Modulation of Iberian Ribbed Newt Complement Component C3 by Stressors Similar to those Encountered during a Stay Onboard the International Space Station

 , ,
, ,
Abstract
:1. Introduction
2. Results
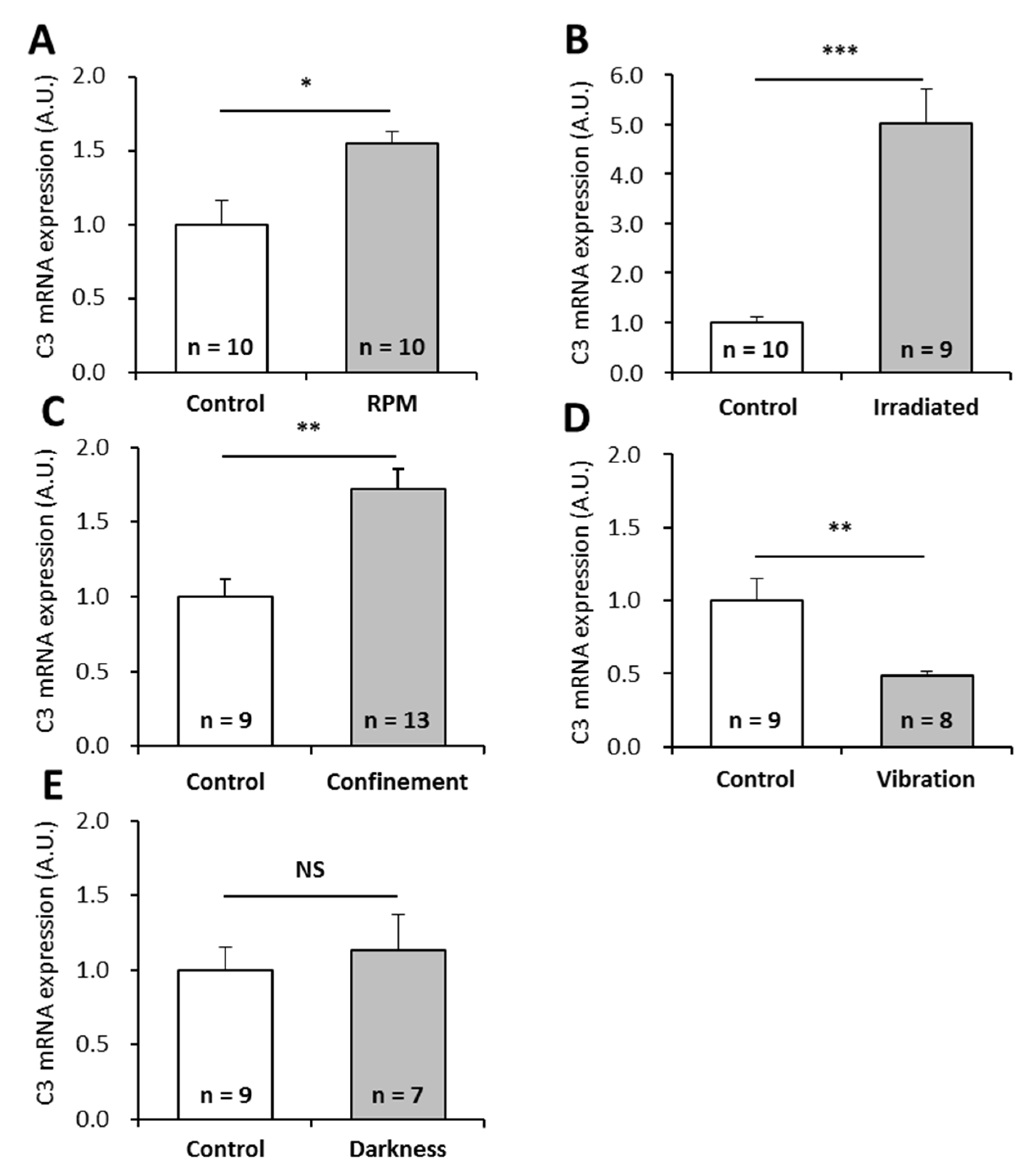
2.1. Effects of Simulated Microgravity, Circadian Rhythm Modification, Confinement, Vibration, and Radiation on P. waltl C3 Transcription
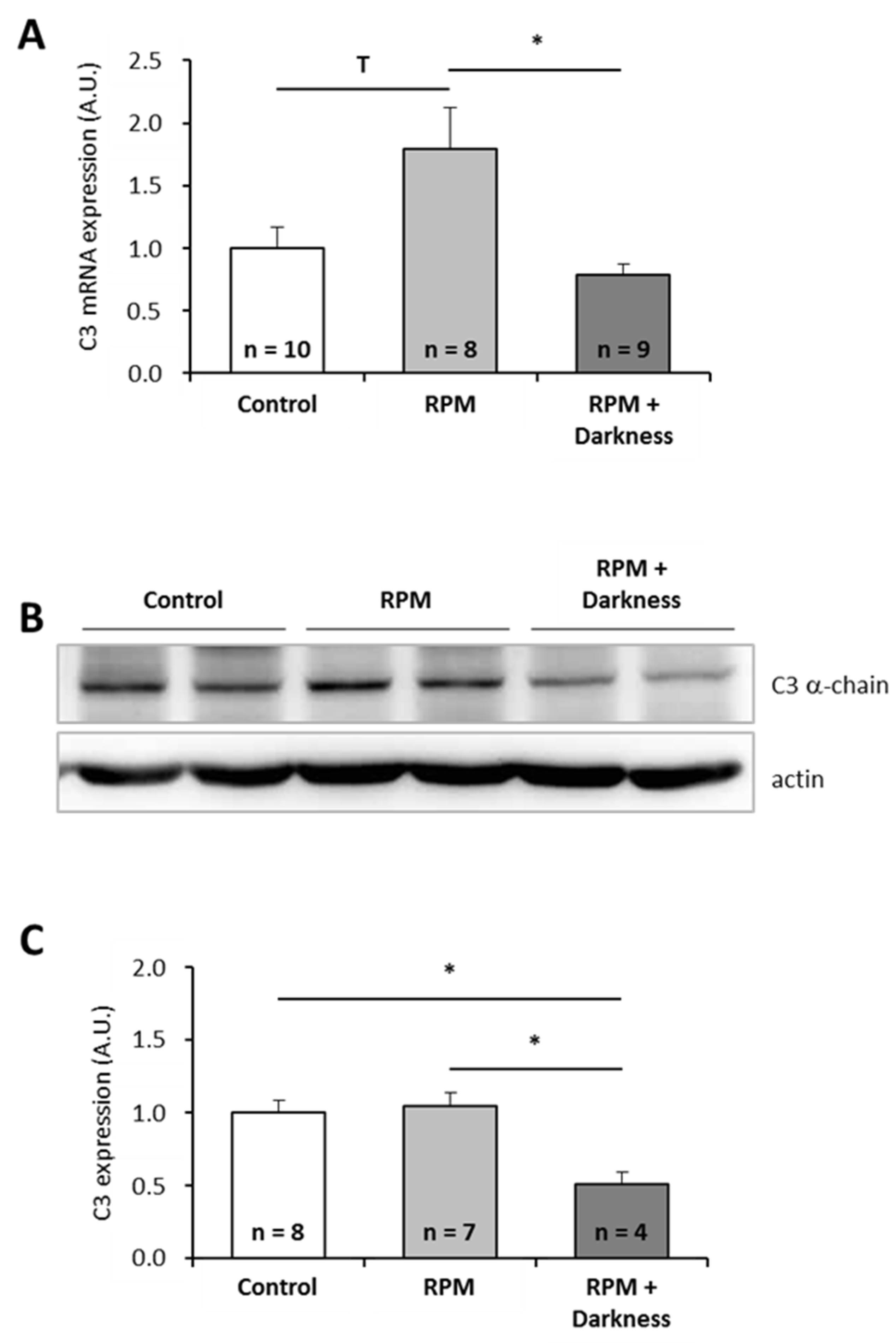
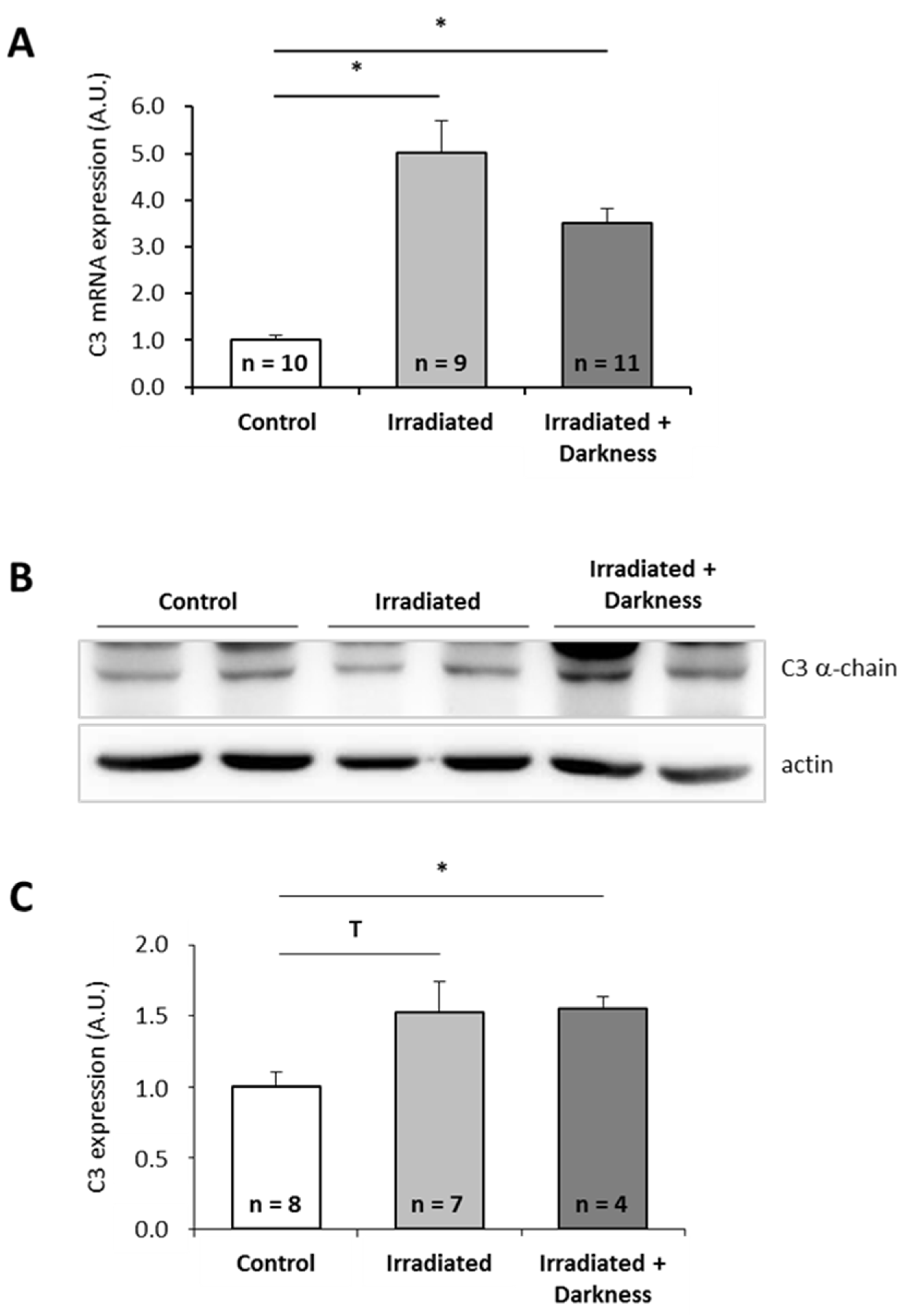
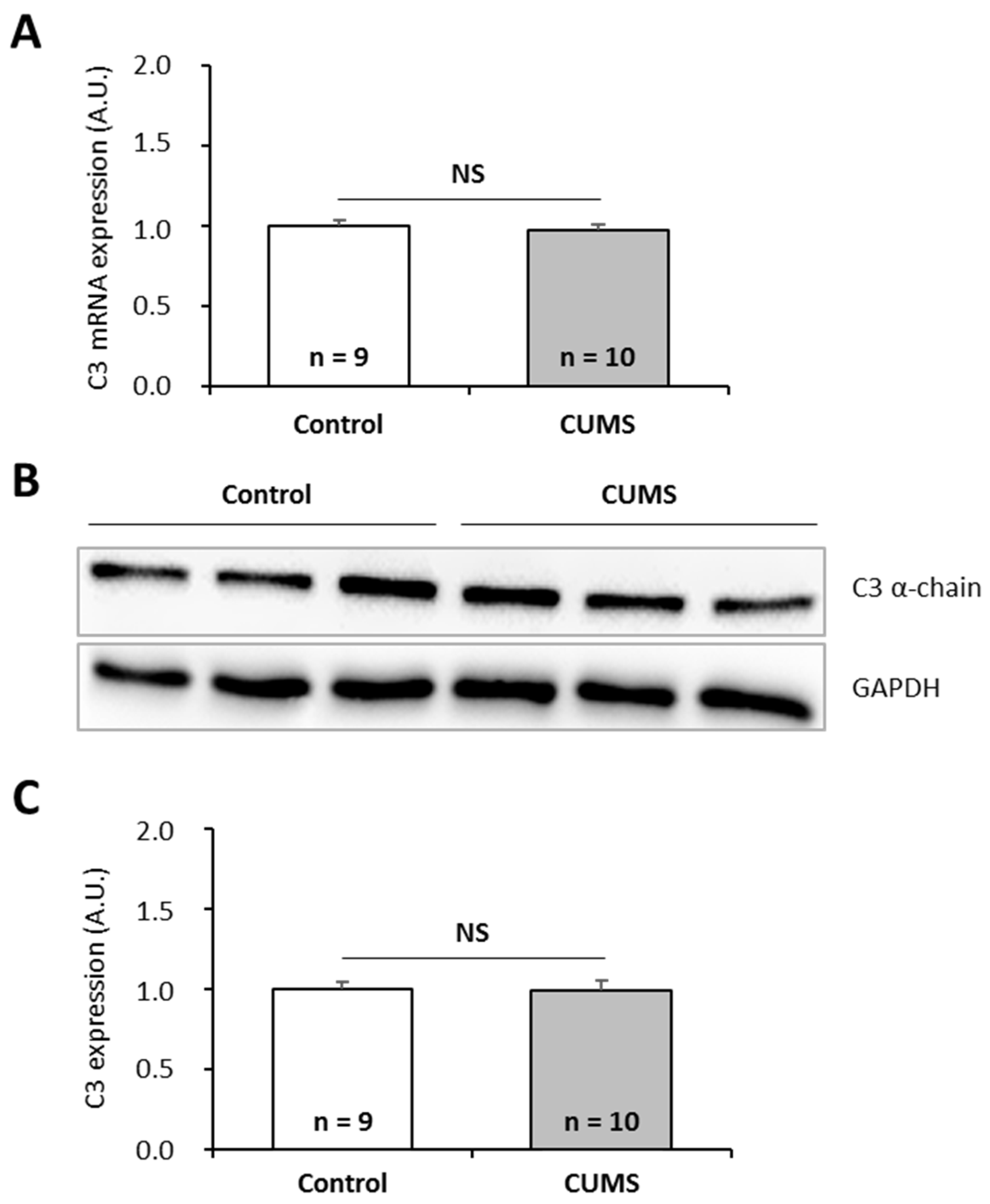
2.2. Effects of Two Combinations of Stressors on P. waltl C3 Expression
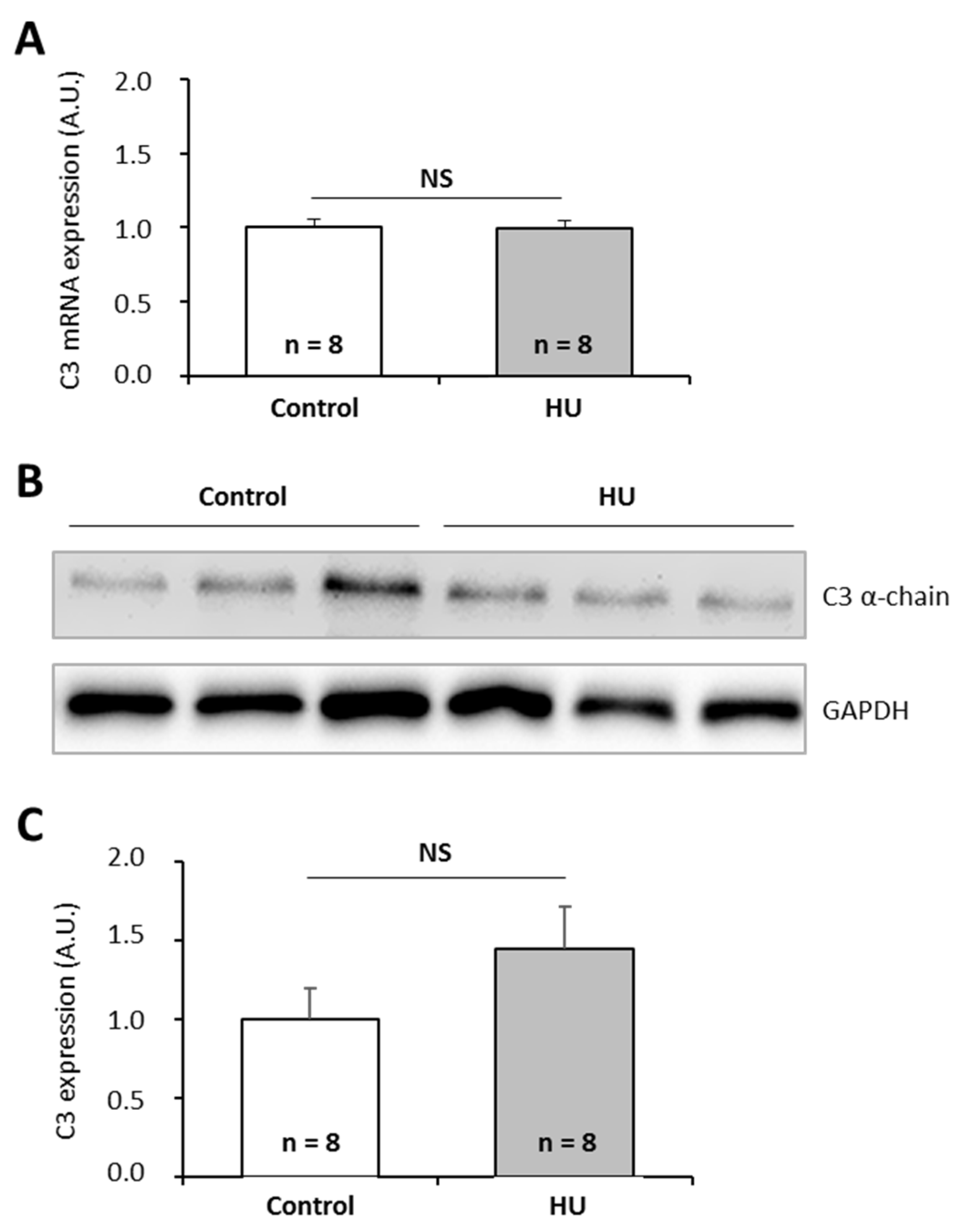
2.3. Effects of Spaceflight Stressors on Murine C3 Expression
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Simulation of Microgravity Using the RPM
4.3. Circadian Rhythm Perturbation
4.4. Simulation of Space Radiation
4.5. Confinement
4.6. Vibration
4.7. Combinations of Stressors
4.8. Simulation of Microgravity using the Hindlimb Unloading Model
4.9. Simulation of Socio-Environmental Stressors Encountered During a Space Mission Using the CUMS Model
4.10. RT-qPCR
4.11. Western Blotting
4.12. Statistics
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| A.U. | Arbitrary unit |
| BLAST | Basic local alignment search tool |
| EADS | European aeronautic defense and space company |
| ERK | Extracellular signal-regulated kinase |
| FXa | Factor Xa |
| FXIa | Factor XIa |
| HU | Hindlimb unloading |
| ISS | International space station |
| MHC | Major histocompatibility complex |
| MML-V | Moloney murine leukemia virus |
| PI3K | Phosphatidylinositol 3-kinase |
| PVDF | Polyvinylidene difluoride |
| RT-qPCR | Reverse transcription-quantitative PCR |
| RPM | Random positioning machine |
| ROS | Reactive oxygen species |
| TLR | Toll-like receptors |
References
- Crucian, B.; Stowe, R.P.; Mehta, S.; Quiriarte, H.; Pierson, D.; Sams, C. Alterations in adaptive immunity persist during long-duration spaceflight. Npj Microgravity 2015, 1, 15013. [Google Scholar] [CrossRef] [Green Version]
- Mehta, S.K.; Laudenslager, M.L.; Stowe, R.P.; Crucian, B.E.; Feiveson, A.H.; Sams, C.F.; Pierson, D.L. Latent virus reactivation in astronauts on the international space station. NPJ Microgravity 2017, 3, 11. [Google Scholar] [CrossRef]
- Crucian, B.; Babiak-Vazquez, A.; Johnston, S.; Pierson, D.L.; Ott, C.M.; Sams, C. Incidence of clinical symptoms during long-duration orbital spaceflight. Int. J. Gen. Med. 2016, 9, 383–391. [Google Scholar] [CrossRef] [Green Version]
- Crucian, B.; Johnston, S.; Mehta, S.; Stowe, R.; Uchakin, P.; Quiriarte, H.; Pierson, D.; Laudenslager, M.L.; Sams, C. A case of persistent skin rash and rhinitis with immune system dysregulation onboard the International Space Station. J. Allergy Clin. Immunol. Pract. 2016, 4, 759–762.e8. [Google Scholar] [CrossRef]
- Crucian, B.; Stowe, R.; Quiriarte, H.; Pierson, D.; Sams, C. Monocyte phenotype and cytokine production profiles are dysregulated by short-duration spaceflight. Aviat. Space Environ. Med. 2011, 82, 857–862. [Google Scholar] [CrossRef] [PubMed]
- Kaur, I.; Simons, E.R.; Kapadia, A.S.; Ott, C.M.; Pierson, D.L. Effect of spaceflight on ability of monocytes to respond to endotoxins of gram-negative bacteria. Clin. Vaccine Immunol. CVI 2008, 15, 1523–1528. [Google Scholar] [CrossRef] [PubMed]
- Rykova, M.P.; Antropova, E.N.; Larina, I.M.; Morukov, B.V. Humoral and cellular immunity in cosmonauts after the ISS missions. Acta Astronaut. 2008, 63, 697–705. [Google Scholar] [CrossRef]
- Brungs, S.; Kolanus, W.; Hemmersbach, R. Syk phosphorylation—a gravisensitive step in macrophage signalling. Cell Commun. Signal. CCS 2015, 13, 9. [Google Scholar] [CrossRef] [PubMed]
- Kaur, I.; Simons, E.R.; Castro, V.A.; Mark Ott, C.; Pierson, D.L. Changes in neutrophil functions in astronauts. Brain. Behav. Immun. 2004, 18, 443–450. [Google Scholar] [CrossRef] [Green Version]
- Cogoli, A.; Tschopp, A.; Fuchs-Bislin, P. Cell sensitivity to gravity. Science 1984, 225, 228–230. [Google Scholar] [CrossRef]
- Cogoli, A. The effect of space flight on human cellular immunity. Environ. Med. Annu. Rep. Res. Inst. Environ. Med. Nagoya Univ. 1993, 37, 107–116. [Google Scholar]
- Gridley, D.S.; Slater, J.M.; Luo-Owen, X.; Rizvi, A.; Chapes, S.K.; Stodieck, L.S.; Ferguson, V.L.; Pecaut, M.J. Spaceflight effects on T lymphocyte distribution, function and gene expression. J. Appl. Physiol. Bethesda Md 1985 2009, 106, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Frippiat, J.-P.; Crucian, B.E.; de Quervain, D.J.-F.; Grimm, D.; Montano, N.; Praun, S.; Roozendaal, B.; Schelling, G.; Thiel, M.; Ullrich, O.; et al. Towards human exploration of space: The THESEUS review series on immunology research priorities. NPJ Microgravity 2016, 2, 16040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guéguinou, N.; Huin-Schohn, C.; Bascove, M.; Bueb, J.-L.; Tschirhart, E.; Legrand-Frossi, C.; Frippiat, J.-P. Could spaceflight-associated immune system weakening preclude the expansion of human presence beyond Earth’s orbit? J. Leukoc. Biol. 2009, 86, 1027–1038. [Google Scholar] [CrossRef] [PubMed]
- Frippiat, J.-P. Contribution of the urodele amphibian Pleurodeles waltl to the analysis of spaceflight-associated immune system deregulation. Mol. Immunol. 2013, 56, 434–441. [Google Scholar] [CrossRef] [PubMed]
- Bascove, M.; Huin-Schohn, C.; Guéguinou, N.; Tschirhart, E.; Frippiat, J.-P. Spaceflight-associated changes in immunoglobulin VH gene expression in the amphibian Pleurodeles waltl. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2009, 23, 1607–1615. [Google Scholar] [CrossRef] [PubMed]
- Boxio, R.; Dournon, C.; Frippiat, J.-P. Effects of a long-term spaceflight on immunoglobulin heavy chains of the urodele amphibian Pleurodeles waltl. J. Appl. Physiol. Bethesda Md 1985 2005, 98, 905–910. [Google Scholar] [CrossRef] [PubMed]
- Bascove, M.; Guéguinou, N.; Schaerlinger, B.; Gauquelin-Koch, G.; Frippiat, J.-P. Decrease in antibody somatic hypermutation frequency under extreme, extended spaceflight conditions. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2011, 25, 2947–2955. [Google Scholar] [CrossRef] [PubMed]
- Rettig, T.A.; Bye, B.A.; Nishiyama, N.C.; Hlavacek, S.; Ward, C.; Pecaut, M.J.; Chapes, S.K. Effects of skeletal unloading on the antibody repertoire of tetanus toxoid and/or CpG treated C57BL/6J mice. PLoS ONE 2019, 14, e0210284. [Google Scholar] [CrossRef] [PubMed]
- Gaignier, F.; Schenten, V.; De Carvalho Bittencourt, M.; Gauquelin-Koch, G.; Frippiat, J.-P.; Legrand-Frossi, C. Three weeks of murine hindlimb unloading induces shifts from B to T and from th to tc splenic lymphocytes in absence of stress and differentially reduces cell-specific mitogenic responses. PLoS ONE 2014, 9, e92664. [Google Scholar] [CrossRef]
- Guéguinou, N.; Bojados, M.; Jamon, M.; Derradji, H.; Baatout, S.; Tschirhart, E.; Frippiat, J.-P.; Legrand-Frossi, C. Stress response and humoral immune system alterations related to chronic hypergravity in mice. Psychoneuroendocrinology 2012, 37, 137–147. [Google Scholar] [CrossRef]
- Vacek, A.; Michurina, T.V.; Serova, L.V.; Rotkovská, D.; Bartonícková, A. Decrease in the number of progenitors of erythrocytes (BFUe, CFUe), granulocytes and macrophages (GM-CFC) in bone marrow of rats after a 14-day flight onboard the Cosmos-2044 Biosatellite. Folia Biol. (Praha) 1991, 37, 35–41. [Google Scholar]
- Ichiki, A.T.; Gibson, L.A.; Jago, T.L.; Strickland, K.M.; Johnson, D.L.; Lange, R.D.; Allebban, Z. Effects of spaceflight on rat peripheral blood leukocytes and bone marrow progenitor cells. J. Leukoc. Biol. 1996, 60, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Davis, T.A.; Wiesmann, W.; Kidwell, W.; Cannon, T.; Kerns, L.; Serke, C.; Delaplaine, T.; Pranger, A.; Lee, K.P. Effect of spaceflight on human stem cell hematopoiesis: Suppression of erythropoiesis and myelopoiesis. J. Leukoc. Biol. 1996, 60, 69–76. [Google Scholar] [CrossRef]
- Ortega, M.T.; Pecaut, M.J.; Gridley, D.S.; Stodieck, L.S.; Ferguson, V.; Chapes, S.K. Shifts in bone marrow cell phenotypes caused by spaceflight. J. Appl. Physiol. Bethesda Md 1985 2009, 106, 548–555. [Google Scholar] [CrossRef]
- Sotnezova, E.V.; Markina, E.A.; Andreeva, E.R.; Buravkova, L.B. Myeloid Precursors in the Bone Marrow of Mice after a 30-Day Space Mission on a Bion-M1 Biosatellite. Bull. Exp. Biol. Med. 2017, 162, 496–500. [Google Scholar] [CrossRef]
- Huin-Schohn, C.; Guéguinou, N.; Schenten, V.; Bascove, M.; Gauquelin-Koch, G.; Baatout, S.; Tschirhart, E.; Frippiat, J.-P. Gravity changes during animal development affect IgM heavy-chain transcription and probably lymphopoiesis. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2013, 27, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Lescale, C.; Schenten, V.; Djeghloul, D.; Bennabi, M.; Gaignier, F.; Vandamme, K.; Strazielle, C.; Kuzniak, I.; Petite, H.; Dosquet, C.; et al. Hind limb unloading, a model of spaceflight conditions, leads to decreased B lymphopoiesis similar to aging. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2015, 29, 455–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tascher, G.; Gerbaix, M.; Maes, P.; Chazarin, B.; Ghislin, S.; Antropova, E.; Vassilieva, G.; Ouzren-Zarhloul, N.; Gauquelin-Koch, G.; Vico, L.; et al. Analysis of femurs from mice embarked on board BION-M1 biosatellite reveals a decrease in immune cell development, including B cells, after 1 wk of recovery on Earth. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2018, fj201801463R. [Google Scholar] [CrossRef]
- Woods, C.C.; Banks, K.E.; Gruener, R.; DeLuca, D. Loss of T cell precursors after spaceflight and exposure to vector-averaged gravity. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2003, 17, 1526–1528. [Google Scholar] [CrossRef] [Green Version]
- Woods, C.C.; Banks, K.E.; Lebsack, T.W.; White, T.C.; Anderson, G.A.; Maccallum, T.; Gruener, R.; DeLuca, D. Use of a microgravity organ culture dish system to demonstrate the signal dampening effects of modeled microgravity during T cell development. Dev. Comp. Immunol. 2005, 29, 565–582. [Google Scholar] [CrossRef] [PubMed]
- Ghislin, S.; Ouzren-Zarhloul, N.; Kaminski, S.; Frippiat, J.-P. Hypergravity exposure during gestation modifies the TCRβ repertoire of newborn mice. Sci. Rep. 2015, 5, 9318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ricklin, D.; Hajishengallis, G.; Yang, K.; Lambris, J.D. Complement: A key system for immune surveillance and homeostasis. Nat. Immunol. 2010, 11, 785–797. [Google Scholar] [CrossRef] [PubMed]
- Hajishengallis, G.; Lambris, J.D. Crosstalk pathways between Toll-like receptors and the complement system. Trends Immunol. 2010, 31, 154–163. [Google Scholar] [CrossRef] [Green Version]
- Dunkelberger, J.R.; Song, W.-C. Complement and its role in innate and adaptive immune responses. Cell Res. 2010, 20, 34–50. [Google Scholar] [CrossRef]
- Jalili, A.; Shirvaikar, N.; Marquez-Curtis, L.; Qiu, Y.; Korol, C.; Lee, H.; Turner, A.R.; Ratajczak, M.Z.; Janowska-Wieczorek, A. Fifth complement cascade protein (C5) cleavage fragments disrupt the SDF-1/CXCR4 axis: Further evidence that innate immunity orchestrates the mobilization of hematopoietic stem/progenitor cells. Exp. Hematol. 2010, 38, 321–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flierman, R.; Daha, M.R. The clearance of apoptotic cells by complement. Immunobiology 2007, 212, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Trouw, L.A.; Blom, A.M.; Gasque, P. Role of complement and complement regulators in the removal of apoptotic cells. Mol. Immunol. 2008, 45, 1199–1207. [Google Scholar] [CrossRef]
- Markiewski, M.M.; DeAngelis, R.A.; Lambris, J.D. Liver inflammation and regeneration: Two distinct biological phenomena or parallel pathophysiologic processes? Mol. Immunol. 2006, 43, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Mastellos, D.C.; Deangelis, R.A.; Lambris, J.D. Complement-triggered pathways orchestrate regenerative responses throughout phylogenesis. Semin. Immunol. 2013, 25, 29–38. [Google Scholar] [CrossRef] [Green Version]
- Markiewski, M.M.; Nilsson, B.; Ekdahl, K.N.; Mollnes, T.E.; Lambris, J.D. Complement and coagulation: Strangers or partners in crime? Trends Immunol. 2007, 28, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Alexander, J.J.; Anderson, A.J.; Barnum, S.R.; Stevens, B.; Tenner, A.J. The complement cascade: Yin-Yang in neuroinflammation--neuro-protection and -degeneration. J. Neurochem. 2008, 107, 1169–1187. [Google Scholar] [CrossRef] [PubMed]
- Shinjyo, N.; Ståhlberg, A.; Dragunow, M.; Pekny, M.; Pekna, M. Complement-derived anaphylatoxin C3a regulates in vitro differentiation and migration of neural progenitor cells. Stem Cells Dayt. Ohio 2009, 27, 2824–2832. [Google Scholar] [CrossRef] [PubMed]
- Nonaka, M.; Yoshizaki, F. Evolution of the complement system. Mol. Immunol. 2004, 40, 897–902. [Google Scholar] [CrossRef]
- Guéguinou, N.; Huin-Schohn, C.; Ouzren-Zarhloul, N.; Ghislin, S.; Frippiat, J.-P. Molecular cloning and expression analysis of Pleurodeles waltl complement component C3 under normal physiological conditions and environmental stresses. Dev. Comp. Immunol. 2014, 46, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Liszewski, M.K.; Kolev, M.; Le Friec, G.; Leung, M.; Bertram, P.G.; Fara, A.F.; Subias, M.; Pickering, M.C.; Drouet, C.; Meri, S.; et al. Intracellular complement activation sustains T cell homeostasis and mediates effector differentiation. Immunity 2013, 39, 1143–1157. [Google Scholar] [CrossRef] [PubMed]
- Berger, T.; Przybyla, B.; Matthiä, D.; Reitz, G.; Burmeister, S.; Labrenz, J.; Bilski, P.; Horwacik, T.; Twardak, A.; Hajek, M.; et al. DOSIS & DOSIS 3D: Long-term dose monitoring onboard the Columbus Laboratory of the International Space Station (ISS). J. Space Weather Space Clim. 2016, 6, A39. [Google Scholar]
- Berger, T.; Burmeister, S.; Matthiä, D.; Przybyla, B.; Reitz, G.; Bilski, P.; Hajek, M.; Sihver, L.; Szabo, J.; Ambrozova, I.; et al. DOSIS & DOSIS 3D: Radiation measurements with the DOSTEL instruments onboard the Columbus Laboratory of the ISS in the years 2009–2016. J. Space Weather Space Clim. 2017, 7, A8. [Google Scholar]
- Globus, R.K.; Morey-Holton, E. Hindlimb unloading: Rodent analog for microgravity. J. Appl. Physiol. Bethesda Md 1985 2016, 120, 1196–1206. [Google Scholar] [CrossRef]
- Gaignier, F.; Legrand-Frossi, C.; Stragier, E.; Mathiot, J.; Merlin, J.-L.; Cohen-Salmon, C.; Lanfumey, L.; Frippiat, J.-P. A Model of Chronic Exposure to Unpredictable Mild Socio-Environmental Stressors Replicates Some Spaceflight-Induced Immunological Changes. Front. Physiol. 2018, 9, 514. [Google Scholar] [CrossRef] [PubMed]
- Schaerlinger, B.; Frippiat, J.-P. IgX antibodies in the urodele amphibian Ambystoma mexicanum. Dev. Comp. Immunol. 2008, 32, 908–915. [Google Scholar] [CrossRef]
- Bascove, M.; Frippiat, J.-P. Molecular characterization of Pleurodeles waltl activation-induced cytidine deaminase. Mol. Immunol. 2010, 47, 1640–1649. [Google Scholar] [CrossRef]
- Fonte, C.; Gruez, A.; Ghislin, S.; Frippiat, J.-P. The urodele amphibian Pleurodeles waltl has a diverse repertoire of immunoglobulin heavy chains with polyreactive and species-specific features. Dev. Comp. Immunol. 2015, 53, 371–384. [Google Scholar] [CrossRef]
- Kolev, M.; Le Friec, G.; Kemper, C. Complement--tapping into new sites and effector systems. Nat. Rev. Immunol. 2014, 14, 811–820. [Google Scholar] [CrossRef]
- Surace, L.; Lysenko, V.; Fontana, A.O.; Cecconi, V.; Janssen, H.; Bicvic, A.; Okoniewski, M.; Pruschy, M.; Dummer, R.; Neefjes, J.; et al. Complement is a central mediator of radiotherapy-induced tumor-specific immunity and clinical response. Immunity 2015, 42, 767–777. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, D.A.; Schwarzenberg, M.; Tkaczuk, J.; Hebrard, S.; Brandenberger, G.; Mauco, G.; Cogoli-Greuter, M.; Abbal, M. Head-down tilt bed rest and immune responses. Pflugers Arch. 2000, 441, R79–R84. [Google Scholar] [CrossRef] [PubMed]
- Reis, E.S.; Lange, T.; Köhl, G.; Herrmann, A.; Tschulakow, A.V.; Naujoks, J.; Born, J.; Köhl, J. Sleep and circadian rhythm regulate circulating complement factors and immunoregulatory properties of C5a. Brain. Behav. Immun. 2011, 25, 1416–1426. [Google Scholar] [CrossRef]
- Hollander, J.; Gore, M.; Fiebig, R.; Mazzeo, R.; Ohishi, S.; Ohno, H.; Ji, L.L. Spaceflight downregulates antioxidant defense systems in rat liver. Free Radic. Biol. Med. 1998, 24, 385–390. [Google Scholar] [CrossRef]
- Rizzo, A.M.; Rossi, F.; Zava, S.; Montorfano, G.; Adorni, L.; Cotronei, V.; Zanini, A.; Berra, B. Antioxidant metabolism in Xenopus laevis embryos is affected by stratospheric balloon flight. Cell Biol. Int. 2007, 31, 716–723. [Google Scholar] [CrossRef] [PubMed]
- De Luca, C.; Deeva, I.; Mariani, S.; Maiani, G.; Stancato, A.; Korkina, L. Monitoring antioxidant defenses and free radical production in space-flight, aviation and railway engine operators, for the prevention and treatment of oxidative stress, immunological impairment, and pre-mature cell aging. Toxicol. Ind. Health 2009, 25, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Tahimic, C.G.T.; Globus, R.K. Redox Signaling and Its Impact on Skeletal and Vascular Responses to Spaceflight. Int. J. Mol. Sci. 2017, 18. [Google Scholar] [CrossRef] [PubMed]
- Collard, C.D.; Lekowski, R.; Jordan, J.E.; Agah, A.; Stahl, G.L. Complement activation following oxidative stress. Mol. Immunol. 1999, 36, 941–948. [Google Scholar] [CrossRef]
- Collard, C.D.; Väkevä, A.; Morrissey, M.A.; Agah, A.; Rollins, S.A.; Reenstra, W.R.; Buras, J.A.; Meri, S.; Stahl, G.L. Complement activation after oxidative stress: Role of the lectin complement pathway. Am. J. Pathol. 2000, 156, 1549–1556. [Google Scholar] [CrossRef]
- Hart, M.L.; Walsh, M.C.; Stahl, G.L. Initiation of complement activation following oxidative stress. In vitro and in vivo observations. Mol. Immunol. 2004, 41, 165–171. [Google Scholar] [CrossRef]
- Baqai, F.P.; Gridley, D.S.; Slater, J.M.; Luo-Owen, X.; Stodieck, L.S.; Ferguson, V.; Chapes, S.K.; Pecaut, M.J. Effects of spaceflight on innate immune function and antioxidant gene expression. J. Appl. Physiol. Bethesda Md 1985 2009, 106, 1935–1942. [Google Scholar] [CrossRef] [Green Version]
- Ishihara, A.; Fujino, H.; Nagatomo, F.; Takeda, I.; Ohira, Y. Gene expression levels of heat shock proteins in the soleus and plantaris muscles of rats after hindlimb suspension or spaceflight. J. Physiol. Sci. JPS 2008, 58, 413–417. [Google Scholar] [CrossRef]
- Stott, B.; Korbelik, M. Activation of complement C3, C5, and C9 genes in tumors treated by photodynamic therapy. Cancer Immunol. Immunother. CII 2007, 56, 649–658. [Google Scholar] [CrossRef]
- McLin, V.A.; Hu, C.-H.; Shah, R.; Jamrich, M. Expression of complement components coincides with early patterning and organogenesis in Xenopus laevis. Int. J. Dev. Biol. 2008, 52, 1123–1133. [Google Scholar] [CrossRef] [PubMed]
- Carmona-Fontaine, C.; Theveneau, E.; Tzekou, A.; Tada, M.; Woods, M.; Page, K.M.; Parsons, M.; Lambris, J.D.; Mayor, R. Complement fragment C3a controls mutual cell attraction during collective cell migration. Dev. Cell 2011, 21, 1026–1037. [Google Scholar] [CrossRef] [PubMed]
- Amara, U.; Flierl, M.A.; Rittirsch, D.; Klos, A.; Chen, H.; Acker, B.; Brückner, U.B.; Nilsson, B.; Gebhard, F.; Lambris, J.D.; et al. Molecular intercommunication between the complement and coagulation systems. J. Immunol. Baltim. Md 1950 2010, 185, 5628–5636. [Google Scholar] [CrossRef] [PubMed]
- Kimzey, S.L.; Ritzmann, S.E.; Mengel, C.E.; Fischer, C.L. Skylab experiment results: Hematology studies. Acta Astronaut. 1975, 2, 141–154. [Google Scholar] [CrossRef]
- Larina, I.M.; Percy, A.J.; Yang, J.; Borchers, C.H.; Nosovsky, A.M.; Grigoriev, A.I.; Nikolaev, E.N. Protein expression changes caused by spaceflight as measured for 18 Russian cosmonauts. Sci. Rep. 2017, 7, 8142. [Google Scholar] [CrossRef]
- Lewis, M.L.; Reynolds, J.L.; Cubano, L.A.; Hatton, J.P.; Lawless, B.D.; Piepmeier, E.H. Spaceflight alters microtubules and increases apoptosis in human lymphocytes (Jurkat). FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 1998, 12, 1007–1018. [Google Scholar] [CrossRef] [Green Version]
- Sharma, C.S.; Sarkar, S.; Periyakaruppan, A.; Ravichandran, P.; Sadanandan, B.; Ramesh, V.; Thomas, R.; Hall, J.C.; Wilson, B.L.; Ramesh, G.T. Simulated microgravity activates apoptosis and NF-kappaB in mice testis. Mol. Cell. Biochem. 2008, 313, 71–78. [Google Scholar] [CrossRef]
- Kang, C.-Y.; Zou, L.; Yuan, M.; Wang, Y.; Li, T.-Z.; Zhang, Y.; Wang, J.-F.; Li, Y.; Deng, X.-W.; Liu, C.-T. Impact of simulated microgravity on microvascular endothelial cell apoptosis. Eur. J. Appl. Physiol. 2011, 111, 2131–2138. [Google Scholar] [CrossRef]
- Lin, S.-C.; Gou, G.-H.; Hsia, C.-W.; Ho, C.-W.; Huang, K.-L.; Wu, Y.-F.; Lee, S.-Y.; Chen, Y.-H. Simulated Microgravity Disrupts Cytoskeleton Organization and Increases Apoptosis of Rat Neural Crest Stem Cells Via Upregulating CXCR4 Expression and RhoA-ROCK1-p38 MAPK-p53 Signaling. Stem Cells Dev. 2016, 25, 1172–1193. [Google Scholar] [CrossRef] [PubMed]
- Strainic, M.G.; Liu, J.; Huang, D.; An, F.; Lalli, P.N.; Muqim, N.; Shapiro, V.S.; Dubyak, G.R.; Heeger, P.S.; Medof, M.E. Locally produced complement fragments C5a and C3a provide both costimulatory and survival signals to naive CD4+ T cells. Immunity 2008, 28, 425–435. [Google Scholar] [CrossRef]
- Weaver, D.J.; Reis, E.S.; Pandey, M.K.; Köhl, G.; Harris, N.; Gerard, C.; Köhl, J. C5a receptor-deficient dendritic cells promote induction of Treg and Th17 cells. Eur. J. Immunol. 2010, 40, 710–721. [Google Scholar] [CrossRef]
- Chang, T.T.; Spurlock, S.M.; Candelario, T.L.T.; Grenon, S.M.; Hughes-Fulford, M. Spaceflight impairs antigen-specific tolerance induction in vivo and increases inflammatory cytokines. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2015, 29, 4122–4132. [Google Scholar] [CrossRef] [Green Version]
- Hawlisch, H.; Belkaid, Y.; Baelder, R.; Hildeman, D.; Gerard, C.; Köhl, J. C5a negatively regulates toll-like receptor 4-induced immune responses. Immunity 2005, 22, 415–426. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Chen, H.; Luo, H.; Zhu, L.; Zhao, Y.; Tian, H.; Wang, R.; Shang, P.; Zhao, Y. Microgravity activates p38 MAPK-C/EBPβ pathway to regulate the expression of arginase and inflammatory cytokines in macrophages. Inflamm. Res. Off. J. Eur. Histamine Res. Soc. Al 2015, 64, 303–311. [Google Scholar] [CrossRef]
- Tauber, S.; Hauschild, S.; Crescio, C.; Secchi, C.; Paulsen, K.; Pantaleo, A.; Saba, A.; Buttron, I.; Thiel, C.S.; Cogoli, A.; et al. Signal transduction in primary human T lymphocytes in altered gravity—results of the MASER-12 suborbital space flight mission. Cell Commun. Signal. CCS 2013, 11, 32. [Google Scholar] [CrossRef]
- Kim, J.; Montagne, K.; Nemoto, H.; Ushida, T.; Furukawa, K.S. Hypergravity down-regulates c-fos gene expression via ROCK/Rho-GTP and the PI3K signaling pathway in murine ATDC5 chondroprogenitor cells. PLoS ONE 2017, 12, e0185394. [Google Scholar] [CrossRef]
- Crucian, B.E.; Choukèr, A.; Simpson, R.J.; Mehta, S.; Marshall, G.; Smith, S.M.; Zwart, S.R.; Heer, M.; Ponomarev, S.; Whitmire, A.; et al. Immune System Dysregulation During Spaceflight: Potential Countermeasures for Deep Space Exploration Missions. Front. Immunol. 2018, 9, 1437. [Google Scholar] [CrossRef]
- Shi, D.L.; Boucaut, J.C. The chronological development of the urodele amphibian Pleurodeles waltl (Michah). Int. J. Dev. Biol. 1995, 39, 427–441. [Google Scholar]
- Herranz, R.; Anken, R.; Boonstra, J.; Braun, M.; Christianen, P.C.M.; de Geest, M.; Hauslage, J.; Hilbig, R.; Hill, R.J.A.; Lebert, M.; et al. Ground-based facilities for simulation of microgravity: Organism-specific recommendations for their use, and recommended terminology. Astrobiology 2013, 13, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Vanhavere, F.; Coeck, M. Comparison between thermoluminescence and electronic dosimetry results at the Belgian Nuclear Research Centre. Radiat. Prot. Dosimetry 2001, 96, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Fonte, C.; Kaminski, S.; Vanet, A.; Lanfumey, L.; Cohen-Salmon, C.; Ghislin, S.; Frippiat, J.-P. Socioenvironmental stressors encountered during spaceflight partially affect the murine TCR-β repertoire and increase its self-reactivity. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2019, 33, 896–908. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, RESEARCH0034. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Sequences | Annealing Temperature (°C) |
|---|---|---|
| P. waltl C3 | F: 5′-TGGTGACAATGACACTGCCT-3′ | 62 |
| R: 5′-CATCCACCCAGATGGAGTCT-3′ | ||
| Murine C3 | F: 5′-AGAGGCAAGTGCTGACCAGT-3′ | 62 |
| R: 5′-ACTGGCTGGAATCTTGATGG-3′ | ||
| P. waltl actin | F: 5′-TGGTCGTGACCTGACTGATT-3′ | 60 |
| R: 5′-TCACGGACAATCTCACGTTC-3′ | ||
| P. waltl TAFII | F: 5′-TTCACGAGCTGTCTGTGGAG-3′ | 60 |
| R: 5′-CCTGGGAAGCATTTGGTAGA-3′ | ||
| P. waltl mtRNA16S | F: 5′-CGTGCAGAAGCGGAGATAA-3′ | 60 |
| R: 5′-TGTCGGGCTGTTGTAGGG-3′ | ||
| P. waltl GAPDH | F: 5′-GAAGGTAGTAAGCAACGCCTCCT-3′ | 65 |
| R: 5′-CACAGCATGTACAGTGGTCATCA-3′ | ||
| Murine Ppia | F: 5′-GTCTCCTTCGAGCTGTTTGC-3′ | 58 |
| R: 5′-GCGTGTAAAGTCACCACCCT-3′ | ||
| Murine Eef2 | F: 5′-GTGGTGGACTGTGTGTCTGG-3′ | 58 |
| R: 5′-CGCTGGAAGGTCTGGTAGAG-3′ | ||
| Murine Rpl13 | F: 5′-GGAAGCGGATGAATACCAAC-3′ | 61 |
| R: 5′-CTTGTCATAGGGTGGAGCGA-3′ | ||
| Murine Eif3f | F: 5′-CATCAAGGCCTATGTCAGCA-3′ | 61 |
| R: 5′-AGGTCAACTCCAATGCGTTC-3′ |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guéguinou, N.; Jeandel, J.; Kaminski, S.; Baatout, S.; Ghislin, S.; Frippiat, J.-P. Modulation of Iberian Ribbed Newt Complement Component C3 by Stressors Similar to those Encountered during a Stay Onboard the International Space Station. Int. J. Mol. Sci. 2019, 20, 1579. https://doi.org/10.3390/ijms20071579
Guéguinou N, Jeandel J, Kaminski S, Baatout S, Ghislin S, Frippiat J-P. Modulation of Iberian Ribbed Newt Complement Component C3 by Stressors Similar to those Encountered during a Stay Onboard the International Space Station. International Journal of Molecular Sciences. 2019; 20(7):1579. https://doi.org/10.3390/ijms20071579
Chicago/Turabian StyleGuéguinou, Nathan, Jérémy Jeandel, Sandra Kaminski, Sarah Baatout, Stéphanie Ghislin, and Jean-Pol Frippiat. 2019. "Modulation of Iberian Ribbed Newt Complement Component C3 by Stressors Similar to those Encountered during a Stay Onboard the International Space Station" International Journal of Molecular Sciences 20, no. 7: 1579. https://doi.org/10.3390/ijms20071579
APA StyleGuéguinou, N., Jeandel, J., Kaminski, S., Baatout, S., Ghislin, S., & Frippiat, J. -P. (2019). Modulation of Iberian Ribbed Newt Complement Component C3 by Stressors Similar to those Encountered during a Stay Onboard the International Space Station. International Journal of Molecular Sciences, 20(7), 1579. https://doi.org/10.3390/ijms20071579