2.1. A Pilot Experiment
As was emphasized in the introduction, the experimental data on the inhalation toxicity of NiO-NPs reported so far have been obtained [
18,
19,
20,
21] for exposures that were not long-term and in this respect not comparable, even for the short lifespan of rats, with possible occupational exposure durations that are of primary interest in the area of preventive industrial toxicology. Indeed, if we assume that the average lifespan of a laboratory rat is equal to 30 months, inhalation experiments that last 4 weeks (i.e., only about 3% of the total lifespan) are negligibly short in comparison with a human working lifetime (which, e.g., in Russia amounts, even under hazardous occupational conditions, to ~50% of the standard human lifetime). Moreover, a tolerable single four-hour exposure of rats in “nose only” inhalation setups is much shorter than the working day, even if it is reduced under hazardous conditions. This circumstance further shortens the total chronic exposure in experiments compared with actual occupational contexts.
For these reasons, we designed a chronic inhalation experiment intended to last 3–10 months and involving an average concentration of 1.0 mg/m
3 of NiO-NPs in the air inhaled by rats. This exposure concentration was considered to be moderate and to allow for the position of regulatory toxicology in relation to metallic nickel and its water-insoluble compounds in workroom air (for example, the US OSHA established a PEL equal just to 1.0 mg/m
3 as a TWA concentration). Assuming that any chemical compound of nickel is likely to be more toxic compared with its iron-containing counterpart, we expected that, for nano-sized nickel-oxide particles, this concentration would be even more toxicologically effective (rather than being innocuous) than for Fe
2O
3 iron oxide nanoparticles in about the same concentration (1.14 ± 0.01 mg/m
3) tested by us previously in a similar inhalation experiment [
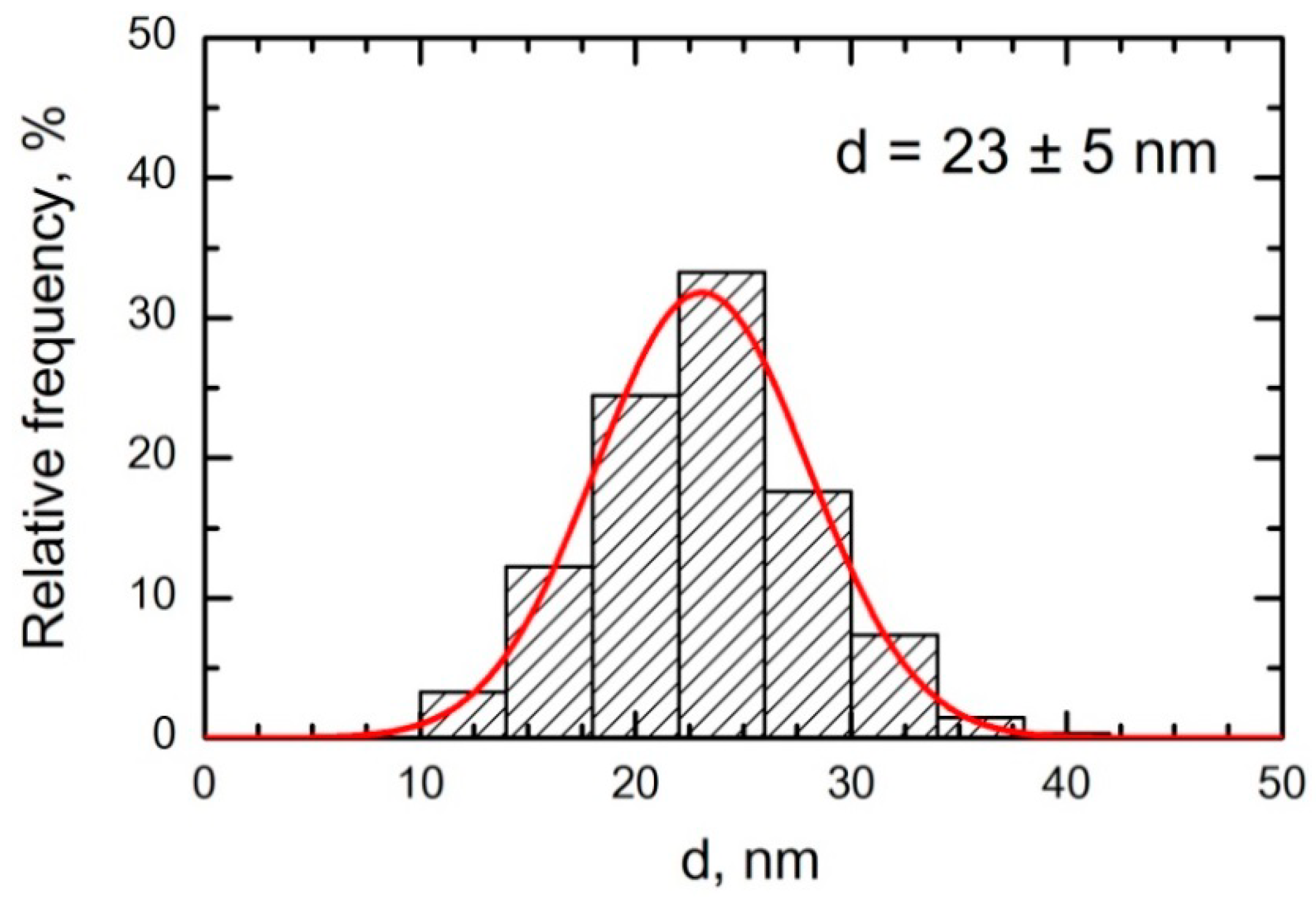
25]. At the same time, we assumed that the contrary role as a factor that determines the comparative toxicity of these two metal-oxide nanoparticle species would be the nearly twice as great diameter of NiO-NPs (23 ± 5 nm) in comparison to Fe
2O
3-NPs (14 ± 4 nm).
Nevertheless, the expectation of a substantial toxicity of nickel-oxide nanoparticles in this concentration unexpectedly came not only true but was even exceeded so much that already after the first 1–2 exposures the clinical picture of an acute intoxication became evident. We were afraid, therefore, that the animals would not live long enough for a chronic intoxication to develop in them. We thus decided to interrupt the experiment after only five daily exposures and to begin a new one with the average concentration of NiO-NPs lowered 4–5 times (see
Section 3).
However, we have measured in these rats at least some of the indices that were planned by the design of the chronic experiment (see
Section 3) in order to use them as predictors of the results in the longer-term experiment. Therefore, we regard the first inhalation experiment as a pilot rather than a failure.
As follows from
Table 1 (which presents mostly those indices for which the exposed group differed from the controls statistically significantly), even though the total exposure period was short, certain functional effects of toxic action did manifest themselves. We noted the following:
- -
an increase in liver mass typical of subchronic and chronic intoxications with practically all metal-oxide nanoparticles ever studied by us;
- -
elevated release of lactate dehydrogenase in the blood due, probably, to the toxic damage to the liver cells and to the pulmonary macrophages;
- -
leukocytosis;
- -
systemic inhibition of the oxidation-reduction energy metabolism, the integrated cytochemical indicator of which is the suppression of succinate dehydrogenase activity in blood lymphocytes, which we repeatedly observed in experiments involving practically any of the toxic metals in any form [
23,
26,
27,
28];
- -
enhanced lipid peroxidation judging by the increased concentration of malondialdehyde in the blood;
- -
stimulation of erythropoiesis suggested by an increase in the erythrocyte count (along with an increased proportion of reticulocytes), hematocrit, and hemoglobin content of the blood.
Such a quasi-beneficial hematological effect is astonishing not only because various metal-induced intoxications go along with this or because of the degree of anemia with different mechanisms of development but also because intraperitoneal subchronic intoxication caused by similar NiO-NPs was previously found by us [
23] to bring about just such anemia. We will consider this apparent contradiction below (
Section 2.2) when discussing the findings of the main chronic inhalation experiment, in which such stimulation was again observed. As for the pilot experiment being considered, it is noteworthy that even one week after its termination the red blood still displayed a response, although it was less pronounced. For instance, the proportion of reticulocytes was equal to 25.86 ± 3.88‰ against 15.22 ± 1.37‰ in the control group (
p < 0.05).
In 24 h after the last exposure, a dramatic increase in both the bronchoalveolar lavage fluid (BALF) total cell count and the number of alveolar macrophages (AMs) and neutrophil leukocytes (NLs) in comparison with the corresponding indices in the control group was observed (
Table 2). The NL/AM ratio was, however, somewhat decreased, which is not characteristic of the immediate pulmonary response to the deposition of any particles and, in particular, of highly cytotoxic ones [
1,
23,
26,
27,
28,
29,
30,
31,
32]. Three weeks later, the differences between the NiO-NP-exposed and control groups in the BALF total cell count and the AM numbers remained statistically significant although somewhat decreased, while the initial neutrophilic response disappeared.
Recently we published (for the first time in nanotoxicology, as far as we know) a cytological analysis of tissue imprints from organs of rats exposed subchronically to intraperitoneally injected metal-oxide nanoparticles in experiments with Al
2O
3-NPs, TiO
2-NPs, and SiO
2-NPs administered separately or in different combinations [
27]. That analysis proved to be rather informative. Simultaneously we explored the potentialities of this approach in the pilot inhalation experiment under discussion now and obtained some interesting results (
Table 3).
In particular, the following findings are noteworthy:
- -
the lungs had increased proportions of explicitly degenerated alveolar macrophages and of eosinophils;
- -
the liver had increased proportions of explicitly degenerated hepatocytes, activated Kupffer’s cells, neutrophils, and eosinophils;
- -
the kidneys had increased proportions of explicitly degenerated cells of proximal (and to a much lesser degree, distal) convoluted tubules;
- -
the spleen displayed a decrease in the proportion of mature lymphocytes together with prolymphocytes and a respective increase in the proportion of other white cells, especially eosinophils.
Thus, notwithstanding rather a short inhalation period and a seemingly low exposure level, we can see some typical adverse effects on target organs along with hyperergic toxic inflammation evidenced by a marked eosinophilic response. It would be an enticing hypothesis to connect the latter with the well-known systemic nickel allergy syndrome if a similar response had not been observed in the above-mentioned experiments with three other NPs of elements, none of which is, unlike nickel, a recognized potent sensitizer [
27]. In this respect, we should mention the urgency of the well-recognized general problem of “nanoparticles and allergy” [
33], which has not yet been solved satisfactorily.
2.2. A Long-Term Low Level Exposure
As will be described in detail in
Section 3, in this experiment, the NiO-NP concentrations were approximately four times lower compared with the pilot one (0.23 ± 0.01 mg/m
3 instead of 1.0 ± 0.1 mg/m
3). In other respects the mode and duration of weekly exposures were the same (4 h a day, 5 times a week), but the total exposure period was prolonged up to 3, 6, or 10 months.
As follows from the comparison of the data presented in
Table 2 and
Table 4, the shifts in the cytological characteristics of the BALF 24 h after the final inhalation exposure to nanoparticles in the chronic inhalation experiment were, unlike the pilot one, of the usual character (an increase in total cell count due to the recruitment of both AMs and NLs with an increased NL/AM ratio in comparison with the control group). Believing that such shifts were determined mainly by the response to the last portions of particles deposited in the lower airways, we did not expect them to increase by increasing the overall duration of the exposure period. Indeed, the cytological characteristic of the BALF after the 6-month and 10-month exposure was in principle the same. However, over both of these experimental periods, the total cell counts and the AM and NL numbers were much lower than after the 3-month exposure.
Our studies demonstrated long ago that the intensity of the protective compensatory recruitment of alveolar macrophages and, particularly, neutrophil leukocytes into the lower airways is higher as the particles become more cytotoxic and thus as the mass of produced macrophage breakdown products (MBPs) regulating this recruitment increases [
29,
34]. However, the recruitment of new echelons of phagocytizing cells in response to an intratracheal instillation of one and the same dose of sterile MBPs (obtained by freezing-thawing or ultrasonication of a macrophage culture in aseptic conditions) greatly depends on the total reactivity of the organism, which may change under the effect of various factors [
31]. Assumingly, in the current experiment, a similar change in the reactivity of the organism took place. This change might be, on the one hand, a result of the chronic intoxication suppressing this reactivity and, on the other hand, a manifestation of the organism’s partial adaptation to the impact of inhaled particles.
At the same time, the increased activity of the BALF supernatant’s enzymes (
Table 5) is likely to reflect the destruction of cells on the free surface of the lungs’ pulmonary region under the impact of cytotoxic particles. Since the exposure to NiO-NPs was constant, it is not surprising that these biochemical indices were relatively stable in all three experimental periods.
In
Table 6 we present all functional indices for the organism’s status measured in both exposed and control (sham-exposed) rats to find whether any of them revealed statistically significant and/or consistent shifts that might be attributed to the NiO-NP toxic action.
In fact, like the above-discussed cytological BALF indices, the systemic toxic action effects of inhaled nanoparticles presented in this table suggest that the rat organism was gradually adapting to some extent to the low-level inhalation exposure. In this respect, of particular interest are the red blood indices. Previously we found [
23] that under a subchronic intraperitoneal exposure to the NiO-NPs they generally developed anemia. At the same time, under low-level inhalation burdens we can see signs of erythropoiesis stimulation both in the short-term pilot experiment (see
Section 2.1) and by the end of the 3rd month in the main experiment. In the latter case, these signs included an elevated hemoglobin content, an increased erythrocyte count with an elevated proportion of reticulocytes, and an elevated hematocrit. However, in the subsequent terms of this investigation, the possible reaction of the marrow bone was evidenced only by a statistically significant increase in the proportion of reticulocytes.
The effect produced by various chemical forms of nickel on erythropoiesis is not something new, although we have not come across any references to such nanoparticle action in this metal. Thus, Sunderman et al. [
35] observed an enhanced generation of erythropoietin with an increase in hematocrit in guinea pigs and rats in response to intrarenal administration of nickel subsulfide. On the contrary, anemia is known to develop in rats under intoxication, for example, with nickel chloride [
36]. Phase-related changes in the red blood indices under a 4-month (every other day) inhalation exposure to micrometer particles of metallic nickel or nickel oxide at a concentration of 350 mg/m
3 were described a long time ago [
37]. An enhancement of erythropoiesis was noted in 2 weeks, lasted for 1.5 months, and then decreased to the initial level or fell below it. Later on, a study involving continuous 3-month inhalation exposure to a rather coarse metallic nickel dust at a concentration of 0.5, 0.1, and 0.02 mg/m
3 also revealed an increase in the erythrocyte count in the peripheral blood after 2.5 months of exposure followed by normalization of this index by the end of the exposure period [
38].
It is interesting that we found a similar alternating pattern of response to chronic inhalation of NiO-NPs for such an important toxic effect as energy metabolism suppression as well. Indeed, the typical decrease in the cytochemical index of SDH activity in blood lymphocytes was pronounced and statistically significant by the end of the 3rd month but was absent in the next month. On the contrary, the intensity of lipid peroxidation estimated by the concentration of malondialdehyde (MDA) in the blood serum increased significantly only towards the end of the long-term exposure period. Meantime, it was significantly reduced in the first term (which is an extremely rare and hard-to-explain effect) and virtually unchanged in the mid-term. No significant changes in reduced glutathione and total SH-groups in the blood-typical effects for different metal intoxications—were observed in this instance.
What is particularly noticeable is the insignificance and inconstancy of many other (including organ-specific) shifts caused by low-level chronic inhalation exposure to NiO-NPs. Thus, none of the usually observed signs of increased inhibition processes in the CNS (longer index of summation of subthreshold impulses, suppression of the behavioral indices of exploratory, and general motor activities) displayed, in the chronic inhalation experiment, any statistically significant difference from the control value. Rather, they generally tended to be oppositely directed, especially at the end of the exposure period. This circumstance is particularly interesting because under inhalation exposure one of the targets for nanoparticles initially deposited in the nasal ways is the brain, to which they move along the fibers of the olfactory nerve [
25,
39,
40,
41]. Indeed, the electron microscopy of olfactory bulb preparations from the rat brain in this experiment revealed both nanoparticle retention (
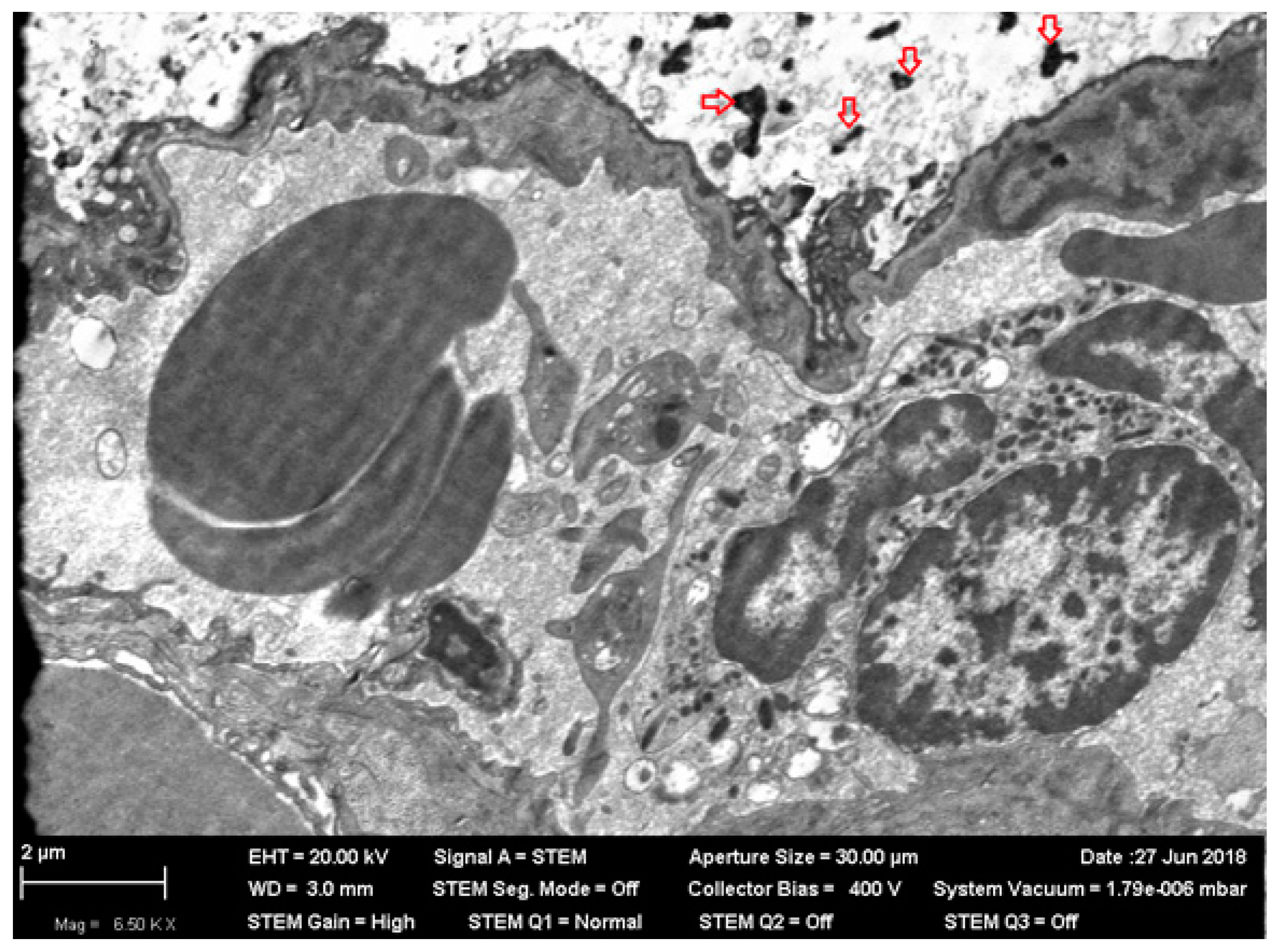
Figure 1) and ultrastructural damage (
Figure 2 and
Figure 3). The averaged nickel content of the homogenized tissues of the entire brain was just slightly increased, which indirectly confirms the local nature of NiO-NP retention limited to the olfactory tract.
There was no noticeable change in the relative mass of the liver, spleen, and kidneys or in indices of liver functioning (the protein and separate protein fractions content in the blood serum and aminotransferase activities) or of kidney functioning (diuresis, urine density, and endogenous creatinine clearance). At the same time, in all three periods of this inhalation exposure, increased bilirubin content in the blood serum and proteinuria after 6 and 10 months of exposure (statistically significant only by the 6th month) was observed.
The low manifestation of the hepato- and nephrotoxicity effects may be explained by the fact that the nickel content of these organs, both total (by the AES data) and as NiO (by EPR-spectroscopy data), virtually did not differ from the control values. Thus, by the end of the 10-month period, for the liver it was equal in the control group to 0.65 ± 0.22 and 0.06 ± 0.02 μg per gram of dry tissue, respectively, while in the exposed group it was equal to 0.73 ± 0.17 and 0.07 ± 0.03 μg/g; for kidneys, it was equal to 0.31 ± 0.08 and 0.03 ± 0.01 μg/g in the control group and 0.35 ± 0.06 and 0.02 ± 0.01 μg/g in the exposed group. No additional accumulation of nickel in the spleen of the exposed rats was observed: 0.25 ± 0.07 μg/g by the AES data and 0.01 ± 0.01 μg/g by the EPR data (against 0.27 ± 0.03 and 0.02 ± 0.00 μg/g in the control group). Meanwhile, both indices of blood nickel content in the exposed rats (0.48 ± 0.05 and 0.53 ± 0.14 μg/g) were higher than the corresponding control values (0.39 ± 0.07 and 0.30 ± 0.07 μg/g), but not by much, and the increase was statistically insignificant.
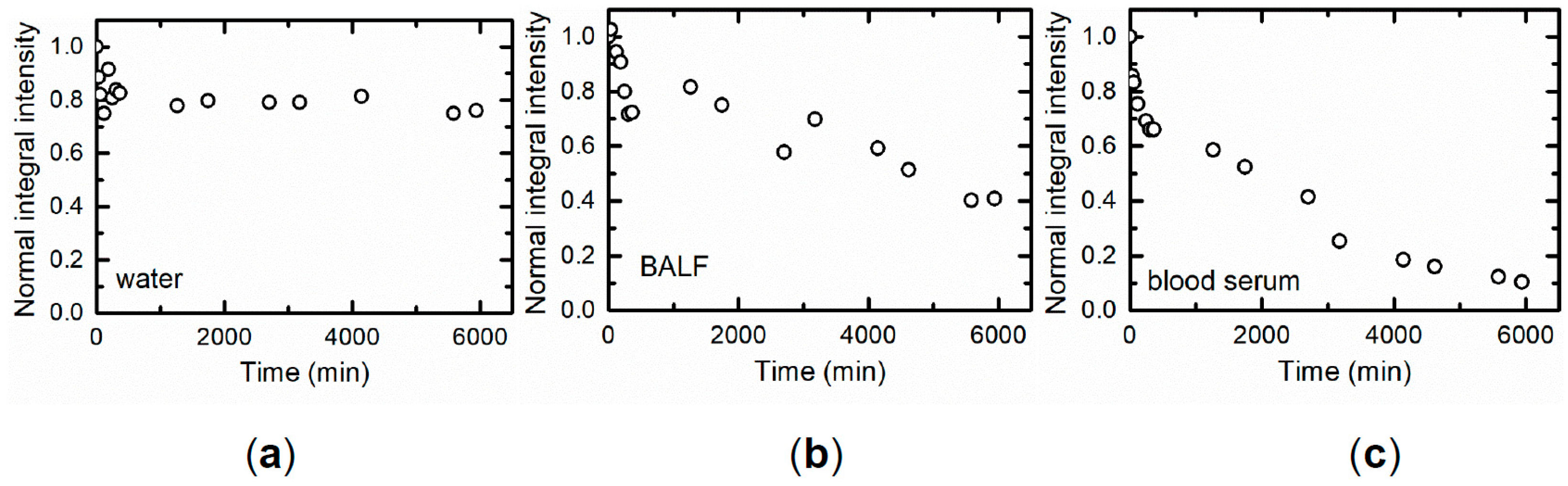
A low chronic accumulation of NiO-NPs in the above-mentioned organs as well as in the lungs (see below) is presumably due mostly to its solubilization in biological milieus, which could be directly proved in vitro (
Figure 4).
It is worth mentioning that in the case of inhalation exposure there is an increase in dry mass of lungs, a typical observation for any experimental pneumoconiosis. In this case, however, this increase grew statistically significant only by the 10th month. By this time, the retention of nickel in the lung tissue was the highest according to both the AES data (3.69 ± 0.35 against 0.96 ± 0.30 μg/kg in the sham-exposed group;
p < 0.05) and the EPR spectroscopy (0.39 ± 0.04 and 0.09 ± 0.03 μg/kg, respectively;
p < 0.05). Nevertheless, electron microscopy of the lungs (
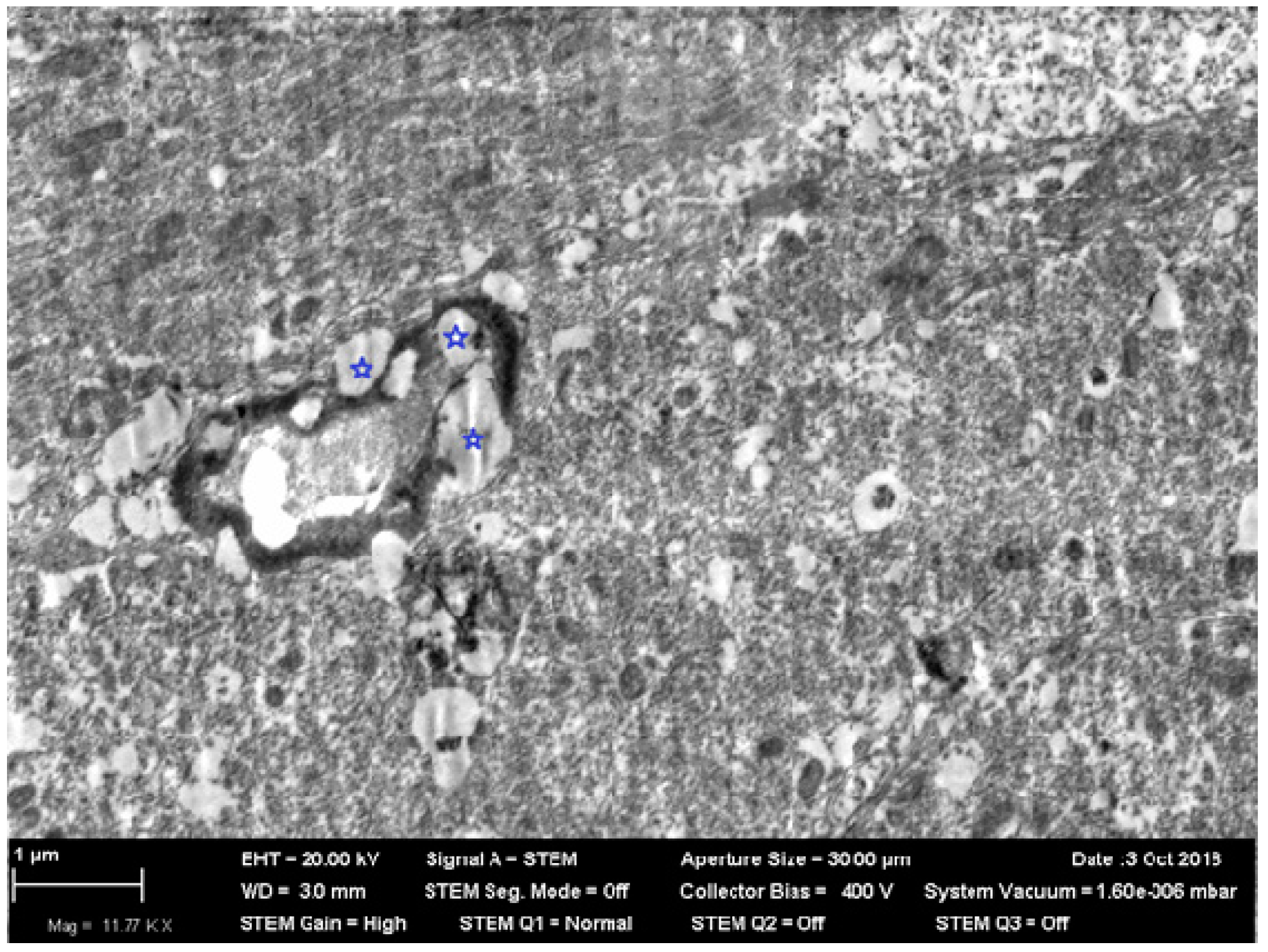
Figure 5) revealed numerous nanoparticles or their nano-sized aggregates in all periods of the study not only as internalized by phagocytic cells but also as located freely inside alveoli, which seems to be a result of their recent deposition from the inhaled air.
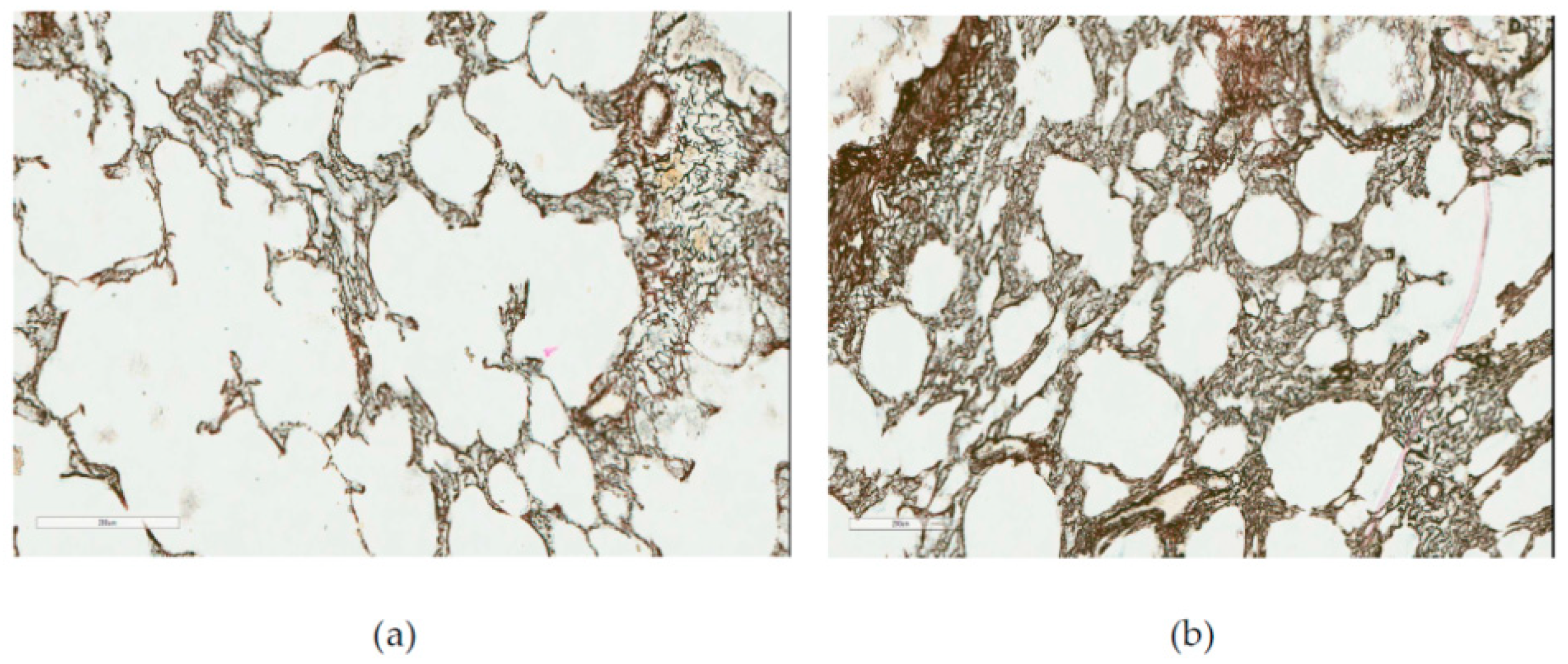
At the same time, a histological investigation of the lung tissue by optical microscopy did not reveal, even towards the end of the experiment, any cellular fibrotic nodules typical of experimental pneumoconioses, or any thickening of interalveolar septae together with their diffuse fibrosis. On the contrary, the septae became thinner or were even destroyed (emphysema), while the reticulin framework was no coarser than in the control rats (
Figure 6).
Besides the above-considered functional indices, all internal organs displayed appreciable, even if moderate, pathological changes in the histological structure confirmed by optical microscopy-based morphometric assessment (
Table 7 and
Table 8). The fact that there was no increase in the proportion of akaryotic hepatocytes after the 3-month exposure is quite consistent with the observation that in this period the retention of nickel in the liver was still negligibly low (according to AES data, 0.10 ± 0.05 against 0.08 ± 0.03 mg/kg in the control group; according to EPR, 0.02 ± 0.02 mg/kg and 0.00, respectively). Thus, the hepatotoxic effect, too, did not display any signs of the organism’s adaptation to the effects of NiO-NPs.
The reduced planimetric ratio of red pulp to white pulp in the spleen usually observed by us in subchronic intraperitoneal experiments with nanoparticles [
1] was found in this inhalation experiment only by the first and last terms of investigation, and this ratio decrease was not statistically significant. However, the response of the white pulp is evidenced by an increase in the average diameter of lymphoid follicles observed in all periods of the experiment. Assumingly, it was a reactive follicular hyperplasia associated not as much with the retention of nanoparticles in this organ as with the probability of an allergic syndrome mentioned above (in the discussion of the data of
Table 3).
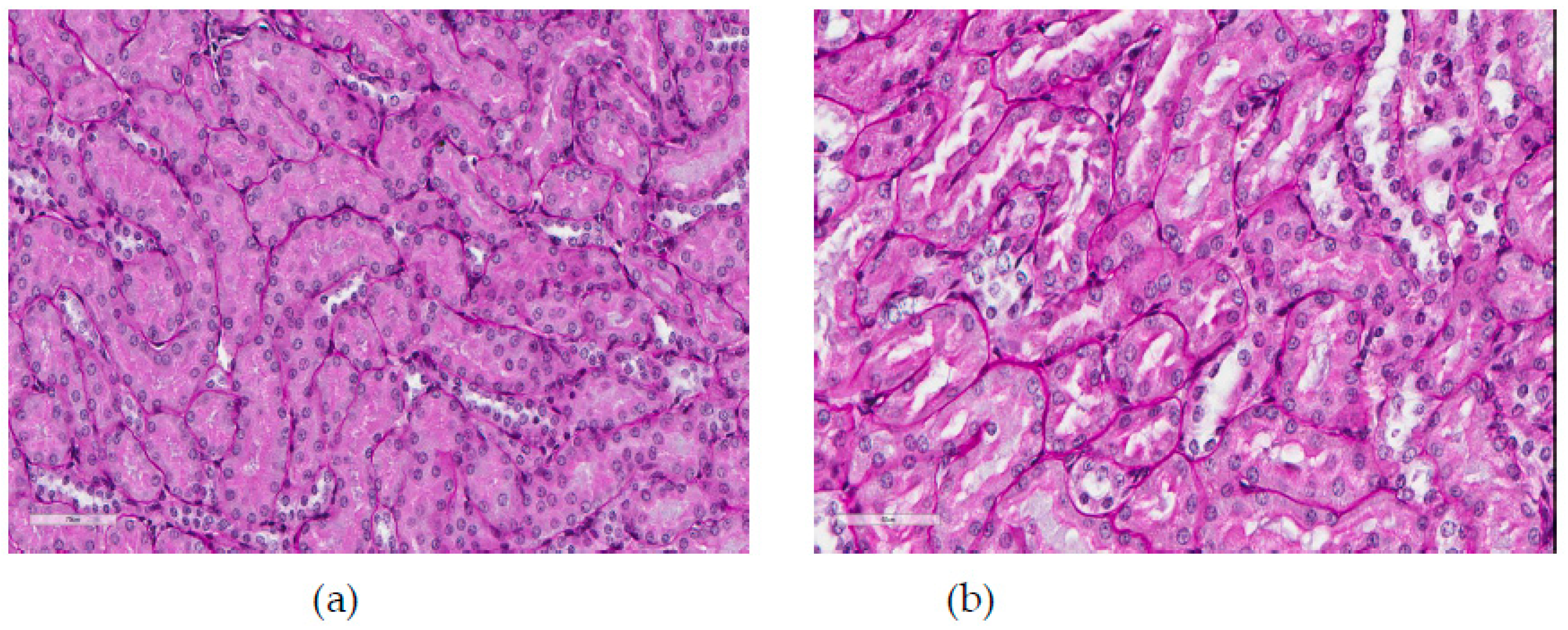
For kidneys, as is common [
1], the most typical adverse effect of NiO-NP toxicity was damage to the tubular epithelium (
Figure 7 and
Table 8). The most obvious signs of such damage are brush border loss in proximal convoluted tubules and full epithelial desquamation. However, although the first of these effects was observed in all terms of the long-term inhalation exposure, it was statistically significant only in the 3-month term. The second of the named effects was statistically significant only in the 10-month term. For this term we only obtained a statistically significant difference in the average diameter of the glomeruli between the exposed and sham-exposed groups. It is easy to see, though, that this indicator in the exposed group was actually the same as in the previous periods, while in the control group it increased unexpectedly. We therefore abstain from its toxicological interpretation.
The cytological changes in tissue imprints (
Table 9) were generally of the same character as in the pilot experiment (
Table 3). At the same time, it is interesting that for some important indices of this kind (for example, the percentage of eosinophils in the liver and spleen imprints), we can see an attenuation of shifts caused by exposure as the duration of exposure increases, although in some cases (in particular, the percentage of degeneratively changed cells in liver and kidney imprints), such attenuation was not observed from the above morphometric characteristics of the histological preparations.



 ,
, 










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}