Histone Acetylation of Immune Regulatory Genes in Human Placenta in Association with Maternal Intake of Olive Oil and Fish Consumption

 ,
,  , , , , , and
, , , , , and 
Abstract
:1. Introduction
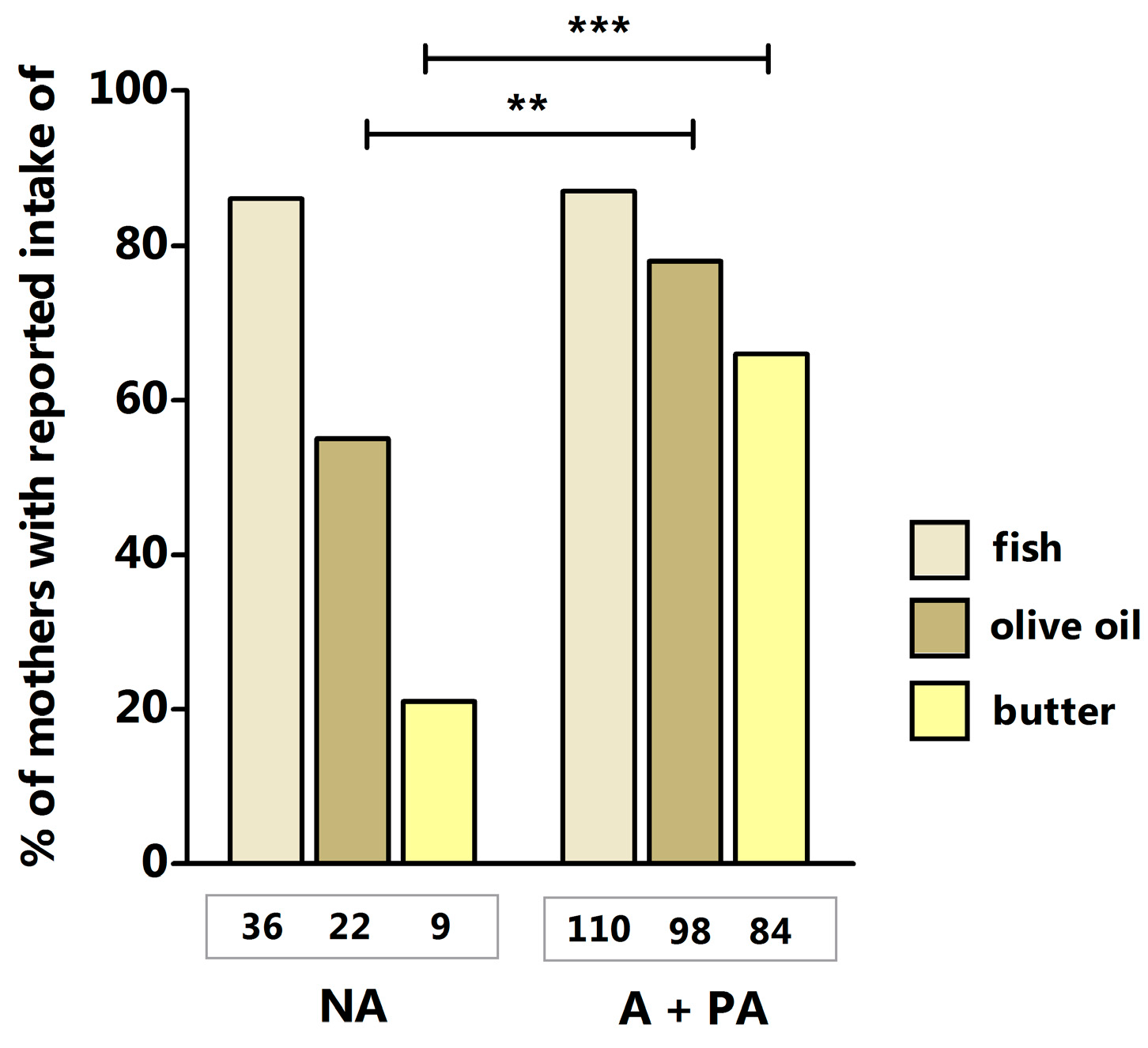
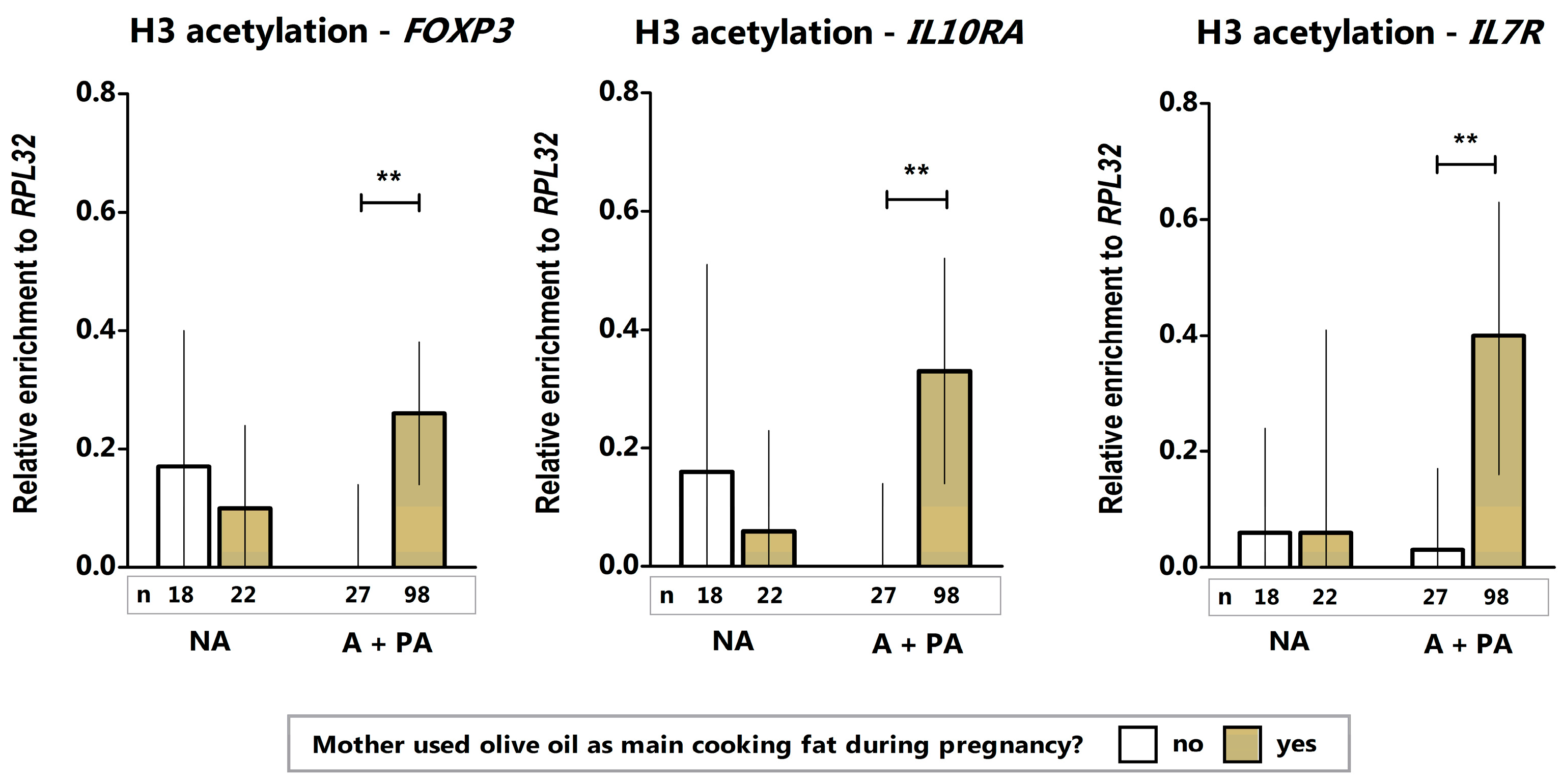
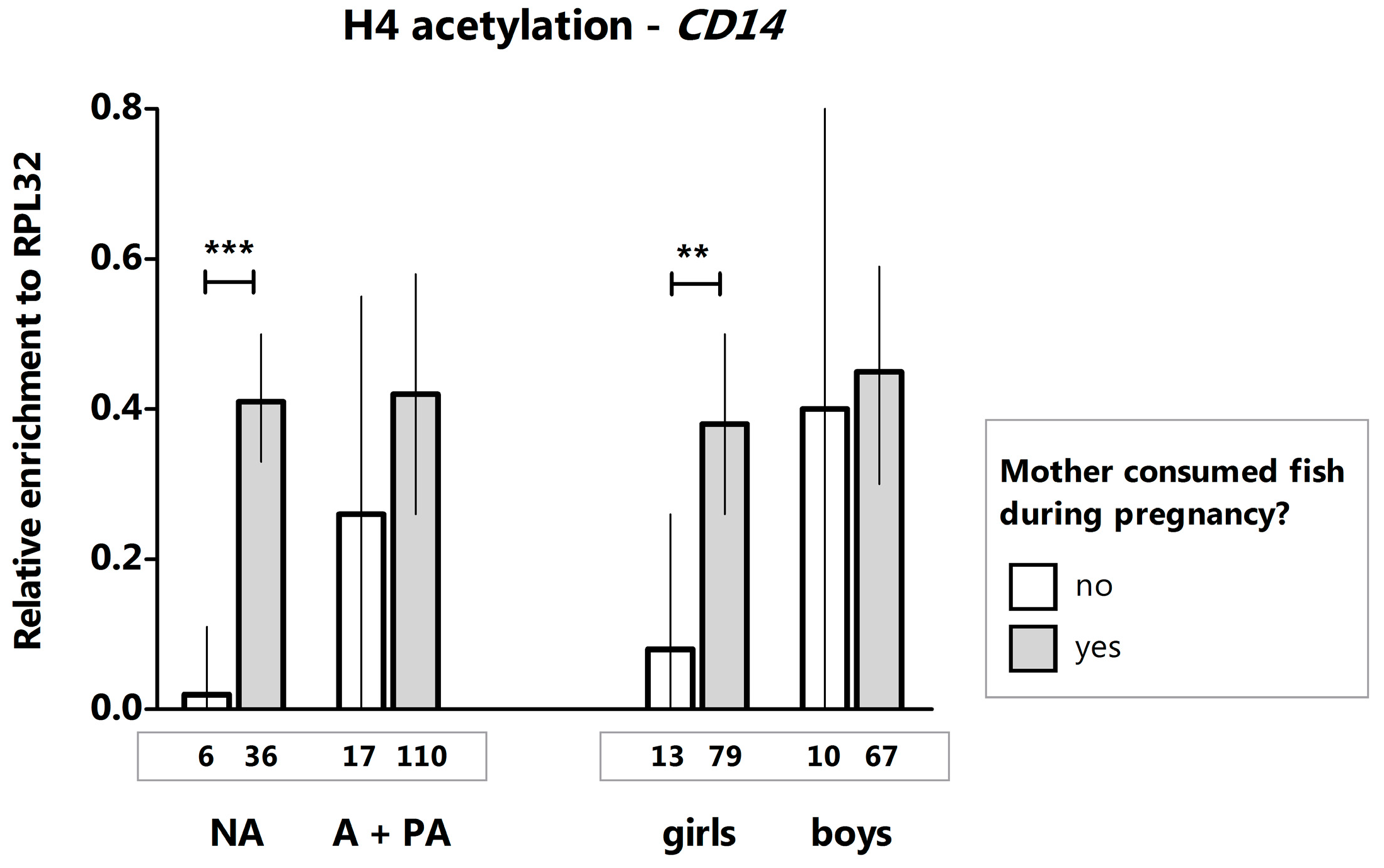
2. Results and Discussion
3. Methods
3.1. Study Population—the ALADDIN Cohort
3.2. Collection of Placenta Specimens and Histopathology Examination
3.3. Isolation of Chromatin, Chromatin Immunoprecipitation, and Quantitative Polymerase Chain Reaction (qPCR)
3.4. Statistics
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lecoutre, S.; Pourpe, C.; Butruille, L.; Marousez, L.; Laborie, C.; Guinez, C.; Lesage, J.; Vieau, D.; Eeckhoute, J.; Gabory, A.; et al. Reduced PPARgamma2 expression in adipose tissue of male rat offspring from obese dams is associated with epigenetic modifications. FASEB J. 2018, 32, 2768–2778. [Google Scholar] [CrossRef] [PubMed]
- Block, T.; El-Osta, A. Epigenetic programming, early life nutrition and the risk of metabolic disease. Atherosclerosis 2017, 266, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S. Impact of Maternal Diet on the Epigenome during In Utero Life and the Developmental Programming of Diseases in Childhood and Adulthood. Nutrients 2015, 7, 9492–9507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.R.; Lazarenko, O.P.; Zhao, H.; Alund, A.W.; Shankar, K. Maternal obesity impairs skeletal development in adult offspring. J. Endocrinol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Isaac, A.R.; da Silva, E.A.N.; de Matos, R.J.B.; Augusto, R.L.; Moreno, G.M.M.; Mendonça, I.P.; de Souza, R.F.; Cabral-Filho, P.E.; Rodrigues, C.G.; Gonçalves-Pimentel, C.; et al. Low omega-6/omega-3 ratio in a maternal protein-deficient diet promotes histone-3 changes in progeny neural cells and favors leukemia inhibitory factor genetranscription. J. Nutr. Biochem. 2018, 55, 229–242. [Google Scholar] [CrossRef] [PubMed]
- Upadhyaya, B.; Larsen, T.; Barwari, S.; Louwagie, E.J.; Baack, M.L.; Dey, M. Prenatal Exposure to a Maternal High-Fat Diet Affects Histone Modification of Cardiometabolic Genes in Newborn Rats. Nutrients 2017, 9, 407. [Google Scholar] [CrossRef] [PubMed]
- Voss, A.K.; Thomas, T. Histone Lysine and Genomic Targets of Histone Acetyltransferases in Mammals. BioEssays 2018, 40, e1800078. [Google Scholar] [CrossRef] [PubMed]
- Brook, P.O.; Perry, M.M.; Adcock, I.M.; Durham, A.L. Epigenome-modifying tools in asthma. Epigenomics 2015, 7, 1017–1032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vahid, F.; Zand, H.; Nosrat-Mirshekarlou, E.; Najafi, R.; Hekmatdoost, A. The role dietary of bioactive compounds on the regulation of histone acetylases and deacetylases: A review. Gene 2015, 562, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Triff, K.; McLean, M.W.; Callaway, E.; Goldsby, J.; Ivanov, I.; Chapkin, R.S. Dietary fat and fiber interact to uniquely modify global histone post-translational epigenetic programming in a rat colon cancer progression model. Int. J. Cancer 2018, 143, 1402–1415. [Google Scholar] [CrossRef] [PubMed]
- Harb, H.; Irvine, J.; Amarasekera, M.; Hii, C.S.; Kesper, D.A.; Ma, Y.F.; D’Vaz, N.; Renz, H.; Potaczek, D.P.; Prescott, S.L.; et al. The role of PKCzeta in cord blood T-cell maturation towards Th1 cytokine profile and its epigenetic regulation by fish oil. Biosci. Rep. 2017, 37, BSR20160485. [Google Scholar] [CrossRef] [PubMed]
- Alaskhar Alhamwe, B.; Khalaila, R.; Wolf, J.; von Bulow, V.; Harb, H.; Alhamdan, F.; Hii, C.S.; Prescott, S.L.; Ferrante, A.; Renz, H.; et al. Histone modifications and their role in epigenetics of atopy and allergic diseases. Allergy Asthma Clin. Immunol. 2018, 14, 39. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.G.; Lim, H.W.; Andrisani, O.M.; Broxmeyer, H.E.; Kim, C.H. Vitamin A metabolites induce gut-homing FoxP3+ regulatory T cells. J. Immunol. 2007, 179, 3724–3733. [Google Scholar] [CrossRef] [PubMed]
- Furusawa, Y.; Obata, Y.; Fukuda, S.; Endo, T.A.; Nakato, G.; Takahashi, D.; Nakanishi, Y.; Uetake, C.; Kato, K.; Kato, T.; et al. Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature 2013, 504, 446–450. [Google Scholar] [CrossRef] [PubMed]
- Arpaia, N.; Campbell, C.; Fan, X.; Dikiy, S.; van der Veeken, J.; de Roos, P.; Liu, H.; Cross, J.R.; Pfeffer, K.; Coffer, P.J.; et al. Metabolites produced by commensal bacteria promote peripheral regulatory T-cell generation. Nature 2013, 504, 451–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thorburn, A.N.; McKenzie, C.I.; Shen, S.; Stanley, D.; Macia, L.; Mason, L.J.; Roberts, L.K.; Wong, C.H.Y.; Shim, R.; et al. Evidence that asthma is a developmental origin disease influenced by maternal diet and bacterial metabolites. Nat. Commun. 2015, 6, 7320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nugent, B.M.; Bale, T.L. The omniscient placenta: Metabolic and epigenetic regulation of fetal programming. Front. Neuroendocrinol. 2015, 39, 28–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panchenko, P.E.; Voisin, S.; Jouin, M.; Jouneau, L.; Prezelin, A.; Lecoutre, S.; Breton, C.; Jammes, H.; Junien, C.; Gabory, A. Expression of epigenetic machinery genes is sensitive to maternal obesity and weight loss in relation to fetal growth in mice. Clin. Epigenet. 2016, 8, 22. [Google Scholar] [CrossRef] [PubMed]
- Ramaiyan, B.; Talahalli, R.R. Dietary Unsaturated Fatty Acids Modulate Maternal Dyslipidemia-Induced DNA Methylation and Histone Acetylation in Placenta and Fetal Liver in Rats. Lipids 2018, 53, 581–588. [Google Scholar] [CrossRef] [PubMed]
- Slaats, G.G.; Reinius, L.E.; Alm, J.; Kere, J.; Scheynius, A.; Joerink, M. DNA methylation levels within the CD14 promoter region are lower in placentas of mothers living on a farm. Allergy 2012, 67, 895–903. [Google Scholar] [CrossRef] [PubMed]
- Stenius, F.; Swartz, J.; Lilja, G.; Borres, M.; Bottai, M.; Pershagen, G.; Scheynius, A.; Alm, J. Lifestyle factors and sensitization in children - the ALADDIN birth cohort. Allergy 2011, 66, 1330–1338. [Google Scholar] [CrossRef] [PubMed]
- Hinz, D.; Bauer, M.; Roder, S.; Olek, S.; Huehn, J.; Sack, U.; Borte, M.; Simon, J.C.; Lehmann, I.; Herberth, G. Cord blood Tregs with stable FOXP3 expression are influenced by prenatal environment and associated with atopic dermatitis at the age of one year. Allergy 2012, 67, 380–389. [Google Scholar] [CrossRef] [PubMed]
- Luccarini, I.; Grossi, C.; Rigacci, S.; Coppi, E.; Pugliese, A.M.; Pantano, D.; la Marca, G.; Ed Dami, T.; Berti, A.; Stefani, M.; et al. Oleuropein aglycone protects against pyroglutamylated-3 amyloid-ss toxicity: Biochemical, epigenetic and functional correlates. Neurobiol. Aging 2015, 36, 648–663. [Google Scholar] [CrossRef] [PubMed]
- Bonvino, N.P.; Ray, N.B.; Luu, V.T.; Liang, J.; Hung, A.; Karagiannis, T.C. Exploration of mechanisms in nutriepigenomics: Identification of chromatin-modifying compounds from Olea Europaea. Hell. J. Nucl. Med. 2015, 18, 51–62. [Google Scholar] [PubMed]
- Serra, G.; Incani, A.; Serreli, G.; Porru, L.; Melis, M.P.; Tuberoso, C.I.G.; Rossin, D.; Biasi, F.; Deiana, M. Olive oil polyphenols reduce oxysterols -induced redox imbalance and pro-inflammatory response in intestinal cells. Redox Biol. 2018, 17, 348–354. [Google Scholar] [CrossRef] [PubMed]
- Lucas, L.; Russell, A.; Keast, R. Molecular mechanisms of inflammation. Anti-inflammatory benefits of virgin olive oil and the phenolic compound oleocanthal. Curr. Pharm. Des. 2011, 17, 754–768. [Google Scholar] [CrossRef] [PubMed]
- Rigacci, S.; Stefani, M. Nutraceutical Properties of Olive Oil Polyphenols. An Itinerary from Cultured Cells through Animal Models to Humans. Int. J. Mol. Sci. 2016, 17, 843. [Google Scholar] [CrossRef] [PubMed]
- Burdge, G.C.; Lillycrop, K.A. Fatty acids and epigenetics. Curr. Opin. Clin. Nutr. Metab. Care 2014, 17, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Goncalves, L.K.; da Silva, I.R.V.; Cechinel, L.R.; Frusciante, M.R.; de Mello, A.S.; Elsner, V.R.; Funchal, C.; Dani, C. Maternal consumption of high-fat diet and grape juice modulates global histone H4 acetylation levels in offspring hippocampus: A preliminary study. Neurosci. Lett. 2017, 661, 29–32. [Google Scholar] [CrossRef] [PubMed]
- Tarrade, A.; Panchenko, P.; Junien, C.; Gabory, A. Placental contribution to nutritional programming of health and diseases: Epigenetics and sexual dimorphism. J. Exp. Biol. 2015, 218, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Gabory, A.; Ferry, L.; Fajardy, I.; Jouneau, L.; Gothie, J.D.; Vigé, A.; Fleur, C.; Mayeur, S.; Gallou-Kabani, C.; Gross, M.S.; et al. Maternal diets trigger sex-specific divergent trajectories of gene expression and epigenetic systems in mouse placenta. PLoS ONE 2012, 7, e47986. [Google Scholar] [CrossRef] [PubMed]
- Kelly, D.; Kotliar, M.; Woo, V.; Jagannathan, S.; Whitt, J.; Moncivaiz, J.; Aronow, B.J.; Dubinsky, M.C.; Hyams, J.S.; Markowitz, J.F.; et al. Microbiota-sensitive epigenetic signature predicts inflammation in Crohn’s disease. JCI Insight 2018, 3, e122104. [Google Scholar] [CrossRef] [PubMed]
- Deckert, J.; Struhl, K. Histone acetylation at promoters is differentially affected by specific activators and repressors. Mol. Cell. Biol. 2001, 21, 2726–2735. [Google Scholar] [CrossRef] [PubMed]
- Harb, H.; Amarasekera, M.; Ashley, S.; Tulic, M.K.; Pfefferle, P.I.; Potaczek, D.P.; Martino, D.; Kesper, D.A.; Prescott, S.L.; ·Renz, H. Epigenetic Regulation in Early Childhood: A Miniaturized and Validated Method to Assess Histone Acetylation. Int. Arch. Allergy Immunol. 2015, 168, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Haring, M.; Offermann, S.; Danker, T.; Horst, I.; Peterhansel, C.; Stam, M. Chromatin immunoprecipitation: Optimization, quantitative analysis and data normalization. Plant Methods 2007, 3, 11. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Characteristic | Non-Anthroposophic (N = 43) | Anthroposophic + Partly Anthroposophic (N = 25 + 105) | p-Value * |
|---|---|---|---|
| Maternal age (years) | 30 (28–33) | 31 (27–34) | 0.46 |
| Parity | |||
| first | 17/43 (40%) | 53/128 (41%) | 0.97 |
| second | 18/43 (42%) | 49/128 (38%) | 0.81 |
| third or more | 8/43 (19%) | 26/128 (20%) | 0.98 |
| Mother vegetarian diet during pregnancy | 2/38 (5%) | 22/126 (17%) | 0.11 |
| Mother smoking during pregnancy | 8/42 (19%) | 9/127 (7%) | 0.05 |
| Father smoking during pregnancy | 11/41 (27%) | 31/125 (25%) | 0.96 |
| Female offspring | 28/43 (65%) | 65/130 (50%) | 0.12 |
| Birth weight (gram) | 3510 (3312–4010) | 3568 (3348–3939) | 0.64 |
| Gestational age in weeks | 39 (38–40) | 40 (39–41) | 0.01 |
| Predictor: Olive Oil (Yes) | β (95% CI), p-Value (Crude) | β (95% CI), p-Value (Adjusted by Offspring Gender) |
|---|---|---|
| H3 FOXP3 | 0.26 (0.08–0.43), p = 0.004 | 0.21 (0.01–0.41), p = 0.03 |
| H3 IL10RA | 0.31 (0.09–0.54), p = 0.008 | 0.31 (0.08–0.54), p = 0.008 |
| H3 IL7R | 0.36 (0.10–0.62), p = 0.007 | 0.36 (0.10–0.61), p = 0.006 |
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| FOXP3 | ATCGTGAGGATGGATGCATTAATA | CCACTGGGAAGGTCCCTAGC |
| IL10RA | GCAACTACCTCCTCCCCATT | GCCTTCGGATCAAAGTGGTC |
| IL7R | AACCCCGTCTCCACTGAAAA | GAGTCTTGCTTTGTTGCCCA |
| CD14 | ATCAGGGTTCACAGAGGA | GACCCCAAGACCCTACAC |
| RPL32 | GGAAGTGCTTGCCTTTTTCC | GGATTGCCACGGATTAACAC |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Acevedo, N.; Frumento, P.; Harb, H.; Alashkar Alhamwe, B.; Johansson, C.; Eick, L.; Alm, J.; Renz, H.; Scheynius, A.; Potaczek, D.P. Histone Acetylation of Immune Regulatory Genes in Human Placenta in Association with Maternal Intake of Olive Oil and Fish Consumption. Int. J. Mol. Sci. 2019, 20, 1060. https://doi.org/10.3390/ijms20051060
Acevedo N, Frumento P, Harb H, Alashkar Alhamwe B, Johansson C, Eick L, Alm J, Renz H, Scheynius A, Potaczek DP. Histone Acetylation of Immune Regulatory Genes in Human Placenta in Association with Maternal Intake of Olive Oil and Fish Consumption. International Journal of Molecular Sciences. 2019; 20(5):1060. https://doi.org/10.3390/ijms20051060
Chicago/Turabian StyleAcevedo, Nathalie, Paolo Frumento, Hani Harb, Bilal Alashkar Alhamwe, Catharina Johansson, Lisa Eick, Johan Alm, Harald Renz, Annika Scheynius, and Daniel P. Potaczek. 2019. "Histone Acetylation of Immune Regulatory Genes in Human Placenta in Association with Maternal Intake of Olive Oil and Fish Consumption" International Journal of Molecular Sciences 20, no. 5: 1060. https://doi.org/10.3390/ijms20051060
APA StyleAcevedo, N., Frumento, P., Harb, H., Alashkar Alhamwe, B., Johansson, C., Eick, L., Alm, J., Renz, H., Scheynius, A., & Potaczek, D. P. (2019). Histone Acetylation of Immune Regulatory Genes in Human Placenta in Association with Maternal Intake of Olive Oil and Fish Consumption. International Journal of Molecular Sciences, 20(5), 1060. https://doi.org/10.3390/ijms20051060