Physiological and Metabolic Responses of Rice to Reduced Soil Moisture: Relationship of Water Stress Tolerance and Grain Production

 ,
,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Rice Cultivar Selection for Differences in Yield Response to Reduced Soil Water Availability
2.2. Irrigation Treatments Resulted in Distinct Differences in Physiological Responses
2.3. Yield Potential of Cultivars under Non-Stressed Conditions Is Not Predictive of Yields under Severe Water Stress
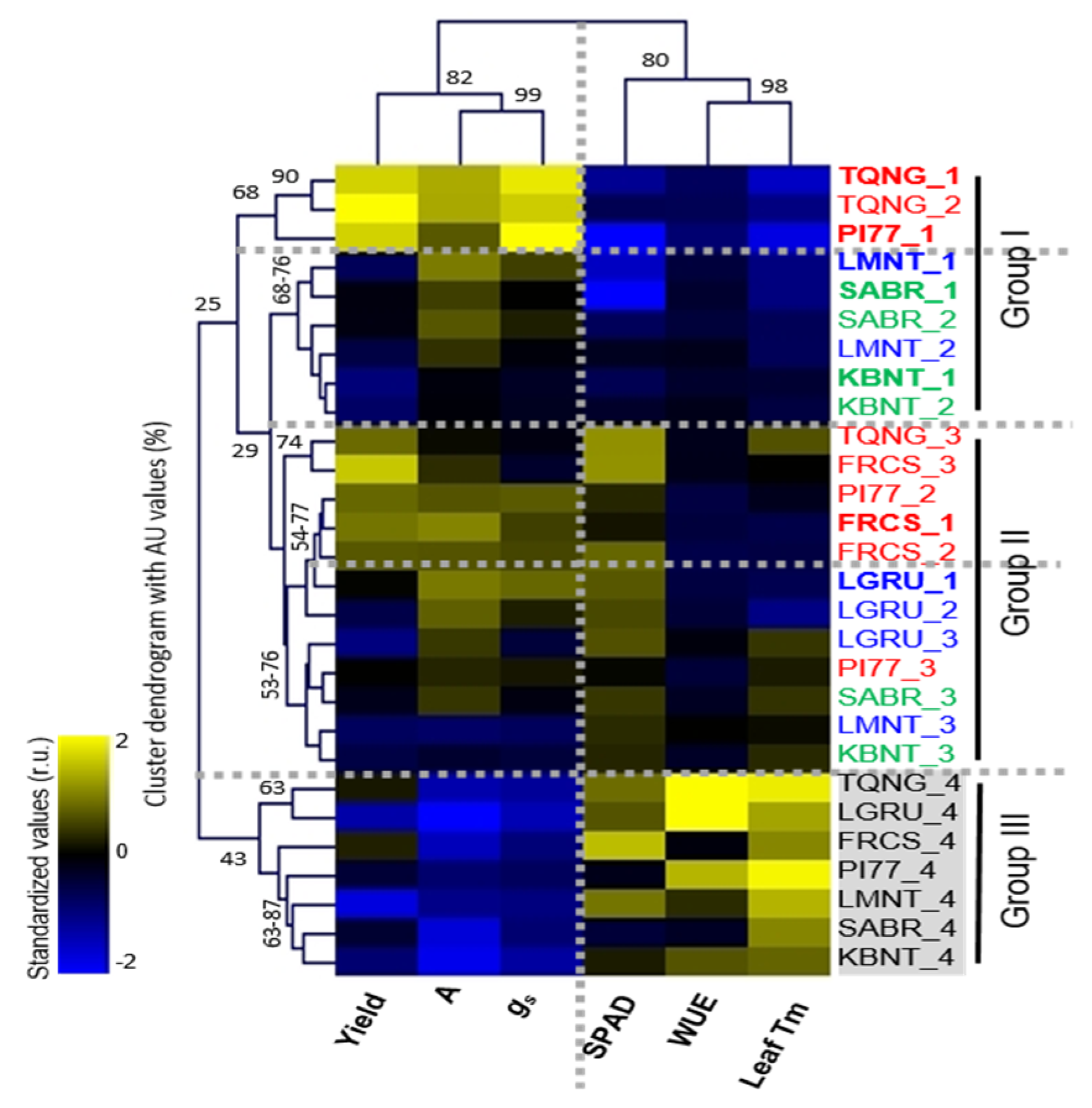
2.4. Association of Physiological Changes among Cultivars That Differ in Yield Response to Water Stress
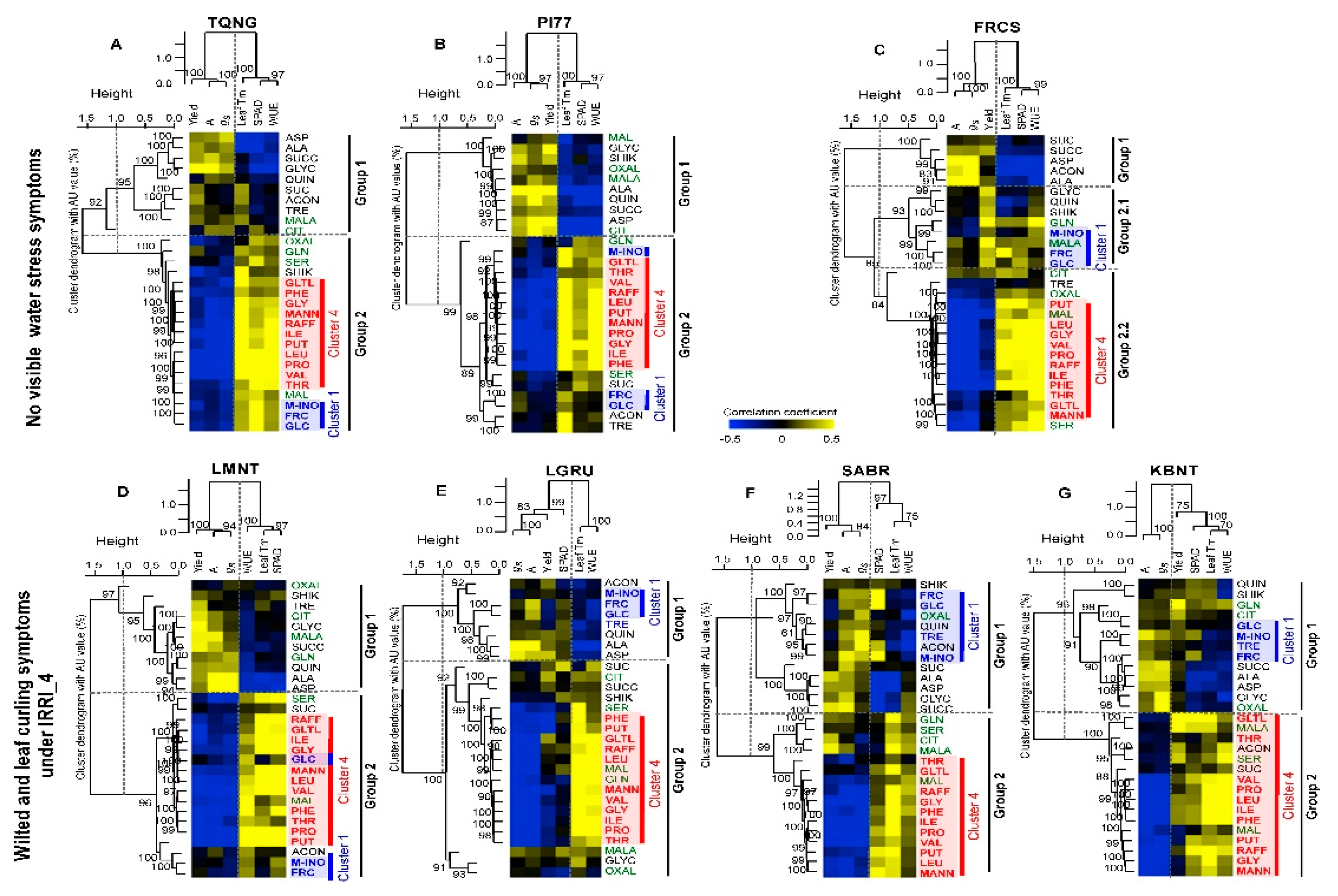
2.5. Metabolic Responses of Seven Cultivars under Reduced Irrigation Levels Resulted in Four Distinctive Cluster Groups
2.6. High vs. Non- or Low Yield Water Stress Response Cultivars Displayed Distinctive Metabolic Patterns
2.7. Cultivars with Little Loss in Yield Due to Reduced Irrigation Levels May have Ability to Regulate Osmo-Protectants
3. Discussion
3.1. Water Management Practice
3.2. Diverse Yield Responses to Water Stress Observed among Different Cultivars
3.3. The Physiological Response of Cultivars to Water Stress
3.4. Cultivar Evaluation: Metabolic Adjustment to Water Stress
3.5. Interrelationship of Physiological and Metabolic Status with Yield Potential and Water Stress
4. Materials and Methods
4.1. Plant Materials And Experimental Conditions
4.2. Field Measurements
4.3. Metabolite Measurements
4.4. Statistical Comparisons
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| VWC | Soil volumetric water content |
| Tm | TemperatureDirectory of open access journals |
| A | Photosynthetic CO2 assimilation |
| gs | Stomatal conductance |
| WUE | Water Use Efficiency |
| IRRI | IRRIgation treatment |
| AWD | Alternate wetting and drying |
| SDI | Subsurface drip irrigation |
| C | Cultivar |
| I | Irrigation |
| C x I | Cultivar by Irrigation |
| RCS | Randomized compete block |
| FC | Field capacity |
| TDT | Time domain transmissometry moisture |
| DAE | Days after emergence |
| SPAD | Relative chlorophyll content |
| VPD | Vapor pressure deficit |
| DW | Dry weight |
| GC-MS | Gas chromatography coupled to mass spectrometry |
| MeV | MultiExperiment Viewer |
| MRVA | Mississippi River Valley Alluvial |
| TQNG | Teqing |
| PI77 | PI312777 |
| LGRU | Lagrue |
| LMNT | Lemont |
| FRCS | Francis |
| SABR | Saber |
| KBNT | Kaybonnet |
| Acon | Aconitate |
| Frc | Fructose |
| Glc | Glucose |
| myo-ino | Myo-inositol |
| Tre | Trehalose |
| Ala | Alanine |
| Asp | Aspartate |
| Succ | Succinate |
| Glyc | Glycerate |
| Quin | Quinate |
| Shik | Shikimate |
| Oxal | Oxalate |
| Ser | Serine |
| Gln | Glutamine |
| Mal | Maltose |
| Cit | Citrate |
| Mala | Malate |
| The | Threonine |
| Gltl | Galactinol |
| Phe | Phenylalanine |
| Ile | Isoleucine |
| Val | Valine |
| Leu | Leucine |
| Pro | Proline |
| Raff | Raffinose |
| Mann | Mannitol |
| Gly | Glycine |
| Put | Putrescine |
References
- Sandhu, N.; Kumar, A. Bridging the rice yield gaps under drought: QTLs, genes, and their use in breeding programs. Agronomy 2017, 7, 27. [Google Scholar] [CrossRef]
- Usda/Ers. Rice: Overview 2017 [Online]. Available online: https://www.ers.usda.gov/topics/crops/rice/ (accessed on 10 April 2019).
- Reba, M.L.; Daniels, M.; Chen, Y.; Sharpley, A.; Bouldin, J.; Teague, T.G.; Daniel, P.; Henry, C.G. A statewide network for monitoring agricultural water quality and water quantity in Arkansas. J. Soil Water Conservation 2013, 68, 45A–49A. [Google Scholar] [CrossRef]
- Massey, J.H.; Walker, T.W.; Anders, M.M.; Smith, M.C.; Avila, L.A. Farmer adaptation of intermittent flooding using multiple-inlet rice irrigation in Mississippi. Agric. Water Manag. 2014, 146, 297–304. [Google Scholar] [CrossRef]
- Ray, D.K.; Mueller, N.D.; West, P.C.; Foley, J.A. Yield trends are insufficient to double global crop production by 2050. PLoS ONE 2013, 8, e66428. [Google Scholar] [CrossRef] [PubMed]
- Camp, C.R. Subsurface drip irrigation: A review. Tran. ASAE 1998, 41, 1353. [Google Scholar] [CrossRef]
- Beecher, H.G.; Dunn, B.W.; Thompson, J.A.; Humphreys, E.; Mathews, S.K.; Timsina, J. Effect of raised beds, irrigation and nitrogen management on growth, water use and yield of rice in south-eastern Australia. Aus. J. Exp. Agr. 2006, 46, 1363–1372. [Google Scholar] [CrossRef]
- Korres, N.E.; Norsworthy, J.K.; Burgos, N.R.; Oosterhuis, D.M. Temperature and drought impacts on rice production: An agronomic perspective regarding short- and long-term adaptation measures. Water Res. and Rural Develop. 2017, 9, 12–27. [Google Scholar] [CrossRef]
- McDonald, P.G.; Fonseca, C.R.; Overton, J.M.; Westoby, M. Leaf-size divergence along rainfall and soil nutrient gradients: Is the method of size-reduction common among clades? Funct. Ecol. 2003, 17, 50–57. [Google Scholar] [CrossRef]
- Okami, M.; Kato, Y.; Kobayashi, N.; Yamagishi, J. Morphological traits associated with vegetative growth of rice (Oryza sativa L.) during the recovery phase after early-season drought. Eur. J. Agron. 2015, 64, 58–66. [Google Scholar] [CrossRef]
- Eldakak, M.; Milad, S.; Nawar, A.; Rohila, J. Proteomics: A biotechnology tool for crop improvement. Front. Plant Sci. 2013, 4, 35. [Google Scholar] [CrossRef]
- Chung, P.J.; Jung, H.; Jeong, D.H.; Ha, S.H.; Choi, Y.D.; Kim, J.K. Transcriptome profiling of drought responsive noncoding RNAs and their target genes in rice. BMC Genomics 2016, 17, 563. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Liao, F.; Wang, F.; Wen, W.; Li, J.; Mei, H.; Luo, L. Identification of Rice Transcription Factors Associated with Drought Tolerance Using the Ecotilling Method. PLoS ONE 2012, 7, e30765. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Lin, S.; Song, Q.; Li, K.; Tao, H.; Huang, J.; Chen, X.; Que, S.; He, H. Genome-wide identification of heat shock proteins (Hsps) and Hsp interactors in rice: Hsp70s as a case study. BMC Genomics 2014, 15, 344. [Google Scholar] [CrossRef] [PubMed]
- Degenkolbe, T.; Do, P.T.; Kopka, J.; Zuther, E.; Hincha, D.K.; Kohl, K.I. Identification of drought tolerance markers in a diverse population of rice cultivars by expression and metabolite profiling. PLoS ONE 2013, 8, e63637. [Google Scholar] [CrossRef] [PubMed]
- Arbona, V.; Manzi, M.; Ollas, C.D.; Gomes-Cadenas, A. Metabolomics as a tool to investigate abiotic stress tolerance in plants. Int. J. Mol. Sci. 2013, 14, 4885–4911. [Google Scholar] [CrossRef]
- Fessenden, M. Metabolomics: Small molecules, single cells. Nature 2016, 540, 153–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alseekh, S.; Bermudez, L.; Haro, L.A.D.; Fernie, A.R.; Carrari, F. Crop metabolomics: From diagnostics to assisted breeding. Metabolomics 2018, 12, 148. [Google Scholar] [CrossRef]
- Swamy, B.M.; Kumar, A. Genomics-based precision breeding approaches to improve drought tolerance in rice. Biotechnol. Adv. 2013, 31, 1308–1318. [Google Scholar] [CrossRef]
- Bernier, J.; Atlin, G.N.; Serraj, R.; Kumar, A.; Spaner, D. Breeding upland rice for drought resistance. J. Sci. Food Agric. 2008, 88, 927–939. [Google Scholar] [CrossRef]
- Zhang, C.; Li, C.; Liu, J.; Lv, Y.; Yu, C.; Li, H.; Zhao, T.; Liu, B. The OsABF1 transcription factor improves drought tolerance by activating the transcription of COR413-TM1 in rice. J. Exp. Bot. 2017, 68, 4695–4707. [Google Scholar] [CrossRef]
- Huang, L.; Wang, Y.; Wang, W.; Zhao, X.; Qin, Q.; Sun, F.; Hu, F.; Zhao, Y.; Li, Z.; Fu, B.; et al. Characterization of Transcription Factor Gene OsDRAP1 Conferring Drought Tolerance in Rice. Front. Plant Sci. 2018, 9, 94. [Google Scholar] [CrossRef] [PubMed]
- Borah, P.; Sharma, E.; Kaur, A.; Chandel, G.; Mohapatra, T.; Kapoor, S.; Khurana, J.P. Analysis of drought-responsive signaling network in two contrasting rice cultivars using transcriptome-based approach. Sci. Rep. 2017, 7, 42131. [Google Scholar] [CrossRef] [PubMed]
- Centritto, M.; Lauteri, M.; Monteverdi, M.C.; Serraj, R. Leaf gas exchange, carbon isotope discrimination, and grain yield in contrasting rice genotypes subjected to water deficits during the reproductive stage. J. Exp. Bot. 2009, 60, 2325–2339. [Google Scholar] [CrossRef] [Green Version]
- Schroeder, J.I.; Allen, G.J.; Hugouvieux, V.; Kwak, J.M.; Waner, D. Guard cell signal transduction. Ann. Rev. Plant Physiol. Plant Mol. Biol. 2001, 52, 627–658. [Google Scholar] [CrossRef] [PubMed]
- Radin, J.W.; Hartung, W.; Kimball, B.A.; Mauney, J.R. Correlation of stomatal conductance with photosynthetic capacity of cotton only in a CO2-enriched atmosphere: Mediation by abscisic acid? Plant Physiol. 1988, 88, 1058–1062. [Google Scholar] [CrossRef] [PubMed]
- Xiong, D.; Chen, J.; Yu, T.; Gao, W.; Ling, X.; Li, Y.; Peng, S.; Huang, J. SPAD-based leaf nitrogen estimation is impacted by environmental factors and crop leaf characteristics. Sci. Rep. 2015, 5, 13389. [Google Scholar] [CrossRef] [Green Version]
- Inamullah; Isoda, A. Adaptive responses of soybean and cotton to water stress II. Changes in CO2 assimilation rate, chlorophyll fluorescence and photochemical reflectance index in relation to leaf temperature. Plant Prod. Sci. 2005, 8, 131–138. [Google Scholar] [CrossRef]
- Silva, M.D.A.; Jifon, J.L.; Silva, J.a.G.D.; Sharma, V. Use of physiological parameters as fast tools to screen for drought tolerance in sugarcane. Braz. J. Plant Physiol. 2007, 19, 193–201. [Google Scholar] [CrossRef]
- Saud, S.; Fahad, S.; Yajun, C.; Ihsan, M.Z.; Hammad, H.M.; Nasim, W.; Amanullah, Jr.; Arif, M.; Albarby, H. Effects of Nitrogen Supply on Water Stress and Recovery Mechanisms in Kentucky Bluegrass Plants. Front. Plant Sci. 2017, 8, 983. [Google Scholar] [CrossRef]
- Syamsia, S.; Idhan, A.; Noerfitryani, N.; Nadir, M.; Reta; Kadir, M. Paddy chlorophyll concentrations in drought stress condition and endophytic fungi application. Earth Environ. Sci. 2018, 156, 012040. [Google Scholar] [CrossRef]
- Sicher, R.C.; Barnaby, J.Y. Impact of carbon dioxide enrichment on the responses of maize leaf transcripts and metabolites to water stress. Physiol. Plant 2012, 144, 238–253. [Google Scholar] [CrossRef]
- Witt, S.; Galicia, L.; Lisec, J.; Cairns, J.; Tiessen, A.; Araus, J.L.; Palacios-Rojas, N.; Fernie, A.R. Metabolic and Phenotypic Responses of Greenhouse-Grown Maize Hybrids to Experimentally Controlled Drought Stress. Mol. Plant 2012, 5, 401–417. [Google Scholar] [CrossRef]
- Barnaby, J.Y.; Kim, M.; Bauchan, G.; Bunce, J.; Reddy, V.; Sicher, R.C. Drought Responses of foliar metabolites in three Maize hybrids differing in water stress tolerance. PLoS ONE 2013, 8, e77145. [Google Scholar] [CrossRef]
- Obata, T.; Witt, S.; Lisec, J.; Palacios-Rojas, N.; Florez-Sarasa, I.; Yousfi, S.; Araus, J.L.; Cairns, J.E.; Fernie, A.R. Metabolite profiles of Maize leaves in drought, heat, and combined stress field trials reveal the relationship between metabolism and grain yield. Plant Physiol. 2015, 169, 2665–2683. [Google Scholar] [CrossRef]
- Warren, C.R.; Aranda, I.; Cano, F.J. Responses to water stress of gas exchange and metabolites in Eucalyptus and Acacia spp. Plant Cell Environ. 2011, 34, 1609–1629. [Google Scholar] [CrossRef]
- Krasensky, J.; Jonak, C. Drought, salt, and temperature stress-induced metabolic rearrangements and regulatory networks. J. Exp. Bot. 2012, 63, 1593–1608. [Google Scholar] [CrossRef] [Green Version]
- Warren, C.R.; Aranda, I.; Cano, F.J. Metabolomics demonstrates divergent responses of two Eucalyptus species to water stress. Metabolomics 2012, 8, 186–200. [Google Scholar] [CrossRef]
- Li, Y.; Xu, S.; Wang, Z.; He, L.; Xu, K.; Wang, G. Glucose triggers stomatal closure mediated by basal signaling through HXK1 and PYR/RCAR receptors in Arabidopsis. J. Exp. Bot. 2018, 69, 1471–1484. [Google Scholar] [CrossRef]
- Kelly, G.; Moshelion, M.; David-Schwartx, R.; Halperin, O.; Wallach, R.; Attia, Z.; Belausov, E.; Granot, D. Hexokinase mediates stomatal closure. Plant J. 2013, 75, 977–988. [Google Scholar] [CrossRef] [Green Version]
- Lu, Z.; Percy, R.G.; Qualset, C.O.; Zeiger, E. Stomatal conductance predicts yields in irrigated Pima cotton and bread wheat grown at high temperatures. J. Exp. Bot. 1998, 49, 453–460. [Google Scholar] [CrossRef]
- Radin, J.W.; Lu, Z.; Percy, R.G.; Zeiger, E. Genetic variability for stomatal conductance in Pima cotton and its relation to improvements of heat adaptation. Proc. Natl. Acad. Sci. USA 1994, 91, 7217–7221. [Google Scholar] [CrossRef] [PubMed]
- Grennan, A.K. The role of trehalose biosynthesis in plants. Plant Physiol. 2007, 144, 3–5. [Google Scholar] [CrossRef] [PubMed]
- Misra, B.B.; Acharya, B.R.; Granot, D.; Assmann, S.M.; Chen, S. The guard cell metabolome: functions in stomatal movement and global food security. Front Plant Sci. 2015, 6, 334. [Google Scholar] [CrossRef] [PubMed]
- Smirnoff, N.; Cumbes, Q.J. Hydroxyl radial scavenging activity of compatible solutes. Phytochemistry 1989, 28, 1057–1060. [Google Scholar] [CrossRef]
- Shen, B.; Jensen, R.G.; Bohnert, H.J. Increased resistance to oxidative stress in transgenic plants by targeting mannitol biosynthesis to chloroplasts. Plant Physiol. 1997, 113, 1177–1183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Widodo; Patterson, J.H.; Newbigin, E.; Tester, M.; Bacic, A.; Roessner, U. Metabolic responses to salt stress of barley (Hordeum vulgare L.) cultivars, Sahara and Clipper, which differ in salinity tolerance. J. Exp. Bot. 2009, 60, 4089–4103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Groppa, M.D.; Benavides, M.P. Polyamines and abiotic stress: Recent advances. Amino Acids 2008, 34, 35–45. [Google Scholar] [CrossRef]
- Alcazar, R.; Cuevas, J.C.; Planas, J.; Zarza, X.; Bortolotti, C.; Carrasco, P.; Salinas, J.; Tiburcio, A.F.; Altabella, T. Integration of polyamines in the cold acclimation response. Plant Sci. 2011, 180, 31–38. [Google Scholar] [CrossRef] [Green Version]
- Hussain, S.S.; Ali, M.; Ahmad, M.; Siddique, K.H. Polyamines: Natural and engineered abiotic and biotic stress tolerance in plants. Biotechnol. Adv. 2011, 29, 300–311. [Google Scholar] [CrossRef]
- Morgan, J.M. Osmoregulation and water stress in higher plants. Annu. Rev. Plant Physiol. 1984, 35, 299–319. [Google Scholar] [CrossRef]
- Zhang, J.; Nguyen, H.T.; Blum, A. Genetic analysis of osmotic adjustment in crop plants. J. Exp. Bot. 1999, 50, 291–302. [Google Scholar] [CrossRef] [Green Version]
- Serraj, R.; Sinclair, T.R. Osmolyte accumulation: Can it really help increase crop yield under drought conditions? Plant Cell Environ. 2002, 25, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Tuberosa, R. Phenotyping for drought tolerance of crops in the genomics era. Front. Physiol. 2012, 3, 347. [Google Scholar] [CrossRef]
- Blum, A. Osmotic adjustment is a prime drought stress adaptive engine in Support of plant production. Plant Cell Environ. 2017, 40, 4–10. [Google Scholar] [CrossRef]
- Vories, E.; Rhine, M.; Straatmann, Z. Investigating irrigation scheduling for rice using variable rate irrigation. Agric. Water Manag. 2017, 179, 314–323. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | Year | Yield under Water Saturated Condition (IRRI_1) | Yield Response (Slope of yield/% VWC) | Yield Performance under Water Stress | ||||
|---|---|---|---|---|---|---|---|---|
| Mean | SE | Estimate | SE | Prob. > F | R2 | |||
| TQNG | 2014 | 38.77 | 4.97 | 2.24 | 0.45 | ** | 0.72 | High response |
| 2015 | 55.80 | 8.05 | 2.37 | 0.82 | * | 0.55 | ||
| 2016 | 47.00 | 2.78 | 1.95 | 0.64 | ** | 0.54 | ||
| PI77 | 2014 | 31.60 | 6.13 | 1.59 | 0.46 | ** | 0.55 | |
| 2015 | 37.30 | 8.99 | 1.00 | 0.52 | ns | 0.35 | ||
| 2016 | 47.00 | 3.51 | 1.57 | 0.45 | ** | 0.56 | ||
| LGRU | 2014 | 25.00 | 1.67 | 1.02 | 0.25 | ** | 0.62 | Intermediate response |
| 2015 | 13.60 | 0.40 | 0.09 | 0.32 | ns | 0.02 | ||
| 2016 | 26.50 | 1.04 | 1.33 | 0.33 | ** | 0.34 | ||
| LMNT | 2014 | 16.93 | 0.73 | 0.80 | 0.27 | ** | 0.47 | |
| 2015 | 15.43 | 3.91 | 0.10 | 0.58 | ns | 0.01 | ||
| 2016 | 17.33 | 2.77 | 0.90 | 0.29 | ** | 0.62 | ||
| FRCS | 2014 | 38.03 | 1.68 | 0.89 | 0.72 | ns | 0.13 | Non or low response |
| 2015 | 31.13 | 5.40 | 0.41 | 0.58 | ns | 0.08 | ||
| 2016 | 37.67 | 6.89 | 0.33 | 0.49 | ns | 0.04 | ||
| KBNT | 2014 | 16.03 | 6.53 | 0.56 | 0.55 | ns | 0.09 | |
| 2015 | 13.10 | 3.41 | 0.49 | 0.20 | ns | 0.50 | ||
| 2016 | 14.17 | 5.13 | −0.10 | 0.32 | ns | 0.01 | ||
| SABR | 2014 | 30.63 | 3.54 | 0.70 | 0.38 | ns | 0.25 | |
| 2015 | 19.00 | 3.06 | 0.09 | 0.24 | ns | 0.02 | ||
| 2016 | 24.50 | 0.76 | 0.21 | 0.22 | ns | 0.08 | ||
| Cultivar | Based on 2014-2016 Results | 2016 Results | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| R | Mean Yield (g) | % Yield Loss from IRRI 1 | % Yield Loss from IRRI 1 | |||||||||
| IRRI1 | IRRI2 | IRRI3 | IRRI4 | Overall | IRRI2 | IRRI3 | IRRI4 | IRRI2 | IRRI3 | IRRI4 | ||
| TQNG | H | 47.19 | 51.51 | 31.55 | 17.56 | 36.95 | −9.16 | 38.75 | 62.80 | −19.15 | 21.99 | 39.36 |
| PI77 | 38.63 | 26.09 | 21.62 | 15.08 | 25.35 | 32.47 | 17.14 | 60.97 | 22.70 | 43.97 | 56.03 | |
| LGRU | I | 21.70 | 19.41 | 16.20 | 10.09 | 16.85 | 10.55 | 16.54 | 53.48 | 28.93 | 49.06 | 64.78 |
| LMNT | 16.57 | 14.02 | 13.40 | 7.40 | 12.85 | 15.36 | 4.44 | 55.33 | −10.58 | 2.88 | 76.73 | |
| FRCS | L | 35.61 | 33.48 | 36.50 | 25.58 | 32.79 | 5.99 | −9.03 | 28.17 | 7.52 | −22.12 | 22.12 |
| KBNT | 14.43 | 11.95 | 18.90 | 8.23 | 13.38 | 17.21 | −58.16 | 42.96 | −14.12 | −36.47 | −5.88 | |
| SABR | 24.71 | 26.66 | 23.27 | 20.18 | 23.70 | −7.87 | 12.71 | 18.35 | 0.68 | 4.76 | 13.61 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barnaby, J.Y.; Rohila, J.S.; Henry, C.G.; Sicher, R.C.; Reddy, V.R.; McClung, A.M. Physiological and Metabolic Responses of Rice to Reduced Soil Moisture: Relationship of Water Stress Tolerance and Grain Production. Int. J. Mol. Sci. 2019, 20, 1846. https://doi.org/10.3390/ijms20081846
Barnaby JY, Rohila JS, Henry CG, Sicher RC, Reddy VR, McClung AM. Physiological and Metabolic Responses of Rice to Reduced Soil Moisture: Relationship of Water Stress Tolerance and Grain Production. International Journal of Molecular Sciences. 2019; 20(8):1846. https://doi.org/10.3390/ijms20081846
Chicago/Turabian StyleBarnaby, Jinyoung Y., Jai S. Rohila, Chris G. Henry, Richard C. Sicher, Vagimalla R. Reddy, and Anna M. McClung. 2019. "Physiological and Metabolic Responses of Rice to Reduced Soil Moisture: Relationship of Water Stress Tolerance and Grain Production" International Journal of Molecular Sciences 20, no. 8: 1846. https://doi.org/10.3390/ijms20081846
APA StyleBarnaby, J. Y., Rohila, J. S., Henry, C. G., Sicher, R. C., Reddy, V. R., & McClung, A. M. (2019). Physiological and Metabolic Responses of Rice to Reduced Soil Moisture: Relationship of Water Stress Tolerance and Grain Production. International Journal of Molecular Sciences, 20(8), 1846. https://doi.org/10.3390/ijms20081846