Rottlerin Reduces cAMP/CREB-Mediated Melanogenesis via Regulation of Autophagy



Abstract
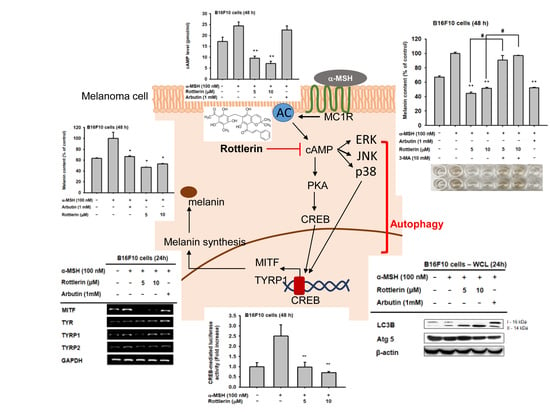
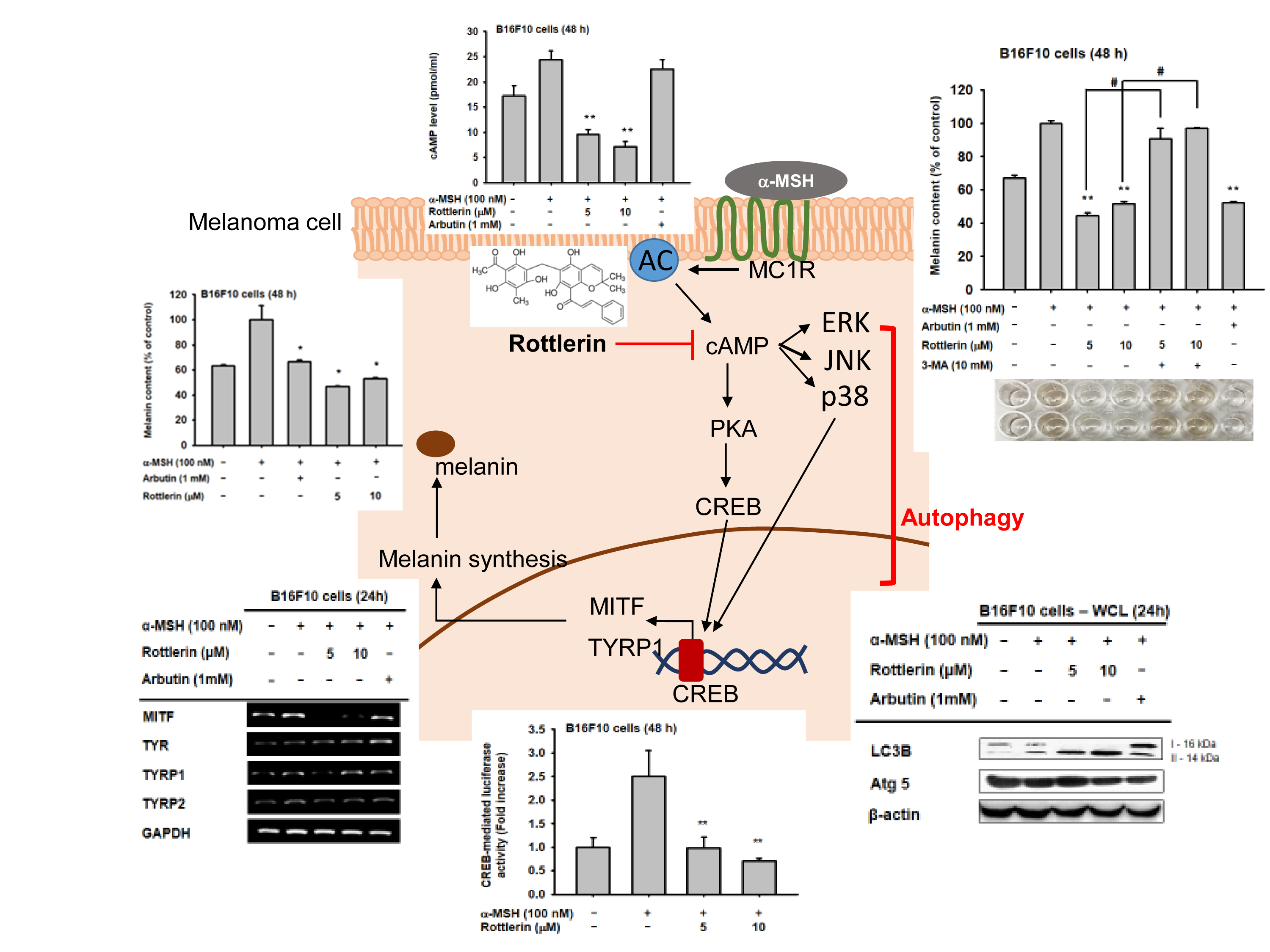
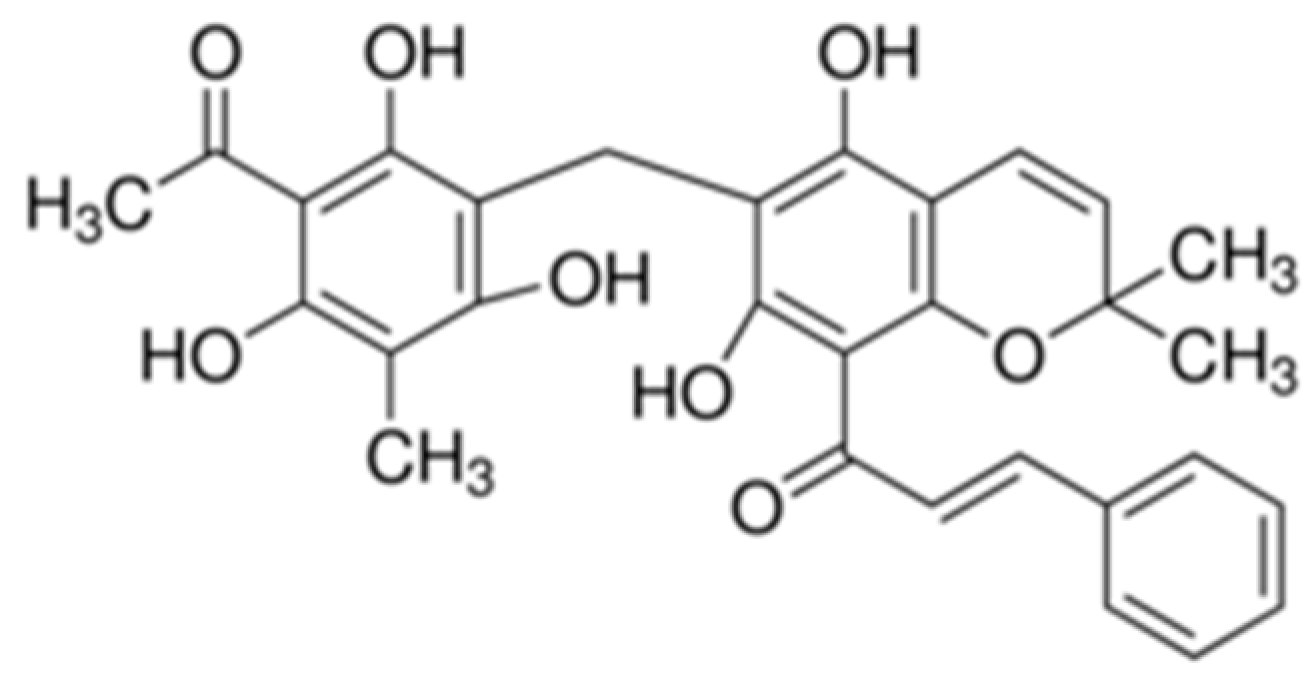
:
1. Introduction
2. Results
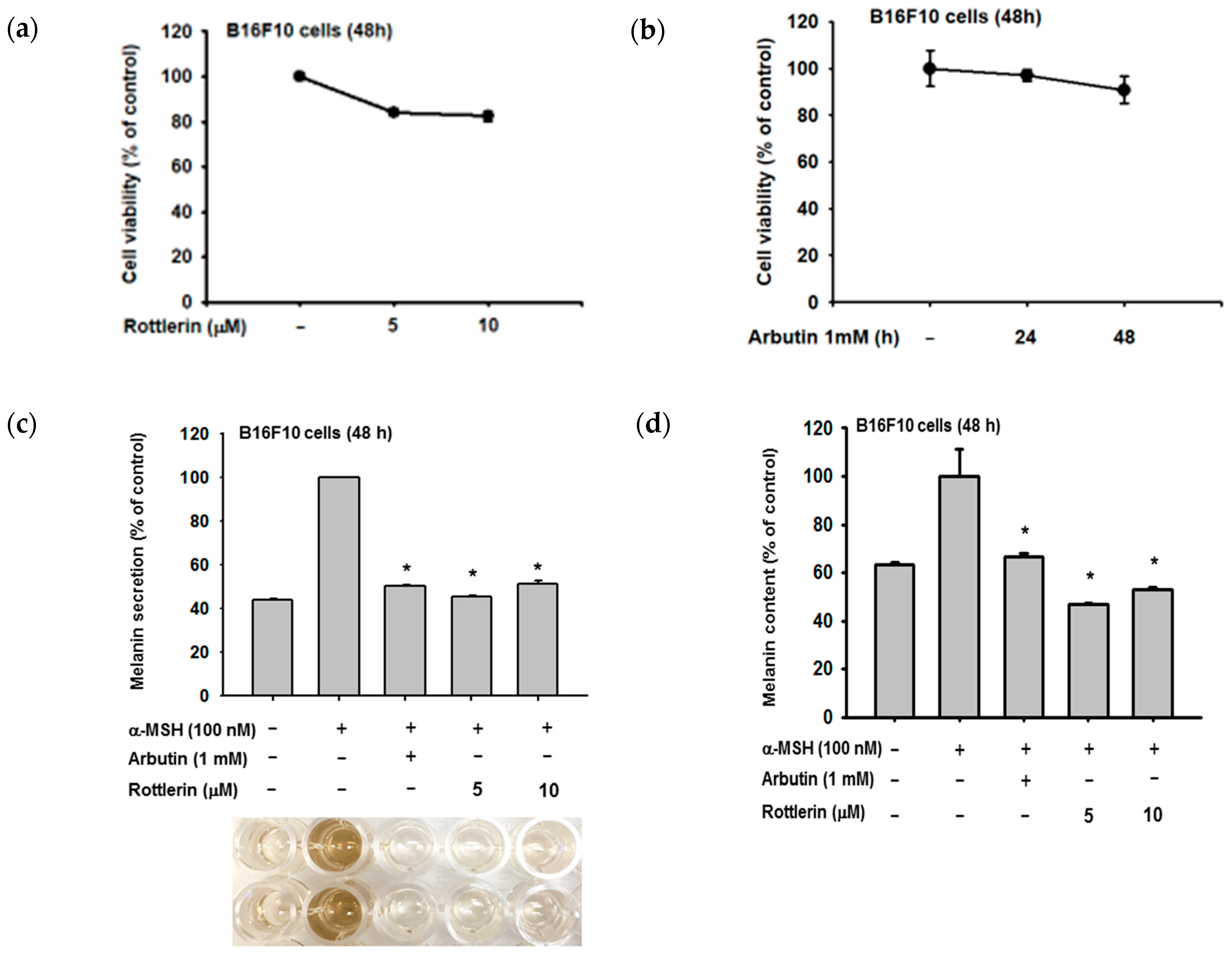
2.1. Effects of Rottlerin on Melanin Produced by Melanoma Cells
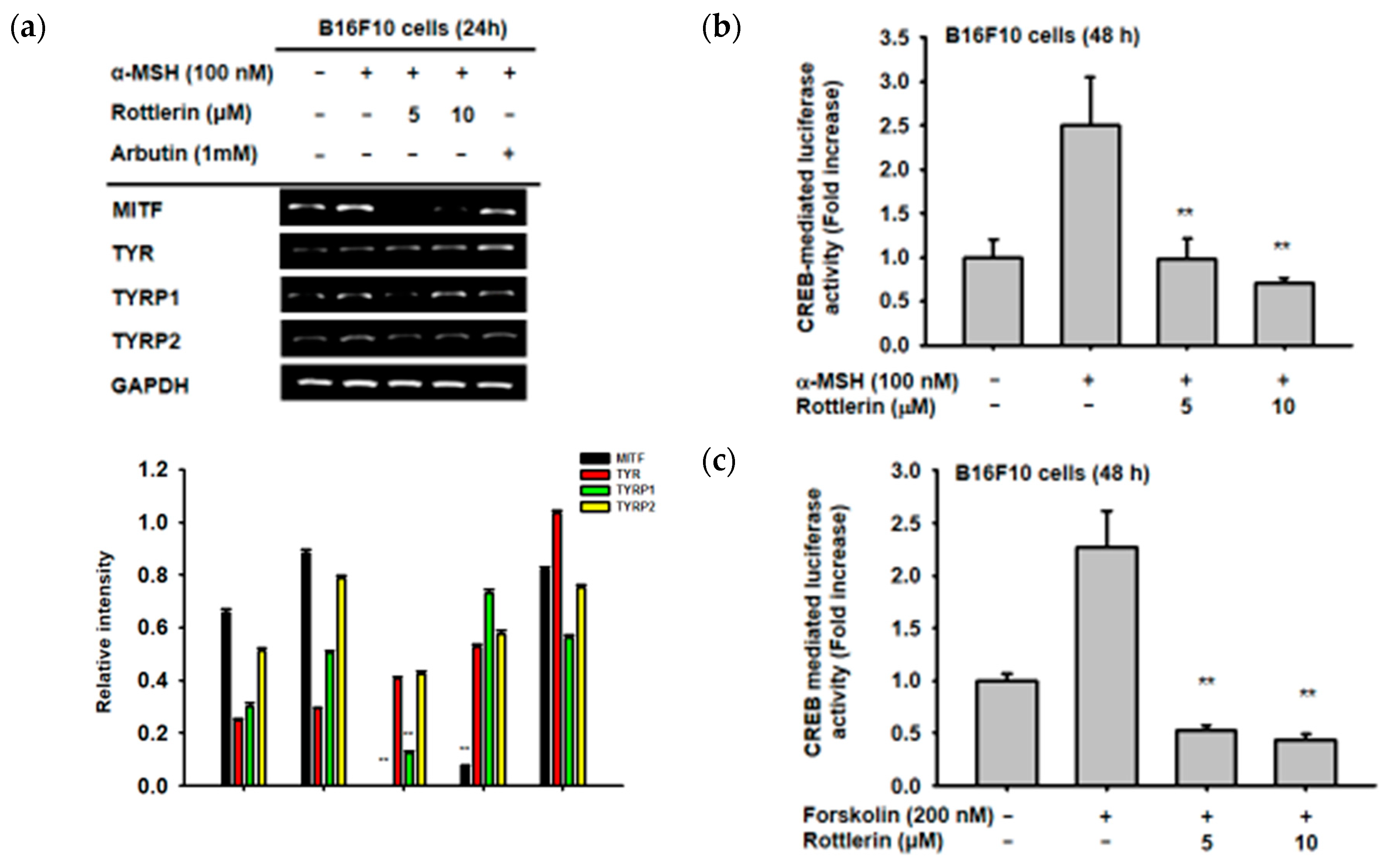
2.2. Effects of Rottlerin on the Expression of Some Genes Related to Melanogenesis
2.3. Effects of Rottlerin on CREB Transcription Factor Activation
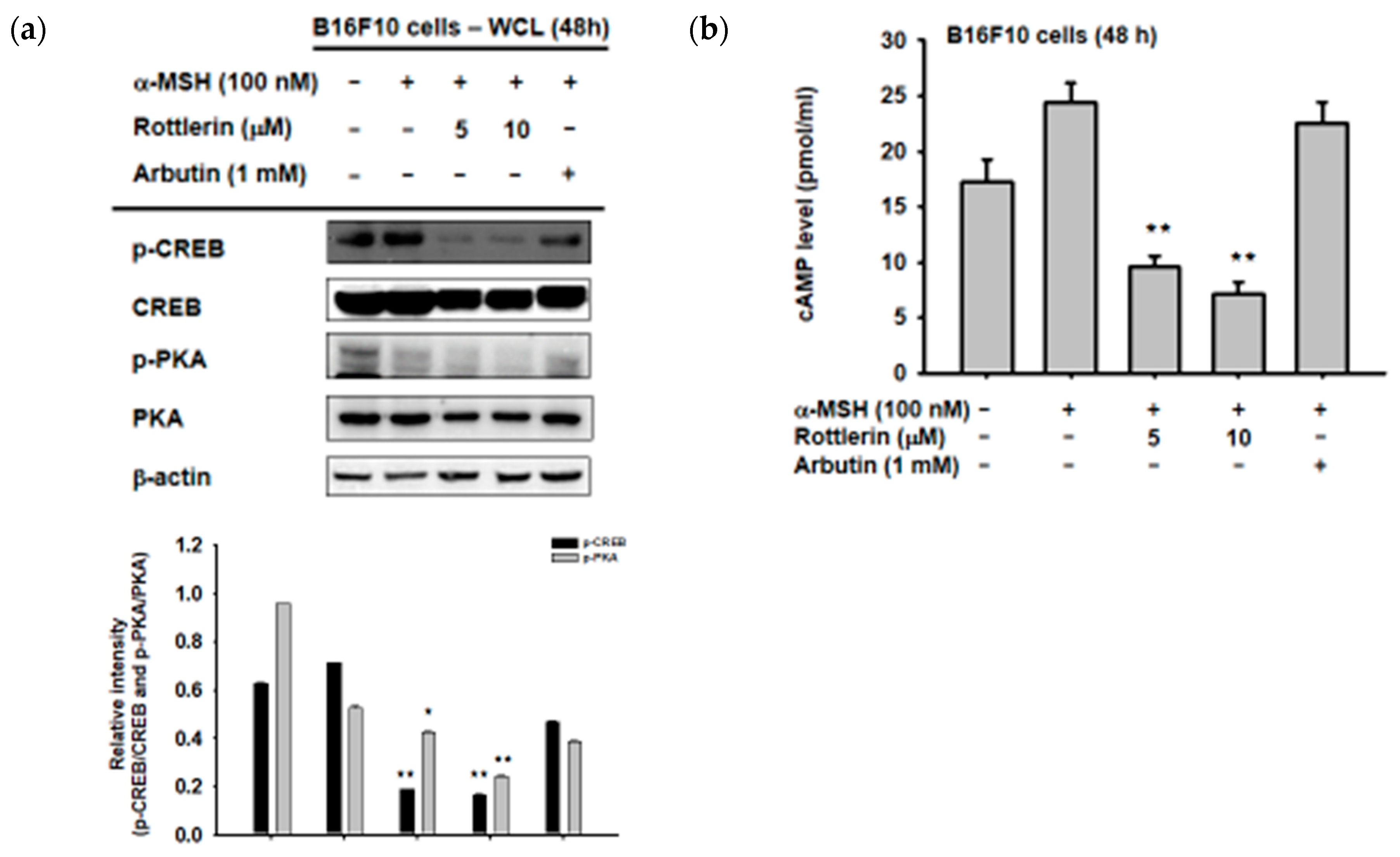
2.4. Effect of Rottlerin on the Melanogenesis-Related Signaling Pathway
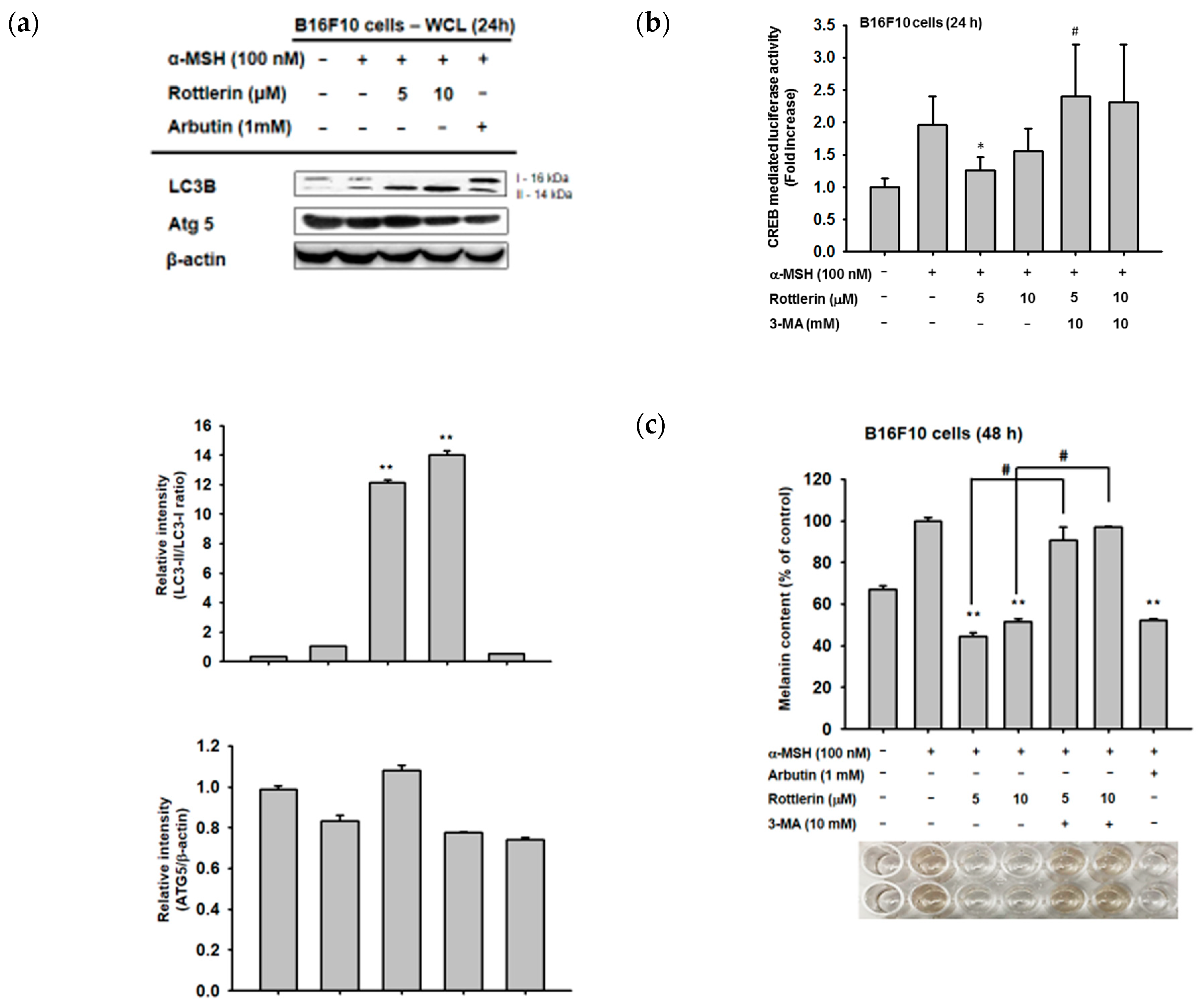
2.5. Effects of Rottlerin on the cAMP/CREB Signaling Pathway by Regulation of Autophagy
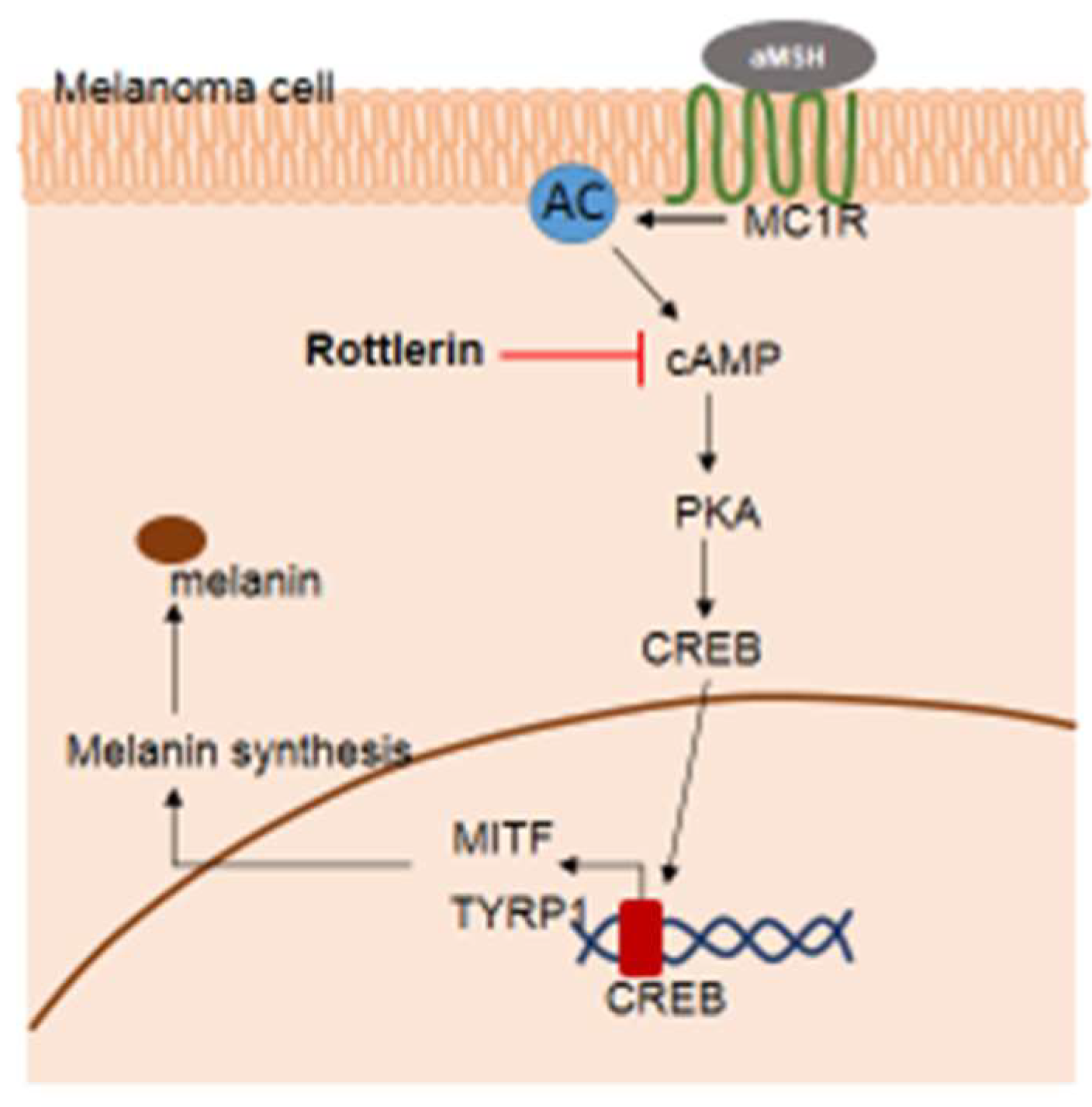
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Drug Preparation, Cell Culture, and Cell Viability Assay
4.3. Determination of Extracellular and Intracellular Melanin Content
4.4. Preparation of Cell Lysate for Immunoblotting
4.5. Measurement of mRNA Levels by Reverse Transcription-Polymerase Chain Reaction
4.6. Luciferase Reporter Gene Assay
4.7. cAMP Assay
4.8. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| UV | Ultraviolet |
| MITF | Microphthalmia-associated transcription factor |
| PCR | Polymerase chain reaction |
| mTOR | Mammalian target of rapamycin |
| mTORC1 | Mammalian target of rapamycin complex 1 |
| α-MSH | Alpha-melanocyte stimulating hormone |
| AMPK | AMP-activated protein kinase |
| TYR | Tyrosinase |
| TYRP1 | Tyrosinase related protein 1 |
| TYRP2 | Tyrosinase related protein 2 |
| cAMP | Cyclic adenosine monophosphate |
| CREB | cAMP response element binding |
| 3-MA | 3-methyladenine |
References
- Kolarsick, P.A.J.; Kolarsick, M.A.; Goodwin, C. Anatomy and physiology of the skin. J. Dermatol. Nurse’s Assoc. 2011, 3, 203–213. [Google Scholar] [CrossRef]
- Dos Santos Videira, I.F.; Moura, D.F.L.; Magina, S. Mechanisms regulating melanogenesis. An. Bras. Dermatol. 2013, 88, 76–83. [Google Scholar] [CrossRef]
- D’Mello, S.A.N.; Finlay, G.J.; Baguley, B.C.; Askarian-Amiri, M.E. Signaling pathways in melanogenesis. Int. J. Mol. Sci. 2016, 17, 1144. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Weng, Q.Y.; Fisher, D.E. UV signaling pathways within the skin. J. Investig. Dermatol. 2014, 134, 2080–2085. [Google Scholar] [CrossRef] [Green Version]
- Chang, T.-S. Natural Melanogenesis Inhibitors Acting Through the Down-Regulation of Tyrosinase Activity. Materials 2012, 5, 1661–1685. [Google Scholar] [Green Version]
- Ho, H.; Ganesan, A.K. The pleiotropic roles of autophagy regulators in melanogenesis. Pigment Cell Melanoma Res. 2011, 24, 595–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maioli, E.; Torricelli, C.; Valacchi, G. Rottlerin and cancer: Novel evidence and mechanisms. Sci. World J. 2012, 2012, 350826. [Google Scholar] [CrossRef]
- Hasima, N.; Ozpolat, B. Regulation of autophagy by polyphenolic compounds as a potential therapeutic strategy for cancer. Cell Death Dis. 2014, 5, e1509. [Google Scholar] [CrossRef] [PubMed]
- Torricelli, C.; Daveri, E.; Salvadori, S.; Valacchi, G.; Ietta, F.; Muscettola, M.; Carlucci, F.; Maioli, E. Phosphorylation-independent mTORC1 inhibition by the autophagy inducer Rottlerin. Cancer Lett. 2015, 360, 17–27. [Google Scholar] [PubMed]
- Singh, B.N.; Kumar, D.; Shankar, S.; Srivastava, R.K. Rottlerin induces autophagy which leads to apoptotic cell death through inhibition of PI3K/Akt/mTOR pathway in human pancreatic cancer stem cells. Biochem. Pharmacol. 2012, 84, 1154–1163. [Google Scholar] [CrossRef] [PubMed]
- Qi, P.; He, Z.; Zhang, L.; Fan, Y.; Wang, Z. Rottlerin-induced autophagy leads to apoptosis in bladder cancer cells. Oncol. Lett. 2016, 12, 4577–4583. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.; Kim, D.; Yoo, S.; Hong, Y.H.; Han, S.Y.; Jeong, S.; Jeong, D.; Kim, J.H.; Cho, J.Y.; Park, J. The skin protective effects of compound K, a metabolite of ginsenoside Rb1 from Panax ginseng. J. Ginseng Res. 2018, 42, 218–224. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.D.; Jin, Y.; Wang, C.; Kim, Y.J.; Perez, Z.E.J.; Baek, N.I.; Mathiyalagan, R.; Markus, J.; Yang, D.C. Rare ginsenoside Ia synthesized from F1 by cloning and overexpression of the UDP-glycosyltransferase gene from Bacillus subtilis: Synthesis, characterization, and in vitro melanogenesis inhibition activity in BL6B16 cells. J. Ginseng Res. 2018, 42, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Choi, E.; Kim, S.; Kim, D.S.; Kim, J.H.; Chang, S.; Choi, J.S.; Park, K.J.; Roh, K.B.; Lee, J.; et al. Oxidative stress-protective and anti-melanogenic effects of loliolide and ethanol extract from fresh water green algae, Prasiola japonica. Int. J. Mol. Sci. 2018, 19, 2825. [Google Scholar] [CrossRef] [PubMed]
- Serre, C.; Busuttil, V.; Botto, J.M. Intrinsic and extrinsic regulation of human skin melanogenesis and pigmentation. Int. J. Cosmet. Sci. 2018, 40, 328–347. [Google Scholar] [CrossRef] [PubMed]
- Im, S.; Moro, O.; Peng, F.; Medrano, E.E.; Cornelius, J.; Babcock, G.; Nordlund, J.J.; Abdel-Malek, Z.A. Activation of the cyclic AMP pathway by α-melanotropin mediates the response of human melanocytes to ultraviolet B radiation. Cancer Res. 1998, 58, 47. [Google Scholar]
- Kim, Y.-M.; Cho, S.-E.; Seo, Y.-K. The activation of melanogenesis by p-CREB and MITF signaling with extremely low-frequency electromagnetic fields on B16-F10 melanoma. Life Sci. 2016, 162, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Song, K.S.; Kim, J.S.; Yun, E.J.; Kim, Y.R.; Seo, K.S.; Park, J.H.; Jung, Y.J.; Park, J.I.; Kweon, G.R.; Yoon, W.H.; et al. Rottlerin induces autophagy and apoptotic cell death through a PKC-delta-independent pathway in HT1080 human fibrosarcoma cells: The protective role of autophagy in apoptosis. Autophagy 2008, 4, 650–658. [Google Scholar] [CrossRef] [Green Version]
- Kumar, D.; Shankar, S.; Srivastava, R.K. Rottlerin induces autophagy and apoptosis in prostate cancer stem cells via PI3K/Akt/mTOR signaling pathway. Cancer Lett. 2014, 343, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Maioli, E.; Daveri, E.; Maellaro, E.; Ietta, F.; Cresti, L.; Valacchi, G. Non-conventional rottlerin anticancer properties. Arch. Biochem. Biophys. 2018, 645, 50–53. [Google Scholar] [CrossRef]
- Tanida, I.; Ueno, T.; Kominami, E. LC3 and Autophagy. In Autophagosome and Phagosome; Deretic, V., Ed.; Humana Press: Totowa, NJ, USA, 2008; pp. 77–88. [Google Scholar]
- Yu, T.; Zuber, J.; Li, J. Targeting autophagy in skin diseases. J. Mol. Med. 2015, 93, 31–38. [Google Scholar] [CrossRef]
- Nishida, Y.; Arakawa, S.; Fujitani, K.; Yamaguchi, H.; Mizuta, T.; Kanaseki, T.; Komatsu, M.; Otsu, K.; Tsujimoto, Y.; Shimizu, S. Discovery of Atg5/Atg7-independent alternative macroautophagy. Nature 2009, 461, 654. [Google Scholar] [CrossRef] [PubMed]
- Takatsuka, C.; Inoue, Y.; Matsuoka, K.; Moriyasu, Y. 3-methyladenine inhibits autophagy in tobacco culture cells under sucrose starvation conditions. Plant Cell Physiol. 2004, 45, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Yang, W.S.; Yu, T.; Yi, Y.-S.; Park, J.G.; Jeong, D.; Kim, J.H.; Oh, J.S.; Yoon, K.; Kim, J.-H.; et al. Novel anti-inflammatory function of NSC95397 by the suppression of multiple kinases. Biochem. Pharmacol. 2014, 88, 201–215. [Google Scholar] [CrossRef]
- Pauwels, R.; Balzarini, J.; Baba, M.; Snoeck, R.; Schols, D.; Herdewijn, P.; Desmyter, J.; De Clercq, E. Rapid and automated tetrazolium-based colorimetric assay for the detection of anti-HIV compounds. J. Virol. Methods 1988, 20, 309–321. [Google Scholar] [CrossRef]
- Qomaladewi, N.P.; Aziz, N.; Kim, M.-Y.; Cho, J.Y. Piper cubeba L. methanol extract has anti-inflammatory activity targeting Src/Syk via NF-B inhibition. Evid.-Based Complement. Altern. Med. 2019, 2019, 1548125. [Google Scholar] [CrossRef]
- Yoo, S.; Kim, M.-Y.; Cho, J.Y. Syk and Src-targeted anti-inflammatory activity of aripiprazole, an atypical antipsychotic. Biochem. Pharmacol. 2018, 148, 1–12. [Google Scholar] [CrossRef]
- Woo, M.-S.; Jung, S.-H.; Kim, S.-Y.; Hyun, J.-W.; Ko, K.-H.; Kim, W.-K.; Kim, H.-S. Curcumin suppresses phorbol ester-induced matrix metalloproteinase-9 expression by inhibiting the PKC to MAPK signaling pathways in human astroglioma cells. Biochem. Biophys. Res. Commun. 2005, 335, 1017–1025. [Google Scholar] [CrossRef]
- Lee, S.E.; Park, S.-H.; Oh, S.W.; Yoo, J.A.; Kwon, K.; Park, S.J.; Kim, J.; Lee, H.S.; Cho, J.Y.; Lee, J. Beauvericin inhibits melanogenesis by regulating cAMP/PKA/CREB and LXR-α/p38 MAPK–mediated pathways. Sci. Rep. 2018, 8, 14958. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Primer | Sequence (5′ to 3′) |
|---|---|---|
| MITF | Forward | AACTCATGCGTGAGCAGATG |
| Reverse | TACCTGGTGCCTCTGAGCTT | |
| TYR | Forward | GTCCACTCACAGGGATAGCAG |
| Reverse | AGAGTCTCTGTTATGGCCGA | |
| TYRP1 | Forward | ATGGAACGGGAGGACAAACC |
| Reverse | TCCTGACCTGGCCATTGAAC | |
| TYRP2 | Forward | CAGTTTCCCCGAGTCTGCAT |
| Reverse | GTCTAAGGCGCCCAAGAACT | |
| GAPDH | Forward | ACCACAGTCCATGCCATCAC |
| Reverse | CCACCACCCTGTTGCTGTAG |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qomaladewi, N.P.; Kim, M.-Y.; Cho, J.Y. Rottlerin Reduces cAMP/CREB-Mediated Melanogenesis via Regulation of Autophagy. Int. J. Mol. Sci. 2019, 20, 2081. https://doi.org/10.3390/ijms20092081
Qomaladewi NP, Kim M-Y, Cho JY. Rottlerin Reduces cAMP/CREB-Mediated Melanogenesis via Regulation of Autophagy. International Journal of Molecular Sciences. 2019; 20(9):2081. https://doi.org/10.3390/ijms20092081
Chicago/Turabian StyleQomaladewi, Nurinanda Prisky, Mi-Yeon Kim, and Jae Youl Cho. 2019. "Rottlerin Reduces cAMP/CREB-Mediated Melanogenesis via Regulation of Autophagy" International Journal of Molecular Sciences 20, no. 9: 2081. https://doi.org/10.3390/ijms20092081
APA StyleQomaladewi, N. P., Kim, M.-Y., & Cho, J. Y. (2019). Rottlerin Reduces cAMP/CREB-Mediated Melanogenesis via Regulation of Autophagy. International Journal of Molecular Sciences, 20(9), 2081. https://doi.org/10.3390/ijms20092081