Muscle-Saturated Bioactive Lipids Are Increased with Aging and Influenced by High-Intensity Interval Training

 ,
,
Abstract
:1. Introduction
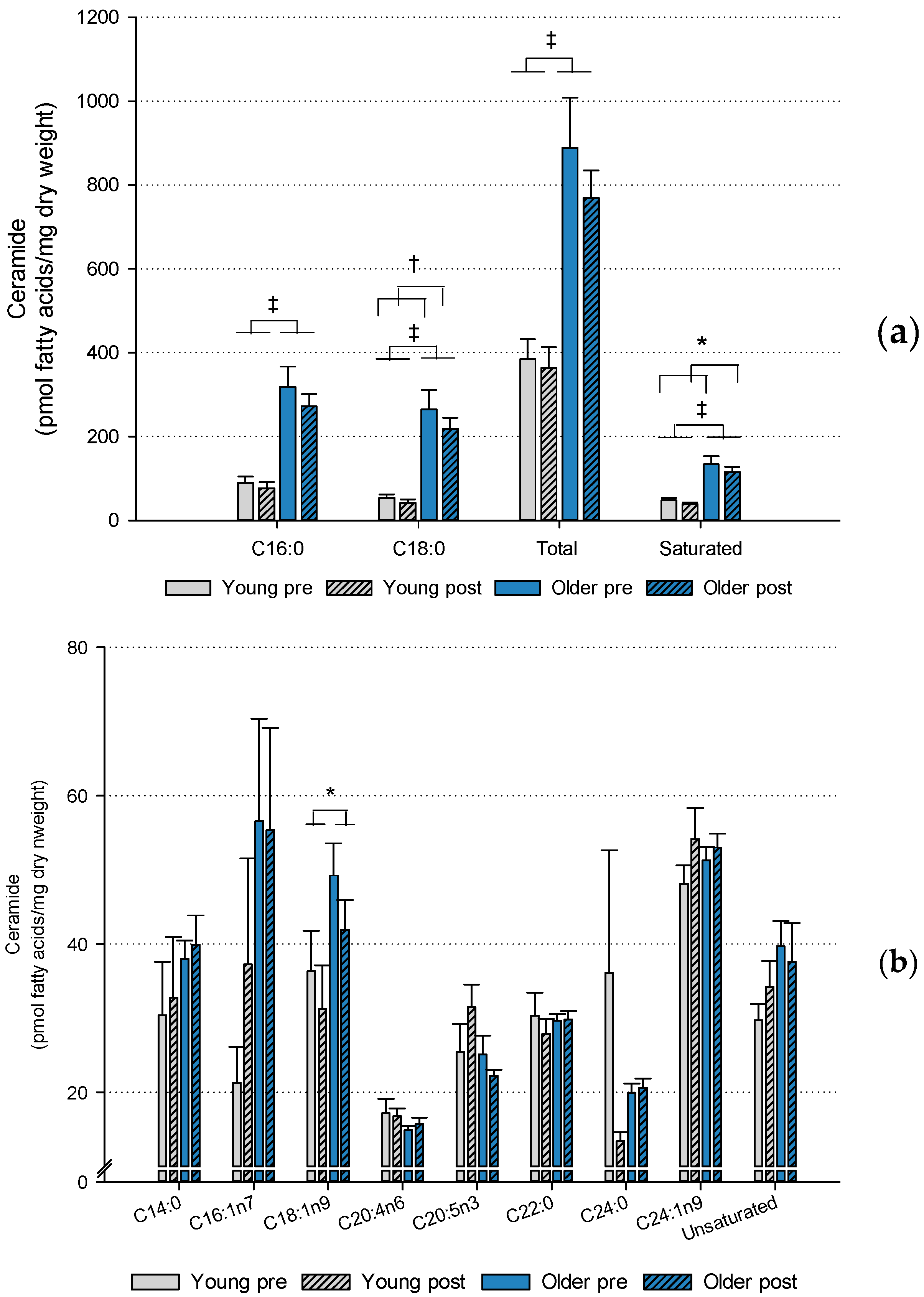
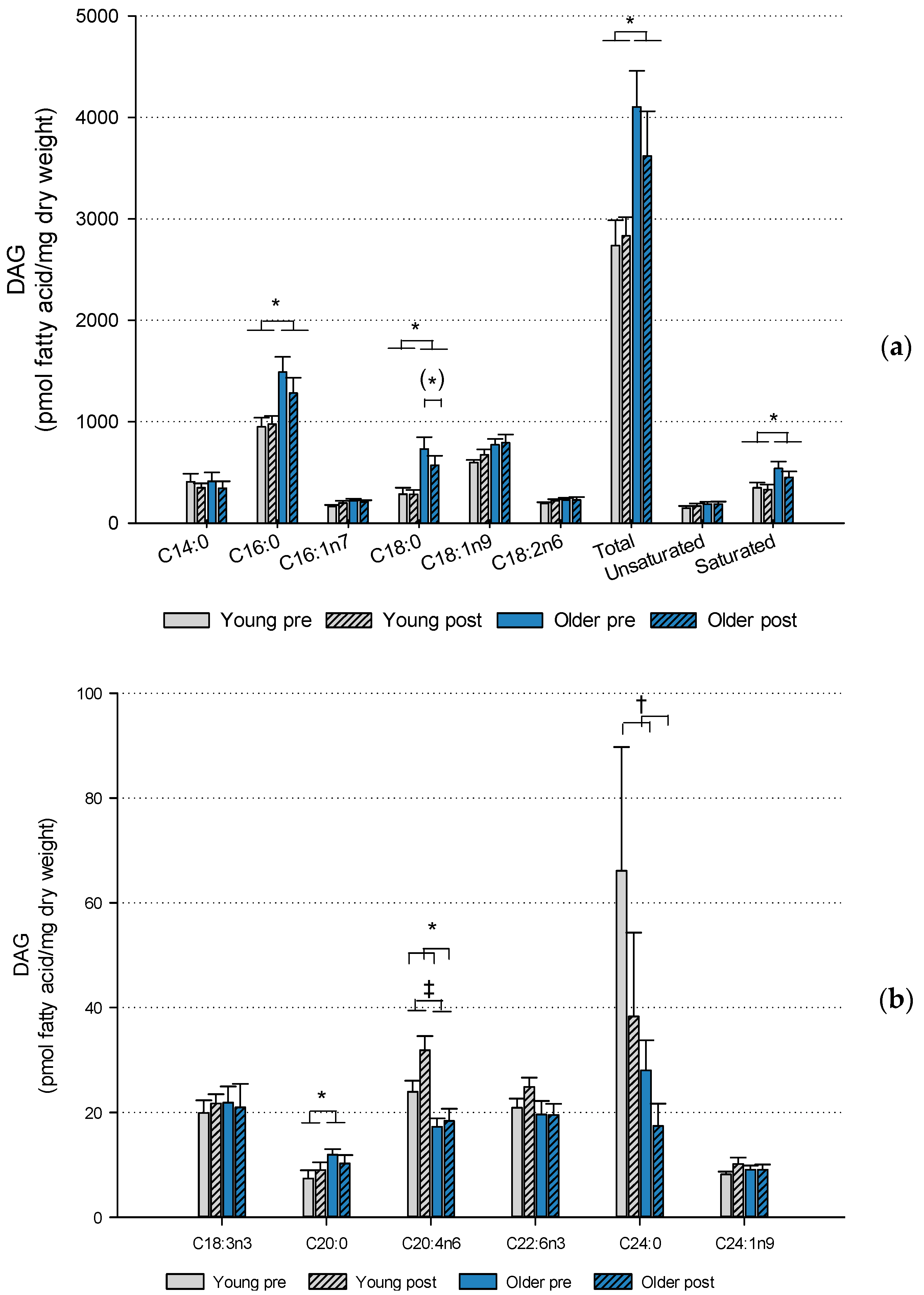
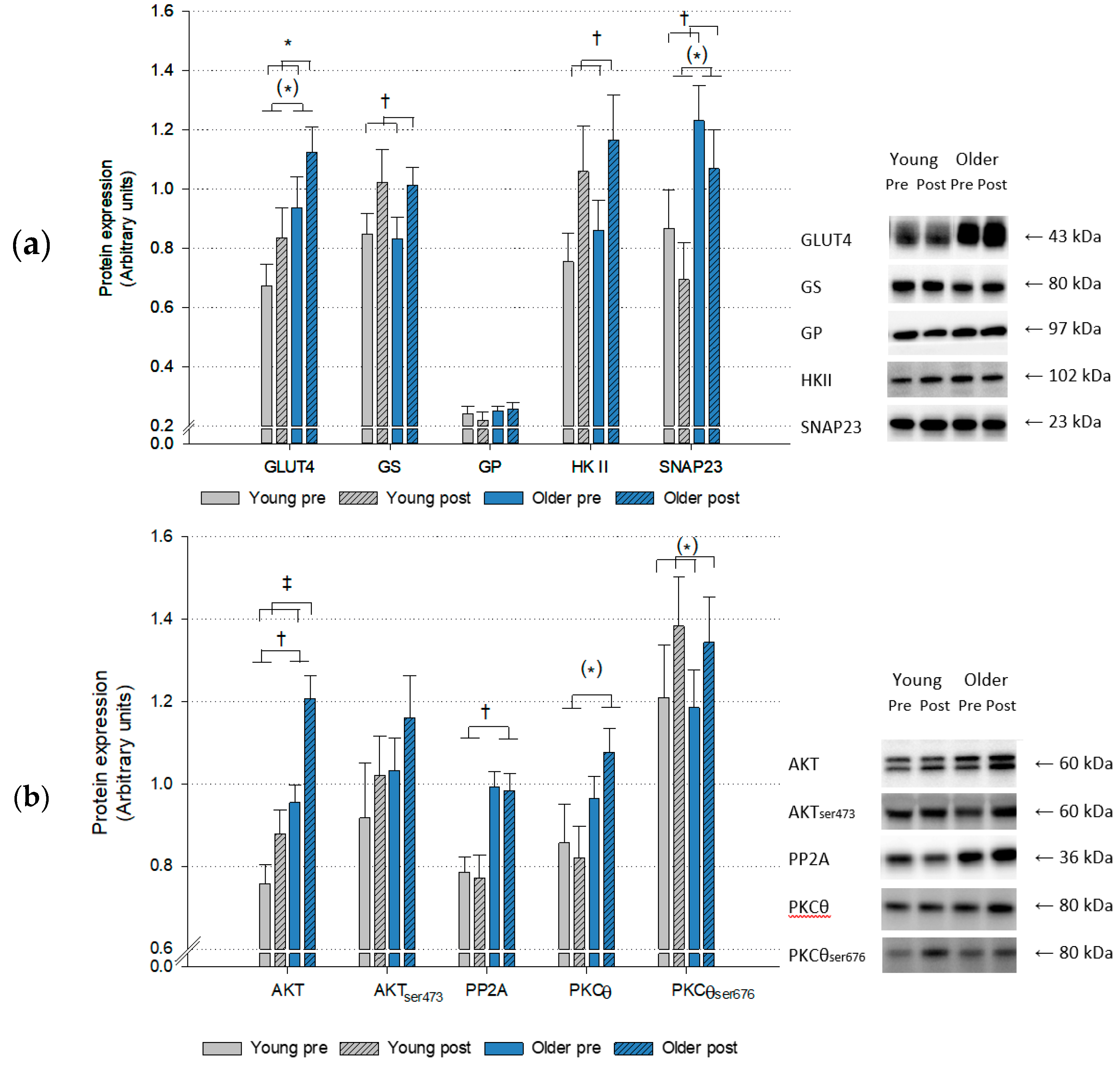
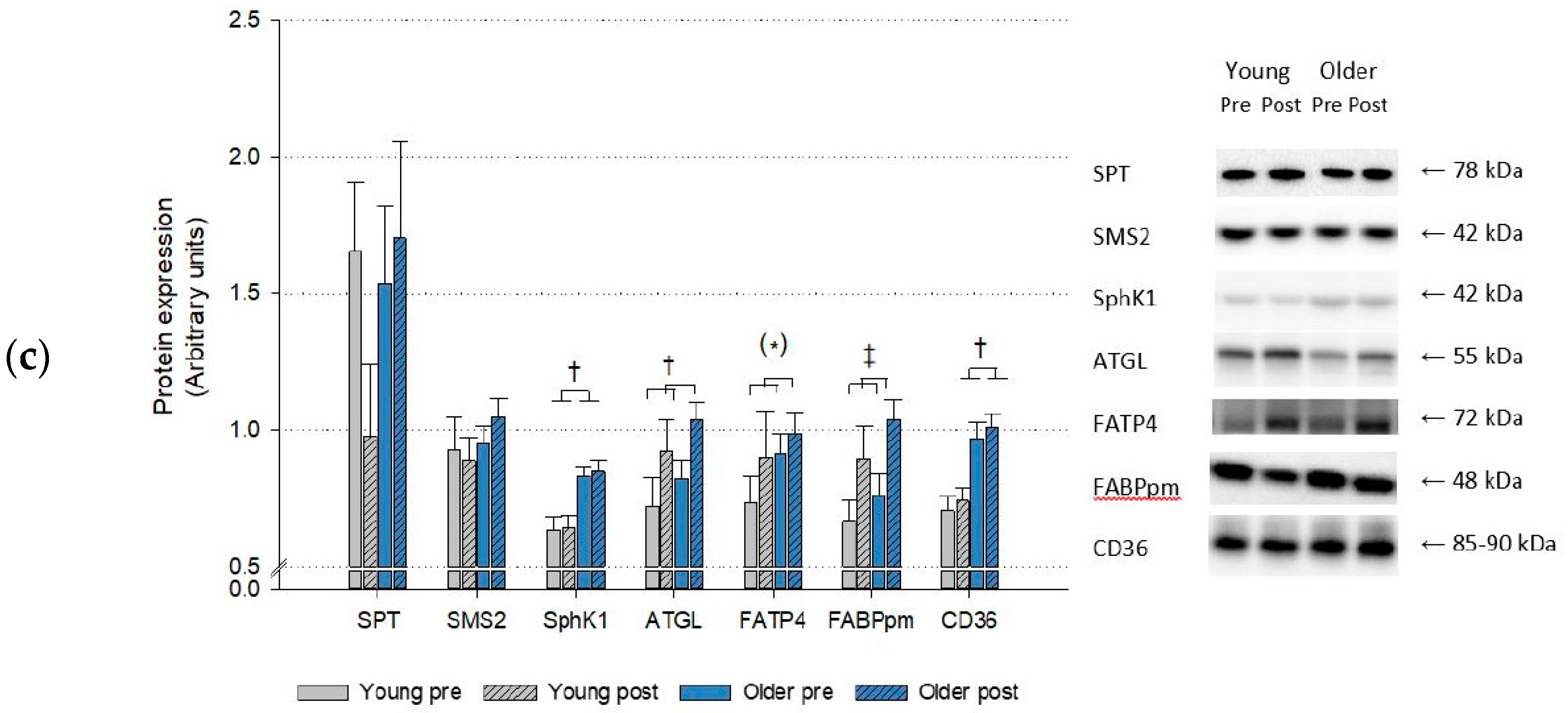
2. Results
2.1. Subject Characteristics
2.2. Muscle Lipids and Glycogen
2.3. Protein Expression
3. Discussion
Limitations
4. Methods and Materials
4.1. Subjects
4.2. Study Design
4.3. High-Intensity Interval Training (HIIT) Protocol
4.4. Maximal Oxygen Uptake
4.5. Blood Analyses
4.6. Muscle Biopsies
4.7. Lipid Analyses
4.8. Intramyocellular Triglyceride and Glycogen
4.9. Citrate Synthase and β-Hydroxyacyl-CoA Dehydrogenase
4.10. Western Blot
4.11. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dela, F.; Mikines, K.J.; Larsen, J.J.; Glabo, H. Training-induced enhancement of insulin action in human skeletal muscle: The influence of aging. J. Gerontol. A Biol. Sci. Med. Sci. 1996, 51, B247–B252. [Google Scholar] [CrossRef]
- Goodpaster, B.H.; Park, S.W.; Harris, T.B.; Kritschevsky, S.B.; Nevitt, M.; Schwarts, A.V.; Simonsick, E.M.; Tylavsky, A.V.; Visser, M.; Newman, A.B. The loss of skeletal muscle strength, mass, and quality in older adults: The health, aging and body composition study. J. Gerontol. A Biol. Sci. Med. Sci. 2006, 61, 1059–1064. [Google Scholar] [CrossRef] [PubMed]
- Bunprajun, T.; Henriksen, T.I.; Scheele, C.; Pedersen, B.K.; Green, C.J. Lifelong Physical Activity Prevents Aging-Associated Insulin Resistance in Human Skeletal Muscle Myotubes via Increased Glucose Transporter Expression. PLoS ONE 2013, 8, e66628. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.C.; Glickman, S.G.; Dengel, D.R.; Brown, M.D.; Supiano, M.A. Abdominal adiposity assessed by dual energy X-ray absorptiometry provides a sex-independent predictor of insulin sensitivity in older adults. J. Gerontol. A Biol. Sci. Med. Sci. 2005, 60, 872–877. [Google Scholar] [CrossRef] [PubMed]
- Hughes, V.A.; Roubenoff, R.; Wood, M.; Frontera, W.R.; Evans, W.J.; Fiatarone Singh, M.A. Anthropometric assessment of 10-y changes in body composition in the elderly. Am. J. Clin. Nutr. 2004, 80, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Kohrt, W.M.; Kirwan, J.P.; Staten, M.A.; Bourey, R.E.; Kings, D.S.; Holloszy, J.S. Insulin resistance in aging is related to abdominal obesity. Diabetes 1993, 42, 273–281. [Google Scholar] [CrossRef]
- Lalia, A.Z.; Dazari, S.; Johnson, M.L.; Robinson, M.M.; Konopka, A.R.; Distelmaier, K.; Port, J.D.; Glavin, M.T.; Esponda, R.R.; Nair, K.S.; et al. Predictors of Whole-Body Insulin Sensitivity Across Ages and Adiposity in Adult Humans. J. Clin. Endocrinol. Metab. 2016, 101, 626–634. [Google Scholar] [CrossRef]
- Petersen, M.C.; Jurczak, M.J. CrossTalk opposing view: Intramyocellular ceramide accumulation does not modulate insulin resistance. J. Physiol. 2016, 594, 3171–3174. [Google Scholar] [CrossRef] [Green Version]
- Summers, S.A.; Goodpaster, B.H. CrossTalk proposal: Intramyocellular ceramide accumulation does modulate insulin resistance. J. Physiol. 2016, 594, 3167–3170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montell, E.; Turini, M.; Marotta, M.; Roberts, M.; Noe, V.; Ciudad, C.J.; Mace, K.; Gomez-Foix, A.M. DAG accumulation from saturated fatty acids desensitizes insulin stimulation of glucose uptake in muscle cells. Am. J. Physiol. Endocrinol. Metab. 2001, 280, E229–E237. [Google Scholar] [CrossRef] [PubMed]
- Stratford, S.; Hoehn, K.L.; Lui, F.; Summers, S.A. Regulation of insulin action by ceramide: Dual mechanisms linking ceramide accumulation to the inhibition of Akt/protein kinase B. J. Biol. Chem. 2004, 279, 36608–36615. [Google Scholar] [CrossRef]
- Griffin, M.E.; Marcucci, M.J.; Cline, G.W.; Bell, K.; Barruci, N.; Lee, D.; Goodyear, L.J.; Kraegen, E.W.; White, M.F.; Shulman, G.I. Free fatty acid-induced insulin resistance is associated with activation of protein kinase C theta and alterations in the insulin signaling cascade. Diabetes 1999, 48, 1270–1274. [Google Scholar] [CrossRef] [PubMed]
- Straczkowski, M.; Kowalska, I.; Nikolajuk, A.; Dzienis-Straczkowska, S.; Kinalska, I.; Baranowski, M.; Zendzian-Piotrowska, M.; Brzezinska, Z.; Gorski, J. Relationship between insulin sensitivity and sphingomyelin signaling pathway in human skeletal muscle. Diabetes 2004, 53, 1215–1221. [Google Scholar] [CrossRef] [PubMed]
- Bergman, B.C.; Brozinick, J.T.; Strauss, A.; Bacon, S.; Kerege, A.; Bui, H.H.; Sanders, P.; Sidall, P.; Wei, T.; Thomas, M.K.; et al. Muscle sphingolipids during rest and exercise: A C18:0 signature for insulin resistance in humans. Diabetologia 2016, 59, 785–798. [Google Scholar] [CrossRef] [PubMed]
- Chee, C.; Shannon, C.E.; Burns, A.; Selby, A.L.; Wilkinson, D.; Smith, K.; Greenhaff, P.L.; Stephens, F.B. Relative Contribution of Intramyocellular Lipid to Whole-Body Fat Oxidation Is Reduced With Age but Subsarcolemmal Lipid Accumulation and Insulin Resistance Are Only Associated With Overweight Individuals. Diabetes 2016, 65, 840–850. [Google Scholar] [CrossRef]
- Moro, C.; Galgani, J.E.; Luu, L.; Pasarica, M.; Mairal, M.; Bajpeyi, S.; Schmitz, G.; Langin, D.; Liebisch, G.; Smith, S.R. Influence of gender, obesity, and muscle lipase activity on intramyocellular lipids in sedentary individuals. J. Clin. Endocrinol. Metab. 2009, 94, 3440–3447. [Google Scholar] [CrossRef]
- Skovbro, M.; Baranowski, M.; Skov-Jensen, C.; Flint, A.; Dela, F.; Gorski, J.; Helge, J.W. Human skeletal muscle ceramide content is not a major factor in muscle insulin sensitivity. Diabetologia 2008, 51, 1253–1260. [Google Scholar] [CrossRef] [Green Version]
- Coen, P.M.; Hames, K.C.; Leachman, E.M.; DeLany, J.P.; Ritov, V.B.; Menshikova, E.V.; Dube, J.J.; Stefanovic-Racic, M.; Toledo, F.G.; Goodpaster, B.H. Reduced skeletal muscle oxidative capacity and elevated ceramide but not diacylglycerol content in severe obesity. Obesity 2013, 21, 2362–2371. [Google Scholar] [CrossRef] [Green Version]
- Straczkowski, M.; Kowalska, I.; Baranowski, M.; Nikolajuk, A.; Otziomek, E.; Zabielski, P.; Adamska, A.; Blachnio, A.; Gorski, J.; Gorska, M. Increased skeletal muscle ceramide level in men at risk of developing type 2 diabetes. Diabetologia 2007, 50, 2366–2373. [Google Scholar] [CrossRef] [Green Version]
- Sogaard, D.; Baranowski, M.; Dela, F.; Helge, J.W. The influence of age and cardiorespiratory fitness on bioactive lipids in muscle. J. Gerontol. A Biol. Sci. Med. Sci. 2018. [Google Scholar] [CrossRef]
- Rivas, D.A.; Morris, E.P.; Harran, P.H.; Pasha, E.P.; Morais Mda, S.; Dolnikowski, G.G.; Philips, E.M.; Fielding, R.A. Increased ceramide content and NFkappaB signaling may contribute to the attenuation of anabolic signaling after resistance exercise in aged males. J. Appl. Physiol. 2012, 113, 1727–1736. [Google Scholar] [CrossRef] [PubMed]
- Dela, F.; Mikines, K.J.; Sonne, B.; Galbo, H. Effect of training on interaction between insulin and exercise in human muscle. J. Appl. Physiol. 1994, 76, 2386–2393. [Google Scholar] [CrossRef]
- Gan, S.K.; Kriketos, A.D.; Ellis, B.A.; Thompson, C.H.; Kraegen, E.W.; Chisholm, D.J. Changes in aerobic capacity and visceral fat but not myocyte lipid levels predict increased insulin action after exercise in overweight and obese men. Diabetes Care 2003, 26, 1706–1713. [Google Scholar] [CrossRef] [PubMed]
- Bruce, C.R.; Thrush, A.B.; Mertz, V.A.; Bezaire, V.; Chabowski, A.; Heigenhauser, G.J.; Dyck, D.J. Endurance training in obese humans improves glucose tolerance and mitochondrial fatty acid oxidation and alters muscle lipid content. Am. J. Physiol. Endocrinol. Metab. 2006, 291, E99–E107. [Google Scholar] [CrossRef] [PubMed]
- Fisher, G.; Brown, A.W.; Bohan Brown, M.M.; Alcorn, A.; Noles, C.; Winwood, L.; Resuehr, H.; George, B.; Jeansonne, M.M.; Allison, D.B. High Intensity Interval- vs Moderate Intensity- Training for Improving Cardiometabolic Health in Overweight or Obese Males: A Randomized Controlled Trial. PLoS ONE 2015, 10, e0138853. [Google Scholar] [CrossRef] [PubMed]
- Terada, T.; Friesen, A.; Chahal, B.S.; Bell, G.J.; McCargar, L.J.; Boule, N.G. Feasibility and preliminary efficacy of high intensity interval training in type 2 diabetes. Diabetes Res. Clin. Pract. 2013, 99, 120–129. [Google Scholar] [CrossRef]
- Martins, C.; Kasakova, I.; Ludviksen, M.; Mehus, I.; Wisloff, U.; Kulseng, B.; Morgan, L.; King, B. High-Intensity Interval Training and Isocaloric Moderate-Intensity Continuous Training Result in Similar Improvements in Body Composition and Fitness in Obese Individuals. Int. J. Sport Nutr. Exerc. Metab. 2015, 26, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Richards, J.C.; Johnson, T.K.; Kuzma, J.N.; Lonac, M.C.; Schweder, M.M.; Voyles, W.F.; Bell, C. Short-term sprint interval training increases insulin sensitivity in healthy adults but does not affect the thermogenic response to beta-adrenergic stimulation. J. Physiol. 2010, 588, 2961–2972. [Google Scholar] [CrossRef]
- Robinson, M.M.; Dasari, S.; Konopka, A.R.; Johnson, M.L.; Manjunatha, S.; Esponda, R.R.; Carter, R.E.; Lanza, I.R.; Nair, K.S. Enhanced Protein Translation Underlies Improved Metabolic and Physical Adaptations to Different Exercise Training Modes in Young and Old Humans. Cell Metab. 2017, 25, 581–592. [Google Scholar] [CrossRef] [Green Version]
- Arad, A.D.; DiMenna, F.J.; Thomas, N.; Tamis-Holland, J.; Weil, R.; Geliebter, A.; Albu, J.B. High-intensity interval training without weight loss improves exercise but not basal or insulin-induced metabolism in overweight/obese African American women. J. Appl. Physiol. 2015, 119, 352–362. [Google Scholar] [CrossRef] [Green Version]
- Bonen, A.; Parolin, M.L.; Steinberg, G.R.; Calles-Escandon, J.; Tandon, N.N.; Glatz, J.F.; Luiken, J.J.; Heigenhauser, G.J.; Dyck, D.J. Triacylglycerol accumulation in human obesity and type 2 diabetes is associated with increased rates of skeletal muscle fatty acid transport and increased sarcolemmal FAT/CD36. FASEB J. 2004, 18, 1144–1146. [Google Scholar] [CrossRef]
- Bruce, C.R.; Risis, S.; Babb, J.R.; Yang, C.; Kowalski, G.M.; Selathurai, A.; Lee-Young, R.S.; Weir, J.R.; Yoshioka, K.; Takuwa, Y.; et al. Overexpression of sphingosine kinase 1 prevents ceramide accumulation and ameliorates muscle insulin resistance in high-fat diet-fed mice. Diabetes 2012, 61, 3148–3155. [Google Scholar] [CrossRef]
- Tonks, K.T.; Coster, A.C.; Christopher, M.J.; Chaudhuri, R.; Xu, A.; Gagnon-Bartsch, J.; Chisholm, D.J.; James, D.E.; Meikle, P.J.; Greenfield, J.R.; et al. Skeletal muscle and plasma lipidomic signatures of insulin resistance and overweight/obesity in humans. Obesity 2016, 24, 908–916. [Google Scholar] [CrossRef] [Green Version]
- Bergman, B.C.; Hunerdosse, D.M.; Kerege, A.; Playdon, M.C.; Perreault, L. Localisation and composition of skeletal muscle diacylglycerol predicts insulin resistance in humans. Diabetologia 2012, 55, 1140–1150. [Google Scholar] [CrossRef] [Green Version]
- Itani, S.I.; Pories, W.J.; MacDonald, K.G.; Dohm, G.L. Increased protein kinase C theta in skeletal muscle of diabetic patients. Metabolism 2001, 50, 553–557. [Google Scholar] [CrossRef]
- Coen, P.M.; Dube, J.J.; Amati, F.; Stefanovic-Racic, M.; Ferrell, R.E.; Toledo, F.G.; Goodpaster, G.H. Insulin resistance is associated with higher intramyocellular triglycerides in type I but not type II myocytes concomitant with higher ceramide content. Diabetes 2010, 59, 80–88. [Google Scholar] [CrossRef]
- Amati, F.; Dube, J.J.; Alvarez-Carnero, E.; Edreira, M.M.; Chomentowski, P.; Coen, P.M.; Switzer, G.E.; Bickel, P.E.; Stefanovic-Racic, M.; Toledo, F.G.; et al. Skeletal muscle triglycerides, diacylglycerols, and ceramides in insulin resistance: Another paradox in endurance-trained athletes? Diabetes 2011, 60, 2588–2597. [Google Scholar] [CrossRef]
- Dube, J.J.; Amati, F.; Toledo, F.G.; Stefanovic-Racic, M.; Rossi, A.; Coen, P.; Goodpaster, B.H. Effects of weight loss and exercise on insulin resistance, and intramyocellular triacylglycerol, diacylglycerol and ceramide. Diabetologia 2011, 54, 1147–1156. [Google Scholar] [CrossRef]
- Dube, J.J.; Amati, F.; Stefanovic-Racic, M.; Toledo, F.G.; Sauers, S.E.; Goodpaster, B.H. Exercise-induced alterations in intramyocellular lipids and insulin resistance: The athlete’s paradox revisited. Am. J. Physiol. Endocrinol. Metab. 2008, 294, E882–E888. [Google Scholar] [CrossRef]
- Louche, K.; Badin, P.M.; Montastier, E.; Laurens, C.; Bourlier, V.; de Glisezinski, I.; Thalamas, C.; Viguerie, N.; Langin, D.; Moro, C. Endurance exercise training up-regulates lipolytic proteins and reduces triglyceride content in skeletal muscle of obese subjects. J. Clin. Endocrinol. Metab. 2013, 98, 4863–4871. [Google Scholar] [CrossRef]
- Devries, M.C.; Samjoo, I.A.; Hamadeh, M.J.; McCready, C.; Raha, S.; Watt, M.J.; Steinberg, G.R.; Tarnapolsky, M.A. Endurance training modulates intramyocellular lipid compartmentalization and morphology in skeletal muscle of lean and obese women. J. Clin. Endocrinol. Metab. 2013, 98, 4852–4862. [Google Scholar] [CrossRef] [PubMed]
- Coen, P.M.; Menshikova, E.V.; Distefano, G.; Zheng, D.; Tanner, C.J.; Standley, R.A.; Helbling, N.L.; Dubis, G.S.; Ritov, V.B.; Xie, H.; et al. Exercise and Weight Loss Improve Muscle Mitochondrial Respiration, Lipid Partitioning, and Insulin Sensitivity After Gastric Bypass Surgery. Diabetes 2015, 64, 3737–3750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hood, M.S.; Little, J.P.; Tarnapolsky, M.A.; Myslik, F.; Gibala, M.J. Low-volume interval training improves muscle oxidative capacity in sedentary adults. Med. Sci. Sports Exerc. 2011, 43, 1849–1856. [Google Scholar] [CrossRef] [PubMed]
- Perry, C.G.; Heigenhauser, G.J.; Bonen, A.; Spriet, L.L. High-intensity aerobic interval training increases fat and carbohydrate metabolic capacities in human skeletal muscle. Appl. Physiol. Nutr. Metab. 2008, 33, 1112–1123. [Google Scholar] [CrossRef] [PubMed]
- Burgomaster, K.A.; Cermak, N.M.; Philips, S.M.; Benton, C.R.; Bonen, A.; Gibala, M.J. Divergent response of metabolite transport proteins in human skeletal muscle after sprint interval training and detraining. Am. J. Physiol. Regul. Integr. Comp Physiol. 2007, 292, R1970–R1976. [Google Scholar] [CrossRef] [PubMed]
- Shaban, N.; Kenno, K.A.; Milne, K.J. The effects of a 2 week modified high intensity interval training program on the homeostatic model of insulin resistance (HOMA-IR) in adults with type 2 diabetes. J. Sports Med. Phys. Fitness 2014, 54, 203–209. [Google Scholar] [PubMed]
- Skleryk, J.R.; Karagounis, L.G.; Hawley, J.A.; Sharman, M.J.; Laursen, P.B.; Watson, G. Two weeks of reduced-volume sprint interval or traditional exercise training does not improve metabolic functioning in sedentary obese men. Diabetes Obes. Metab. 2013, 15, 1146–1153. [Google Scholar] [CrossRef] [PubMed]
- Sogaard, D.; Lund, M.T.; Scheuer, C.M.; Dehlbaek, M.S.; Dideriksen, S.G.; Abildskov, C.V.; Christensen, K.K.; Dohlmann, T.L.; Larsen, S.; Vigelso, A.H.; et al. High-intensity interval training improves insulin sensitivity in older individuals. Acta Physiol. 2018, 222, e13009. [Google Scholar] [CrossRef] [PubMed]
- Bergstrom, J. Muscle-biopsy needles. Lancet 1979, 1, 153. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Stanley, G.H.S. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar]
- Yano, M.; Kishida, E.; Muneyuki, Y.; Masusawa, Y. Quantitative analysis of ceramide molecular species by high performance liquid chromatography. J. Lipid Res. 1998, 39, 2091–2098. [Google Scholar] [PubMed]
- Nawrocki, A.; Gorski, J. Effect of plasma free fatty acid concentration on the content and composition of the free fatty acid fraction in rat skeletal muscles. Horm. Metab. Res. 2004, 36, 601–606. [Google Scholar] [CrossRef] [PubMed]
- Frayn, K.N.; Maycock, P.F. Skeletal muscle triacylglycerol in the rat: Methods for sampling and measurement, and studies of biological variability. J. Lipid Res. 1980, 21, 139–144. [Google Scholar] [PubMed]
- Kiens, B.; Richter, E.A. Types of carbohydrate in an ordinary diet affect insulin action and muscle substrates in humans. Am. J. Clin. Nutr. 1996, 63, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Larsen, S.; Danielsen, J.H.; Sondergard, S.D.; Sogaard, D.; Vigelsoe, A.; Dybboe, R.; Skaaby, S.; Dela, F.; Helge, J.W. The effect of high-intensity training on mitochondrial fat oxidation in skeletal muscle and subcutaneous adipose tissue. Scand. J. Med. Sci. Sports 2015, 25, e59–e69. [Google Scholar] [CrossRef] [PubMed]
- Sogaard, D.; Ostergard, T.; Blachnio-Zabielska, A.U.; Baranowski, M.; Vigelso, A.H.; Andersen, J.L.; Dela, F.; Helge, J.W. Training Does Not Alter Muscle Ceramide and Diacylglycerol in Offsprings of Type 2 Diabetic Patients Despite Improved Insulin Sensitivity. J. Diabetes Res. 2016, 2016, 2372741. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Young (n = 14) Pre Post | Older (n = 22) Pre Post | Main Effect (p-value) Age Time | Interaction (p-value) Group x Time | ||||
|---|---|---|---|---|---|---|---|
| Gender (F/M) | 5/9 | 11/11 | |||||
| Age (yrs) | 32 ± 2 | 63 ± 1 | |||||
| Height (m) | 1.78 ± 0.02 | 1.70 ± 0.02 | 0.014 | NS | NS | ||
| Weight (kg) | 110 ± 4 | 110 ± 4 | 88.7 ± 2.6 | 88.4 ± 2.6 | <0.001 | NS | NS |
| BMI (kg·m−2) | 34.8 ± 1.0 | 34.6 ± 1.0 | 30.7 ± 0.7 | 30.6 ± 0.7 | 0.003 | NS | NS |
| LBM (kg) | 63.8 ± 2.1 | 64.7 ± 2.3 | 51.5 ± 2.1 | 51.8 ± 2.1 | <0.001 | <0.001 | 0.099 |
| Fat mass (kg) | 40.3 ± 3.1 | 39.3 ± 3.3 | 34.0 ± 1.6 | 33.3 ± 1.7 | NS | 0.016 | NS |
| Fat % | 39.2 ± 2.1 | 38.2 ± 2.3 | 39.8 ± 1.6 | 39.1 ± 1.6 | NS | <0.001 | NS |
| Visceral fat (kg) | 1.67 ± 0.25 | 1.56 ± 0.24 | 1.90 ± 0.16 | 1.81 ± 0.16 | NS | 0.024 | NS |
| HbA1c (%) | 5.3 ± 0.1 | 5.3 ± 0.1 | 5.7 ± 0.1 | 5.6 ± 0.1 | 0.002 | NS | NS |
| HOMA-IR (AU) | 2.14 ± 0.24 | 2.31 ± 0.38 | 1.88 ± 0.23 | 1.99 ± 0.30 | NS | NS | NS |
| Glucose, fasting (mmol·L−1) | 4.5 ± 0.1 | 4.5 ± 0.1 | 6.1 ± 0.2 | 6.0 ± 0.2 | <0.001 | NS | NS |
| Insulin, fasting (pmol L−1) | 69.7 ± 9,5 | 67.2 ± 8.9 | 40.9 ± 4.8 | 42.6 ± 5.8 | 0.008 | NS | NS |
| IMTG (mmol·kg−1 dw) | 126 ± 27 | 118 ± 21 | 156 ± 23 | 119 ± 12 | NS | NS | NS |
| Glycogen (nmol·kg−1 dw) | 236 ± 30 | 474 ± 46 | 323 ± 24 | 483 ± 23 | NS | <0.001 | NS |
| HAD (µmol·g−1·min−1) | 116 ± 7 | 130 ± 7 | 112 ± 10 | 141 ± 5 | NS | <0.001 | NS |
| CS (µmol·g−1·min−1) | 132 ± 7 | 165 ± 8 | 122 ± 10 | 169 ± 10 | NS | <0.001 | NS |
| VO2max (mL·min−1) | 3068 ± 131 | 3186 ± 118 | 2234 ± 106 | 2361 ± 134 | <0.001 | 0.021 | NS |
| VO2max (mL·min−1·kg−1) | 28.3 ± 1.2 | 29.7 ± 1.5 | 25.2 ± 1.0 | 26.7 ± 1.1 | NS | 0.007 | NS |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Søgaard, D.; Baranowski, M.; Larsen, S.; Taulo Lund, M.; Munk Scheuer, C.; Vestergaard Abildskov, C.; Greve Dideriksen, S.; Dela, F.; Wulff Helge, J. Muscle-Saturated Bioactive Lipids Are Increased with Aging and Influenced by High-Intensity Interval Training. Int. J. Mol. Sci. 2019, 20, 1240. https://doi.org/10.3390/ijms20051240
Søgaard D, Baranowski M, Larsen S, Taulo Lund M, Munk Scheuer C, Vestergaard Abildskov C, Greve Dideriksen S, Dela F, Wulff Helge J. Muscle-Saturated Bioactive Lipids Are Increased with Aging and Influenced by High-Intensity Interval Training. International Journal of Molecular Sciences. 2019; 20(5):1240. https://doi.org/10.3390/ijms20051240
Chicago/Turabian StyleSøgaard, Ditte, Marcin Baranowski, Steen Larsen, Michael Taulo Lund, Cathrine Munk Scheuer, Carina Vestergaard Abildskov, Sofie Greve Dideriksen, Flemming Dela, and Jørn Wulff Helge. 2019. "Muscle-Saturated Bioactive Lipids Are Increased with Aging and Influenced by High-Intensity Interval Training" International Journal of Molecular Sciences 20, no. 5: 1240. https://doi.org/10.3390/ijms20051240
APA StyleSøgaard, D., Baranowski, M., Larsen, S., Taulo Lund, M., Munk Scheuer, C., Vestergaard Abildskov, C., Greve Dideriksen, S., Dela, F., & Wulff Helge, J. (2019). Muscle-Saturated Bioactive Lipids Are Increased with Aging and Influenced by High-Intensity Interval Training. International Journal of Molecular Sciences, 20(5), 1240. https://doi.org/10.3390/ijms20051240