Dehydroabietic Acid Suppresses Inflammatory Response Via Suppression of Src-, Syk-, and TAK1-Mediated Pathways

 , ,
, , 
Abstract
:1. Introduction
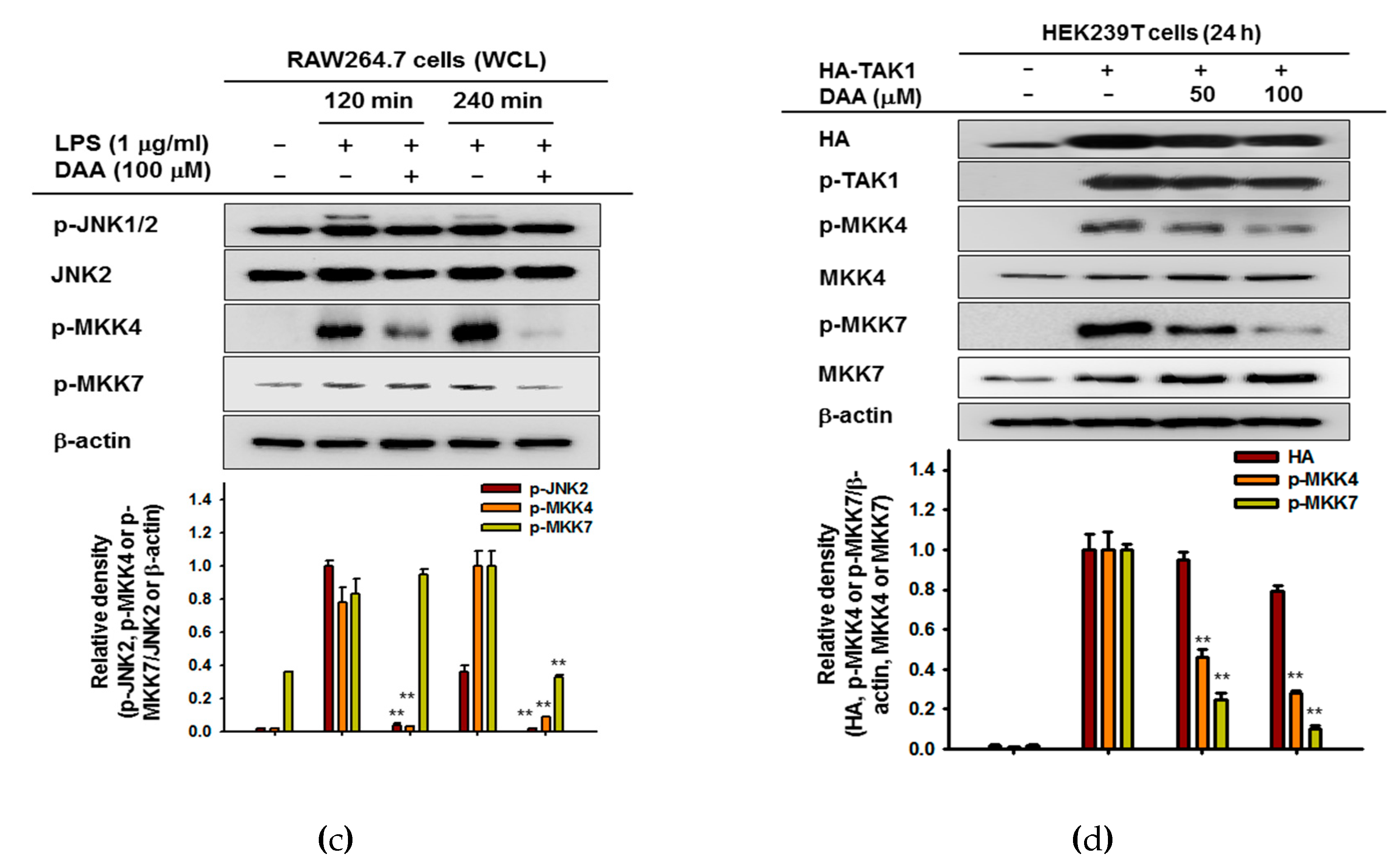
2. Results
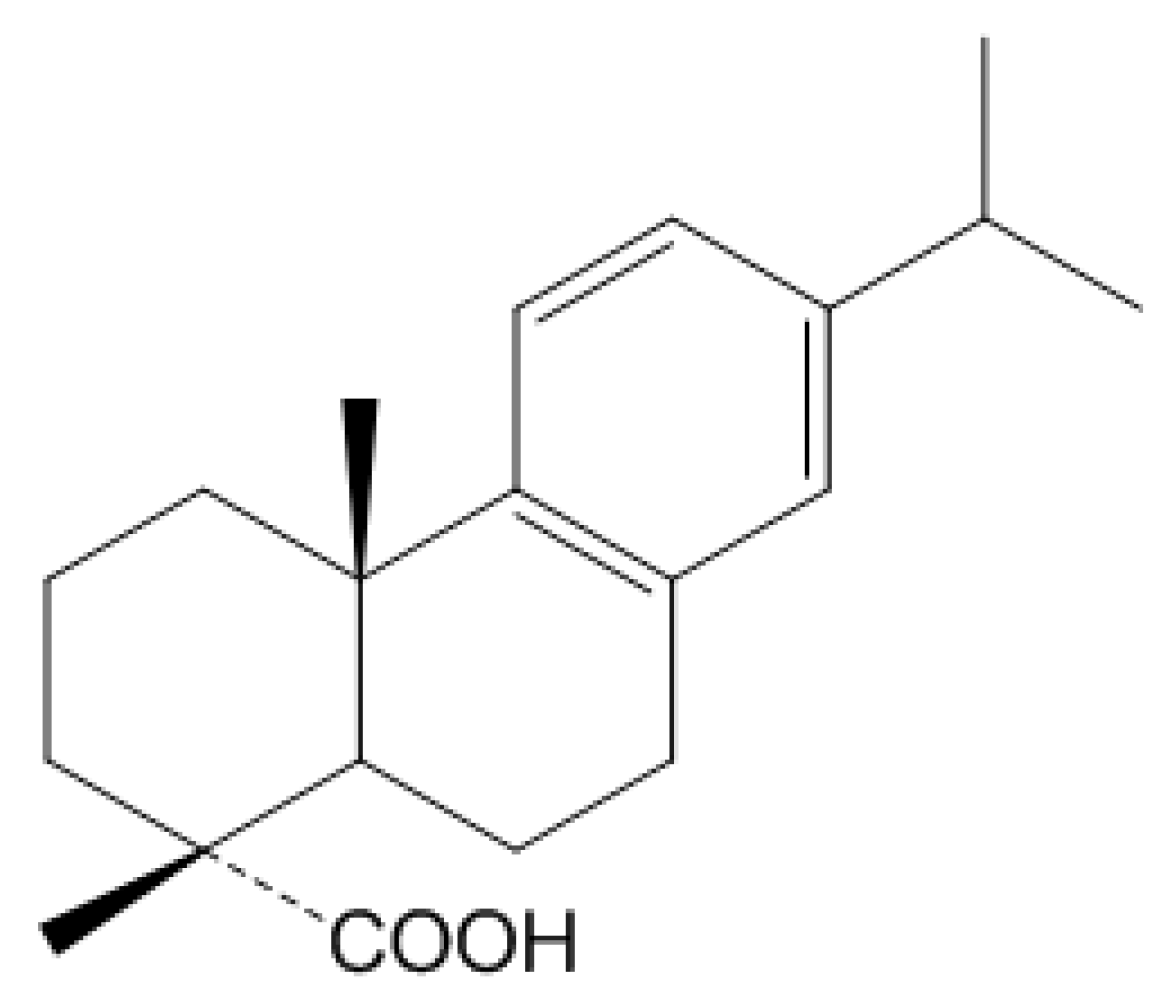
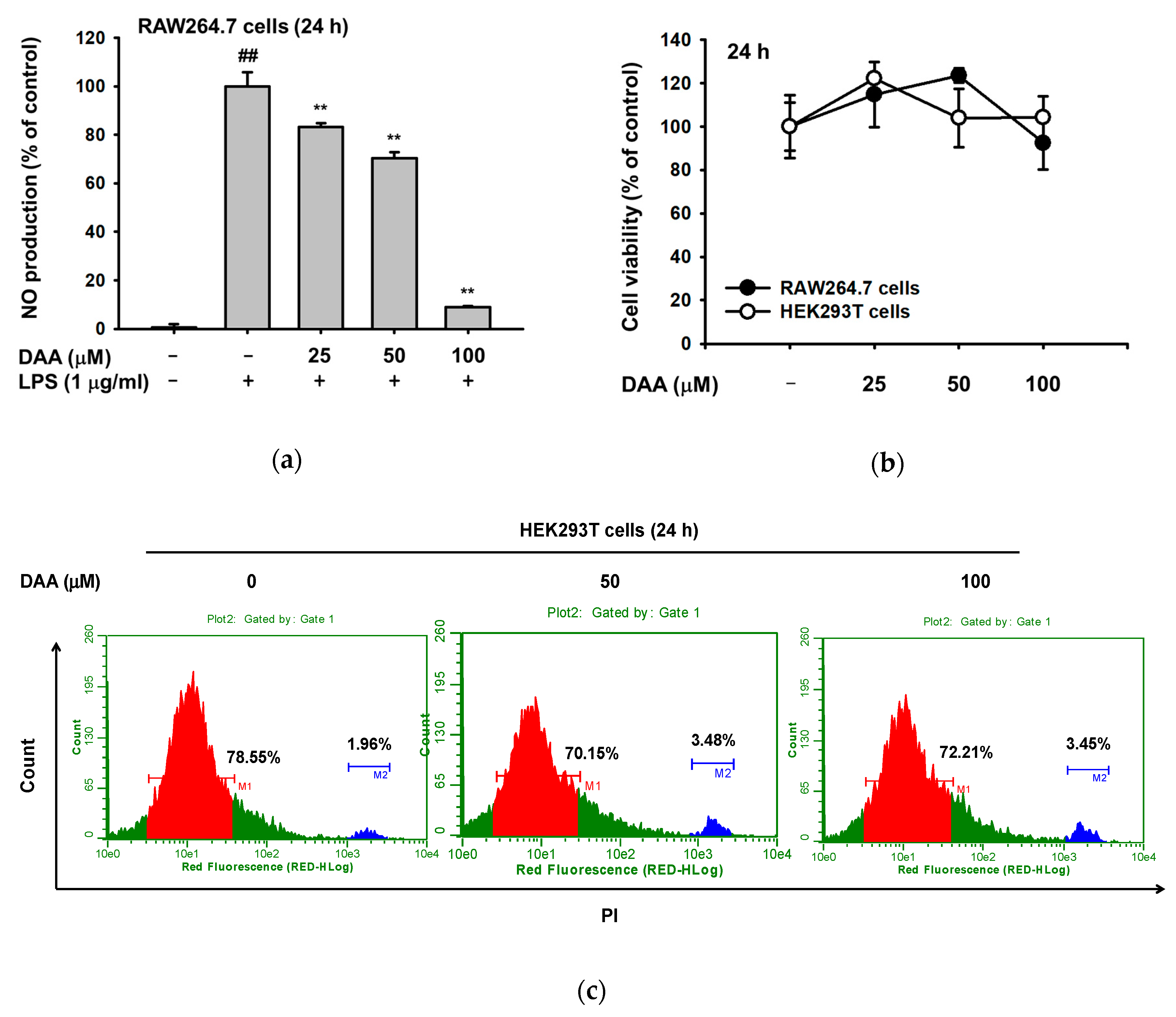
2.1. The Effect of DAA on Nitric Oxide Production
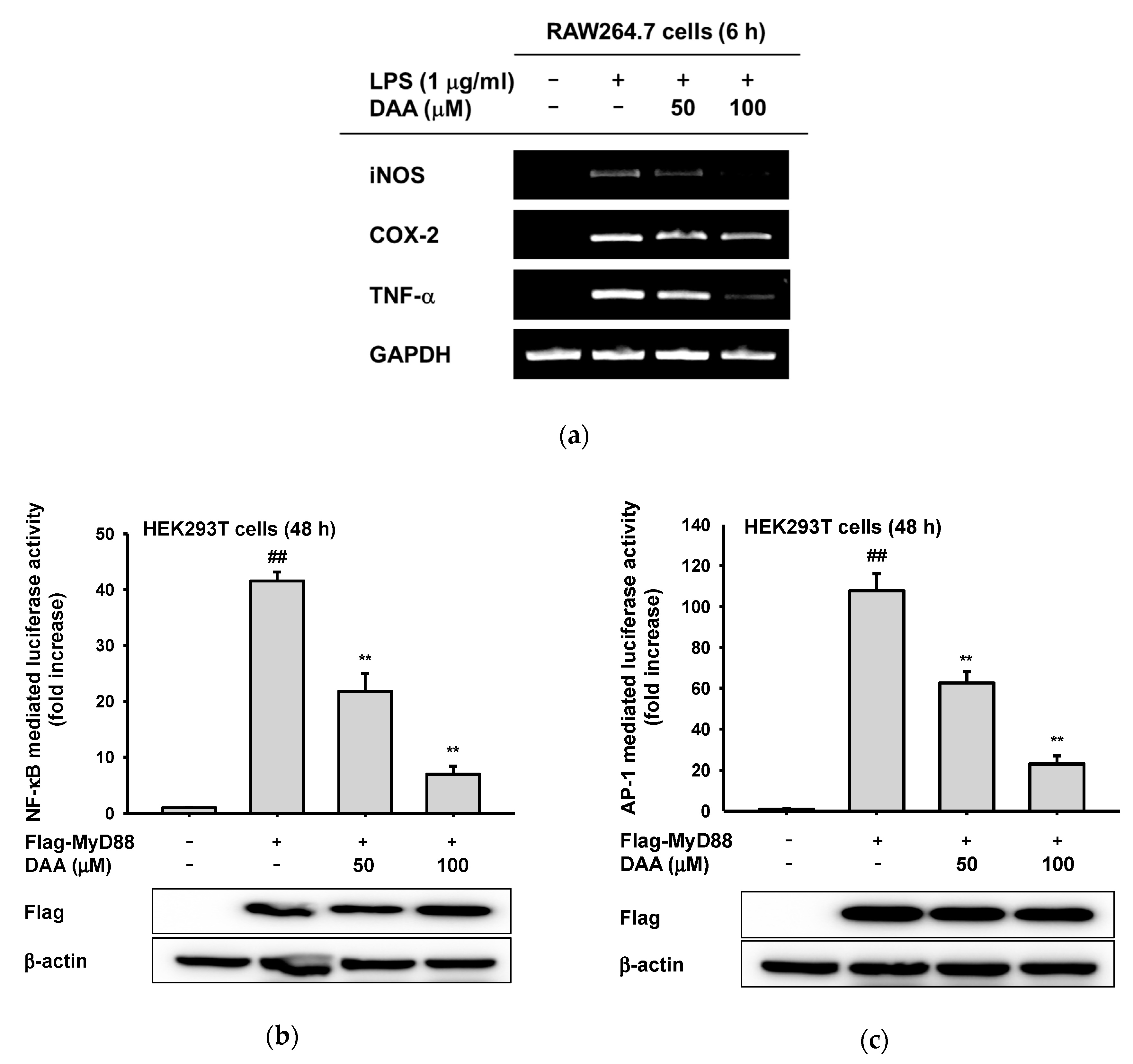
2.2. The Anti-Inflammatory Effect of DAA at the Transcriptional Level
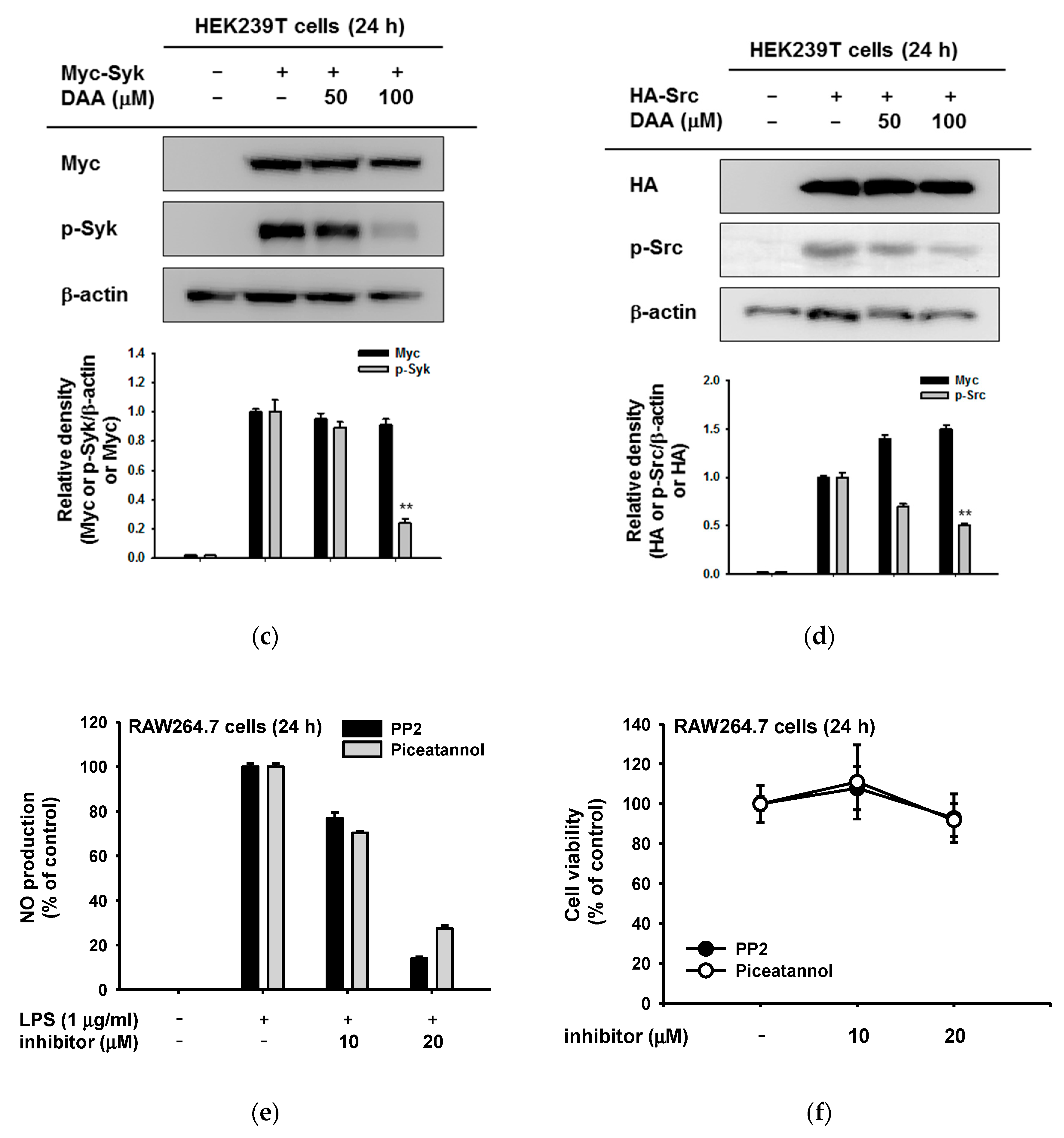
2.3. The Anti-Inflammatory Effect of DAA on the NF-κB Signaling Pathway
2.4. The Anti-Inflammatory Effect of DAA in the AP-1 Signaling Pathway
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture
4.3. NO Production and Griess Assay
4.4. Cell Viability Assay
4.5. Preparation of mRNA and Semi-Quantitative PCR
4.6. Plasmid Transfcetion and Luciferase Assay
4.7. Preparation of Cell Lysates and Immunoblotting Analysis
4.8. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| DAA | dehydroabietic acid |
| iNOS | inducible nitric oxide synthase |
| COX | cycloocygenase |
| TNF | tumor necrosis factor |
| PAMPs | pattern-associated molecular patterns |
| PRRs | pattern-recognition receptors |
| TLRs | toll-like receptors |
| MyD88 | myeloid differentiation primary response 88 |
| IKK | IκB kinase |
| IL | interleukin |
| MAPKs | mitogen-activated protein kinases |
| ERK | extracellular signal-regulated kinase |
| JNK | c-Jun N-terminal kinase |
| STAT | signal transducer and activator of transcription protein |
| MEK | MAPK/ERK kinase |
| MKK | mitogen-activated protein kinase kinase |
| TAK1 | transforming growth factor beta-activated kinase 1 |
| LPS | lipopolysaccharide |
| poly(I:C) | polyinosinic-polycytidylic acid |
| PGN | peptidoglycan |
| NO | nitric oxide |
| L-NAME | NG-nitro-L-arginine methyl ester |
| PEI | polyethylenimine |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| FBS | fetal bovine serum |
| EDTA | ethylenediaminetetraacetic acid |
| EGTA | ethyleneglycotetraacetic acid |
References
- Wake, A.; Morgan, H.R. Inflammation, as a Defense Mechanism. In Host-Parasite Relationships and the Yersinia Model; Springer: New York, NY, USA, 1986; pp. 159–163. [Google Scholar]
- Mogensen, T.H. Pathogen recognition and inflammatory signaling in innate immune defenses. Clin. Microbiol. Rev. 2009, 22, 240–273. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Yi, Y.S.; Kim, M.Y.; Cho, J.Y. Role of ginsenosides, the main active components of Panax ginseng, in inflammatory responses and diseases. J. Ginseng Res. 2017, 41, 435–443. [Google Scholar] [CrossRef]
- Bagaitkar, J. Cellular dynamics of resolving inflammation. Blood 2014, 124, 1701–1703. [Google Scholar] [CrossRef]
- Helming, L. Inflammation: Cell recruitment versus local proliferation. Curr. Biol. 2011, 21, R548–R550. [Google Scholar] [CrossRef]
- Kawai, T.; Akira, S. The roles of TLRs, RLRs and NLRs in pathogen recognition. Int. Immunol. 2009, 21, 317–337. [Google Scholar] [CrossRef]
- Christmas, P. Toll-like receptors: Sensors that detect infection. Nat. Educ. 2010, 3, 85. [Google Scholar]
- Ivashkiv, L.B. Inflammatory signaling in macrophages: Transitions from acute to tolerant and alternative activation states. Eur. J. Immunol. 2011, 41, 2477–2481. [Google Scholar] [CrossRef] [PubMed]
- Lowell, C.A. Src-family and Syk kinases in activating and inhibitory pathways in innate immune cells: Signaling cross talk. Cold Spring Harb. Perspect. Biol. 2010, a002352. [Google Scholar] [CrossRef]
- Kanarek, N.; Ben-Neriah, Y. Regulation of NF-κB by ubiquitination and degradation of the IκBs. Immunol. Rev. 2012, 246, 77–94. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.S.; Jeong, D.; Nam, G.; Yi, Y.-S.; Yoon, D.H.; Kim, T.W.; Park, Y.C.; Hwang, H.; Rhee, M.H.; Hong, S. AP-1 pathway-targeted inhibition of inflammatory responses in LPS-treated macrophages and EtOH/HCl-treated stomach by Archidendron clypearia methanol extract. J. Ethnopharmacol. 2013, 146, 637–644. [Google Scholar] [CrossRef] [PubMed]
- Newton, K.; Dixit, V.M. Signaling in innate immunity and inflammation. Cold Spring Harb. Perspect. Biol. 2012, a006049. [Google Scholar] [CrossRef] [PubMed]
- Hess, J.; Angel, P.; Schorpp-Kistner, M. AP-1 subunits: Quarrel and harmony among siblings. J. Cell Sci. 2004, 117, 5965–5973. [Google Scholar] [CrossRef]
- Qiao, Y.; He, H.; Jonsson, P.; Sinha, I.; Zhao, C.; Dahlman-Wright, K. AP-1 is a key regulator of proinflammatory cytokine TNFα-mediated triple-negative breast cancer progression. J. Biol. Chem. 2016, 291, 5068–5079. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; Yang, Y.; Kwak, Y.S.; Song, G.G.; Kim, M.Y.; Rhee, M.H.; Cho, J.Y. Ginsenoside Rc from Panax ginseng exerts anti-inflammatory activity by targeting TANK-binding kinase 1/interferon regulatory factor-3 and p38/ATF-2. J. Ginseng Res. 2017, 41, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Yi, Y.S.; Son, Y.J.; Han, S.Y.; Kim, D.H.; Nam, G.; Hossain, M.A.; Kim, J.H.; Park, J.; Cho, J.Y. BIOGF1K, a compound K-rich fraction of ginseng, plays an antiinflammatory role by targeting an activator protein-1 signaling pathway in RAW264.7 macrophage-like cells. J. Ginseng Res. 2018, 42, 233–237. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Zhang, S.; Geng, Z.; Wang, D.; Liu, F.; Zhang, M.; Bian, H.; Xu, W. Analysis of abietic acid and dehydroabietic acid residues in raw ducks and cooked ducks. Poult. Sci. 2014, 93, 2663–2667. [Google Scholar] [CrossRef]
- Tanaka, R.; Tokuda, H.; Ezaki, Y. Cancer chemopreventive activity of “rosin” constituents of Pinus spez. and their derivatives in two-stage mouse skin carcinogenesis test. Phytomedicine 2008, 15, 985–992. [Google Scholar] [CrossRef]
- Gonçalves, M.D.; Bortoleti, B.; Tomiotto-Pellissier, F.; Miranda-Sapla, M.M.; Assolini, J.P.; Carloto, A.C.M.; Carvalho, P.; Tudisco, E.T.; Urbano, A.; Ambrósio, S.R. Dehydroabietic acid isolated from Pinus elliottii exerts in vitro antileishmanial action by pro-oxidant effect, inducing ROS production in promastigote and downregulating Nrf2/ferritin expression in amastigote forms of Leishmania amazonensis. Fitoterapia 2018, 128, 224–232. [Google Scholar] [CrossRef]
- Kang, M.-S.; Hirai, S.; Goto, T.; Kuroyanagi, K.; Lee, J.-Y.; Uemura, T.; Ezaki, Y.; Takahashi, N.; Kawada, T. Dehydroabietic acid, a phytochemical, acts as ligand for PPARs in macrophages and adipocytes to regulate inflammation. Biochem. Biophys. Res. Commun. 2008, 369, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Zhaoyu, L.; Xiangming, F.; Chunyi, L.; Jiayu, P.; Lu, S.; Jitao, C.; Liangcai, C.; Jifang, L. Abietic acid attenuates allergic airway inflammation in a mouse allergic asthma model. Int. Immunopharmacol. 2016, 38, 261–266. [Google Scholar] [CrossRef]
- González, M.A.; Correa-Royero, J.; Agudelo, L.; Mesa, A.; Betancur-Galvis, L. Synthesis and biological evaluation of abietic acid derivatives. Eur. J. Med. Chem. 2009, 44, 2468–2472. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Kim, M.-Y.; Kim, J.-H.; Cho, J.Y. Fisetin suppresses macrophage-mediated inflammatory responses by blockade of Src and Syk. Biomol. Ther. 2015, 23, 414. [Google Scholar] [CrossRef]
- Byeon, S.E.; Yi, Y.-S.; Oh, J.; Yoo, B.C.; Hong, S.; Cho, J.Y. The role of Src kinase in macrophage-mediated inflammatory responses. Mediat. Inflamm. 2012, 2012. [Google Scholar] [CrossRef]
- Lee, J.-O.; Yang, W.S.; Park, J.G.; Jeong, D.; Kim, H.G.; Yoon, K.D.; Aravinthan, A.; Kim, J.-H.; Kim, E.; Cho, J.Y. Src and Syk contribute to the anti-inflammatory activities of Achyranthes aspera ethanolic extract. J. Ethnopharmacol. 2017, 206, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Costa, M.S.; Rego, A.; Ramos, V.; Afonso, T.B.; Freitas, S.; Preto, M.; Lopes, V.; Vasconcelos, V.; Magalhães, C.; Leão, P.N. The conifer biomarkers dehydroabietic and abietic acids are widespread in Cyanobacteria. Sci. Rep. 2016, 6, 23436. [Google Scholar] [CrossRef]
- Kim, J.; Kang, Y.-G.; Lee, J.-Y.; Choi, D.-H.; Cho, Y.-U.; Shin, J.-M.; Park, J.S.; Lee, J.H.; Kim, W.G.; Seo, D.B. The natural phytochemical dehydroabietic acid is an anti-aging reagent that mediates the direct activation of SIRT1. Mol. Cell. Endocrinol. 2015, 412, 216–225. [Google Scholar] [CrossRef]
- Leandro, L.F.; Cardoso, M.J.O.; Silva, S.D.C.; Souza, M.G.M.; Veneziani, R.C.S.; Ambrosio, S.R.; Martins, C.H.G. Antibacterial activity of Pinus elliottii and its major compound, dehydroabietic acid, against multidrug-resistant strains. J. Med. Microbiol. 2014, 63, 1649–1653. [Google Scholar] [CrossRef]
- Takahashi, N.; Yao, R.; Kang, M.S.; Senda, M.; Ando, C.; Nishimura, K.; Goto, T.; Hirai, S.; Ezaki, Y.; Kawada, T. Dehydroabietic acid activates peroxisome proliferator-activated receptor-γ and stimulates insulin-dependent glucose uptake into 3T3-L1 adipocytes. BioFactors 2011, 37, 309–314. [Google Scholar] [CrossRef]
- Okutani, D.; Lodyga, M.; Han, B.; Liu, M. Src protein tyrosine kinase family and acute inflammatory responses. Am. J. Physiol. Lung Cell. Mol. Physiol. 2006, 291, L129–L141. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S.; Moon, C.; Lee, H.W.; Park, E.-M.; Cho, M.-S.; Kang, J.L. Src tyrosine kinases mediate activations of NF-κB and integrin signal during lipopolysaccharide-induced acute lung injury. J. Immunol. 2007, 179, 7001–7011. [Google Scholar] [CrossRef] [PubMed]
- Jalal, D.I.; Kone, B.C. Src Activation of NF-κB Augments IL-1β–Induced Nitric Oxide Production in Mesangial Cells. J. Am. Soc. Nephrol. 2006, 17, 99–106. [Google Scholar] [CrossRef]
- Ley, K. Fueling the Fire: Src Family Kinases Drive Inflammation; Rockefeller University Press: New York, NY, USA, 2014. [Google Scholar]
- Jiao, H.; Zhang, Y.; Yan, Z.; Wang, Z.-G.; Liu, G.; Minshall, R.D.; Malik, A.B.; Hu, G. Caveolin-1 Tyr14 phosphorylation induces interaction with TLR4 in endothelial cells and mediates MyD88-dependent signaling and sepsis-induced lung inflammation. J. Immunol. 2013, 1300873. [Google Scholar] [CrossRef]
- Geahlen, R.L. Getting Syk: Spleen tyrosine kinase as a therapeutic target. Trends Pharmacol. Sci. 2014, 35, 414–422. [Google Scholar] [CrossRef] [PubMed]
- Takada, Y.; Aggarwal, B.B. TNF activates Syk protein tyrosine kinase leading to TNF-induced MAPK activation, NF-κB activation, and apoptosis. J. Immunol. 2004, 173, 1066–1077. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, D.; Gutierrez, H.; Gavalda, N.; O’Keeffe, G.; Hay, R.; Davies, A.M. Nuclear factor-κB activation via tyrosine phosphorylation of inhibitor κB-α is crucial for ciliary neurotrophic factor-promoted neurite growth from developing neurons. J. Neurosci. 2007, 27, 9664–9669. [Google Scholar] [CrossRef] [PubMed]
- Sakurai, H. Targeting of TAK1 in inflammatory disorders and cancer. Trends Pharmacol. Sci. 2012, 33, 522–530. [Google Scholar] [CrossRef]
- Kim, H.G.; Choi, S.; Lee, J.; Hong, Y.H.; Jeong, D.; Yoon, K.; Yoon, D.H.; Sung, G.H.; Lee, S.; Hong, S.; et al. Src is a prime target inhibited by Celtis choseniana Methanol extract in its anti-inflammatory action. Evid. Based Complement. Alternat. Med. 2018, 2018, 3909038. [Google Scholar] [CrossRef]
- Kim, Y.J.; Deok, J.; Kim, S.; Yoon, D.H.; Sung, G.H.; Aravinthan, A.; Lee, S.; Lee, M.N.; Hong, S.; Kim, J.H.; et al. Anti-inflammatory effect of Piper attenuatum methanol extract in LPS-stimulated inflammatory responses. Evid. Based Complement. Alternat. Med. 2017, 2017, 4606459. [Google Scholar] [CrossRef]
- Lamb, A.; Yang, X.D.; Tsang, Y.H.N.; Li, J.D.; Higashi, H.; Hatakeyama, M.; Peek, R.M.; Blanke, S.R.; Chen, L.F. Helicobacter pylori CagA activates NF-κB by targeting TAK1 for TRAF6-mediated Lys 63 ubiquitination. EMBO Rep. 2009, 10, 1242–1249. [Google Scholar]
- Pathak, S.; Borodkin, V.S.; Albarbarawi, O.; Campbell, D.G.; Ibrahim, A.; Van Aalten, D.M. O-GlcNAcylation of TAB1 modulates TAK1-mediated cytokine release. EMBO J. 2012, 31, 1394–1404. [Google Scholar] [CrossRef]
- Gu, W.; Miao, T.-T.; Hua, D.-W.; Jin, X.-Y.; Tao, X.-B.; Huang, C.-B.; Wang, S.-F. Synthesis and in vitro cytotoxic evaluation of new 1H-benzo [d] imidazole derivatives of dehydroabietic acid. Bioorganic Med. Chem. Lett. 2017, 27, 1296–1300. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.-Z.; Liang, G.-B.; Huang, X.-C.; Zhang, B.; Zhou, M.-M.; Liao, Z.-X.; Wang, H.-S. Discovery of dehydroabietic acid sulfonamide based derivatives as selective matrix metalloproteinases inactivators that inhibit cell migration and proliferation. Eur. J. Med. Chem. 2017, 138, 979–992. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.Y.; Hong, J.T.; Han, S.B.; Park, Y.H.; Son, D.J. Effect of Ixeris dentata Nakai extract on nitric oxide production and prostaglandin E2 generation in LPS-stimulated RAW264. 7 Cells. Immune Netw. 2015, 15, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Byeon, S.E.; Chung, J.Y.; Lee, Y.G.; Kim, B.H.; Kim, K.H.; Cho, J.Y. In vitro and in vivo anti-inflammatory effects of taheebo, a water extract from the inner bark of Tabebuia avellanedae. J. Ethnopharmacol. 2008, 119, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Bak, M.J.; Truong, V.-L.; Ko, S.-Y.; Nguyen, X.N.G.; Jun, M.; Hong, S.-G.; Lee, J.-W.; Jeong, W.-S. Induction of Nrf2/ARE-mediated cytoprotective genes by red ginseng oil through ASK1–MKK4/7–JNK and p38 MAPK signaling pathways in HepG2 cells. J. Ginseng Res. 2016, 40, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.K.; Lee, H.S.; Cho, J.Y.; Shin, W.C.; Rhee, M.H.; Kim, T.G.; Kang, J.H.; Kim, S.H.; Hong, S.; Kang, S.Y. Inhibitory effect of curcumin on nitric oxide production from lipopolysaccharide-activated primary microglia. Life Sci. 2006, 79, 2022–2031. [Google Scholar] [CrossRef]
- Burnette, W.N. “Western blotting”: Electrophoretic transfer of proteins from sodium dodecyl sulfate-polyacrylamide gels to unmodified nitrocellulose and radiographic detection with antibody and radioiodinated protein A. Anal. Biochem. 1981, 112, 195–203. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5′ to 3′) | |
|---|---|---|
| iNOS | F | CCCTTCCGAAGTTTCTGGCAGCAG |
| R | GGCTGTCAGAGCCTCGTGGCTTTGG | |
| TNF-α | F | TTGACCTCAGCGCTGAGTTG |
| R | CCTGTAGCCCACGTCGTAGC | |
| COX-2 | F | CACTACATCCTGACCCACTT |
| R | ATGCTCCTGCTTGAGTATGT | |
| GAPDH | F | CACTCACGGCAAATTCAACGGCA |
| R | GACTCCACGACATACTCAGCAC |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, E.; Kang, Y.-G.; Kim, Y.-J.; Lee, T.R.; Yoo, B.C.; Jo, M.; Kim, J.H.; Kim, J.-H.; Kim, D.; Cho, J.Y. Dehydroabietic Acid Suppresses Inflammatory Response Via Suppression of Src-, Syk-, and TAK1-Mediated Pathways. Int. J. Mol. Sci. 2019, 20, 1593. https://doi.org/10.3390/ijms20071593
Kim E, Kang Y-G, Kim Y-J, Lee TR, Yoo BC, Jo M, Kim JH, Kim J-H, Kim D, Cho JY. Dehydroabietic Acid Suppresses Inflammatory Response Via Suppression of Src-, Syk-, and TAK1-Mediated Pathways. International Journal of Molecular Sciences. 2019; 20(7):1593. https://doi.org/10.3390/ijms20071593
Chicago/Turabian StyleKim, Eunji, Young-Gyu Kang, Yong-Jin Kim, Tae Ryong Lee, Byong Chul Yoo, Minkyeong Jo, Ji Hye Kim, Jong-Hoon Kim, Donghyun Kim, and Jae Youl Cho. 2019. "Dehydroabietic Acid Suppresses Inflammatory Response Via Suppression of Src-, Syk-, and TAK1-Mediated Pathways" International Journal of Molecular Sciences 20, no. 7: 1593. https://doi.org/10.3390/ijms20071593
APA StyleKim, E., Kang, Y. -G., Kim, Y. -J., Lee, T. R., Yoo, B. C., Jo, M., Kim, J. H., Kim, J. -H., Kim, D., & Cho, J. Y. (2019). Dehydroabietic Acid Suppresses Inflammatory Response Via Suppression of Src-, Syk-, and TAK1-Mediated Pathways. International Journal of Molecular Sciences, 20(7), 1593. https://doi.org/10.3390/ijms20071593