AHK3-Mediated Cytokinin Signaling Is Required for the Delayed Leaf Senescence Induced by SSPP


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
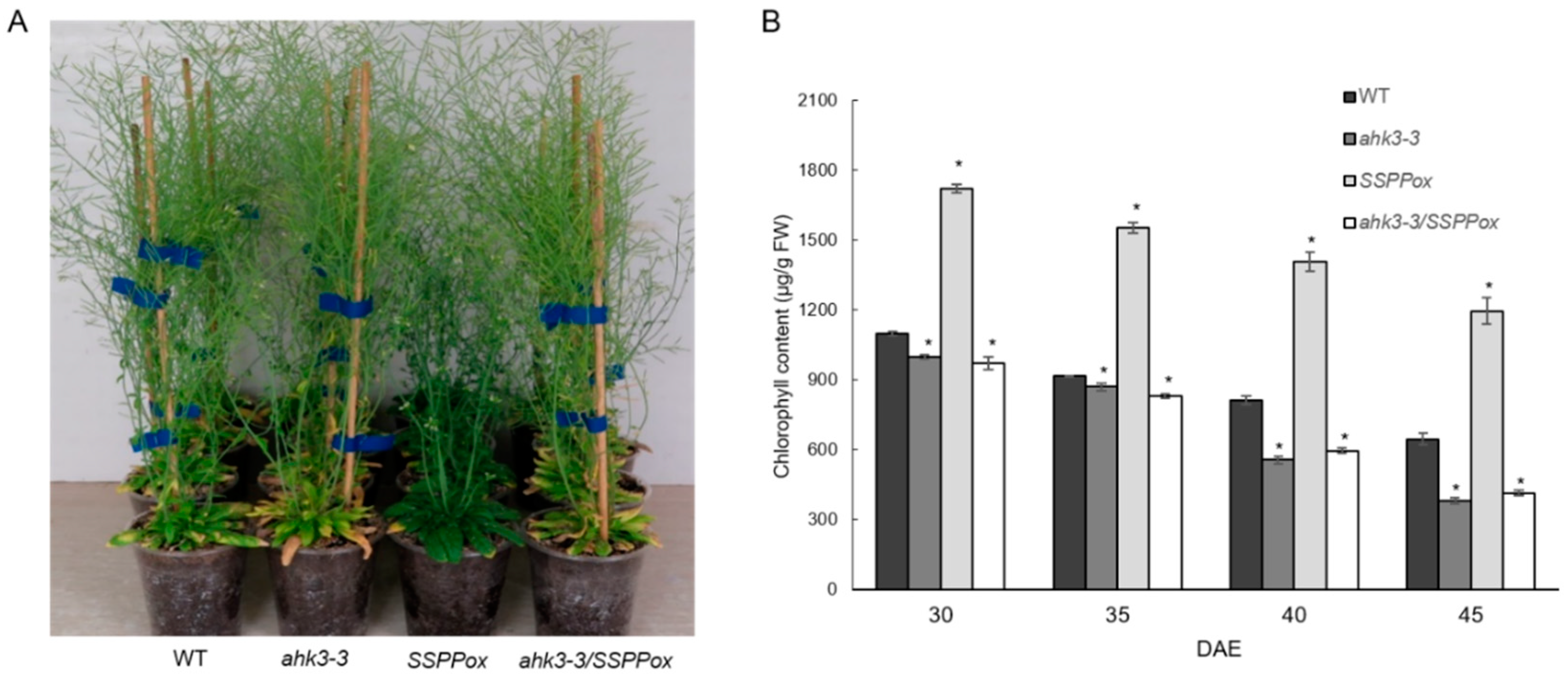
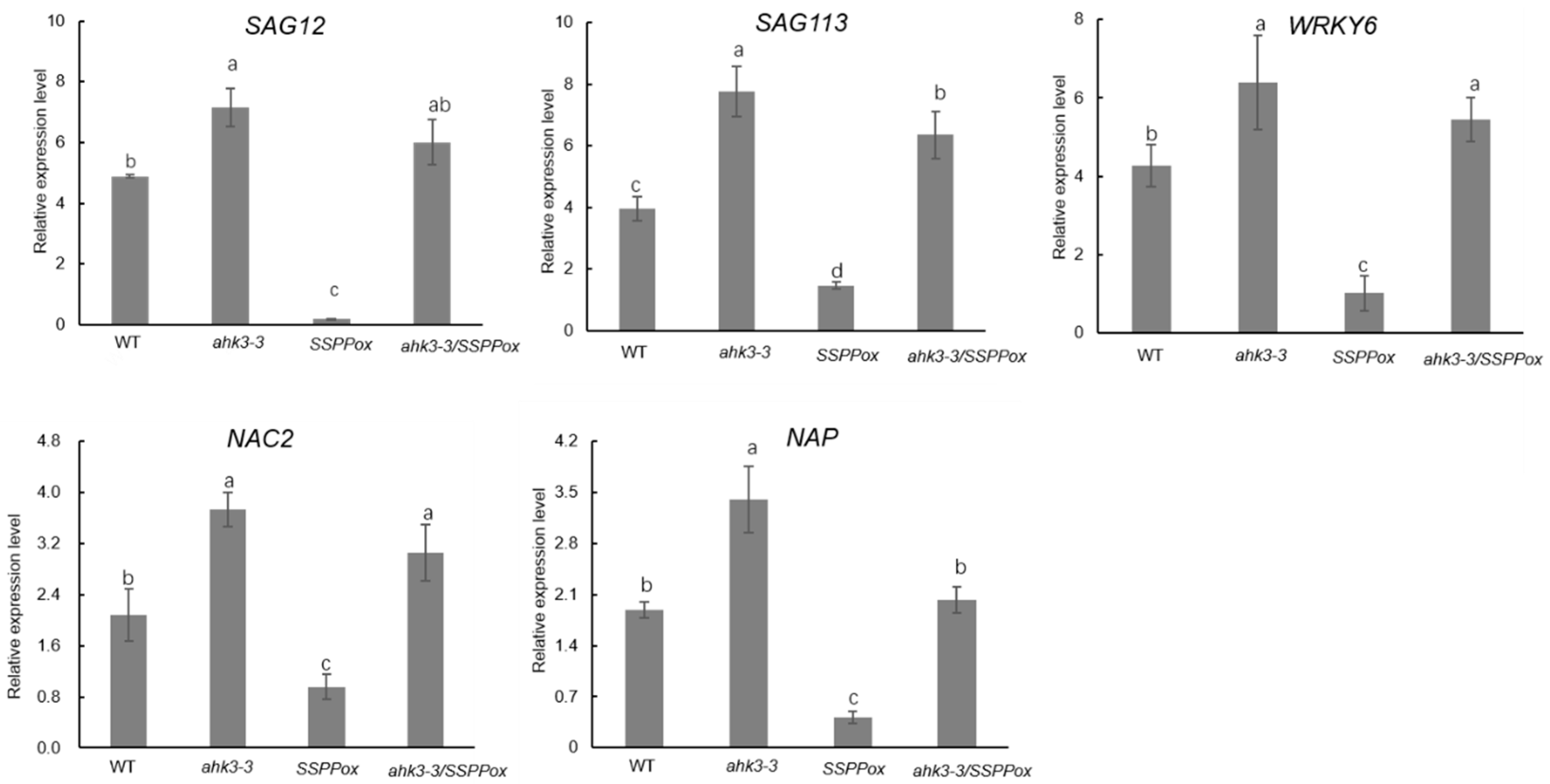
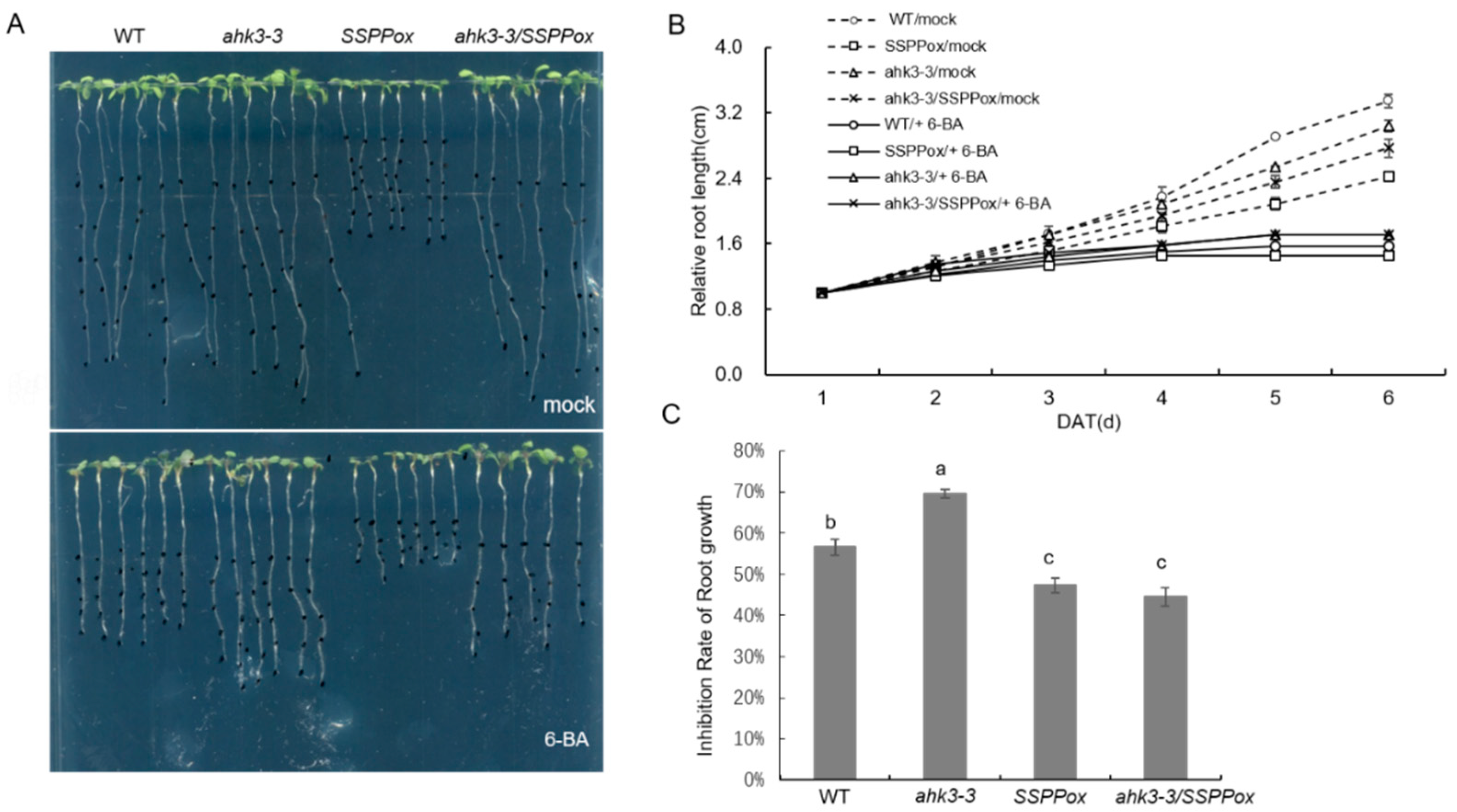
2. Results and Discussion
3. Materials and Methods
3.1. Plant Material and Growth Condition
3.2. Generation of Hybrid Arabidopsis
3.3. Measurements of Chlorophyll Contents
3.4. RNA Extraction and RT-PCR Analysis of Gene Expression
3.5. Cytokinin Response Assay
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| DAE | Days After Emergence |
| DAT | Days After Treatment |
| LRR-RLK | Leucine-Rich Repeat Receptor-Like Protein Kinase |
References
- Kim, J.; Kim, J.H.; Lyu, J.I.; Woo, H.R.; Lim, P.O. New insights into the regulation of leaf senescence in Arabidopsis. J. Exp. Bot. 2017, 69, 787–799. [Google Scholar] [CrossRef]
- Lim, P.O.; Nam, H.G. Aging and senescence of the leaf organ. J. Exp. Bot. 2007, 50, 291–300. [Google Scholar] [CrossRef]
- Zhao, L.; Xia, Y.; Wu, X.Y.; Schippers, J.H.M.; Jing, H.C. Phenotypic Analysis and Molecular Markers of Leaf Senescence. In Plant Senescence. Methods in Molecular Biology; Guo, Y., Ed.; Humana Press: New York, NY, USA, 2018; Volume 1744. [Google Scholar]
- Bowman, J.L.; Kohchi, T.; Yamato, K.T.; Jenkins, J.; Shu, S.Q.; Ishizaki, K.; Yamato, S.; Nishihama, R.; Nakamura, Y.; Berger, F.; et al. Insights into Land Plant Evolution Garnered from the Marchantia polymorpha Genome. Cell 2017, 171, 287–304. [Google Scholar] [CrossRef] [PubMed]
- Jibran, R.; Hunter, D.A.; Dijkwel, P.P. Hormonal regulation of leaf senescence through integration of developmental and stress signals. Plant Mol. Biol. 2013, 82, 547–561. [Google Scholar] [CrossRef]
- Inoue, T.; Higuchi, M.; Hashimoto, Y.; Seki, M.; Kobayashi, M.; Kato, T.; Tabata, S.; Shinozaki, K.; Kakimoto, T. Identification of CRE1 as a cytokinin receptor from Arabidopsis. Nature 2001, 409, 1060–1063. [Google Scholar] [CrossRef]
- Suzuki, T.; Miwa, K.; Ishikawa, K.; Yamada, H.; Aiba, H.; Mizunoet, T. The Arabidopsis sensor His-kinase, AHK4, can respond to cytokinins. Plant Cell Physiol. 2001, 42, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Ueguchi, C.; Koizumi, H.; Suzuki, T.; Mizuno, T. Novel family of sensor histidine kinase genes in Arabidopsis thaliana. Plant Cell Physiol. 2001, 42, 231–235. [Google Scholar] [CrossRef]
- Kim, H.J.; Ryu, H.; Hong, S.H.; Woo, H.R.; Lim, P.O.; Lee, I.C.; Sheen, J.; Nam, H.G.; Hwang, I. Cytokinin-mediated control of leaf longevity by AHK3 through phosphorylation of ARR2 in Arabidopsis. Proc. Natl. Acad. Sci. USA 2006, 103, 814–819. [Google Scholar] [CrossRef] [PubMed]
- Zwack, P.J.; Rashotte, A.M. Cytokinin inhibition of leaf senescence. Plant Signal Behav. 2013, 8, e24737. [Google Scholar] [CrossRef] [PubMed]
- Raines, T.; Shanks, C.; Cheng, C.Y.; McPherson, D.; Argueso, C.T.; Kim, H.J.; Franco-Zorrilla, J.M.; López-Vidriero, I.; Solano, R.; Vaňková, R.; et al. The cytokinin response factors modulate root and shoot growth and promote leaf senescence in Arabidopsis. Plant J. 2016, 85, 134–147. [Google Scholar] [CrossRef]
- Balibrea Lara, M.E.; Gonzalez Garcia, M.C.; Fatima, T.; Ehness, R.; Lee, T.K.; Proels, R.; Tanner, W.; Roitsch, T. Extracellular Invertase Is an Essential Component of Cytokinin-Mediated Delay of Senescence. Plant Cell 2004, 16, 1276–1287. [Google Scholar] [CrossRef]
- Xiao, D.; Cui, Y.J.; Xu, F.; Xu, X.X.; Gao, G.X.; Wang, Y.X.; Guo, Z.X.; Wang, D.; Wang, N.N. The protein phosphatase SSPP directly interacts with the cytoplasmic domain of AtSARK and negatively regulates leaf senescence in Arabidopsis. Plant Physiol. 2015, 169, 1275–1291. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, M.; Pischke, M.S.; Mahonen, A.P.; Miyawaki, K.; Hashimoto, Y.; Seki, M.; Kobayashi, M.; Shinozaki, K.; Kato, T.; Tabata, S.; et al. In planta functions of the Arabidopsis cytokinin receptor family. Proc. Natl. Acad. Sci. USA 2004, 101, 8821–8826. [Google Scholar] [CrossRef]
- Gan, S.; Amasino, R.M. Inhibition of Leaf Senescence by Autoregulated Production of Cytokinin. Science 1995, 270, 1986–1988. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Gan, S. An abscisic acid-AtNAP transcription factor-SAG113 protein phosphatase 2C regulatory chain for controlling dehydration in senescing Arabidopsis leaves. Plant Physiol. 2012, 158, 961–969. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Gan, S. AtNAP, a NAC family transcription factor, has an important role in leaf senescence. Plant J. 2006, 46, 601–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robatzek, S.; Somssich, I.E. Targets of AtWRKY6 regulation during plant senescence and pathogen defense. Genes Dev. 2002, 16, 1139–1149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.J.; Woo, H.R.; Kim, J.; Lim, P.O.; Lee, I.C.; Choi, S.H.; Hwang, D.; Nam, H.G. Trifurcate feed-forward regulation of age-dependent cell death involving miR164 in Arabidopsis. Science 2009, 323, 1053–1057. [Google Scholar] [CrossRef]
- Danilova, M.N.; Kudryakova, N.V.; Doroshenko, A.S.; Zabrodin, D.A.; Rakhmankulova, Z.F.; Oelmüller, R.; Kusnetsov, V.V. Opposite roles of the Arabidopsis cytokinin receptors AHK2 and AHK3 in the expression of plastid genes and genes for the plastid transcriptional machinery during senescence. Plant Mol. Biol. 2017, 93, 533–546. [Google Scholar] [CrossRef] [PubMed]
- Caesar, K.; Thamm, A.M.; Witthöft, J. Evidence for the localization of the Arabidopsis cytokinin receptors AHK3 and AHK4 in the endoplasmic reticulum. J. Exp. Bot. 2011, 62, 5571–5580. [Google Scholar] [CrossRef]
- Xu, F.; Meng, T.; Li, P.; Yu, Y.; Cui, Y.J.; Wang, Y.X.; Gong, Q.Q.; Wang, N.N. A Soybean Dual-Specificity Kinase, GmSARK, and Its Arabidopsis Homolog, AtSARK, Regulate Leaf Senescence through Synergistic Actions of Auxin and Ethylene. Plant Physiol. 2011, 157, 2131–2153. [Google Scholar] [CrossRef] [PubMed]
- Arnon, D.I. Copper enzymes in isolated chloroplasts: Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Gong, Q.; Ma, Y.; Li, P.; Li, J.; Yang, S.; Yuan, L.; Yu, Y.; Pan, D.; Xu, F.; et al. cpSecA, a thylakoid protein translocase subunit, is essential for photosynthetic development in Arabidopsis. J. Exp. Bot. 2010, 61, 1655–1669. [Google Scholar] [CrossRef] [PubMed]
- Xiong, L.; Xiao, D.; Xu, X.; Guo, Z.; Wang, N.N. The non-catalytic N-terminal domain of ACS7 is involved in the post-translational regulation of this gene in Arabidopsis. J. Exp. Bot. 2014, 65, 4397–4408. [Google Scholar] [CrossRef] [PubMed]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Zhang, X.; Cui, Y.; Li, L.; Wang, D.; Mei, Y.; Wang, N.N. AHK3-Mediated Cytokinin Signaling Is Required for the Delayed Leaf Senescence Induced by SSPP. Int. J. Mol. Sci. 2019, 20, 2043. https://doi.org/10.3390/ijms20082043
Wang Y, Zhang X, Cui Y, Li L, Wang D, Mei Y, Wang NN. AHK3-Mediated Cytokinin Signaling Is Required for the Delayed Leaf Senescence Induced by SSPP. International Journal of Molecular Sciences. 2019; 20(8):2043. https://doi.org/10.3390/ijms20082043
Chicago/Turabian StyleWang, Yanan, Xiyu Zhang, Yanjiao Cui, Lei Li, Dan Wang, Yuanyuan Mei, and Ning Ning Wang. 2019. "AHK3-Mediated Cytokinin Signaling Is Required for the Delayed Leaf Senescence Induced by SSPP" International Journal of Molecular Sciences 20, no. 8: 2043. https://doi.org/10.3390/ijms20082043
APA StyleWang, Y., Zhang, X., Cui, Y., Li, L., Wang, D., Mei, Y., & Wang, N. N. (2019). AHK3-Mediated Cytokinin Signaling Is Required for the Delayed Leaf Senescence Induced by SSPP. International Journal of Molecular Sciences, 20(8), 2043. https://doi.org/10.3390/ijms20082043