharan, R.; Mundayoor, S. Whole-genome sequences of two clinical isolates of Mycobacterium tuberculosis from Kerala, South India. J. Bacteriol. 2012, 194, 4430. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, C.; Liu, J.; Deng, H.; Pan, A.; Zhang, L.; Zhao, X.; Huang, M.; Lu, B.; Dong, H.; et al. Complete genome sequences of Mycobacterium tuberculosis strains CCDC5079 and CCDC5080, which belong to the Beijing family. J. Bacteriol. 2011, 193, 5591–5592. [Google Scholar] [CrossRef] [PubMed]
- Tang, B.; Wang, Q.; Yang, M.; Xie, F.; Zhu, Y.; Zhuo, Y.; Wang, S.; Gao, H.; Ding, X.; Zhang, L.; et al. ContigScape: A Cytoscape plugin facilitating microbial genome gap closing. BMC Genom. 2013, 14, 289. [Google Scholar] [CrossRef] [PubMed]
- Ilina, E.N.; Shitikov, E.A.; Ikryannikova, L.N.; Alekseev, D.G.; Kamashev, D.E.; Malakhova, M.V.; Parfenova, T.V.; Afanas’ev, M.V.; Ischenko, D.S.; Bazaleev, N.A.; et al. Comparative genomic analysis of Mycobacterium tuberculosis drug resistant strains from Russia. PLoS ONE 2013, 8, e56577. [Google Scholar] [CrossRef] [PubMed]
- Isaza, J.P.; Duque, C.; Gomez, V.; Robledo, J.; Barrera, L.F.; Alzate, J.F. Whole genome shotgun sequencing of one Colombian clinical isolate of Mycobacterium tuberculosis reveals DosR regulon gene deletions. FEMS Microbiol. Lett. 2012, 330, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Miyoshi-Akiyama, T.; Matsumura, K.; Iwai, H.; Funatogawa, K.; Kirikae, T. Complete annotated genome sequence of Mycobacterium tuberculosis Erdman. J. Bacteriol. 2012, 194, 2770. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, S.; Deshpande, U. Whole-genome sequences of four clinical isolates of Mycobacterium tuberculosis from Tamil Nadu, south India. Genome Announc. 2013, 1, e00186-13. [Google Scholar] [CrossRef] [PubMed]
- Roetzer, A.; Diel, R.; Kohl, T.A.; Rückert, C.; Nübel, U.; Blom, J.; Wirth, T.; Jaenicke, S.; Schuback, S.; Rüsch-Gerdes, S.; et al. Whole genome sequencing versus traditional genotyping for investigation of a Mycobacterium tuberculosis outbreak: A longitudinal molecular epidemiological study. PLoS Med. 2013, 10, e1001387. [Google Scholar] [CrossRef] [PubMed]
- Al Rashdi, A.S.A.; Jadhav, B.L.; Deshpande, T.; Deshpande, U. Whole-genome sequencing and annotation of a clinical isolate of Mycobacterium tuberculosis from Mumbai, India. Genome Announc. 2014, 2, e00154-14. [Google Scholar] [CrossRef] [PubMed]
- Garnier, T.; Eiglmeier, K.; Camus, J.C.; Medina, N.; Mansoor, H.; Pryor, M.; Duthoy, S.; Grondin, S.; Lacroix, C.; Monsempe, C.; et al. The complete genome sequence of Mycobacterium bovis. Proc. Natl. Acad. Sci. USA 2003, 100, 7877–7882. [Google Scholar] [CrossRef] [PubMed]
- Brosch, R.; Gordon, S.V.; Garnier, T.; Eiglmeier, K.; Frigui, W.; Valenti, P.; Dos Santos, S.; Duthoy, S.; Lacroix, C.; Garcia-Pelayo, C.; et al. Genome plasticity of BCG and impact on vaccine efficacy. Proc. Natl. Acad. Sci. USA 2007, 104, 5596–5601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seki, M.; Honda, I.; Fujita, I.; Yano, I.; Yamamoto, S.; Koyama, A. Whole genome sequence analysis of Mycobacterium bovis bacillus Calmette–Guerin (BCG) Tokyo 172: A comparative study of BCG vaccine substrains. Vaccine 2009, 27, 1710–1716. [Google Scholar] [CrossRef] [PubMed]
- Orduña, P.; Cevallos, M.A.; de León, S.P.; Arvizu, A.; Hernández-González, I.L.; Mendoza-Hernández, G.; López-Vidal, Y. Genomic and proteomic analyses of Mycobacterium bovis BCG Mexico 1931 reveal a diverse immunogenic repertoire against tuberculosis infection. BMC Genom. 2011, 12, 493. [Google Scholar] [CrossRef] [PubMed]
- Joung, S.M.; Jeon, S.J.; Lim, Y.J.; Lim, J.S.; Choi, B.S.; Choi, I.Y.; Yu, J.H.; Na, K.I.; Cho, E.H.; Shin, S.S.; et al. Complete genome sequence of Mycobacterium bovis BCG Korea, the Korean vaccine strain for substantial production. Genome Announc. 2013, 1, e00069-13. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Yang, X.; Duan, J.; Lu, N.; Leung, A.S.; Tran, V.; Hu, Y.; Wu, N.; Liu, D.; Wang, Z.; et al. The whole genome sequence of four BCG vaccine strains. J. Bacteriol. 2011, 193, 3152–3153. [Google Scholar] [CrossRef] [PubMed]
- Bentley, S.D.; Comas, I.; Bryant, J.M.; Walker, D.; Smith, N.H.; Harris, S.R.; Thurston, S.; Gagneux, S.; Wood, J.; Antonio, M.; et al. The genome of Mycobacterium africanum West African 2 reveals a lineage-specific locus and genome erosion common to the M. tuberculosis complex. PLoS Negl. Trop. Dis. 2012, 6, e1552. [Google Scholar] [CrossRef] [PubMed]
- Supply, P.; Marceau, M.; Mangenot, S.; Roche, D.; Rouanet, C.; Khanna, V.; Majlessi, L.; Criscuolo, A.; Tap, J.; Pawlik, A.; et al. Genomic analysis of smooth tubercle bacilli provides insights into ancestry and pathoadaptation of Mycobacterium tuberculosis. Nat. Genet. 2013, 45, 172. [Google Scholar] [CrossRef] [PubMed]
- Cole, S.T.; Eiglmeier, K.; Parkhill, J.; James, K.D.; Thomson, N.R.; Wheeler, P.R.; Honore, N.; Garnier, T.; Churcher, C.; Harris, D.; et al. Massive gene decay in the leprosy bacillus. Nature 2001, 409, 1007. [Google Scholar] [CrossRef] [PubMed]
- Monot, M.; Honoré, N.; Garnier, T.; Zidane, N.; Sherafi, D.; Paniz-Mondolfi, A.; Matsuoka, M.; Taylor, G.M.; Donoghue, H.D.; Bouwman, A.; et al. Comparative genomic and phylogeographic analysis of Mycobacterium leprae. Nat. Genet. 2009, 41, 1282–1289. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Bannantine, J.P.; Zhang, Q.; Amonsin, A.; May, B.J.; Alt, D.; Banerji, N.; Kanjilal, S.; Kapur, V. The complete genome sequence of Mycobacterium avium subspecies paratuberculosis. Proc. Natl. Acad. Sci. USA 2005, 102, 12344–12349. [Google Scholar] [CrossRef] [PubMed]
- Bannantine, J.P.; Li, L.; Mwangi, M.; Cote, R.; Garay, J.A.R.; Kapur, V. Complete genome sequence of Mycobacterium avium subsp. paratuberculosis, isolated from human breast milk. Genome Announc. 2014, 2, e01252-13. [Google Scholar] [CrossRef] [PubMed]
- Amin, A.S.; Hsu, C.Y.; Darwish, S.F.; Ghosh, P.; AbdEl-Fatah, E.M.; Behour, T.S.; Talaat, A.M. Ecology and genomic features of infection with Mycobacterium avium subspecies paratuberculosis in Egypt. Microbiology 2015, 161, 807–818. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.J.; Choi, B.S.; Lim, J.S.; Choi, I.Y.; Lee, J.H.; Chun, J.; Kook, Y.H.; Kim, B.J. Complete genome sequence of Mycobacterium intracellulare clinical strain MOTT-02. J. Bacteriol. 2012, 194, 2771. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.J.; Choi, B.S.; Lim, J.S.; Choi, I.Y.; Kook, Y.H.; Kim, B.J. Complete genome sequence of Mycobacterium intracellulare clinical strain MOTT-64, belonging to the INT1 genotype. J. Bacteriol. 2012, 194, 3268. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.J.; Choi, B.S.; Lim, J.S.; Choi, I.Y.; Lee, J.H.; Chun, J.; Kook, Y.H.; Kim, B.J. Complete genome sequence of Mycobacterium intracellulare strain ATCC 13950T. J. Bacteriol. 2012, 194, 2750. [Google Scholar] [CrossRef] [PubMed]
- Saini, V.; Raghuvanshi, S.; Khurana, J.P.; Ahmed, N.; Hasnain, S.E.; Tyagi, A.K.; Tyagi, A.K. Massive gene acquisitions in Mycobacterium indicus pranii provide a perspective on mycobacterial evolution. Nucleic Acids Res. 2012, 40, 10832–10850. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.J.; Kim, B.R.; Lee, S.Y.; Seok, S.H.; Kook, Y.H.; Kim, B.J. Whole-genome sequence of a novel species, Mycobacterium yongonense DSM 45126T. Genome Announc. 2013, 1, e00604-13. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.J.; Choi, B.S.; Choi, I.Y.; Lee, J.H.; Chun, J.; Hong, S.H.; Kook, Y.H.; Kim, B.J. Complete genome sequence of Mycobacterium intracellulare clinical strain MOTT-36Y, belonging to the INT5 genotype. J. Bacteriol. 2012, 194, 4141–4142. [Google Scholar] [CrossRef] [PubMed]
- Gallien, S.; Perrodou, E.; Carapito, C.; Deshayes, C.; Reyrat, J.M.; Van Dorsselaer, A.; Poch, O.; Schaeffer, C.; Lecompte, O. Ortho-proteogenomics: Multiple proteomes investigation through orthology and a new MS-based protocol. Genome Res. 2009, 19, 128–135. [Google Scholar] [CrossRef] [PubMed]
- Mohan, A.; Padiadpu, J.; Baloni, P.; Chandra, N. Complete genome sequences of a Mycobacterium smegmatis laboratory strain (MC2 155) and isoniazid-resistant (4XR1/R2) mutant strains. Genome Announc. 2015, 3, e01520-14. [Google Scholar] [CrossRef] [PubMed]
- Kallimanis, A.; Karabika, E.; Mavromatis, K.; Lapidus, A.; LaButti, K.M.; Liolios, K.; Ivanova, N.; Goodwin, L.; Woyke, T.; Velentzas, A.D.; et al. Complete genome sequence of Mycobacterium sp. strain (Spyr1) and reclassification to Mycobacterium gilvum Spyr1. Stand. Genom. Sci. 2011, 5, 144. [Google Scholar] [CrossRef] [PubMed]
- Bragin, E.Y.; Shtratnikova, V.Y.; Dovbnya, D.V.; Schelkunov, M.I.; Pekov, Y.A.; Malakho, S.G.; Egorova, O.V.; Ivashina, T.V.; Sokolov, S.L.; Ashapkin, V.V.; et al. Comparative analysis of genes encoding key steroid core oxidation enzymes in fast-growing Mycobacterium spp. strains. J. Steroid Biochem. Mol. Biol. 2013, 138, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Kato, H.; Ogawa, N.; Ohtsubo, Y.; Oshima, K.; Toyoda, A.; Mori, H.; Nagata, Y.; Kurokawa, K.; Hattori, M.; Fujiyama, A.; et al. Complete genome sequence of a phenanthrene degrader, Mycobacterium sp. strain EPa45 (NBRC 110737), isolated from a phenanthrene-degrading consortium. Genome Announc. 2015, 3, e00782-15. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Tao, F.; Li, F.; Hou, J.; Tang, H.; Ma, C.; Xu, P. Complete genome sequence of Mycobacterium goodii X7B, a facultative thermophilic biodesulfurizing bacterium with industrial potential. J. Biotechnol. 2015, 212, 56–57. [Google Scholar] [CrossRef] [PubMed]
- Costa, K.C.; Bergkessel, M.; Saunders, S.; Korlach, J.; Newman, D.K. Enzymatic degradation of phenazines can generate energy and protect sensitive organisms from toxicity. mBio 2015, 6, e01520-15. [Google Scholar] [CrossRef] [PubMed]
- Stinear, T.P.; Seemann, T.; Pidot, S.; Frigui, W.; Reysset, G.; Garnier, T.; Meurice, G.; Simon, D.; Bouchier, C.; Ma, L.; et al. Reductive evolution and niche adaptation inferred from the genome of Mycobacterium ulcerans, the causative agent of Buruli ulcer. Genome Res. 2007, 17, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.Y.; Sun, Z.Q.; Wang, Z.L.; Wen, Z.L.; Sun, Q.W.; Zhu, Z.Q.; Song, Y.Z.; Zhao, J.W.; Wang, H.H.; Zhang, S.L.; et al. Complete genome sequence of a novel clinical isolate, the nontuberculous Mycobacterium strain JDM601. J. Bacteriol. 2011, 193, 4300–4301. [Google Scholar] [CrossRef] [PubMed]
- Stinear, T.P.; Seemann, T.; Harrison, P.F.; Jenkin, G.A.; Davies, J.K.; Johnson, P.D.; Abdellah, Z.; Arrowsmith, C.; Chillingworth, T.; Churcher, C.; et al. Insights from the complete genome sequence of Mycobacterium marinum on the evolution of Mycobacterium tuberculosis. Genome Res. 2008, 18, 729–741. [Google Scholar] [CrossRef] [PubMed]
- Tobias, N.J.; Doig, K.D.; Medema, M.H.; Chen, H.; Haring, V.; Moore, R.; Seemann, T.; Stinear, T.P. Complete genome sequence of the frog pathogen Mycobacterium ulcerans ecovar Liflandii. J. Bacteriol. 2013, 195, 556–564. [Google Scholar] [CrossRef] [PubMed]
- Tufariello, J.M.; Kerantzas, C.A.; Vilchèze, C.; Calder, R.B.; Nordberg, E.K.; Fischer, J.A.; Hartman, T.E.; Yang, E.; Driscoll, T.; Cole, L.E.; et al. The complete genome sequence of the emerging pathogen Mycobacterium haemophilum explains its unique culture requirements. mBio 2015, 6, e01313-15. [Google Scholar] [CrossRef] [PubMed]
- Ripoll, F.; Pasek, S.; Schenowitz, C.; Dossat, C.; Barbe, V.; Rottman, M.; Macheras, E.; Heym, B.; Herrmann, J.L.; Daffé, M.; et al. Non mycobacterial virulence genes in the genome of the emerging pathogen Mycobacterium abscessus. PLoS ONE 2009, 4, e5660. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.J.; Kim, B.R.; Hong, S.H.; Seok, S.H.; Kook, Y.H.; Kim, B.J. Complete genome sequence of Mycobacterium massiliense clinical strain Asan 50594, belonging to the type II genotype. Genome Announc. 2013, 1, e00429-13. [Google Scholar] [CrossRef] [PubMed]
- Raiol, T.; Ribeiro, G.M.; Maranhão, A.Q.; Bocca, A.L.; Silva-Pereira, I.; Junqueira-Kipnis, A.P.; de Macedo Brigido, M.; Kipnis, A. Complete genome sequence of Mycobacterium massiliense. J. Bacteriol. 2012, 194. [Google Scholar] [CrossRef] [PubMed]
- Sekizuka, T.; Kai, M.; Nakanaga, K.; Nakata, N.; Kazumi, Y.; Maeda, S.; Makino, M.; Hoshino, Y.; Kuroda, M. Complete genome sequence and comparative genomic analysis of Mycobacterium massiliense JCM 15300 in the Mycobacterium abscessus group reveal a conserved genomic island MmGI-1 related to putative lipid metabolism. PLoS ONE 2014, 9, e114848. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene Number | Protein Name |
|---|---|---|
| mce4E/lprN | Rv3495c a,c,d | Mce4 transport system |
| mce4C | Rv3497c a,c,d | Mce4 transport system |
| mce4A | Rv3499c a,c,d | Mce4 transport system |
| yrb4A/YrbE4A/supA | Rv3501c a,c,d | possible ABC transporter (Sterol uptake permease subunit) |
| hsd4A | Rv3502c a,c,d | 17β-hydroxysteroid dehydrogenase (17β-HSD) |
| kshA | Rv3526 a,c,d | kerosteroid-9α-hydroxylase, oxygenase |
| hsaF | Rv3534c a,c,d | probable 4-hydroxy-2-oxovalerate aldolase / 4-hydroxy-2-ketovalerate aldolase |
| kstD | Rv3537 b,c,d | 3-ketosteroid-Δ1-dehydrogenase (Δ1-KSTD) |
| fadE28 | Rv3544c a,b,c | probable acyl-CoA dehydrogenase |
| ipdA | Rv3551 a,b,c | ATP-dependent CoA transferase α subunit |
| fadE30 | Rv3560c a,b,c | probable acyl-CoA dehydrogenase |
| fadE32 | Rv3563 a,b,c | probable acyl-CoA dehydrogenase |
| hsaC | Rv3568c a,c,d | 3,4-DHSA dioxygenase |
| hsaD | Rv3569c b,c,d | 4,9-DHSA hydrolase |
| hsaA | Rv3570c b,c,d | 3-hydroxy-9,10-seconandrost-1,3,5(10)-triene-9,17-dione hydroxylase (3-HSA hydroxylase, reductase) |
| kshB | Rv3571 a,c,d | ketosteroid-9α-hydroxylase, reductase |
| mce4F | Rv3494c c,d | Mce4 transport system |
| mce4D | Rv3496c c,d | Mce4 transport system |
| mce4B | Rv3498c c,d | Mce4 transport system |
| yrb4B/YrbE4B/supB | Rv3500c c,d | possible ABC transporter (Sterol uptake permease subunit) |
| fadD19 | Rv3515c c,d | probable fatty-acid-CoA ligase |
| ltp3 | Rv3523 a,d | probable ketoacyl-CoA thiolase |
| hsaE | Rv3536c c,d | probable hydratase / 2-hydroxypentadienoate hydratase |
| ltp2 | Rv3540c a,c | probable ketoacyl-CoA thiolase |
| Rv3542c a,c | CHP / putative enoyl-CoA hydratase | |
| cyp125 | Rv3545c a,c | cytochrome P450 |
| fadA5 | Rv3546 a,c | acetoacetyl-CoA thiolase |
| fadA6 | Rv3556c a,b | acetoacetyl-CoA thiolase |
| ppiA | Rv0009 c | iron-regulated peptidyl-prolyl cis-trans isomerase A |
| fadD10 | Rv0099 e | fatty acid-CoA synthase |
| ptbB | Rv0153c c | phosphotyrosine protein phosphatase PTPB (protein-tyrosine-phosphatase) (PTPase) |
| mmpL11 | Rv0202c c | transmembrane transport protein MmpL11 |
| fadE5 | Rv0244c c | acyl-CoA dehydrogenase |
| mgtE | Rv0362 c | Mg2+ transport transmembrane protein MgtE |
| metZ | Rv0391 c | O-succinylhomoserine sulfhydrylase |
| mmpL4 | Rv0450c c | transmembrane transport protein MmpL4 |
| fadB2 | Rv0468 e | hydroxybutyryl-CoA dehydrogenase |
| Rv0485 c | transcriptional regulatory protein | |
| Rv0495c c | HP | |
| mkl | Rv0655 c | ribonucleotide ABC transporter ATP-binding protein |
| pqqE | Rv0693 c | coenzyme PQQ synthesis protein E |
| lldD1 | Rv0694 c | L-lactate dehydrogenase (cytochrome) LldD1 |
| Rv0695 c | HP | |
| Rv0696 c | membrane sugar transferase | |
| adhB | Rv0761c c | zinc-containing alcohol dehydrogenase NAD dependent ADHB |
| Rv0805 c | HP | |
| Rv0876c c | transmembrane protein | |
| echA9 | Rv1071c c | 3-hydroxyisobutyryl-CoA hydrolase |
| Rv1084 c | HP | |
| Rv1096 c | glycosyl hydrolase | |
| Rv1106c e | 3β-HSD | |
| Rv1129c c | transcriptional regulator protein | |
| Rv1130 c | HP | |
| gltA1 | Rv1131 c | citrate synthase |
| mmpL10 | Rv1183 c | transmembrane transport protein MmpL10 |
| fadD36 | Rv1193 c | acyl-CoA synthetase |
| mbtN (fadE14) | Rv1346 e | acyl-CoA dehydrogenase |
| Rv1428c c | HP | |
| Rv1432 c | dehydrogenase | |
| bcpB | Rv1608c c | peroxidoxin BcpB |
| Rv1626 c | two-component system transcriptional regulator | |
| Rv1627c c | lipid-transfer protein | |
| fadB3 | Rv1715 e | hydroxybutyryl-CoA dehydrogenase |
| Rv1798 c | HP | |
| Rv1906c c | HP | |
| Rv1919c c | HP | |
| mce3R | Rv1963c c | transcriptional repressor (probably TETR-family) MCE3R |
| pks12 | Rv2048c c | polyketide synthase pks12 |
| Rv2118c c | RNA methyltransferase | |
| Rv2206 c | transmembrane protein | |
| Rv2239c c | HP | |
| eis | Rv2416c c | HP |
| tig | Rv2462c c | trigger factor |
| Rv2506 c | TetR family transcriptional regulator | |
| fadD9 | Rv2590 e | fatty acid-CoA synthase |
| Rv2668 c | HP | |
| Rv2681 c | HP | |
| arsA | Rv2684 c | arsenic-transport integral membrane protein ArsA |
| sigB | Rv2710 c | RNA polymerase sigma factor SigB |
| Rv2799 c | HP | |
| pknI | Rv2914c c | transmembrane serine/threonine-protein kinase I |
| mutT1 | Rv2985 c | hydrolase MutT1 |
| Rv3050c c | AsnC family transcriptional regulator | |
| fadE22 | Rv3061c e | acyl-CoA dehydrogenase |
| fadE24 | Rv3139 e | acyl-CoA dehydrogenase |
| fadE23 | Rv3140 e | acyl-CoA dehydrogenase |
| fadE25 | Rv3274c c | acyl-CoA dehydrogenase FADE25 |
| choD | Rv3409c d | cholesterol oxidase |
| gcp | Rv3419c c | putative DNA-binding/iron metalloprotein/AP endonuclease |
| Rv3421c c | HP | |
| Rv3492c c | CHP MCE associated protein | |
| Rv3493c c | CHP MCE associated protein | |
| fdxD | Rv3503c e | probable ferredoxin |
| fadE26 | Rv3504 d | probable acyl-CoA dehydrogenase |
| fadE27 | Rv3505 d | probable acyl-CoA dehydrogenase |
| fadD17 | Rv3506 d | possible fatty-acid-CoA ligase |
| PE PGRS53 | Rv3507 e | PE PGRS family |
| PE PGRS54 | Rv3508 e | PE PGRS family |
| ilvX | Rv3509c e | probable acetohydroxy-acid synthase |
| Rv3510c e | CHP | |
| PE PGRS55 | Rv3511 e | PE PGRS family |
| PE PGRS56 | Rv3512 e | PE PGRS family |
| fadD18 | Rv3513c e | possible fatty-acid-CoA ligase |
| PE PGRS57 | Rv3514 e | PE PGRS family |
| echA19 | Rv3516 d | possible enoyl-CoA hydratase |
| whiB3 | Rv3517 e | conserved hypothetical protein (CHP) / transcription factor |
| cyp142 | Rv3518c e | cytochrome P450 |
| Rv3519 a | CHP | |
| Rv3520c e | coenzyme F420-dependent oxidoreductase | |
| Rv3521 e | CHP | |
| ltp4 | Rv3522 d | probable ketoacyl-CoA thiolase |
| Rv3524 e | probable conserved membrane protein | |
| Rv3525c e | possible siderophore binding protein | |
| Rv3527 a | hypothetical protein (HP) | |
| Rv3528c e | HP | |
| Rv3529c e | CHP | |
| Rv3530c e | possible oxidoreductase | |
| Rv3531c c | hypothetical protein | |
| PPE61 | Rv3532 e | PPE family |
| PPE62 | Rv3533c e | PPE family |
| hsaG | Rv3535c d | probable aldehyde dehydrogenase |
| hsd4B | Rv3538 d | probable enoyl-CoA hydratase |
| PPE63 | Rv3539 e | PE |
| Rv3541c a | CHP / putative enoyl-CoA hydratase | |
| fadE29 | Rv3543c c | probable acyl-CoA dehydrogenase |
| Rv3547 e | CHP | |
| Rv3548c c | probable short chain dehydrogenase/reductase | |
| Rv3549c c | probable short chain dehydrogenase/reductase | |
| echA20 | Rv3550 e | possible enoyl-CoA hydratase |
| ipdB | Rv3552 a | ATP-dependent CoA transferase β subunit |
| Rv3553 c | possible oxidoreductase / 2-nitropropane dioxygenase | |
| fdxB | Rv3554 e | possible electron transfer protein / ferredoxin |
| Rv3555c e | CHP | |
| kstR2 | Rv3557c e | Tet-R transcriptional regulator (repressor) |
| PPE64 | Rv3558 e | PPE |
| Rv3559c c | probable oxidoreductase | |
| fadD3 | Rv3561 c | acyl-CoA synthetase (AMP forming) |
| fadE31 | Rv3562 e | probable acyl-CoA dehydrogenase |
| fadE33 | Rv3564 c | probable acyl-CoA dehydrogenase |
| aspB | Rv3565 e | possible aspartate aminotransferase |
| Rv3566A e | CHP | |
| nhoA/nat | Rv3566c e | arylamine N-acetyltransferase |
| hsaB | Rv3567c d | 3-hydroxy-9,10-seconandrost-1,3,5(10)-triene-9,17-dione hydroxylase (3-HSA hydroxylase, reductase) |
| Rv3572 c | HP | |
| fadE34 | Rv3573c c | probable acyl-CoA dehydrogenase |
| kstR | Rv3574 a | Tet-R transcriptional regulator (repressor) |
| Rv3575c c | transcriptional regulatory protein LacI-family | |
| Rv3779 c | transmembrane protein alanine and leucine rich | |
| papA2 | Rv3820c c | polyketide synthase associated protein PapA2 |
| papA1 | Rv3824c c | polyketide synthase associated protein |
| pks2 | Rv3825c c | polyketide synthase PKS2 |
| sigM | Rv3911 c | RNA polymerase sigma factor SigM |
| Organism Code | H37Rv Homolog(s) Not Found Relating to Cholesterol Catabolism | Ability to Degrade Cholesterol | |||
|---|---|---|---|---|---|
| Proven to Be Essential | Predicted to Be Essential or Specifically Required | Predicted to Be Involved | Involved but Not Proven or Predicted to Be Essential | ||
| Mycobacterium tuberculosis complex (MTBC) | |||||
| mtu | None | None | None | None | Positive |
| mtv | None | None | None | None | Positive |
| mtc | None | None | None | Rv3555c | Positive |
| mra | None | None | None | None | Positive |
| mtf | None | None | None | Rv3566A | Positive |
| mtb | None | None | None | Rv3566A | Positive |
| mtk | None | None | None | Rv3566A | Positive |
| mtz | None | None | None | Rv3566A | Positive |
| mtg | None | Rv1084 Rv2799 | None | None | No prediction |
| mti | Rv3526 | Rv0153c Rv0485 Rv0805 Rv0876c Rv2416c Rv2681 Rv3526 Rv3531c | Rv3526 | None | No prediction |
| mte | None | Rv0805 Rv1919c | None | Rv3566A | No prediction |
| mtur | None | None | None | None | Positive |
| mtl | None | Rv0805 Rv1919c | None | Rv3566A | No prediction |
| mto | None | None | None | None | Positive |
| mtd | None | None | None | None | Positive |
| mtn | None | Rv0805 | None | Rv3566A | No prediction |
| mtj | None | None | None | Rv3566A | Positive |
| mtub | None | None | None | None | Positive |
| mtuc | None | Rv0485 Rv0695 Rv1084 Rv1130 Rv1432 Rv2416c Rv2681 Rv3536c Rv3779 | Rv3536c | Rv3521 Rv3566A | No prediction |
| mtue | None | Rv2681 | None | Rv3566A | No prediction |
| mtx | None | None | None | None | Positive |
| mtuh | None | Rv0485 Rv0876c Rv1084 Rv1096 Rv1129c Rv2416c Rv3531c | None | None | No prediction |
| mtul | None | None | None | Rv3566A | Positive |
| mtut | None | None | None | None | Positive |
| mtuu | None | None | None | None | Positive |
| mtq | None | None | None | None | Positive |
| mbo | None | None | None | None | Positive |
| mbb | None | None | None | None | Positive |
| mbt | None | None | None | None | Positive |
| mbm | None | None | None | None | Positive |
| mbk | None | None | None | Rv3566A | Positive |
| mbx | None | Rv0805 Rv2206 | None | Rv3566A Rv3566c | No prediction |
| mbz | None | None | None | None | Positive |
| maf | None | None | None | Rv3528c | Positive |
| mce | None | Rv1130 | None | None | No prediction |
| mcq | None | None | None | None | Positive |
| mcv | None | None | None | None | Positive |
| mcx | None | None | Rv3566A | Positive | |
| mcz | None | None | None | Rv3517 Rv3528c Rv3566A | Positive |
| Mycobacterium chelonae-abscessus complex (MCAC) | |||||
| mab | Rv3519 | Rv0876c Rv1906c Rv2684 | None | Rv3507 Rv3508 Rv3511 Rv3514 Rv3524 Rv3528c Rv3566A | No prediction |
| mabb | Rv3519 | Rv0876c Rv1906c Rv2684 Rv3575c | None | Rv3507 Rv3508 Rv3511 Rv3514 Rv3524 Rv3528c Rv3566A | No prediction |
| mmv | Rv3519 | Rv0876c Rv2684 Rv3575c | None | Rv3507 Rv3508 Rv3511 Rv3514 Rv3524 Rv3528c Rv3566A | No prediction |
| may | Rv3519 | Rv1906c Rv2684 | None | Rv3507 Rv3508 Rv3511 Rv3514 Rv3524 Rv3528c Rv3566A | No prediction |
| mabo | Rv3519 | Rv1906c Rv2684 | None | Rv3507 Rv3508 Rv3511 Rv3514 Rv3524 Rv3528c Rv3566A | No prediction |
| mabl | Rv3519 | Rv0876c Rv1906c Rv2684 Rv3575c | None | Rv3507 Rv3508 Rv3511 Rv3514 Rv3517 Rv3524 Rv3528c Rv3566A | No prediction |
| maz | Rv3519 | Rv1906c Rv2684 | None | Rv3507 Rv3508 Rv3511 Rv3514 Rv3524 Rv3528c Rv3566A | No prediction |
| mak | Rv3519 | Rv1906c Rv2684 Rv3575c | None | Rv3507 Rv3508 Rv3511 Rv3514 Rv3524 Rv3528c Rv3566A | No prediction |
| mys | Rv3519 | Rv2684 Rv3575c | None | Rv3507 Rv3508 Rv3511 Rv3514 Rv3524 Rv3528c Rv3566A | No prediction |
| myc | Rv3519 | Rv2684 Rv3575c | None | Rv3507 Rv3508 Rv3511 Rv3514 Rv3524 Rv3528c Rv3566A | No prediction |
| Mycobacterium avium complex (MAC) | |||||
| mpa | None | None | None | Rv3528c Rv3566A | Positive |
| mao | None | Rv0153c | None | Rv3528c Rv3566A | No prediction |
| mavi | None | Rv0153c Rv1084 | None | Rv3528c Rv3566A | No prediction |
| mavu | None | None | None | Rv3528c Rv3566A | Positive |
| mav | None | Rv3779 | None | Rv3528c Rv3566A | No prediction |
| mavd | None | Rv0153c | None | Rv3528c Rv3566A | No prediction |
| mavr | None | None | None | Rv3528c Rv3566A | Positive |
| mava | None | None | None | Rv3528c Rv3566A | Positive |
| mit | Rv3519 | None | None | Rv3528c Rv3566A | No prediction |
| mir | None | None | None | Rv3528c Rv3566A | Positive |
| mia | None | None | None | Rv3528c Rv3566A | Positive |
| mie | None | None | None | Rv3528c Rv3566A | Positive |
| mid | None | None | None | Rv3528c Rv3566A | Positive |
| myo | None | None | None | Rv3528c Rv3566A | Positive |
| mmm | None | None | None | Rv3528c Rv3566A | Positive |
| Mycobacteria causing leprosy (MCL) | |||||
| mle | Rv3523 Rv3526 Rv3540c Rv3551 Rv3568c Rv3571 Rv3519 Rv3527 Rv3552 | Rv0153c Rv0485 Rv0693 Rv0695 Rv1084 Rv1129c Rv1130 Rv2416c Rv2668 Rv2799 Rv3492c Rv3493c Rv3526 Rv3531c Rv3536c Rv3540c Rv3551 Rv3553 Rv3568c Rv3571 | Rv3523 Rv3526 Rv3535c Rv3536c Rv3568c Rv3571 | Rv3503c Rv3510c Rv3517 Rv3521 Rv3524 Rv3528c Rv3529c Rv3554 Rv3555c Rv3566A Rv3566c | Negative |
| mlb | Rv3523 Rv3526 Rv3540c Rv3551 Rv3568c Rv3571 Rv3519 Rv3527 Rv3552 | Rv0153c Rv0485 Rv0693 Rv0695 Rv1084 Rv1129c Rv1130 Rv2416c Rv2668 Rv2799 Rv3492c Rv3493c Rv3526 Rv3531c Rv3536c Rv3540c Rv3551 Rv3553 Rv3568c Rv3571 | Rv3523 Rv3526 Rv3535c Rv3536c Rv3568c Rv3571 | Rv3503c Rv3510c Rv3517 Rv3521 Rv3524 Rv3528c Rv3529c Rv3554 Rv3555c Rv3566A Rv3566c | Negative |
| Non-tuberculosis Mycobacterium (NTM) | |||||
| mul | None | Rv2416c | None | Rv3517 Rv3528c Rv3566A | No prediction |
| mjd | None | Rv3575c | None | Rv3528c Rv3566A | No prediction |
| mmi | None | None | None | Rv3528c Rv3566A | Positive |
| mli | None | None | None | Rv3528c Rv3566A | Positive |
| mkn | None | None | None | Rv3528c Rv3566A | Positive |
| mks | None | Rv2462c | None | Rv3528c Rv3566A | No prediction |
| mki | None | Rv2462c | None | Rv3528c Rv3566A | No prediction |
| mhad | Rv3534c | Rv1130 Rv3534c | Rv3534c | Rv3528c Rv3566A | No prediction |
| Saprophytes (SAP) | |||||
| msm | None | Rv0805 Rv3572 Rv3779 | None | Rv3507 Rv3508 Rv3511 Rv3514 Rv3528c Rv3566A | No prediction |
| msg | None | Rv0805 Rv3572 Rv3779 | None | Rv3507 Rv3508 Rv3511 Rv3514 Rv3528c Rv3566A | No prediction |
| msb | None | Rv0805 Rv3572 Rv3779 | None | Rv3507 Rv3508 Rv3511 Rv3514 Rv3528c Rv3566A | No prediction |
| msn | None | Rv0805 Rv3493c Rv3572 Rv3779 | None | Rv3507 Rv3508 Rv3511 Rv3514 Rv3528c Rv3566A | No prediction |
| msh | None | Rv0805 Rv3572 Rv3779 | None | Rv3507 Rv3508 Rv3511 Rv3514 Rv3528c Rv3566A | No prediction |
| msa | None | Rv1130 | None | Rv3507 Rv3508 Rv3511 Rv3514 Rv3517 Rv3528c | No prediction |
| mva | None | Rv0805 Rv1130 Rv3572 Rv3779 | None | Rv3507 Rv3508 Rv3511 Rv3514 Rv3517 Rv3528c Rv3566A | No prediction |
| mgi | None | Rv0805 Rv1130 Rv3572 Rv3779 | None | Rv3507 Rv3508 Rv3514 Rv3528c Rv3566A | No prediction |
| msp | None | Rv0805 Rv1084 Rv1130 Rv1919c Rv3492c Rv3572 Rv3779 | None | Rv3507 Rv3508 Rv3511 Rv3514 Rv3528c Rv3566A | No prediction |
| mmc | None | Rv0805 Rv1130 Rv3572 Rv3779 | None | Rv3507 Rv3508 Rv3511 Rv3514 Rv3528c Rv3566A | No prediction |
| mkm | None | Rv0805 Rv1130 Rv3572 Rv3779 | None | Rv3507 Rv3508 Rv3511 Rv3514 Rv3528c Rv3566A | No prediction |
| mjl | None | Rv0805 Rv1130 Rv3572 Rv3779 | None | Rv3507 Rv3508 Rv3511 Rv3514 Rv3528c Rv3566A | No prediction |
| mrh | None | Rv0805 Rv1130 Rv3572 Rv3779 | None | Rv3507 Rv3508 Rv3514 Rv3528c Rv3566A | No prediction |
| mcb | None | Rv1130 Rv2416c | None | Rv3507 Rv3508 Rv3511 Rv3514 Rv3528c Rv3566A Rv3566c | No prediction |
| mne | None | Rv0805 Rv3572 Rv3779 | None | Rv3507 Rv3508 Rv3511 Rv3514 Rv3517 Rv3528c Rv3566A | No prediction |
| myv | None | Rv0805 Rv3572 | None | Rv3507 Rv3508 Rv3511 Rv3514 Rv3528c | No prediction |
| mye | None | Rv0876c Rv1130 Rv2416c | None | Rv3507 Rv3508 Rv3511 Rv3517 Rv3528c Rv3566A Rv3566c | No prediction |
| mgo | None | Rv0805 Rv0876c Rv3572 | None | Rv3507 Rv3508 Rv3511 Rv3514 Rv3528c Rv3566A | No prediction |
| mft | None | Rv3572 | None | Rv3507 Rv3508 Rv3511 Rv3514 Rv3528c | No prediction |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
van Wyk, R.; van Wyk, M.; Mashele, S.S.; Nelson, D.R.; Syed, K. Comprehensive Comparative Analysis of Cholesterol Catabolic Genes/Proteins in Mycobacterial Species. Int. J. Mol. Sci. 2019, 20, 1032. https://doi.org/10.3390/ijms20051032
van Wyk R, van Wyk M, Mashele SS, Nelson DR, Syed K. Comprehensive Comparative Analysis of Cholesterol Catabolic Genes/Proteins in Mycobacterial Species. International Journal of Molecular Sciences. 2019; 20(5):1032. https://doi.org/10.3390/ijms20051032
Chicago/Turabian Stylevan Wyk, Rochelle, Mari van Wyk, Samson Sitheni Mashele, David R. Nelson, and Khajamohiddin Syed. 2019. "Comprehensive Comparative Analysis of Cholesterol Catabolic Genes/Proteins in Mycobacterial Species" International Journal of Molecular Sciences 20, no. 5: 1032. https://doi.org/10.3390/ijms20051032
APA Stylevan Wyk, R., van Wyk, M., Mashele, S. S., Nelson, D. R., & Syed, K. (2019). Comprehensive Comparative Analysis of Cholesterol Catabolic Genes/Proteins in Mycobacterial Species. International Journal of Molecular Sciences, 20(5), 1032. https://doi.org/10.3390/ijms20051032