Copper Utilization, Regulation, and Acquisition by Aspergillus fumigatus



Abstract
:1. Introduction
2. Copper Metabolic Processes
2.1. Enzymes Using Copper as A Cofactor
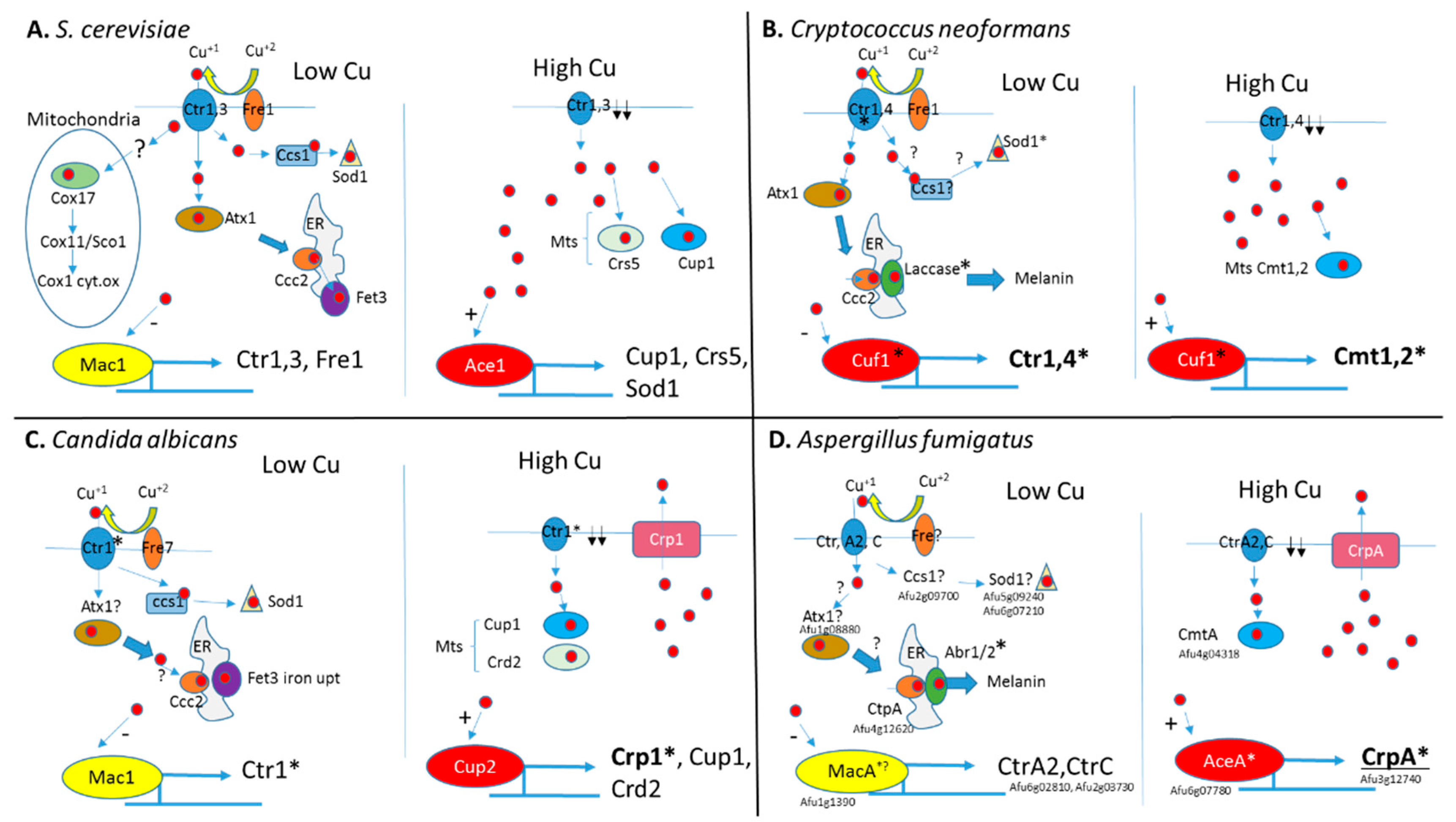
2.2. Copper Transporters and Copper-dependent Transcription Factors
3. Copper Toxicity
4. Infection Biology and Copper
4.1. Fungal Response to Copper Limitation
4.2. Fungal Response to Copper Excess
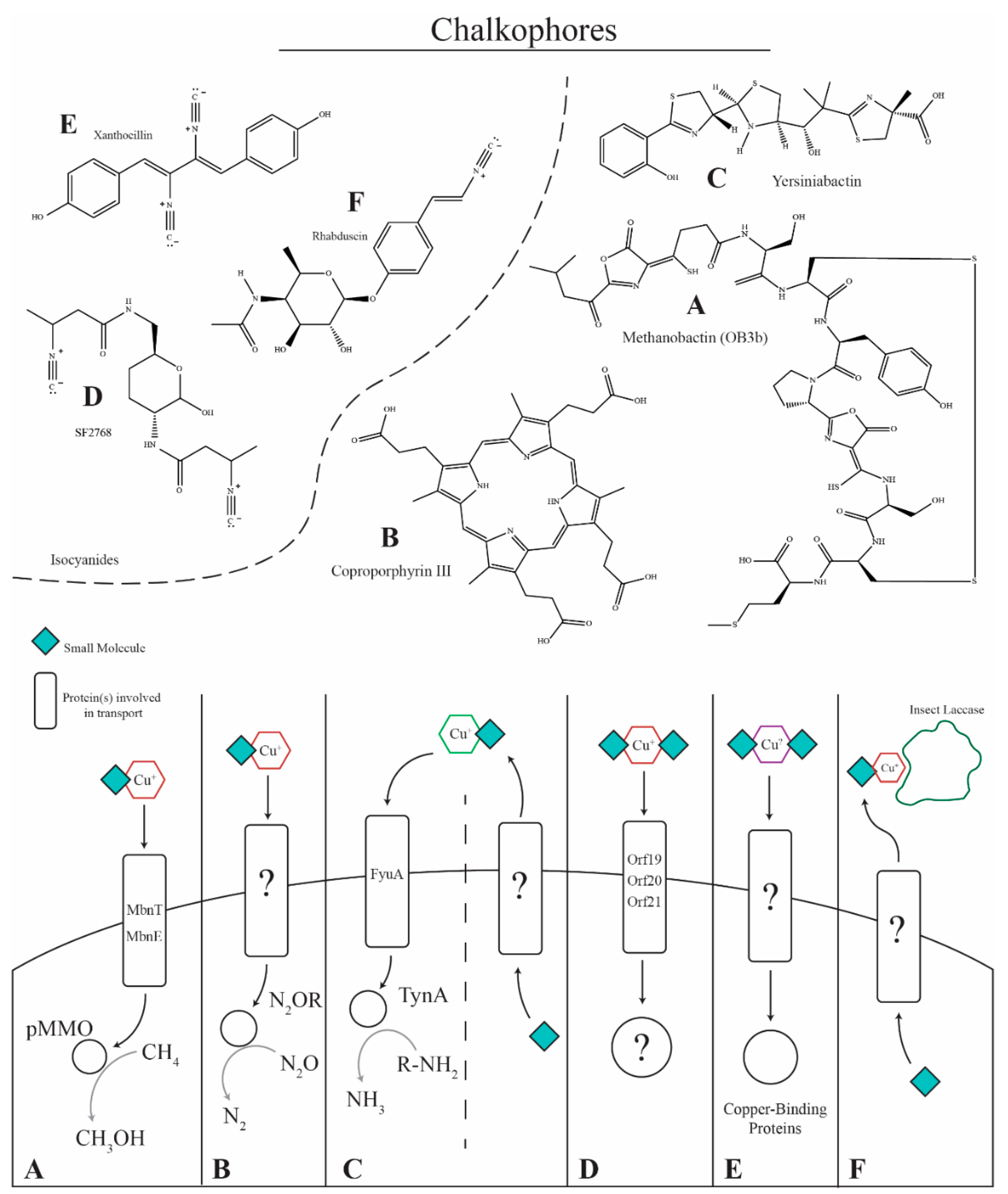
5. Copper-Binding Secondary Metabolites
6. Future Directions
Funding
Acknowledgments
Conflicts of Interest
References
- Holliday, G.L.; Thornton, J.M. Metal Ions in Biological Catalysis: From Enzyme Databases to General Principles. J. Biol. Inorg. Chem. 2008, 13, 1205–1218. [Google Scholar] [CrossRef]
- Waldron, K.J.; Robinson, N.J. How Do Bacterial Cells Ensure That Metalloproteins Get the Correct Metal? Nat. Rev. Microbiol. 2009, 7, 25–35. [Google Scholar] [CrossRef]
- Ganz, T. Iron and Infection. Int. J. Hematol. 2018, 107, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Cassat, J.E.; Skaar, E.P. Review Iron in Infection and Immunity. Cell Host Microbe 2013, 13, 509–519. [Google Scholar] [CrossRef] [PubMed]
- Irving, H.; Williams, R.J.P. The Stability of Transition-Metal Complexes. J. Chem. Soc. 1953, 0, 3192–3210. [Google Scholar] [CrossRef]
- Macomber, L.; Imlay, J.A. The Iron-Sulfur Clusters of Dehydratases Are Primary Intracellular Targets of Copper Toxicity. Proc. Natl. Acad. Sci. USA 2009, 106, 8344–8349. [Google Scholar] [CrossRef] [PubMed]
- Heinekamp, T.; Schmidt, H.; Lapp, K.; Pähtz, V. Interference of Aspergillus Fumigatus with the Immune Response. Semin. Immunopathol. 2015, 37, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Wolschendorf, F.; Ackart, D.; Shrestha, T.B.; Hascall-dove, L.; Nolan, S.; Lamichhane, G. Copper Resistance Is Essential for Virulence of Mycobacterium Tuberculosis. Proc. Natl. Acad. Sci. USA 2010, 1–6. [Google Scholar] [CrossRef]
- Naaraayan, A.; Kavian, R.; Lederman, J.; Basak, P.; Jesmajian, S. Invasive Pulmonary Aspergillosis—Case Report and Review of Literature. J. Community Hosp. Intern. Med. Perspect. 2015, 5, 3–6. [Google Scholar] [CrossRef]
- Pena, M.M.O.; Lee, J.; Thiele, D.J. Critical Review a Delicate Balance: Homeostatic Control of Copper. Am. Soc. Nutr. Sci. 1999, 1251–1260. [Google Scholar]
- Smith, A.D.; Logeman, B.L.; Thiele, D.J. Copper Acquisition and Utilization in Fungi. Annu. Rev. Microbiol. 2017, 71, 597–623. [Google Scholar] [CrossRef] [PubMed]
- Gerwien, F.; Skrahina, V.; Kasper, L.; Hube, B.; Brunke, S. Metals in Fungal Virulence. FEMS Microbiol. Rev. 2018, 42, 1–21. [Google Scholar] [CrossRef]
- Little, A.G.; Lau, G.; Mathers, K.E.; Leary, S.C.; Moyes, C.D. Comparative Biochemistry and Physiology, Part B Comparative Biochemistry of Cytochrome c Oxidase in Animals. Comp. Biochem. Physiol. Part B 2018, 224, 170–184. [Google Scholar] [CrossRef]
- Grahl, N.; Dinamarco, T.M.; Willger, S.D.; Goldman, G.H.; Cramer, R.A. Aspergillus Fumigatus Mitochondrial Electron Transport Chain Mediates Oxidative Stress Homeostasis, Hypoxia Responses and Fungal Pathogenesis. Mol. Microbiol. 2012, 84, 383–399. [Google Scholar] [CrossRef]
- Strogolova, V.; Hoang, N.H.; Hosler, J.; Stuart, R.A. The Yeast Mitochondrial Proteins Rcf1 and Rcf2 Support the Enzymology of the Cytochrome c Oxidase Complex and Generation of the Proton Motive Force. J. Biol. Chem. 2019, 294, 4867–4877. [Google Scholar] [CrossRef]
- Beatriz Flecha, D.B. Metabolic Soruces of Hydrogen Peroxide in Aerobically Growing E. coli. J. Biol. Chem. 1995, 270, 13681. [Google Scholar] [CrossRef]
- Gonzalez-flecha, B.; Boveris, A. Mitochondrial Sites of Hydrogen Peroxide Production in Reperfused Rat Kidney Cortex. Biochim. Biophys. Acta 1995, 1243, 361–366. [Google Scholar] [CrossRef]
- Jitsukawa, K.; Harata, M.; Arii, H.; Sakurai, H. SOD Activities of the Copper Complexes with Tripodal Polypyridylamine Ligands Having a Hydrogen Bonding Site. Inorg. Chim. Acta 2001, 324, 108–116. [Google Scholar] [CrossRef]
- Lambou, K.; Lamarre, C.; Beau, R.; Dufour, N.; Latge, J. Functional Analysis of the Superoxide Dismutase Family in Aspergillus Fumigatus. Mol. Microbiol. 2010, 75, 910–923. [Google Scholar] [CrossRef]
- Long, A.; Song, B.; Fridey, K.; Silva, A. Detection and Diversity of Copper Containing Nitrite Reductase Genes (NirK) in Prokaryotic and Fungal Communities of Agricultural Soils. FEMS Microbiol. Ecol. 2015, 91, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Cánovas, D.; Marcos, J.F.; Marcos, A.T.; Strauss, J. Nitric Oxide in Fungi: Is There NO Light at the End of the Tunnel? Curr. Genet. 2016, 62, 513–518. [Google Scholar] [CrossRef]
- Li, Y.; Hodak, M.; Bernholc, J. Enzymatic Mechanism of Copper-Containing Nitrite Reductase. Biochemistry 2015, 54, 1233–1242. [Google Scholar] [CrossRef]
- Schrettl, M.; Bignell, E.; Kragl, C.; Joechl, C.; Rogers, T.; Arst, H.N.; Haynes, K.; Haas, H. Siderophore Biosynthesis But Not Reductive Iron Assimilation Is Essential for Aspergillus Fumigatus Virulence. J. Exp. Med. 2004, 200, 1213–1219. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, S.; Torres, G.; Lin, X. Laccases Involved in 1,8-Dihydroxynaphthalene Melanin Biosynthesis in Aspergillus Fumigatus Are Regulated by Developmental Factors and Copper Homeostasis. Eukaryot. Cell 2013, 12, 1641–1652. [Google Scholar] [CrossRef] [PubMed]
- Brenelli, L.; Squina, F.M.; Felby, C.; Cannella, D. Biotechnology for Biofuels Laccase-Derived Lignin Compounds Boost Cellulose Oxidative Enzymes AA9. Biotechnol. Biofuels 2018, 1–12. [Google Scholar] [CrossRef]
- Wiemann, P.; Perevitsky, A.; Lim, F.Y.; Shadkchan, Y.; Knox, B.P.; Landero Figueora, J.A.; Choera, T.; Niu, M.; Steinberger, A.J.; Wüthrich, M.; et al. Aspergillus Fumigatus Copper Export Machinery and Reactive Oxygen Intermediate Defense Counter Host Copper-Mediated Oxidative Antimicrobial Offense. Cell Rep. 2017, 19, 1008–1021. [Google Scholar] [CrossRef]
- Cai, Z.; Du, W.; Zeng, Q.; Long, N.; Dai, C.; Lu, L. Cu-Sensing Transcription Factor Mac1 Coordinates with the Ctr Transporter Family to Regulate Cu Acquisition and Virulence in Aspergillus Fumigatus. Fungal Genet. Biol. 2017, 107, 31–43. [Google Scholar] [CrossRef]
- Park, Y.; Lian, H.; Chang, M.; Kang, C.; Yun, C. Identification of High-Affinity Copper Transporters in Aspergillus Fumigatus. Fungal Genet. Biol. 2014, 73, 29–38. [Google Scholar] [CrossRef]
- Lin, S.J.; O’Halloran, T.V.; Pufahl, R.A.; Culotta, V.C.; Dancis, A. A Role for the Saccharomyces Cerevisiae ATX1 Gene in Copper Trafficking and Iron Transport. J. Biol. Chem. 1997, 272, 9215–9220. [Google Scholar] [CrossRef]
- Brown, N.M.; Torres, A.S.; Doan, P.E.; O’Halloran, T.V. Oxygen and the Copper Chaperone CCS Regulate Posttranslational Activation of Cu,Zn Superoxide Dismutase. Proc. Natl. Acad. Sci. USA 2004, 101, 5518–5523. [Google Scholar] [CrossRef]
- Bakti, F.; Sasse, C.; Heinekamp, T.; Pócsi, I.; Braus, G.H. Heavy Metal-Induced Expression of PcaA Provides Cadmium Tolerance to Aspergillus Fumigatus and Supports Its Virulence in the Galleria Mellonella Model. Front. Microbiol. 2018, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Glerum, M.; Shtanko, A.; Tzagoloff, A. Characterization of COX1 7, a Yeast Gene Involved in Copper Metabolism and Assembly of Cytochrome Oxidase. J. Biol. Chem. 1996, 24, 14504–14509. [Google Scholar] [CrossRef]
- Jomova, K.; Lawson, M.; Drostinova, L.; Lauro, P.; Poprac, P.; Brezova, V.; Michalik, M.; Lukes, V.; Valko, M. Protective Role of Quercetin against Copper (II)-Induced Oxidative Stress: A Spectroscopic, Theoretical and DNA Damage Study. Food Chem. Toxicol. 2017, 110, 340–350. [Google Scholar] [CrossRef]
- Prousek, J. Fenton Chemistry in Biology and Medicine. Pure Appl. Chem. 2007, 79, 2325–2338. [Google Scholar] [CrossRef]
- Bruska, M.K.; Stiebritz, M.T.; Reiher, M. Binding of Reactive Oxygen Species at Fe-S Cubane Clusters. Comput. Chem. 2015, 21, 19081–19089. [Google Scholar] [CrossRef]
- Jang, S.; Imlay, J.A. Micromolar Intracellular Hydrogen Peroxide Disrupts Metabolism by Damaging Iron-Sulfur Enzymes. J. Biol. Chem. 2007, 282, 929–937. [Google Scholar] [CrossRef]
- Murakami, K.; Yoshino, M. Inactivation of Aconitase in yeast exposed to oxidative stress. Biochem. Mol. Biol. 1997, 41, 481–486. [Google Scholar] [CrossRef]
- Imlay, J.A. The Mismetallation of Enzymes during Oxidative Stress. J. Biol. Chem. 2014, 289, 28121–28128. [Google Scholar] [CrossRef]
- Vallieres, C.; Holland, S.L.; Avery, S.V. Mitochondrial Ferredoxin Determines Vulnerability of Cells to Copper Excess Article Mitochondrial Ferredoxin Determines Vulnerability of Cells to Copper Excess. Cell Chem. Biol. 2017, 24, 1228–1237. [Google Scholar] [CrossRef]
- De Freitas, J.M.; Valentine, J.S.; Gralla, E.B.; Liba, A.; Meneghini, R. Yeast Lacking Cu-Zn Superoxide Dismutase Show Altered Iron Homeostasis. J. Biol. Chem. 2002, 275, 11645–11649. [Google Scholar] [CrossRef]
- White, C.; Lee, J.; Kambe, T.; Fritsche, K.; Petris, M.J. A Role for the ATP7A Copper-Transporting ATPase in Macrophage Bactericidal Activity. J. Biol. Chem. 2009, 284, 33949–33956. [Google Scholar] [CrossRef] [PubMed]
- Ding, C.; Festa, R.A.; Sun, T.S.; Wang, Z.Y. Iron and Copper as Virulence Modulators in Human Fungal Pathogens. Mol. Microbiol. 2014, 93, 10–23. [Google Scholar] [CrossRef] [PubMed]
- Li, C.X.; Gleason, J.E.; Zhang, S.X.; Bruno, V.M.; Cormack, B.P.; Culotta, V.C. Candida Albicans Adapts to Host Copper during Infection by Swapping Metal Cofactors for Superoxide Dismutase. Proc. Natl. Acad. Sci. USA 2015, 112, E5336–E5342. [Google Scholar] [CrossRef]
- Mackie, J.; Szabo, E.K.; Urgast, D.S.; Ballou, E.R.; Childers, D.S.; MacCallum, D.M.; Feldmann, J.; Brown, A.J.P. Host-Imposed Copper Poisoning Impacts Fungal Micronutrient Acquisition during Systemic Candida Albicans Infections. PLoS ONE 2016, 11, 1–18. [Google Scholar] [CrossRef]
- Sun, T.-S.; Ju, X.; Gao, H.-L.; Wang, T.; Thiele, D.J.; Li, J.-Y.; Wang, Z.-Y.; Ding, C. Reciprocal Functions of Cryptococcus Neoformans Copper Homeostasis Machinery during Pulmonary Infection and Meningoencephalitis. Nat. Commun. 2014, 5, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Georgatsou, E.; Mavrogiannis, L.A.; Fragiadakis, G.S.; Alexandraki, D. The Yeast Fre1p/Fre2p Cupric Reductases Facilitate Copper Uptake and Are Regulated by the Copper-Modulated Mac1p Activator. J. Biol. Chem. 1997, 272, 13786–13792. [Google Scholar] [CrossRef]
- Labbé, S.; Zhu, Z.; Thiele, D.J. Copper-Specific Transcriptional Repression of Yeast Genes Encoding Critical Components in the Copper Transport Pathway. J. Biol. Chem. 1997, 272, 15951–15958. [Google Scholar] [CrossRef] [PubMed]
- Jensen, L.T.; Winge, D.R. Identification of a Copper-Induced Intramolecular Interaction in the Transcription Factor Mac1 from Saccharomyces Cerevisiae. EMBO J. 1998, 17, 5400–5408. [Google Scholar] [CrossRef] [PubMed]
- Culotta, V.C.; Howard, W.R.; Liu, X.F. CRSS Encodes a Metallothionein-like Protein. J. Biol. Chem. 1994, 2, 25295–25302. [Google Scholar]
- Corazza, A.; Harvey, I.; Sadler, P.J. 1H,13C-NMR and X-Ray Absorption Studies of Copper(I) Glutathione Complexes. Eur. J. Biochem. 1996, 236, 697–705. [Google Scholar] [CrossRef]
- Jensen, L.T.; Howard, W.R.; Strain, J.J.; Winge, D.R.; Valeria, C.C. Enhanced Effectiveness of Copper Ion Buffering by CUP1 Metallothionein Compared with CRS5 Metallothionein in Saccharomyces Cerevisiae. J. Biol. Chem. 1996, 271, 18514–18519. [Google Scholar] [CrossRef]
- Punter, F.A.; Glerum, D.M. Mutagenesis Reveals a Specific Role for Cox17p in Copper Transport to Cytochrome Oxidase. J. Biol. Chem. 2003, 278, 30875–30880. [Google Scholar] [CrossRef] [Green Version]
- Boyd, S.D.; Liu, L.; Bulla, L.; Winkler, D.D. Quantifying the Interaction between Copper-Zinc Superoxide Dismutase (Sod1) and Its Copper Chaperone (Ccs1). J. Proteomics Bioinform. 2018, 11, 99–103. [Google Scholar] [CrossRef]
- Woodacre, A.; Mason, R.P.; Jeeves, R.E.; Cashmore, A.M. Copper-Dependent Transcriptional Regulation by Candida Albicans Mac1p. Microbiology 2008, 154, 1502–1512. [Google Scholar] [CrossRef] [PubMed]
- Waterman, S.R.; Hacham, M.; Zhu, X.; Park, Y.-D.; Hu, G.; Shin, S.; Panepinto, J.; Valyi-Nagy, T.; Beam, C.; Husain, S.; et al. Role of a CUF1/CTR4 Copper Regulatory Axis in the Virulence of Cryptococcus Neoformans. J. Clin. Investig. 2007, 117, 794–802. [Google Scholar] [CrossRef] [PubMed]
- Ding, C.; Festa, R.A.; Chen, Y.L.; Espart, A.; Palacios, Ò.; Espín, J.; Capdevila, M.; Atrian, S.; Heitman, J.; Thiele, D.J. Cryptococcus Neoformans Copper Detoxification Machinery Is Critical for Fungal Virulence. Cell Host Microbe 2013, 13, 265–276. [Google Scholar] [CrossRef]
- Kusuya, Y.; Hagiwara, D.; Sakai, K.; Yaguchi, T.; Gonoi, T.; Takahashi, H. Transcription Factor Afmac1 Controls Copper Import Machinery in Aspergillus Fumigatus. Curr. Genet. 2017, 63, 777–789. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.; Du, W.; Zhang, Z.; Guan, L.; Zeng, Q.; Chai, Y.; Dai, C.; Lu, L. The Aspergillus Fumigatus Transcription Factor AceA Is Involved Not Only in Cu but Also in Zn Detoxification through Regulating Transporters CrpA and ZrcA. Cell. Microbiol. 2018, 20, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.-S.; Kim, T.-H.; Yun, C.-W. Functional Characterization of the Copper Transcription Factor AfMac1 from Aspergillus Fumigatus. Biochem. J. 2017, 474, 2365–2378. [Google Scholar] [CrossRef] [PubMed]
- Hover, B.M.; Kim, S.H.; Katz, M.; Charlop-Powers, Z.; Owen, J.G.; Ternei, M.A.; Maniko, J.; Estrela, A.B.; Molina, H.; Park, S.; et al. Culture-Independent Discovery of the Malacidins as Calcium-Dependent Antibiotics with Activity against Multidrug-Resistant Gram-Positive Pathogens. Nat. Microbiol. 2018, 3, 415–422. [Google Scholar] [CrossRef]
- Park, Y.S.; Kang, S.; Seo, H.; Yun, C.W. A Copper Transcription Factor, AfMac1, Regulates Both Iron and Copper Homeostasis in the Opportunistic Fungal Pathogen Aspergillus Fumigatus. Biochem. J. 2018, 475, 2831–2845. [Google Scholar] [CrossRef]
- Keller, N.P. Heterogeneity Confounds Establishment of “a” Model Microbial Strain. MBio 2017, 8, e00135-17. [Google Scholar] [CrossRef]
- Fürst, P.; Hu, S.; Hackett, R.; Hamer, D. Copper Activates Metallothionein Gene Transcription by Altering the Conformation of a Specific DNA Binding Protein. Cell 1988, 55, 705–717. [Google Scholar] [CrossRef]
- Culotta, C.V.; Hsu, T.; Hu, S.; Furst, P. Copper and the ACEl Regulatory Protein Reversibly Induce Yeast Metallothionein Gene Transcription in a Mouse Extract (Metal Ions/Transcription Factor/DNA-Binding Protein). Proc. Natl. Acad. Sci. USA 1989, 86, 8377–8381. [Google Scholar] [CrossRef]
- Gralla, E.B.; Thiele, D.J.; Silar, P.; Valentine, J.S. ACE1, a Copper-Dependent Transcription Factor, Activates Expression of the Yeast Copper, Zinc Superoxide Dismutase Gene. Proc. Natl. Acad. Sci. USA 2006, 88, 8558–8562. [Google Scholar] [CrossRef]
- Thiele, D.J. ACE1 Regulates Expression of the Saccharomyces Cerevisiae Metallothionein Gene. Mol. Cell. Biol. 1988, 8, 2745–2752. [Google Scholar] [CrossRef]
- Culotta, V.; Joh, H.; Lin, S.; Slekar, K.; Strain, J. A Physiological Role for Saccharomyces Cerevisiae Copper/Zinc Superoxide Dismutase in Copper Buffering. J. Biol. Chem. 1995, 270, 29991–29997. [Google Scholar]
- Weissman, Z.; Berdicevsky, I.; Cavari, B.-Z.; Kornitzer, D. The High Copper Tolerance of Candida Albicans Is Mediated by a P-Type ATPase. Proc. Natl. Acad. Sci. USA 2000, 97, 3520–3525. [Google Scholar] [CrossRef]
- Antsotegi-Uskola, M.; Markina-Iñarrairaegui, A.; Ugalde, U. Copper Resistance in Aspergillus Nidulans Relies on the PI-Type ATPase CrpA, Regulated by the Transcription Factor AceA. Front. Microbiol. 2017, 8, 1–15. [Google Scholar] [CrossRef]
- Yang, K.; Shadkchan, Y.; Tannous, J.; Landero Figueroa, J.A.; Wiemann, P.; Osherov, N.; Wang, S.; Keller, N.P. Contribution of ATPase Copper Transporters in Animal but Not Plant Virulence of the Crossover Pathogen Aspergillus Flavus. Virulence 2018, 9, 1273–1286. [Google Scholar] [CrossRef]
- Wiemann, P.; Lechner, B.E.; Baccile, J.A.; Velk, T.A.; Yin, W.B.; Bok, J.W.; Pakala, S.; Losada, L.; Nierman, W.C.; Schroeder, F.C.; et al. Perturbations in Small Molecule Synthesis Uncovers an Iron-Responsive Secondary Metabolite Network in Aspergillus Fumigatus. Front. Microbiol. 2014, 5, 1–16. [Google Scholar] [CrossRef]
- Haas, H. Fungal Siderophore Metabolism with a Focus on Aspergillus Fumigatus. Nat. Prod. Rep. 2014, 31, 1266–1276. [Google Scholar] [CrossRef]
- Bignell, E.; Cairns, T.C.; Throckmorton, K.; Nierman, W.C.; Keller, N.P. Secondary Metabolite Arsenal of an Opportunistic Pathogenic Fungus. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20160023. [Google Scholar] [CrossRef] [PubMed]
- Schrettl, M.; Bignell, E.; Kragl, C.; Sabiha, Y.; Loss, O.; Eisendle, M.; Wallner, A.; Arst, H.N.; Haynes, K.; Haas, H. Distinct Roles for Intra- and Extracellular Siderophores during Aspergillus Fumigatus Infection. PLoS Pathog. 2007, 3, 1195–1207. [Google Scholar] [CrossRef] [PubMed]
- Yin, W.B.; Baccile, J.A.; Bok, J.W.; Chen, Y.; Keller, N.P.; Schroeder, F.C. A Nonribosomal Peptide Synthetase-Derived Iron(III) Complex from the Pathogenic Fungus Aspergillus Fumigatus. J. Am. Chem. Soc. 2013, 135, 2064–2067. [Google Scholar] [CrossRef] [PubMed]
- Kenney, G.E.; Rosenzweig, A.C. Chalkophores. Annu. Rev. Biochem. 2018, 87, 645–676. [Google Scholar] [CrossRef] [PubMed]
- Bergaust, L.; Van Spanning, R.J.M.; Bakken, L.R. Expression of Nitrous Oxide Reductase in Paracoccus Denitrificans Is Regulated by Oxygen and Nitric Oxide through FnrP and NNR. Microbiology 2012, 158, 826–834. [Google Scholar] [CrossRef] [PubMed]
- Anttila, J.; Heinonen, P.; Nenonen, T.; Pino, A.; Iwaï, H.; Kauppi, E.; Soliymani, R.; Baumann, M.; Saksi, J.; Suni, N.; et al. Biochimica et Biophysica Acta Is Coproporphyrin III a Copper-Acquisition Compound in Paracoccus Denitrificans? BBA Bioenerg. 2011, 1807, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Chaturvedi, K.S.; Hung, C.S.; Giblin, D.E.; Urushidani, S.; Austin, A.M.; Dinauer, M.C.; Henderson, J.P. Cupric Yersiniabactin Is a Virulence-Associated Superoxide Dismutase Mimic. ACS Chem. Biol. 2014, 9, 551–561. [Google Scholar] [CrossRef]
- Nolan, E.M. A Noncanonical Role for Yersiniabactin in Bacterial Copper Acquisition. Biochemistry 2017, 56, 6073–6074. [Google Scholar] [CrossRef] [PubMed]
- Chaturvedi, K.S.; Hung, C.S.; Crowley, J.R.; Stapleton, A.E.; Henderson, J.P. The Siderophore Yersiniabactin Binds Copper to Protect Pathogens during Infection. Nat. Chem. Biol. 2012, 8, 731–736. [Google Scholar] [CrossRef] [PubMed]
- Koh, E.I.; Robinson, A.E.; Bandara, N.; Rogers, B.E.; Henderson, J.P. Copper Import in Escherichia Coli by the Yersiniabactin Metallophore System. Nat. Chem. Biol. 2017, 13, 1016–1021. [Google Scholar] [CrossRef]
- Wang, L.; Zhu, M.; Zhang, Q.; Zhang, X.; Yang, P.; Liu, Z.; Deng, Y.; Zhu, Y.; Huang, X.; Han, L.; et al. Diisonitrile Natural Product SF2768 Functions As a Chalkophore That Mediates Copper Acquisition in Streptomyces Thioluteus. ACS Chem. Biol. 2017, 12, 3067–3075. [Google Scholar] [CrossRef] [PubMed]
- Hahn, E. The Coordination Chemistry of Multidentate Isocyanide Ligands. Angew. Chem. Int. Ed. 1993, 33, 650–665. [Google Scholar] [CrossRef]
- Crawford, J.M.; Portmann, C.; Zhang, X.; Roeffaers, M.B.J.; Clardy, J. Small Molecule Perimeter Defense in Entomopathogenic Bacteria. Proc. Natl. Acad. Sci. USA 2012, 109, 10821–10826. [Google Scholar] [CrossRef] [PubMed]
- Harris, N.C.; Sato, M.; Herman, N.A.; Twigg, F.; Cai, W.; Liu, J.; Zhu, X.; Downey, J.; Khalaf, R.; Martin, J.; et al. Biosynthesis of Isonitrile Lipopeptides by Conserved Nonribosomal Peptide Synthetase Gene Clusters in Actinobacteria. Proc. Natl. Acad. Sci. USA 2017, 114, 7025–7030. [Google Scholar] [CrossRef] [PubMed]
- Lim, F.Y.; Won, T.H.; Raffa, N.; Baccile, J.A.; Wisecaver, J.; Rokas, A.; Schroeder, F.C.; Keller, N.P. Fungal Isocyanide Synthases and Xanthocillin Biosynthesis in Aspergillus Fumigatus. MBio 2018, 9, e00785-18. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Copper-Binding Proteins in Aspergillus fumigatus | ||||
|---|---|---|---|---|
| Designation (AFUA) | Name | Function | Reference | |
| Known function | 6g07780 | AceA | Copper-Toxicity Transcription Factor | [26] |
| 1g13190 | MacA | Copper-Deficiency Transcription Factor | [27] | |
| 2g01190 | CufA | Unknown Function Transcription Factor | [26] | |
| 6g02810 | CtrA2 | High-Affinity Copper Importer | [28] | |
| 2g03730 | CtrC | High-Affinity Copper Importer | [28] | |
| 3G13660 | CtrA1 | Copper Importer | [28] | |
| 3g08180 | Ctr2 | Low-Affinity Importer | [28] | |
| 3g12740 | CrpA | Copper P-Type ATPase Exporter | [26] | |
| 4g12620 | CtpA | Intracellular Copper ATPase | [24] | |
| 4g04318 | CmtA | Copper Metallothionein | [26] | |
| 5g09240 | Sod1 | Cytoplasmic Superoxide Dismutase | [19] | |
| 5g03790 | FetC | Ferrioxidase Involved in Iron Import | [23] | |
| 2g17540 | Abr1 | Laccase | [24] | |
| 2g17530 | Abr2 | Laccase | [24] | |
| 2g13110 | CycA | Cytochrome C oxidase | [14] | |
| Putative function | 3g14950 | NirK * | Copper-Binding Nitrite with Reductase Activity | [20] |
| 1g08880 | Atx1 * | Copper Chaperone | [29] | |
| 2g09700 | Ccs1 * | Sod1 Copper Chaperone | [30] | |
| 1g16130 | Pca1 * | Cd-Exporting ATPase | [31] | |
| 3g07690 | Cox17 * | Copper Chaperone to Cytochrome C Oxidase | [32] | |
| 5g01470 | Amine Oxidase | |||
| 3g14590 | Amine Oxidase | |||
| 3g00680 | Copper Amine Oxidase | |||
| 7g08470 | Copper Amine Oxidase | |||
| 5g07360 | Peroxisomal Amine Oxidase | |||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raffa, N.; Osherov, N.; Keller, N.P. Copper Utilization, Regulation, and Acquisition by Aspergillus fumigatus. Int. J. Mol. Sci. 2019, 20, 1980. https://doi.org/10.3390/ijms20081980
Raffa N, Osherov N, Keller NP. Copper Utilization, Regulation, and Acquisition by Aspergillus fumigatus. International Journal of Molecular Sciences. 2019; 20(8):1980. https://doi.org/10.3390/ijms20081980
Chicago/Turabian StyleRaffa, Nicholas, Nir Osherov, and Nancy P. Keller. 2019. "Copper Utilization, Regulation, and Acquisition by Aspergillus fumigatus" International Journal of Molecular Sciences 20, no. 8: 1980. https://doi.org/10.3390/ijms20081980
APA StyleRaffa, N., Osherov, N., & Keller, N. P. (2019). Copper Utilization, Regulation, and Acquisition by Aspergillus fumigatus. International Journal of Molecular Sciences, 20(8), 1980. https://doi.org/10.3390/ijms20081980