Receptor Tyrosine Kinases in Development: Insights from Drosophila


Abstract
:1. The Receptor Tyrosine Kinase Protein Superfamily
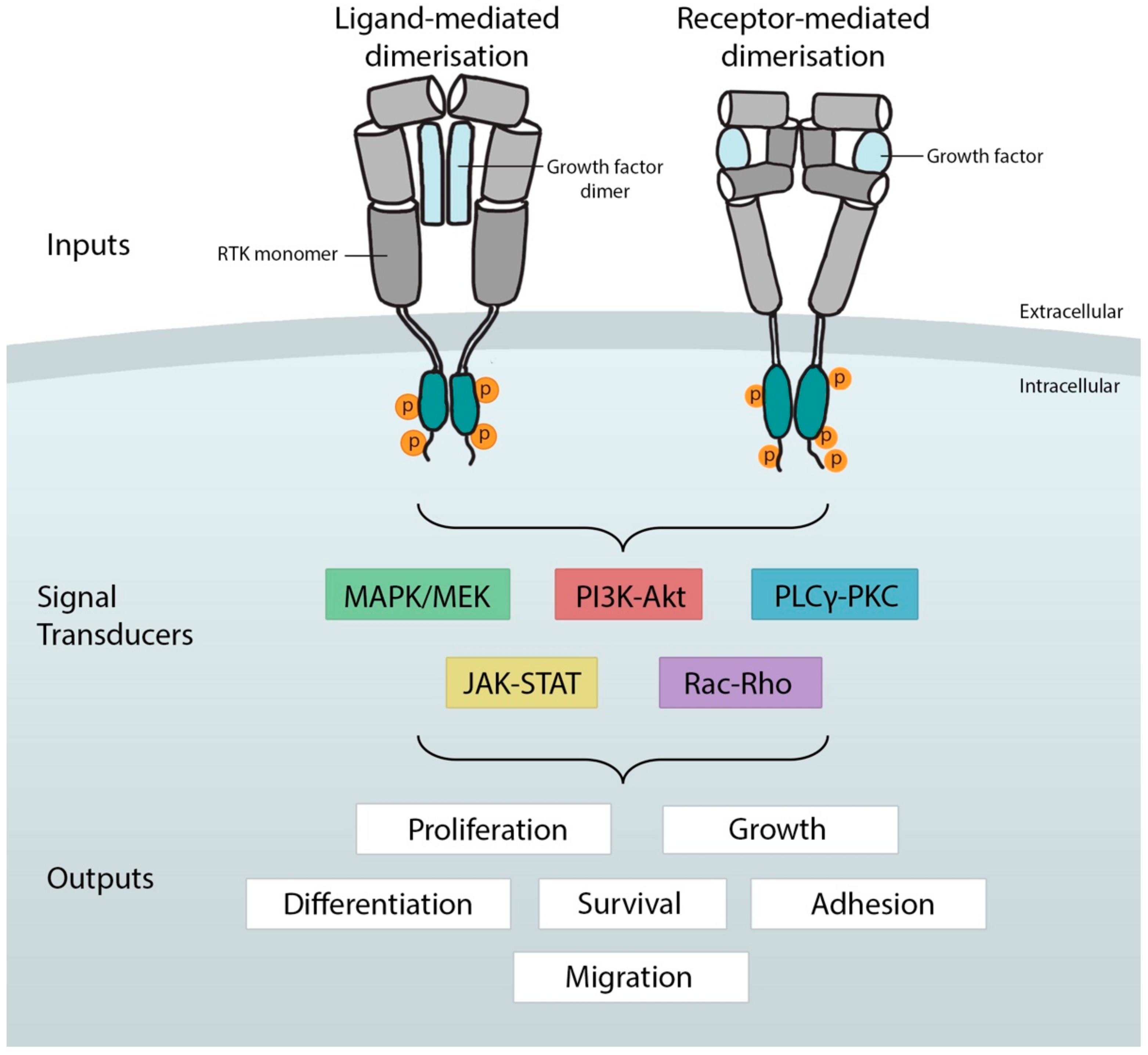
1.1. RTK Structure, Function, and Signaling
1.2. General Mechanisms That Control RTK Activity in Development
2. Drosophila melanogaster as a Model to Study RTK Function in Development
3. Epidermal Growth Factor Receptor
4. Insulin-Like Receptor
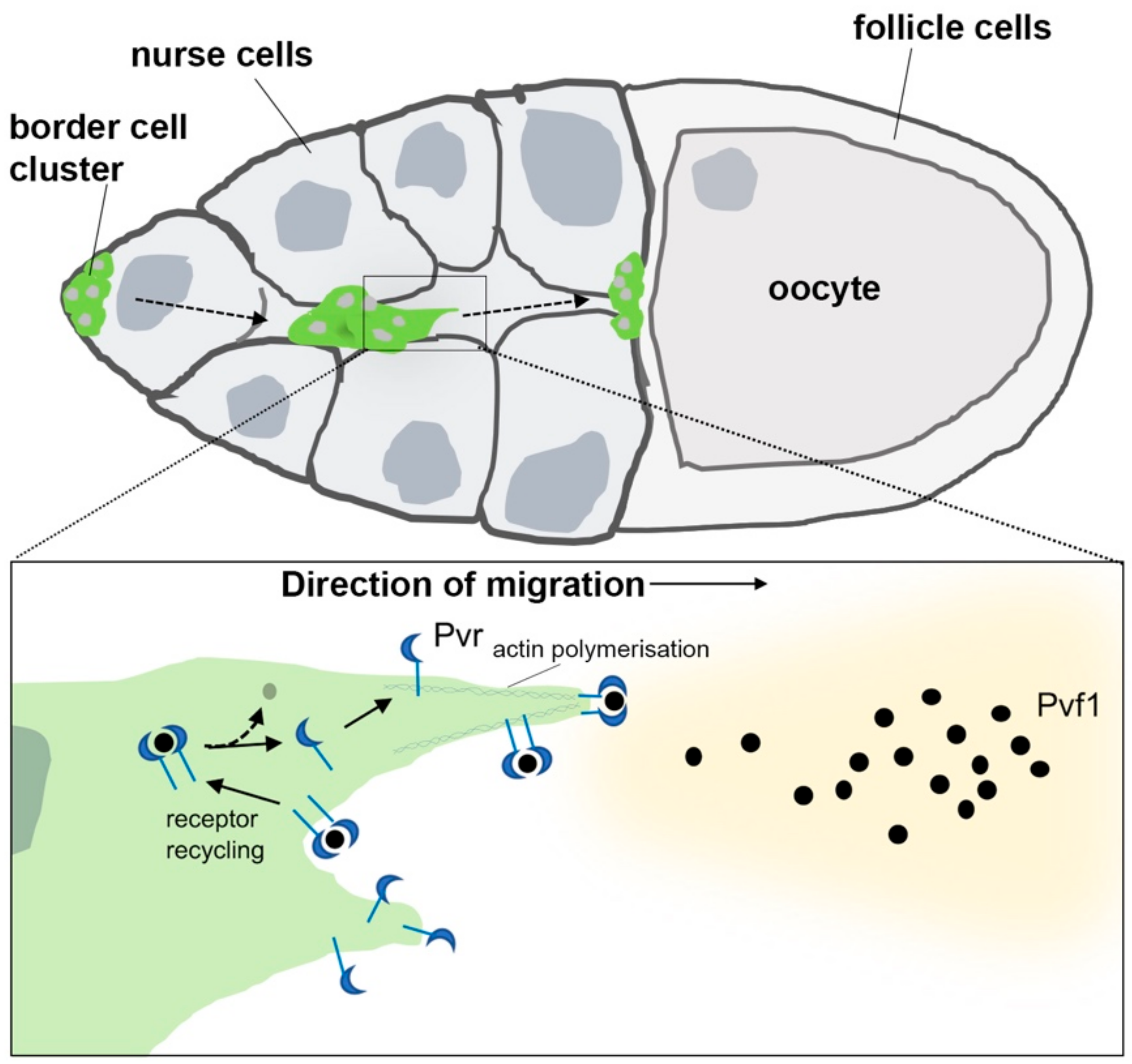
5. The PDGF/VEGF Receptor
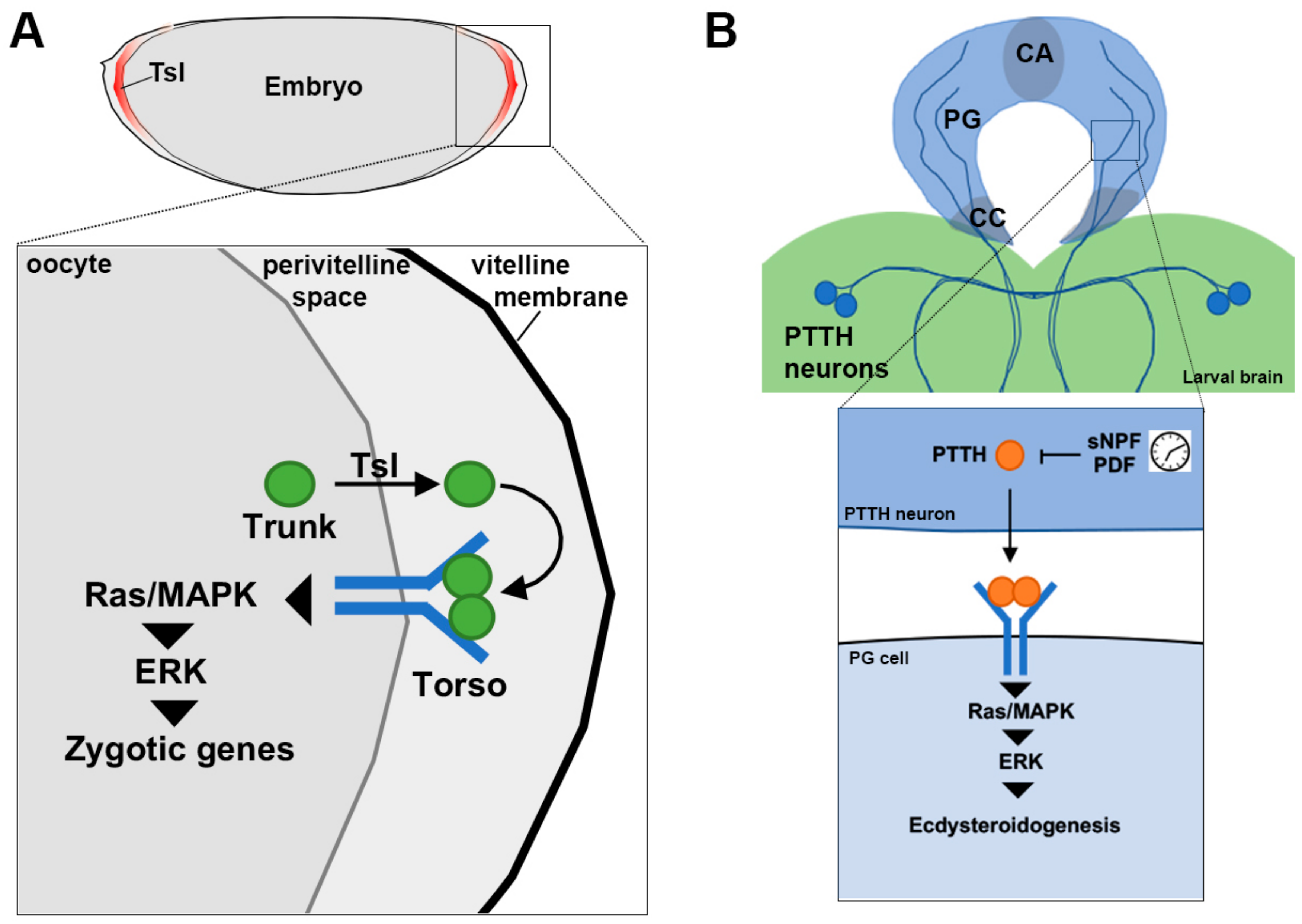
6. Torso in Embryonic Patterning and the Initiation of Metamorphosis
7. Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Schlessinger, J. Receptor tyrosine kinases: Legacy of the first two decades. Cold Spring Harb. Perspect. Biol. 2014, 6. [Google Scholar] [CrossRef] [Green Version]
- Lemmon, M.A.; Schlessinger, J. Cell Signaling by Receptor Tyrosine Kinases. Cell 2010, 141, 1117–1134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perrimon, N.; Pitsouli, C.; Shilo, B.Z. Signaling Mechanisms Controlling Cell Fate and Embryonic Patterning. CSH Perspect. Biol. 2012, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, I.; Lee, H.S. EphB/ephrinB Signaling in Cell Adhesion and Migration. Mol. Cells 2015, 38, 14–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Jack, J.; Garofalo, R.S. The Drosophila insulin receptor is required for normal growth. Endocrinology 1996, 137, 846–856. [Google Scholar] [CrossRef] [PubMed]
- Matsuoka, S.; Hiromi, Y.; Asaoka, M. Egfr signaling controls the size of the stem cell precursor pool in the Drosophila ovary. Mech. Dev. 2013, 130, 241–253. [Google Scholar] [CrossRef]
- Bruckner, K.; Kockel, L.; Duchek, P.; Luque, C.M.; Rorth, P.; Perrimon, N. The PDGF/VEGF receptor controls blood cell survival in Drosophila. Dev. Cell 2004, 7, 73–84. [Google Scholar] [CrossRef] [Green Version]
- Hu, K.; Olsen, B.R. The roles of vascular endothelial growth factor in bone repair and regeneration. Bone 2016, 91, 30–38. [Google Scholar] [CrossRef] [Green Version]
- Du, Z.; Lovly, C.M. Mechanisms of receptor tyrosine kinase activation in cancer. Mol. Cancer 2018, 17, 58. [Google Scholar] [CrossRef]
- Ahluwalia, M.S.; Becker, K.; Levy, B.P. Epidermal Growth Factor Receptor Tyrosine Kinase Inhibitors for Central Nervous System Metastases from Non-Small Cell Lung Cancer. Oncologist 2018, 23, 1199–1209. [Google Scholar] [CrossRef] [Green Version]
- Shen, G.S.; Zheng, F.C.; Ren, D.F.; Du, F.; Dong, Q.X.; Wang, Z.Y.; Zhao, F.X.; Ahmad, R.; Zhao, J.D. Anlotinib: A novel multi-targeting tyrosine kinase inhibitor in clinical development. J. Hematol. Oncol. 2018, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Odawara, M.; Kadowaki, T.; Yamamoto, R.; Shibasaki, Y.; Tobe, K.; Accili, D.; Bevins, C.; Mikami, Y.; Matsuura, N.; Akanuma, Y.; et al. Human Diabetes Associated with a Mutation in the Tyrosine Kinase Domain of the Insulin-Receptor. Science 1989, 245, 66–68. [Google Scholar] [CrossRef] [PubMed]
- Almendro, V.; Garcia-Recio, S.; Gascon, P. Tyrosine kinase receptor transactivation associated to G protein-coupled receptors. Curr. Drug Targets 2010, 11, 1169–1180. [Google Scholar] [CrossRef] [PubMed]
- Mustonen, T.; Alitalo, K. Endothelial Receptor Tyrosine Kinases Involved in Angiogenesis. J. Cell Biol. 1995, 129, 895–898. [Google Scholar] [CrossRef]
- McDonell, L.M.; Kernohan, K.D.; Boycott, K.M.; Sawyer, S.L. Receptor tyrosine kinase mutations in developmental syndromes and cancer: Two sides of the same coin. Hum. Mol. Genet. 2015, 24, R60–R66. [Google Scholar] [CrossRef] [Green Version]
- Choura, M.; Rebai, A. Receptor tyrosine kinases: From biology to pathology. J. Recept. Signal Transduct. 2011, 31, 387–394. [Google Scholar] [CrossRef]
- Pawson, T. Regulation and targets of receptor tyrosine kinases. Eur. J. Cancer 2002, 38, S3–S10. [Google Scholar] [CrossRef]
- Kroiher, M.; Miller, M.A.; Steele, R.E. Deceiving appearances: Signaling by “dead” and “fractured” receptor protein tyrosine kinases. Bioessays 2001, 23, 69–76. [Google Scholar] [CrossRef]
- Shi, X.; Hapiak, V.; Zheng, J.; Muller-Greven, J.; Bowman, D.; Lingerak, R.; Buck, M.; Wang, B.C.; Smith, A.W. A role of the SAM domain in EphA2 receptor activation. Sci. Rep. 2017, 7, 45084. [Google Scholar] [CrossRef] [Green Version]
- Molina, L.; Marino-Buslje, C.; Quinn, D.R.; Siddle, K. Structural domains of the insulin receptor and IGF receptor required for dimerisation and ligand binding. FEBS Lett. 2000, 467, 226–230. [Google Scholar] [CrossRef] [Green Version]
- Hanks, S.K.; Quinn, A.M.; Hunter, T. The Protein-Kinase Family—Conserved Features and Deduced Phylogeny of the Catalytic Domains. Science 1988, 241, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Manning, G.; Plowman, G.D.; Hunter, T.; Sudarsanam, S. Evolution of protein kinase signaling from yeast to man. Trends Biochem. Sci. 2002, 27, 514–520. [Google Scholar] [CrossRef]
- Bae, J.H.; Schlessinger, J. Asymmetric tyrosine kinase arrangements in activation or autophosphorylation of receptor tyrosine kinases. Mol. Cells 2010, 29, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Maruyama, I.N. Nerve growth factor receptor TrkA exists as a preformed, yet inactive, dimer in living cells. FEBS Lett. 2011, 585, 295–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wehrman, T.; He, X.L.; Raab, B.; Dukipatti, A.; Blau, H.; Garcia, K.C. Structural and mechanistic insights into nerve growth factor interactions with the TrkA and p75 receptors. Neuron 2007, 53, 25–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burgess, A.W.; Cho, H.S.; Eigenbrot, C.; Ferguson, K.M.; Garrett, T.P.J.; Leahy, D.J.; Lemmon, M.A.; Sliwkowski, M.X.; Ward, C.W.; Yokoyama, S. An open-and-shut case? Recent insights into the activation of EGF/ErbB receptors. Mol. Cell 2003, 12, 541–552. [Google Scholar] [CrossRef]
- Yayon, A.; Klagsbrun, M.; Esko, J.D.; Leder, P.; Ornitz, D.M. Cell-Surface, Heparin-Like Molecules Are Required for Binding of Basic Fibroblast Growth-Factor to Its High-Affinity Receptor. Cell 1991, 64, 841–848. [Google Scholar] [CrossRef]
- Yuzawa, S.; Opatowsky, Y.; Zhang, Z.T.; Mandiyan, V.; Lax, I.; Schlessinger, J. Structural basis for activation of the receptor tyrosine kinase KIT by stem cell factor. Cell 2007, 130, 323–334. [Google Scholar] [CrossRef] [Green Version]
- Pawson, T.; Gish, G.D.; Nash, P. SH2 domains, interaction modules and cellular wiring. Trends Cell Biol. 2001, 11, 504–511. [Google Scholar] [CrossRef]
- Hubbard, S.R. Juxtamembrane autoinhibition in receptor tyrosine kinases. Nat. Rev. Mol. Cell Biol. 2004, 5, 464–470. [Google Scholar] [CrossRef]
- Sun, Y.; Liu, W.Z.; Liu, T.; Feng, X.; Yang, N.; Zhou, H.F. Signaling pathway of MAPK/ERK in cell proliferation, differentiation, migration, senescence and apoptosis. J. Recept. Signal Transduct. 2015, 35, 600–604. [Google Scholar] [CrossRef] [PubMed]
- Krishna, M.; Narang, H. The complexity of mitogen-activated protein kinases (MAPKs) made simple. Cell. Mol. Life Sci. 2008, 65, 3525–3544. [Google Scholar] [CrossRef] [PubMed]
- Manning, B.D.; Cantley, L.C. AKT/PKB signaling: Navigating downstream. Cell 2007, 129, 1261–1274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, T.L.; Wulf, G.; Burga, L.; Cantley, L.C. Cell-to-Cell Variability in PI3K Protein Level Regulates PI3K-AKT Pathway Activity in Cell Populations. Curr. Biol. 2011, 21, 173–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bunney, T.D.; Katan, M. PLC regulation: Emerging pictures for molecular mechanisms. Trends Biochem. Sci. 2011, 36, 88–96. [Google Scholar] [CrossRef]
- Shuai, K. Modulation of STAT signaling by STAT-interacting proteins. Oncogene 2000, 19, 2638–2644. [Google Scholar] [CrossRef] [Green Version]
- Wittes, J.; Schupbach, T. A Gene Expression Screen in Drosophila melanogaster Identifies Novel JAK/STAT and EGFR Targets During Oogenesis. G3-Genes Genomes Genet. 2019, 9, 47–60. [Google Scholar] [CrossRef] [Green Version]
- Xu, N.; Wang, S.Q.; Tan, D.; Gao, Y.W.; Lin, G.N.; Xi, R.W. EGFR, Wingless and JAK/STAT signaling cooperatively maintain Drosophila intestinal stem cells. Dev. Biol. 2011, 354, 31–43. [Google Scholar] [CrossRef] [Green Version]
- Simon, A.R.; Takahashi, S.; Severgnini, M.; Fanburg, B.L.; Cochran, B.H. Role of the JAK-STAT pathway in PDGF-stimulated proliferation of human airway smooth muscle cells. Am. J. Physiol. Lung Cell. 2002, 282, L1296–L1304. [Google Scholar] [CrossRef] [Green Version]
- Li, W.X.; Agaisse, H.; Mathey-Prevot, B.; Perrimon, N. Differential requirement for STAT by gain-of-function and wild-type receptor tyrosine kinase Torso in Drosophila. Development 2002, 129, 4241–4248. [Google Scholar]
- Casaletto, J.B.; McClatchey, A.I. Spatial regulation of receptor tyrosine kinases in development and cancer. Nat. Rev. Cancer 2012, 12, 387–400. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, G.; Smith, A.W.; Jones, B. Discoidin domain receptors: Micro insights into macro assemblies. BBA-Mol. Cell Res. 2019, 1866. [Google Scholar] [CrossRef] [PubMed]
- Delos Santos, R.C.; Garay, C.; Antonescu, C.N. Charming neighborhoods on the cell surface: Plasma membrane microdomains regulate receptor tyrosine kinase signaling. Cell. Signal. 2015, 27, 1963–1976. [Google Scholar] [CrossRef] [PubMed]
- Volinsky, N.; Kholodenko, B.N. Complexity of Receptor Tyrosine Kinase Signal Processing. CSH Perspect. Biol. 2013, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simons, M.; Gordon, E.; Claesson-Welsh, L. Mechanisms and regulation of endothelial VEGF receptor signalling. Nat. Rev. Mol. Cell Biol. 2016, 17, 611–625. [Google Scholar] [CrossRef] [PubMed]
- Zinkle, A.; Mohammadi, M. A threshold model for receptor tyrosine kinase signaling specificity and cell fate determination. F1000Research 2018, 7. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, M.; Nakayama, A.; van Lessen, M.; Yamamoto, H.; Hoffmann, S.; Drexler, H.C.A.; Itoh, N.; Hirose, T.; Breier, G.; Vestweber, D.; et al. Spatial regulation of VEGF receptor endocytosis in angiogenesis. Nat. Cell Biol. 2013, 15, 249–260. [Google Scholar] [CrossRef] [Green Version]
- Klein, D.E.; Nappi, V.M.; Reeves, G.T.; Shvartsman, S.Y.; Lemmon, M.A. Argos inhibits epidermal growth factor receptor signalling by ligand sequestration. Nature 2004, 430, 1040–1044. [Google Scholar] [CrossRef]
- Matkar, S.; An, C.Y.; Hua, X.X. Kinase inhibitors of HER2/AKT pathway induce ERK phosphorylation via a FOXO-dependent feedback loop. Am. J. Cancer Res. 2017, 7, 1476–1485. [Google Scholar]
- Zhang, X.C.; Lavoie, G.; Meant, A.; Aubert, L.; Cargnello, M.; Haman, A.; Hoang, T.; Roux, P.P. Extracellular Signal-Regulated Kinases 1 and 2 Phosphorylate Gab2 To Promote a Negative-Feedback Loop That Attenuates Phosphoinositide 3-Kinase/Akt Signaling. Mol. Cell. Biol. 2017, 37. [Google Scholar] [CrossRef] [Green Version]
- Freeman, M. Reiterative use of the EGF receptor triggers differentiation of all cell types in the Drosophila eye. Cell 1996, 87, 651–660. [Google Scholar] [CrossRef] [Green Version]
- Jekely, G.; Sung, H.H.; Luque, C.M.; Rorth, P. Regulators of endocytosis maintain localized receptor tyrosine kinase signaling in guided migration. Dev. Cell 2005, 9, 197–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, B.; Dsilva, C.J.; Levario, T.J.; Lu, H.; Schupbach, T.; Kevrekidis, I.G.; Shvartsman, S.Y. Dynamics of Inductive ERK Signaling in the Drosophila Embryo. Curr. Biol. 2015, 25, 1784–1790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oldham, S.; Stocker, H.; Laffargue, M.; Wittwer, F.; Wymann, M.; Hafen, E. The Drosophila insulin/IGF receptor controls growth and size by modulating PtdInsP(3) levels. Development 2002, 129, 4103–4109. [Google Scholar] [PubMed]
- Urban, S.; Lee, J.R.; Freeman, M. Drosophila Rhomboid-1 defines a family of putative intramembrane serine proteases. Cell 2001, 107, 173–182. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.Q.; Tsarouhas, V.; Xylourgidis, N.; Sabri, N.; Tiklova, K.; Nautiyal, N.; Gallio, M.; Samakovlis, C. The tyrosine kinase Stitcher activates Grainy head and epidermal wound healing in Drosophila. Nat. Cell Biol. 2009, 11, 890. [Google Scholar] [CrossRef]
- Cheng, L.; Baonza, A.; Grifoni, D. Drosophila Models of Human Disease. BioMed Res. Int. 2018. [Google Scholar] [CrossRef]
- Rubin, G.M.; Yandell, M.D.; Wortman, J.R.; Miklos, G.L.G.; Nelson, C.R.; Hariharan, I.K.; Fortini, M.E.; Li, P.W.; Apweiler, R.; Fleischmann, W.; et al. Comparative genomics of the eukaryotes. Science 2000, 287, 2204–2215. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, S.P.; Hastings, J.F.; Han, J.Z.R.; Croucher, D.R. The Under-Appreciated Promiscuity of the Epidermal Growth Factor Receptor Family. Front. Cell Dev. Biol. 2016, 4. [Google Scholar] [CrossRef] [Green Version]
- Shilo, B.Z. Signaling by the Drosophila epidermal growth factor receptor pathway during development. Exp. Cell Res. 2003, 284, 140–149. [Google Scholar] [CrossRef]
- Vivekanand, P. Lessons from Drosophila Pointed, an ETS family transcription factor and key nuclear effector of the RTK signaling pathway. Genesis 2018, 56. [Google Scholar] [CrossRef] [PubMed]
- Zak, N.B.; Wides, R.J.; Schelter, E.D.; Raz, E.; Shilo, B.Z. Localization of the Der/Flb Protein in Embryos—Implications on the Faint Little Ball Lethal Phenotype. Development 1990, 109, 865–874. [Google Scholar] [PubMed]
- Shilo, B.Z. Regulating the dynamics of EGF receptor signaling in space and time. Development 2005, 132, 4017–4027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casanova, J.; Struhl, G. Localized Surface-Activity of Torso, a Receptor Tyrosine Kinase, Specifies Terminal Body Pattern in Drosophila. Genes Dev. 1989, 3, 2025–2038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sprenger, F.; Stevens, L.M.; Nussleinvolhard, C. The Drosophila Gene Torso Encodes a Putative Receptor Tyrosine Kinase. Nature 1989, 338, 478–483. [Google Scholar] [CrossRef]
- Buff, E.; Carmena, A.; Gisselbrecht, S.; Jimenez, F.; Michelson, A.M. Signalling by the Drosophila epidermal growth factor receptor is required for the specification and diversification of embryonic muscle progenitors. Development 1998, 125, 2075–2086. [Google Scholar]
- Duchek, P.; Rorth, P. Guidance of cell migration by EGF receptor signaling during Drosophila oogenesis. Science 2001, 291, 131–133. [Google Scholar] [CrossRef]
- Skeath, J.B. The Drosophila EGF receptor controls the formation and specification of neuroblasts along the dorsal-ventral axis of the Drosophila embryo. Development 1998, 125, 3301–3312. [Google Scholar]
- Gisselbrecht, S.; Skeath, J.B.; Doe, C.Q.; Michelson, A.M. Heartless encodes a fibroblast growth factor receptor (DFR1/DFGF-R2) involved in the directional migration of early mesodermal cells in the Drosophila embryo. Genes Dev. 1996, 10, 3003–3017. [Google Scholar] [CrossRef] [Green Version]
- Beiman, M.; Shilo, B.Z.; Volk, T. Heartless, a Drosophila FGF receptor homolog, is essential for cell migration and establishment of several mesodermal lineages. Genes Dev. 1996, 10, 2993–3002. [Google Scholar] [CrossRef] [Green Version]
- Klambt, C.; Glazer, L.; Shilo, B.Z. Breathless, a Drosophila Fgf Receptor Homolog, Is Essential for Migration of Tracheal and Specific Midline Glial-Cells. Genes Dev. 1992, 6, 1668–1678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitadate, Y.; Shigenobu, S.; Arita, K.; Kobayashi, S. Boss/Sev signaling from germline to soma specify the germline stem cell niche in Drosophila male gonads. Zool. Sci. 2006, 23, 1183. [Google Scholar]
- Fernandez, R.; Tabarini, D.; Azpiazu, N.; Frasch, M.; Schlessinger, J. The Drosophila Insulin-Receptor Homolog—A Gene Essential for Embryonic-Development Encodes 2 Receptor Isoforms with Different Signaling Potential. EMBO J. 1995, 14, 3373–3384. [Google Scholar] [CrossRef] [PubMed]
- Duchek, P.; Somogyi, K.; Jekely, G.; Beccari, S.; Rorth, P. Guidance of cell migration by the Drosophila PDGF/VEGF receptor. Cell 2001, 107, 17–26. [Google Scholar] [CrossRef] [Green Version]
- Heino, T.I.; Karpanen, T.; Wahlstrom, G.; Pulkkinen, M.; Eriksson, U.; Alitalo, K.; Roos, C. The Drosophila VEGF receptor homolog is expressed in hemocytes. Mech. Dev. 2001, 109, 69–77. [Google Scholar] [CrossRef]
- McDonald, J.A.; Pinheiro, E.M.; Montell, D.J. PVF1, a PDGF/VEGF homolog, is sufficient to guide border cells and interacts genetically with Taiman. Development 2003, 130, 3469–3478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callahan, C.A.; Muralidhar, M.G.; Lundgren, S.E.; Scully, A.L.; Thomas, J.B. Control of Neuronal Pathway Selection by a Drosophila Receptor Protein-Tyrosine Kinase Family Member. Nature 1995, 376, 171–174. [Google Scholar] [CrossRef]
- Harris, K.E.; Beckendorf, S.K. Different Wnt signals act through the Frizzled and RYK receptors during Drosophila salivary gland migration. Development 2007, 134, 2017–2025. [Google Scholar] [CrossRef] [Green Version]
- Oates, A.C.; Bonkovsky, J.L.; Irvine, D.V.; Kelly, L.E.; Thomas, J.B.; Wilks, A.F. Embryonic expression and activity of doughnut, a second RYK homolog in Drosophila. Mech. Dev. 1998, 78, 165–169. [Google Scholar] [CrossRef]
- Englund, C.; Loren, C.E.; Grabbe, C.; Varshney, G.K.; Deleuil, F.; Hallberg, B.; Palmer, R.H. Jeb signals through the Alk receptor tyrosine kinase to drive visceral muscle fusion. Nature 2003, 425, 512–516. [Google Scholar] [CrossRef]
- Gouzi, J.Y.; Moressis, A.; Walker, J.A.; Apostolopoulou, A.A.; Palmer, R.H.; Bernards, A.; Skoulakis, E.M.C. The Receptor Tyrosine Kinase Alk Controls Neurofibromin Functions in Drosophila Growth and Learning. PLoS Genet. 2011, 7, e1002281. [Google Scholar] [CrossRef] [PubMed]
- Paganoni, S.; Ferreira, A. Neurite extension in central neurons: A novel role for the receptor tyrosine kinases Ror1 and Ror2. J. Cell Sci. 2005, 118, 433–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oishi, I.; Sugiyama, S.; Liu, Z.J.; Yamamura, H.; Nishida, Y.; Minami, Y. A novel Drosophila receptor tyrosine kinase expressed specifically in the nervous system—Unique structural features and implication in developmental signaling. J. Biol. Chem. 1997, 272, 11916–11923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winberg, M.L.; Tamagnone, L.; Bai, J.W.; Comoglio, P.M.; Montell, D.; Goodman, C.S. The transmembrane protein off-track associates with plexins and functions downstream of semaphorin signaling during axon guidance. Neuron 2001, 32, 53–62. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.J.; Bo, J.Y.; Bridges, T.; Dugan, K.D.; Pan, T.C.; Chodosh, L.A.; Montell, D.J. Analysis of cell migration using whole-genome expression profiling of migratory cells in the Drosophila ovary. Dev. Cell 2006, 10, 483–495. [Google Scholar] [CrossRef] [Green Version]
- Tomlinson, A.; Bowtell, D.D.L.; Hafen, E.; Rubin, G.M. Localization of the Sevenless Protein, a Putative Receptor for Positional Information, in the Eye Imaginal Disk of Drosophila. Cell 1987, 51, 143–150. [Google Scholar] [CrossRef]
- Baonza, A.; Casci, T.; Freeman, M. A primary role for the epidermal growth factor receptor in ommatidial spacing in the Drosophila eye. Curr. Biol. 2001, 11, 396–404. [Google Scholar] [CrossRef] [Green Version]
- Kubota, K.; Goto, S.; Eto, K.; Hayashi, S. EGF receptor attenuates Dpp signaling and helps to distinguish the wing and leg cell fates in Drosophila. Development 2000, 127, 3769–3776. [Google Scholar]
- Munier, A.I.; Doucet, D.; Perrodou, E.; Zachary, D.; Meister, M.; Hoffmann, J.A.; Janeway, C.A., Jr.; Lagueux, M. PVF2, a PDGF/VEGF-like growth factor, induces hemocyte proliferation in Drosophila larvae. EMBO Rep. 2002, 3, 1195–1200. [Google Scholar] [CrossRef] [Green Version]
- Zettervall, C.J.; Anderl, I.; Williams, M.J.; Palmer, R.; Kurucz, E.; Ando, I.; Hultmark, D. A directed screen for genes involved in Drosophila blood cell activation. Proc. Natl. Acad. Sci. USA 2004, 101, 14192–14197. [Google Scholar] [CrossRef] [Green Version]
- Emori, Y.; Saigo, K. Distinct Expression of 2 Drosophila Homologs of Fibroblast Growth-Factor Receptors in Imaginal Disks. FEBS Lett. 1993, 332, 111–114. [Google Scholar] [CrossRef] [Green Version]
- Franzdottir, S.R.; Engelen, D.; Yuva-Aydemir, Y.; Schmidt, I.; Aho, A.; Klambt, C. Switch in FGF signalling initiates glial differentiation in the Drosophila eye. Nature 2009, 460, 758. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, T.; Choi, I.; Banerjee, U. Genetic Analysis of Fibroblast Growth Factor Signaling in the Drosophila Eye. G3-Genes Genomes Genet. 2012, 2, 23–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garofalo, R.S.; Rosen, O.M. Tissue Localization of Drosophila-Melanogaster Insulin-Receptor Transcripts during Development. Mol. Cell. Biol. 1988, 8, 1638–1647. [Google Scholar] [CrossRef] [Green Version]
- Cheng, L.Y.; Bailey, A.P.; Leevers, S.J.; Ragan, T.J.; Driscoll, P.C.; Gould, A.P. Anaplastic Lymphoma Kinase Spares Organ Growth during Nutrient Restriction in Drosophila. Cell 2011, 146, 435–447. [Google Scholar] [CrossRef] [Green Version]
- Cafferty, P.; Yu, L.; Rao, Y. The receptor tyrosine kinase off-track is required for layer-specific neuronal connectivity in Drosophila. Development 2004, 131, 5287–5295. [Google Scholar] [CrossRef] [Green Version]
- Dearborn, R.; He, Q.; Kunes, S.; Dai, Y. Eph receptor tyrosine kinase-mediated formation of a topographic map in the Drosophila visual system. J. Neurosci. 2002, 22, 1338–1349. [Google Scholar] [CrossRef] [Green Version]
- Rewitz, K.F.; Yamanaka, N.; Gilbert, L.I.; O’Connor, M.B. The Insect Neuropeptide PTTH Activates Receptor Tyrosine Kinase Torso to Initiate Metamorphosis. Science 2009, 326, 1403–1405. [Google Scholar] [CrossRef]
- Del Alamo, D.; Terriente, J.; Diaz-Benjumea, F.J. Spitz/EGFr signalling via the Ras/MAPK pathway mediates the induction of bract cells in Drosophila legs. Development 2002, 129, 1975–1982. [Google Scholar]
- Galindo, M.I.; Bishop, S.A.; Greig, S.; Couso, J.P. Leg patterning driven by proximal-distal interactions and EGFR signaling. Science 2002, 297, 256–259. [Google Scholar] [CrossRef]
- Huang, Z.; Shilo, B.Z.; Kunes, S. A retinal axon fascicle uses spitz, an EGF receptor ligand, to construct a synaptic cartridge in the brain of Drosophila. Cell 1998, 95, 693–703. [Google Scholar] [CrossRef] [Green Version]
- Jung, S.H.; Evans, C.J.; Uemura, C.; Banerjee, U. The Drosophila lymph gland as a developmental model of hematopoiesis. Development 2005, 132, 2521–2533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosin, D.; Schejter, E.; Volk, T.; Shilo, B.Z. Apical accumulation of the Drosophila PDGF/VEGF receptor ligands provides a mechanism for triggering localized actin polymerization. Development 2004, 131, 1939–1948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dutta, D.; Shaw, S.; Maqbool, T.; Pandya, H.; VijayRaghavan, K. Drosophila heartless acts with Heartbroken/Dof in muscle founder differentiation. PLoS Biol. 2005, 3, 1789–1800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeitouni, B.; Senatore, S.; Severac, D.; Aknin, C.; Semeriva, M.; Perrin, L. Signalling pathways involved in adult heart formation revealed by gene expression profiling in Drosophila. PLoS Genet. 2007, 3, 1907–1921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, S.M.; Baker, B.S. Sex-specific deployment of FGF signaling in Drosophila recruits mesodermal cells into the male genital imaginal disc. Cell 2002, 109, 651–661. [Google Scholar] [CrossRef] [Green Version]
- Sato, M.; Kornberg, T.B. FGF is an essential mitogen and chemoattractant for the air sacs of the Drosophila tracheal system. Dev. Cell 2002, 3, 195–207. [Google Scholar] [CrossRef]
- Bazigou, E.; Apitz, H.; Johansson, J.; Loren, C.E.; Hirst, E.M.A.; Chen, P.L.; Palmer, R.H.; Salecker, I. Anterograde jelly belly and Alk receptor tyrosine kinase signaling mediates retinal axon targeting in Drosophila. Cell 2007, 128, 961–975. [Google Scholar] [CrossRef] [Green Version]
- Michaut, L.; Flister, S.; Neeb, M.; White, K.P.; Certa, U.; Gehring, W.J. Analysis of the eye developmental pathway in Drosophila using DNA microarrays. Proc. Natl. Acad. Sci. USA 2003, 100, 4024–4029. [Google Scholar] [CrossRef] [Green Version]
- Buchon, N.; Broderick, N.A.; Kuraishi, T.; Lemaitre, B. Drosophila EGFR pathway coordinates stem cell proliferation and gut remodeling following infection. BMC Biol. 2010, 8. [Google Scholar] [CrossRef] [Green Version]
- Goode, S.; Wright, D.; Mahowald, A.P. The Neurogenic Locus Brainiac Cooperates with the Drosophila Egf Receptor to Establish the Ovarian Follicle and to Determine Its Dorsal-Vental Polarity. Development 1992, 116, 177–192. [Google Scholar] [PubMed]
- Kiger, A.A.; White-Cooper, H.; Fuller, M.T. Somatic support cells restrict germline stem cell self-renewal and promote differentiation. Nature 2000, 407, 750–754. [Google Scholar] [CrossRef] [PubMed]
- Parrott, B.B.; Hudson, A.; Brady, R.; Schulz, C. Control of Germline Stem Cell Division Frequency—A Novel, Developmentally Regulated Role for Epidermal Growth Factor Signaling. PLoS ONE 2012, 7, e36460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, N.H.; Kim, J.G.; Yang, D.J.; Kim, Y.S.; Yoo, M.A. Age-related changes in Drosophila midgut are associated with PVF2, a PDGF/VEGF-like growth factor. Aging Cell 2008, 7, 318–334. [Google Scholar] [CrossRef] [Green Version]
- Veenstra, J.A.; Agricola, H.J.; Sellami, A. Regulatory peptides in fruit fly midgut. Cell Tissue Res. 2008, 334, 499–516. [Google Scholar] [CrossRef] [PubMed]
- Dura, J.M.; Taillebourg, E.; Preat, T. The Drosophila Learning and Memory Gene Linotte Encodes a Putative Receptor Tyrosine Kinase Homologous to the Human Ryk Gene-Product. FEBS Lett. 1995, 370, 250–254. [Google Scholar] [CrossRef] [Green Version]
- Srahna, M.; Leyssen, M.; Choi, C.M.; Fradkin, L.G.; Noordermeer, J.N.; Hassan, B.A. A signaling network for patterning of neuronal connectivity in the Drosophila brain. PLoS Biol. 2006, 4, 2076–2090. [Google Scholar] [CrossRef]
- Adrain, C.; Freeman, M. Regulation of receptor tyrosine kinase ligand processing. Cold Spring Harb. Perspect. Biol. 2014, 6. [Google Scholar] [CrossRef] [Green Version]
- Steinhauer, J.; Liu, H.H.; Miller, E.; Treisman, J.E. Trafficking of the EGFR ligand Spitz regulates its signaling activity in polarized tissues. J. Cell Sci. 2013, 126, 4469–4478. [Google Scholar] [CrossRef] [Green Version]
- Wasserman, J.D.; Freeman, M. An autoregulatory cascade of EGF receptor signaling patterns the Drosophila egg. Cell 1998, 95, 355–364. [Google Scholar] [CrossRef] [Green Version]
- Dominguez, M.; Wasserman, J.D.; Freeman, M. Multiple functions of the EGF receptor in Drosophila eye development. Curr. Biol. 1998, 8, 1039–1048. [Google Scholar] [CrossRef] [Green Version]
- Sapir, A.; Schweitzer, R.; Shilo, B.Z. Sequential activation of the EGF receptor pathway during Drosophila oogenesis establishes the dorsoventral axis. Development 1998, 125, 191–200. [Google Scholar] [PubMed]
- Golembo, M.; Schweitzer, R.; Freeman, M.; Shilo, B.Z. argos transcription is induced by the Drosophila EGF receptor pathway to form an inhibitory feedback loop. Development 1996, 122, 223–230. [Google Scholar] [PubMed]
- Schweitzer, R.; Howes, R.; Smith, R.; Shilo, B.Z.; Freeman, M. Inhibition of Drosophila Egf Receptor Activation by the Secreted Protein Argos. Nature 1995, 376, 699–702. [Google Scholar] [CrossRef] [PubMed]
- Freeman, M.; Klambt, C.; Goodman, C.S.; Rubin, G.M. The Argos Gene Encodes a Diffusible Factor That Regulates Cell Fate Decisions in the Drosophila Eye. Cell 1992, 69, 963–975. [Google Scholar] [CrossRef]
- Avraham, R.; Yarden, Y. Feedback regulation of EGFR signalling: Decision making by early and delayed loops. Nat. Rev. Mol. Cell Biol. 2011, 12, 104–117. [Google Scholar] [CrossRef] [PubMed]
- Nassel, D.R.; Liu, Y.T.; Luo, J.N. Insulin/IGF signaling and its regulation in Drosophila. Gen. Comp. Endocrinol. 2015, 221, 255–266. [Google Scholar] [CrossRef]
- Tatar, M.; Kopelman, A.; Epstein, D.; Tu, M.P.; Yin, C.M.; Garofalo, R.S. A mutant Drosophila insulin receptor homolog that extends life-span and impairs neuroendocrine function. Science 2001, 292, 107–110. [Google Scholar] [CrossRef] [Green Version]
- Brogiolo, W.; Stocker, H.; Ikeya, T.; Rintelen, F.; Fernandez, R.; Hafen, E. An evolutionarily conserved function of the Drosophila insulin receptor and insulin-like peptides in growth control. Curr. Biol. 2001, 11, 213–221. [Google Scholar] [CrossRef] [Green Version]
- Junger, M.A.; Rintelen, F.; Stocker, H.; Wasserman, J.D.; Vegh, M.; Radimerski, T.; Greenberg, M.E.; Hafen, E. The Drosophila forkhead transcription factor FOXO mediates the reduction in cell number associated with reduced insulin signaling. J. Biol. 2003, 2, 20. [Google Scholar] [CrossRef] [Green Version]
- Bohni, R.; Riesgo-Escovar, J.; Oldham, S.; Brogiolo, W.; Stocker, H.; Andruss, B.F.; Beckingham, K.; Hafen, E. Autonomous control of cell and organ size by CHICO, a Drosophila homolog of vertebrate IRS1-4. Cell 1999, 97, 865–875. [Google Scholar] [CrossRef] [Green Version]
- Kido, Y.; Nakae, J.; Accili, D. Clinical review 125—The insulin receptor and its cellular targets. J. Clin. Endocrinol. Metab. 2001, 86, 972–979. [Google Scholar] [CrossRef] [PubMed]
- Slaidina, M.; Delanoue, R.; Gronke, S.; Partridge, L.; Leopold, P. A Drosophila Insulin-like Peptide Promotes Growth during Nonfeeding States. Dev. Cell 2009, 17, 874–884. [Google Scholar] [CrossRef] [PubMed]
- Colombani, J.; Andersen, D.S.; Leopold, P. Secreted Peptide Dilp8 Coordinates Drosophila Tissue Growth with Developmental Timing. Science 2012, 336, 582–585. [Google Scholar] [CrossRef]
- Nassel, D.R.; Vanden Broeck, J. Insulin/IGF signaling in Drosophila and other insects: Factors that regulate production, release and post-release action of the insulin-like peptides. Cell. Mol. Life Sci. 2016, 73, 271–290. [Google Scholar] [CrossRef]
- Ikeya, T.; Galic, M.; Belawat, P.; Nairz, K.; Hafen, E. Nutrient-dependent expression of insulin-like peptides from neuroendocrine cells in the CNS contributes to growth regulation in Drosophila. Curr. Biol. 2002, 12, 1293–1300. [Google Scholar] [CrossRef] [Green Version]
- Gronke, S.; Clarke, D.F.; Broughton, S.; Andrews, T.D.; Partridge, L. Molecular Evolution and Functional Characterization of Drosophila Insulin-Like Peptides. PLoS Genet. 2010, 6. [Google Scholar] [CrossRef] [Green Version]
- Rulifson, E.J.; Kim, S.K.; Nusse, R. Ablation of insulin-producing neurons in flies: Growth and diabetic phenotypes. Science 2002, 296, 1118–1120. [Google Scholar] [CrossRef]
- Kim, J.; Neufeld, T.P. Dietary sugar promotes systemic TOR activation in Drosophila through AKH-dependent selective secretion of Dilp3. Nat. Commun. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Honegger, B.; Galic, M.; Kohler, K.; Wittwer, F.; Brogiolo, W.; Hafen, E.; Stocker, H. Imp-L2, a putative homolog of vertebrate IGF-binding protein 7, counteracts insulin signaling in Drosophila and is essential for starvation resistance. J. Biol. 2008, 7, 10. [Google Scholar] [CrossRef] [Green Version]
- Shibuya, M. VEGFR and type-V RTK activation and signaling. Cold Spring Harb. Perspect. Biol. 2013, 5, a009092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrae, J.; Gallini, R.; Betsholtz, C. Role of platelet-derived growth factors in physiology and medicine. Genes Dev. 2008, 22, 1276–1312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shibuya, M. Vascular endothelial growth factor and its receptor system: Physiological functions in angiogenesis and pathological roles in various diseases. J. Biochem. 2013, 153, 13–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, N.K.; Keyes, L.; Johnson, E.; Heller, J.; Ryner, L.; Karim, F.; Krasnow, M.A. Developmental control of blood cell migration by the Drosophila VEGF pathway. Cell 2002, 108, 865–876. [Google Scholar] [CrossRef] [Green Version]
- Olofsson, B.; Page, D.T. Condensation of the central nervous system in embryonic Drosophila is inhibited by blocking hemocyte migration or neural activity. Dev. Biol. 2005, 279, 233–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, W.; Faria, C.; Jacinto, A. Distinct mechanisms regulate hemocyte chemotaxis during development and wound healing in Drosophila melanogaster. J. Cell Biol. 2006, 173, 405–416. [Google Scholar] [CrossRef] [Green Version]
- Learte, A.R.; Forero, M.G.; Hidalgo, A. Gliatrophic and gliatropic roles of PVF/PVR signaling during axon guidance. Glia 2008, 56, 164–176. [Google Scholar] [CrossRef]
- Bunt, S.; Hooley, C.; Hu, N.; Scahill, C.; Weavers, H.; Skaer, H. Hemocyte-secreted type IV collagen enhances BMP signaling to guide renal tubule morphogenesis in Drosophila. Dev. Cell 2010, 19, 296–306. [Google Scholar] [CrossRef] [Green Version]
- Harris, K.E.; Schnittke, N.; Beckendorf, S.K. Two ligands signal through the Drosophila PDGF/VEGF receptor to ensure proper salivary gland positioning. Mech. Dev. 2007, 124, 441–448. [Google Scholar] [CrossRef] [Green Version]
- Bond, D.; Foley, E. Autocrine Platelet-derived Growth Factor-Vascular Endothelial Growth Factor Receptor-related (Pvr) Pathway Activity Controls Intestinal Stem Cell Proliferation in the Adult Drosophila Midgut. J. Biol. Chem. 2012, 287, 27359–27370. [Google Scholar] [CrossRef] [Green Version]
- Joukov, V.; Sorsa, T.; Kumar, V.; Jeltsch, M.; ClaessonWelsh, L.; Cao, Y.H.; Saksela, O.; Kalkkinen, N.; Alitalo, K. Proteolytic processing regulates receptor specificity and activity of VEGF-C. EMBO J. 1997, 16, 3898–3911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parsons, B.; Foley, E. The Drosophila Platelet-derived Growth Factor and Vascular Endothelial Growth Factor-Receptor Related (Pvr) Protein Ligands Pvf2 and Pvf3 Control Hemocyte Viability and Invasive Migration. J. Biol. Chem. 2013, 288, 20173–20183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siekhaus, D.; Haesemeyer, M.; Moffitt, O.; Lehmann, R. RhoL controls invasion and Rap1 localization during immune cell transmigration in Drosophila. Nat. Cell Biol. 2010, 12, 605–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strecker, T.R.; Halsell, S.R.; Fisher, W.W.; Lipshitz, H.D. Reciprocal Effects of Hyperactivity and Hypoactivity Mutations in the Drosophila Pattern Gene Torso. Science 1989, 243, 1062–1066. [Google Scholar] [CrossRef] [PubMed]
- Casanova, J.; Furriols, M.; McCormick, C.A.; Struhl, G. Similarities between trunk and spatzle, putative extracellular ligands specifying body pattern in Drosophila. Genes Dev. 1995, 9, 2539–2544. [Google Scholar] [CrossRef] [Green Version]
- Savant-Bhonsale, S.; Montell, D.J. Torso-Like Encodes the Localized Determinant of Drosophila Terminal Pattern-Formation. Genes Dev. 1993, 7, 2548–2555. [Google Scholar] [CrossRef]
- Jimenez, G.; Gonzalez-Reyes, A.; Casanova, J. Cell surface proteins Nasrat and Polehole stabilize the Torso-like extracellular determinant in Drosophila oogenesis. Genes Dev. 2002, 16, 913–918. [Google Scholar] [CrossRef] [Green Version]
- Stevens, L.M.; Beuchle, D.; Jurcsak, J.; Tong, X.L.; Stein, D. The Drosophila embryonic patterning determinant torsolike is a component of the eggshell. Curr. Biol. 2003, 13, 1058–1063. [Google Scholar] [CrossRef] [Green Version]
- Martin, J.R.; Raibaud, A.; Ollo, R. Terminal Pattern Elements in Drosophila Embryo Induced by the Torso-Like Protein. Nature 1994, 367, 741–745. [Google Scholar] [CrossRef]
- Sprenger, F.; Nusslein Volhard, C. Torso Receptor Activity Is Regulated by a Diffusible Ligand Produced at the Extracellular Terminal Regions of the Drosophila Egg. Cell 1992, 71, 987–1001. [Google Scholar] [CrossRef]
- Duncan, E.J.; Benton, M.A.; Dearden, P.K. Canonical terminal patterning is an evolutionary novelty. Dev. Biol. 2013, 377, 245–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goyal, Y.; Schupbach, T.; Shvartsman, S.Y. A quantitative model of developmental RTK signaling. Dev. Biol. 2018, 442, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Ponting, C.P. Chlamydial homologues of the MACPF (MAC/perforin) domain. Curr. Biol. 1999, 9, R911–R913. [Google Scholar] [CrossRef] [Green Version]
- Rosado, C.J.; Buckle, A.M.; Law, R.H.P.; Butcher, R.E.; Kan, W.T.; Bird, C.H.; Ung, K.; Browne, K.A.; Baran, K.; Bashtannyk-Puhalovich, T.A.; et al. A common fold mediates vertebrate defense and bacterial attack. Science 2007, 317, 1548–1551. [Google Scholar] [CrossRef] [Green Version]
- Johnson, T.K.; Henstridge, M.A.; Warr, C.G. MACPF/CDC proteins in development: Insights from Drosophila torso-like. Semin. Cell Dev. Biol. 2017, 72, 163–170. [Google Scholar] [CrossRef]
- Casali, A.; Casanova, J. The spatial control of Torso RTK activation: A C-terminal fragment of the Trunk protein acts as a signal for Torso receptor in the Drosophila embryo. Development 2001, 128, 1709–1715. [Google Scholar]
- Johnson, T.K.; Henstridge, M.A.; Herr, A.; Moore, K.A.; Whisstock, J.C.; Warr, C.G. Torso-like mediates extracellular accumulation of Furin-cleaved Trunk to pattern the Drosophila embryo termini. Nat. Commun. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Mineo, A.; Fuentes, E.; Furriols, M.; Casanova, J. Holes in the Plasma Membrane Mimic Torso-Like Perforin in Torso Tyrosine Kinase Receptor Activation in the Drosophila Embryo. Genetics 2018, 210, 257–262. [Google Scholar] [CrossRef] [Green Version]
- Kondos, S.C.; Hatfaludi, T.; Voskoboinik, I.; Trapani, J.A.; Law, R.H.P.; Whisstock, J.C.; Dunstone, M.A. The structure and function of mammalian membrane-attack complex/perforin-like proteins. Tissue Antigens 2010, 76, 341–351. [Google Scholar] [CrossRef]
- McBrayer, Z.; Ono, H.; Shimell, M.; Parvy, J.P.; Beckstead, R.B.; Warren, J.T.; Thummel, C.S.; Dauphin-Villemant, C.; Gilbert, L.I.; O’Connor, M.B. Prothoracicotropic hormone regulates developmental timing and body size in Drosophila. Dev. Cell 2007, 13, 857–871. [Google Scholar] [CrossRef] [Green Version]
- Selcho, M.; Millan, C.; Palacios-Munoz, A.; Ruf, F.; Ubillo, L.; Chen, J.; Bergmann, G.; Ito, C.; Silva, V.; Wegener, C.; et al. Central and peripheral clocks are coupled by a neuropeptide pathway in Drosophila. Nat. Commun. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Jenni, S.; Goyal, Y.; von Grotthuss, M.; Shvartsman, S.Y.; Klein, D.E. Structural Basis of Neurohormone Perception by the Receptor Tyrosine Kinase Torso. Mol. Cell 2015, 60, 941–952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coppey, M.; Boettiger, A.N.; Berezhkovskii, A.M.; Shvartsman, S.Y. Nuclear trapping shapes the terminal gradient in the Drosophila embryo. Curr. Biol. 2008, 18, 915–919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helman, A.; Cinnamon, E.; Mezuman, S.; Hayouka, Z.; Von Ohlen, T.; Orian, A.; Jimenez, G.; Paroush, Z. Phosphorylation of Groucho Mediates RTK Feedback Inhibition and Prolonged Pathway Target Gene Expression. Curr. Biol. 2011, 21, 1102–1110. [Google Scholar] [CrossRef] [Green Version]
- Vinayagam, A.; Kulkarni, M.M.; Sopko, R.; Sun, X.Y.; Hu, Y.H.; Nand, A.; Villalta, C.; Moghimi, A.; Yang, X.M.; Mohr, S.E.; et al. An Integrative Analysis of the InR/PI3K/Akt Network Identifies the Dynamic Response to Insulin Signaling. Cell Rep. 2016, 16, 3062–3074. [Google Scholar] [CrossRef] [Green Version]
- Harvey, C.D.; Ehrhardt, A.G.; Cellurale, C.; Zhong, H.N.; Yasuda, R.; Davis, R.J.; Svoboda, K. A genetically encoded fluorescent sensor of ERK activity. Proc. Natl. Acad. Sci. USA 2008, 105, 19264–19269. [Google Scholar] [CrossRef] [Green Version]
- Swift, J.L.; Godin, A.G.; Dore, K.; Freland, L.; Bouchard, N.; Nimmo, C.; Sergeev, M.; De Koninck, Y.; Wiseman, P.W.; Beaulieu, J.M. Quantification of receptor tyrosine kinase transactivation through direct dimerization and surface density measurements in single cells. Proc. Natl. Acad. Sci. USA 2011, 108, 7016–7021. [Google Scholar] [CrossRef] [Green Version]
- De la Cova, C.; Townley, R.; Regot, S.; Greenwald, I. A Real-Time Biosensor for ERK Activity Reveals Signaling Dynamics during C. elegans Cell Fate Specification. Dev. Cell 2017, 42, 542–553. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Lee, S.; Jung, K.; Oh, W.C.; Kim, N.; Son, S.; Jo, Y.; Kwon, H.B.; Heo, W.D. Intensiometric biosensors visualize the activity of multiple small GTPases in vivo. Nat. Commun. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Leopold, A.V.; Chernov, K.G.; Shemetov, A.A.; Verkhusha, V.V. Neurotrophin receptor tyrosine kinases regulated with near-infrared light. Nat. Commun. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Goglia, A.G.; Wilson, M.Z.; DiGiorno, D.B.; Toettcher, J.E. Optogenetic Control of Ras/Erk Signaling Using the Phy-PIF System. Methods Mol. Biol. 2017, 1636, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Grusch, M.; Schelch, K.; Riedler, R.; Reichhart, E.; Differ, C.; Berger, W.; Ingles-Prieto, A.; Janovjak, H. Spatio-temporally precise activation of engineered receptor tyrosine kinases by light. EMBO J. 2014, 33, 1713–1726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krueger, D.; Izquierdo, E.; Viswanathan, R.; Hartmann, J.; Cartes, C.P.; De Renzis, S. Principles and applications of optogenetics in developmental biology. Development 2019, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, H.E.; Toettcher, J.E. Illuminating developmental biology with cellular optogenetics. Curr. Opin. Biotechnol. 2018, 52, 42–48. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Mammalian RTK Family Symbol | Mammalian RTK Family Members | Drosophila RTK Homolog |
|---|---|---|
| ALK | ALK, LTK | Anaplastic lymphoma kinase (Alk) |
| FGFR | FGFR(1–4) | Breathless (Btl), Heartless (Htl) |
| DDR1 and DDR2 | DDR1, DDR2 | Discoidin domain receptor (Ddr) |
| RYK | RYK | Doughnut on 2 (Dnt), Derailed (Drl)†, Derailed 2 (Drl-2) |
| EGFR | EGFR, ERBB(2–4) | Epidermal growth factor receptor (EGFR) |
| EPH | EphA(1–8), EphA10 | Erythropoietin-producing human hepatocellular carcinoma cell line (Eph) |
| INSR/IGF1R | INSR, IGF1R, IGF2R | Insulin-like receptor (Inr) |
| MuSK | MuSK | Neurotrophic receptor kinase (Nrk) |
| TRK | TrkA, TrkB, TrkC | Offtrack (Otk)† |
| PDGFR | PDGFRα, PDGFR β, Kit, CSF-1R | PDGF- and VEGF-receptor related (Pvr) |
| VEGFR | VEGFR(1–3) | PDGF- and VEGF-receptor related (Pvr) |
| RET | RET | Ret oncogene (Ret) |
| ROR1 and ROR2 | ROR1, ROR2 | RTK-like orphan receptors (Ror) |
| TIE | TIE1, TIE 2 | Tie-like receptor (Tie) |
| Sevenless (Sev) | ||
| Torso (Tor) | ||
| Stitcher (Cad96Ca) |
| Life Stage | Receptor (Gene Symbol) | Ligand (Gene Symbol) | Function | References |
|---|---|---|---|---|
| Embryo | tor | trk | Terminal patterning (maternal) | [64,65] |
| Egfr | spi, vn | Embryonic patterning, cell recruitment, specification, proliferation, cell attachment | [60,66,67,68] | |
| htl | pyr, ths | Visceral mesoderm specification, migration during gastrulation (cardiogenesis) | [69,70] | |
| btl | bnl | Tracheal cell migration, central nervous system patterning | [71] | |
| sev | boss | Male gonad stem cell niche restriction | [72] | |
| InR | Ilp2, 4, 7 | Epidermal and neural cell growth | [73] | |
| Pvr | Pvf2, 3 | Hemocyte migration | [74,75,76] | |
| dnt, drl, drl-2 | wnt5 | Axon targeting, salivary gland cell migration | [77,78,79] | |
| Alk | jeb | Muscle cell specification, neuronal differentiation | [80,81] | |
| Ror | − | Neural cell growth | [82] | |
| Nrk | − | Neural cell growth† | [83] | |
| otk | wnt4 | Neuroblast migration | [84] | |
| Cad96Ca | collagen | Wound healing, axon patterning† | [56] | |
| Tie | Pvf1† | Border cell migration | [85] | |
| Larva | sev | boss | Photoreceptor specification | [86] |
| Egfr | spi, vn | Imaginal and wing disc determination, proliferation of eye disc cells | [51,87,88] | |
| Pvr | Pvf2 | Hemocyte proliferation | [89,90] | |
| htl | pyr, ths | Imaginal disc determination | [91,92] | |
| btl | bnl | Retinal patterning and glial migration | [93] | |
| InR | Ilps 1–6 | Neural cell and imaginal disc growth | [94] | |
| Alk | jeb | Spares organ growth during starvation | [95] | |
| otk | - | Photoreceptor axon guidance | [96] | |
| Eph | Eph | Photoreceptor axon pathfinding | [97] | |
| tor | Ptth | Initiation of metamorphosis | [98] | |
| Pupa | Egfr | spi, vn | Neuron differentiation, leg patterning, specification of bract cell fate | [99,100,101] |
| Pvr | Pvf1, 3 | Hemocyte proliferation and differentiation, maintenance of cell adhesion | [102,103] | |
| htl | pyr, ths | Heart muscle remodeling, leg and thoracic muscle cell differentiation | [104,105] | |
| btl | bnl | Imaginal tracheoblast remodeling, eye disc remodeling, male genital disc migration | [93,106,107] | |
| Alk | jeb | Photoreceptor axon migration | [108] | |
| Nrk | − | Nervous system restructuring | [83] | |
| Tie | - | Retinal cell differentiation† | [109] | |
| Adult | Egfr | grk, spi | Midgut maintenance, spermatogenesis, oogenesis, germline stem cell attenuation (testes), border cell migration (ovary) | [110,111,112,113] |
| Pvr | Pvf1, 2 | Midgut maintenance, border cell migration (ovary) | [74,114] | |
| InR | Ilp2, 3, 5 | Brain, thorax, abdomen, and gut cell maintenance; body growth | [5,115] | |
| drl | wnt5 | Axon targeting | [116] | |
| Btl | bnl | Axon retraction (visual system) | [117] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mele, S.; Johnson, T.K. Receptor Tyrosine Kinases in Development: Insights from Drosophila. Int. J. Mol. Sci. 2020, 21, 188. https://doi.org/10.3390/ijms21010188
Mele S, Johnson TK. Receptor Tyrosine Kinases in Development: Insights from Drosophila. International Journal of Molecular Sciences. 2020; 21(1):188. https://doi.org/10.3390/ijms21010188
Chicago/Turabian StyleMele, Sarah, and Travis K. Johnson. 2020. "Receptor Tyrosine Kinases in Development: Insights from Drosophila" International Journal of Molecular Sciences 21, no. 1: 188. https://doi.org/10.3390/ijms21010188
APA StyleMele, S., & Johnson, T. K. (2020). Receptor Tyrosine Kinases in Development: Insights from Drosophila. International Journal of Molecular Sciences, 21(1), 188. https://doi.org/10.3390/ijms21010188